
Species diversity of plant communities and its main influencing factors in Horqin Sandy Land, Inner Mongolia of northern China
-
摘要:目的科尔沁沙地地处我国北方农牧交错带,其植物多样性组成与格局对人为干扰及其导致的环境因子变化极其敏感。近年来,科尔沁地区农牧业发展迅速,人为干扰对自然植被的影响不断加大,但是探讨放牧干扰与环境因子对植物群落不同水平物种多样性相对影响的研究还比较缺乏。方法本研究基于44个调查点,放牧干扰数据以及环境数据,通过单因素方差分析、一元线性回归、多项式回归和方差分解等方法,分析科尔沁沙地植物群落α、β、γ多样性分布特征及其主要影响因素。结果(1)群落α、β、γ多样性均随放牧强度的增加而逐渐下降,不同放牧强度之间均存在显著差异(P < 0.05),重度放牧下群落α、β、γ多样性均显著低于围封区(P < 0.05),在围封区达到最高。(2)群落α多样性与放牧强度和最热月均温呈极显著负相关关系(P < 0.01),与气温日较差、土壤有机碳和土壤全磷含量呈极显著的正相关关系(P < 0.01),随年均降水量的增加而呈显著的先减后增的变化趋势(P < 0.05),并随着最冷月均温和土壤全氮含量的增加呈显著的先增后减的变化趋势(P < 0.05)。群落β多样性与放牧强度、潜在蒸散量和最冷月均温呈显著的负相关关系(P < 0.05),并与年均降水量呈显著的正相关关系(P < 0.05)。群落γ多样性与放牧强度呈极显著的负相关关系(P < 0.001),与土壤有机碳和土壤全氮含量呈极显著的正相关关系(P < 0.01)。(3)放牧干扰是群落α和β多样性的主要影响因素,放牧干扰与土壤因子的协同作用是群落γ多样性的主要影响因素。结论放牧干扰对科尔沁沙地植物群落不同水平物种多样性起着关键作用。因此需要制定科学的放牧管理制度、加强监管力度,对严重退化的区域实行围封禁牧,在降低放牧强度的同时打击偷牧行为,能够有效促进科尔沁沙地植物多样性保护和退化生态系统自然恢复。Abstract:ObjectiveThe Horqin Sandy Land is located in agro-pastoral ecotone of northern China, and its species composition and diversity patterns are quite vulnerable to the changes of human disturbance and environmental factors. Because of the fast development of agriculture and animal husbandry industry in recent years, human disturbance has become an essential role in the Horqin Sandy Land. However, further systematic studies are needed to understand the relative influence of grazing disturbance and environmental factors on the α, β and γ diversity of plant communities in the Horqin Sandy Land.MethodThe grazing disturbance data and environmental data of 44 sampling points were used to study the distribution characteristics of the α, β and γ diversity of plant communities and their main influencing factors in the Horqin Sandy Land by one-way ANOVA, simple linear regression analysis, polynomial regression analysis, and variance decomposition.Result(1) The α, β and γ diversity of plant communities declined as grazing intensity increased, and the peak species diversity appeared under grazing exclusion. There was a significant difference in the α, β and γ diversity of plant communities among different grazing intensities (P <0.05). The α, β and γ diversity under heavy grazing intensity were significantly lower than that under grazing exclusion (P < 0.05). (2) The α diversity of plant communities had a significantly negative correlation with grazing intensity and mean temperature of the hotest month (P < 0.01), and a positive correlation with mean diurnal range, soil organic carbon and soil total phosphorus (P < 0.01); it had a significant U-shaped relationship with mean annual precipitation, and a hump-shaped relationship with mean temperature of the coldest month and soil total nitrogen (P < 0.05). The β diversity of plant communities had a significantly negative correlation with grazing intensity, potential evapotranspiration, and mean temperature of the coldest month (P < 0.05), and a positive correlation with mean annual precipitation (P < 0.05). The γ diversity of plant communities had a significantly negative correlation with grazing intensity (P < 0.001), and a positive correlation with soil organic carbon and soil total nitrogen (P < 0.01). (3) Grazing disturbance was the main influencing factor affecting the α and β diversity of plant communities; the synergism between grazing disturbance and soil factors was the main influencing factor affecting the γ diversity of plant communities.ConclusionGrazing disturbance has a critical effect on species diversity of plant communities in the Horqin Sandy Land. Therefore, in order to enhance biodiversity conservation and restore the degraded ecosystem in the Horqin Sandy Land, local government should formulate polices to forbid overgrazing and enhance supervision to reduce the occurrence of grazing without permission.
-
Keywords:
- Horqin Sandy Land /
- α diversity /
- β diversity /
- γ diversity /
- grazing disturbance /
- environmental factor
-
植物是依赖于光合作用生存的光能自养型生物,而光合作用的主要场所为叶绿体。叶绿体拥有自身的遗传物质DNA[1],其基因组较小,仅占植株总基因组的10% ~ 20%,呈双链闭环形式,一般为四分体结构[2],包括小单拷贝区(SSC,small single copy region),大单拷贝区(LSC,large single copy region),和两个大小相等、方向相反的反向重复区(IR,inverted repeat region)。大单拷贝区和小单拷贝区被两个反向重复区在中间隔开。叶绿体是半自主细胞器,具有独立的遗传系统,作为光合作用的中心,同时进行着多种生化反应[3]。
密码子在核酸和蛋白质之间起传递的作用[4],其偏好性会受到自然选择、基因突变、碱基组成、遗传进化、基因漂移等多种因素影响[5]。密码子偏好性可影响物种的分子进化、环境适应和基因组特征,进而影响外源基因的表达、反映物种间的进化关系,还可为引种驯化提供重要依据。通过分析密码子中碱基的使用情况,可以推断影响其使用偏好的原因。叶绿体基因组密码子对植物光合作用的机制和代谢调控具有重要意义。此前,已有研究对忍冬属(Lonicera)[6]、车轴草属(Trifolium)[7]、赖草属(Leymus)[8]、梁山慈竹(Dendrocalamus farinosus)[9]、人参(Panax ginseng)和三七(Panax notoginseng)[10]等植物的叶绿体基因组密码子偏好性进行了探讨。目前,关于油茶叶绿体基因组密码子偏好性分析的报道仅限于普通油茶(Camellia oleifera)[11]和香花油茶(Camellia osmantha)[12],研究发现其密码子偏好性不仅受到突变作用的影响,更多地受到自然选择的作用。
油茶属于山茶科(Theaceae)山茶属(Camellia),灌木或中乔木,嫩枝有粗毛,叶革质,花顶生,蒴果球形或卵圆形,与油橄榄(Olea europaea)、油棕(Elaeis guineensis)、椰子(Cocos nucifera)并称世界四大木本油料植物[13],与乌桕(Triadica sebifera)、油桐(Vernicia fordii)、核桃(Juglans regia)并称中国四大木本油料植物[14]。油茶不仅可提炼食用油,还可入药,茶枯还可作为农药、肥料,有提高农田蓄水能力和防治稻田害虫的功能[15]。小果油茶(Camellia meiocarpa)又称小子茶和江西子,因其种子形状与鸡心相似,故又称鸡心子[16]。其生长习性与普通油茶相似,适宜种植在亚热带季风气候的中低海拔山地、丘陵地区。越南油茶(Camellia vietnamensis)分布于中国广东南部、海南、广西靠近越南边界各县,以及越南和老挝。越南油茶是一种在中国广泛栽培的油料树种,栽培面积仅次于普通油茶和小果油茶。小果油茶和越南油茶是油茶资源中的两个重要栽培类型,其栽培面积在全国范围内位居前三,两者在植物学特性、地理分布、果实特征、栽培利用和亲缘关系等方面既存在显著差异,又具有一定的内在联系。借助基因测序、分子标记等现代生物技术手段,能够深入剖析小果油茶和越南油茶之间的遗传差异,为其遗传改良提供坚实的理论基础。通过对小果油茶与越南油茶叶绿体基因组密码子偏好性的研究,可以揭示二者在进化规律和突变方式上的特点,进而阐释这两个品种适应外界环境的分子机制。在此研究基础上,通过对目标基因密码子进行优化,能够显著提高基因的表达效率。这不仅为后续良种培育提供了理论依据,更为将小果油茶的适应性强、抗炭疽病能力强、产量稳定等优良性状与越南油茶生长速度快、种仁含油率高这一优势相结合,培育出更具市场竞争力的新品种奠定了基础。
截至目前,尚未有针对小果油茶和越南油茶这两个油茶近源物种叶绿体基因组密码子使用偏好性的对比研究报道。本研究以市场上常见的小果油茶和越南油茶为研究对象,拟通过相关软件对其叶绿体基因组密码子进行碱基组成分析、中性绘图、ENC-plot绘图、PR2-plot偏倚、最优密码子分析,构建高低表达库,揭示叶绿体基因组密码子的使用偏好性,并探讨影响其偏好性的原因,进而筛选出两物种的最优密码子。研究结果旨在为油茶基因组研究及其良种培育提供理论依据。
1. 材料与方法
1.1 序列材料
在NCBI数据库中下载小果油茶和越南油茶的叶绿体全基因组CDS序列,小果油茶[17]登陆号为NC_058881.1,共85条基因序列156 550 bp。越南油茶[18]登录号为NC_060778.1,共87条基因序列156 999 bp。为减少结果误差,将两个物种的CDS序列进行筛选,剔除重复和长度小于300 bp的序列后[19],选择包含起始密码子(ATG)和终止密码子(TAA、TAG、TGA)的序列[20],最终两物种以各52条CDS序列作为此次分析的基础数据,利用Origin、Excel等软件绘制相关图表[21]。
1.2 研究方法
1.2.1 密码子组成分析
运用Codon W 1.4.2软件、CUSP在线软件(http://emboss.toulouse.inra.fr/cgi-bin/emboss/cusp)、MEGA软件[22]等分析处理剔除相关序列后的CDS序列,得出氨基酸密码子组成数量、同义密码子相对使用度(relative synonymous codon usage,RSCU)、有效密码子数(effective number of codon,ENC)、密码子适应指数(codon bias index,CAI)、各碱基(A、T、G、C)在每一位置的含量以及每一位置的G + C含量。其中,A3、T3、G3、C3为基因中所有密码子第3位碱基A、T、G、C的含量;GC1、GC2、GC3分别为基因中所有密码子(除蛋氨酸、色氨酸和终止密码子外)第1位、第2位、第3位碱基G + C的含量。ENC计算公式为
ENC=2+t+29/(t2+(1-t2)2) 式中:t 为密码子第3位G + C的含量(GC3)。
1.2.2 中性绘图分析
以GC3为横坐标,以 GC1 和 GC2 的均值(GC12)为纵坐标绘制散点图,图中每 1个点代表1个基因的位置,直观体现3个密码子位置之间的相关性[23],初步判断密码子使用偏好性的影响因素[24−25]。若回归曲线斜率较低,表明基因突变对密码子偏好性的影响较小,自然选择在其中发挥了更重要的作用。
表 1 小果油茶与越南油茶叶绿体基因组中GC含量、ENC值和CAI值Table 1. GC content, ENC value and CAI value in chloroplast genes of Camellia meiocarpa and C. vietnamensis物种 A3/% T3/% G3/% C3/% GC1/% GC2/% GC3/% GCall/% ENC CAI 小果油茶 Camellia meiocarpa 39.73 36.96 27.85 23.22 45.81 38.00 28.64 37.49 48.561 0.158 越南油茶 Camellia vietnamensis 39.82 38.74 26.28 22.75 45.88 38.02 28.59 37.50 48.529 0.160 注:T3、C3、A3、G3分别代表基因组中所有密码子第3位碱基T、C、A、G的含量;GC1、GC2、GC3分别代表基因组中所有密码子(除蛋氨酸、色氨酸和终止密码子外)第1位、第2位、第3位碱基G + C的含量;GCall:基因中所有密码子第1位、第2位、第3位碱基G + C含量的平均值;ENC表示有效密码子数;CAI表示密码子适应指数。下同。 1.2.3 ENC-plot绘图分析
以GC3为横坐标,以ENC值为纵坐标使用Origin绘制散点图,并在散点图中绘制ENC值的标准曲线。ENC常被用来衡量单个基因的密码子偏好性大小,取值范围在 20 ~ 61 之间。其值越接近20,认为该基因具有强密码子偏好性,越接近61,认为该基因无密码子偏好性。
当基因ENC值在标准曲线附近时,表明基因密码子使用偏好受突变压力的影响较大;而当 ENC 值偏离标准曲线较远时,表明密码子使用模式受选择压力的影响较大[26−28]。以ENC检测值和ENC标准值两者的差值与ENC标准值的比值作为各基因有效密码子比值,并进行油茶叶绿体基因组有效密码子比值频率分布分析。
1.2.4 PR2-plot偏倚分析
以A3/(A3 + T3)为纵坐标,G3/( G3 + C3)为横坐标绘制散点图,并画出散点图的中心点(A = T且C = G),表示基因两条互补链间不存在任何突变或自然选择上的偏倚,从中心点发出的矢量距离代表该碱基偏移的程度和方向[29]。如果密码子第 3 位碱基使用AT 的频率等于 GC 的频率,则表明基因的密码子使用偏好仅受到突变压力的影响;否则,说明自然选择或两者共同作用影响了密码子的偏好性使用[30]。
1.2.5 最优密码子分析
以ENC值作为高表达和低表达基因的参考依据,将所有基因的CDS序列按照ENC值进行排序,排序前后两端各选取5条基因序列,分别建立高、低表达组。随后计算高、低表达组的RSCU,以 RSCU > 1 的密码子为高频密码子,并计算△RSCU(高、低两组RSCU的差值),△RSCU ≥ 0.08的密码子作为高表达密码子。高频(RSCU > 1)且高表达(△RSCU ≥ 0.08)的密码子确定为油茶叶绿体基因组的最优密码子[31]。
2. 结果与分析
2.1 小果油茶和越南油茶密码子组成特征
小果油茶和越南油茶叶绿体基因组密码子中GC分布不均,均呈现GC1 > GC2 > GC3;两物种A3/T3均远大于G3/C3,表明两种油茶编码基因的密码子中碱基偏好以A/T结尾;CAI理论取值在0 ~ 1之间[32],两物种的CAI值均小于0.17,表明小果油茶与越南油茶的适应性弱(表1)。
在小果油茶与越南油茶叶绿体基因组不同的基因中,ENC值分布在35.640 ~ 61.000之间,两物种ENC均值为48.561和48.529(表2)。ENC值可决定偏好性强弱,以35为标准,低于35为偏好性强,高于35为偏好性弱[33]。因两物种各52条叶绿体基因组序列ENC均大于35,可知两物种叶绿体基因组密码子的偏好性均比较弱。
表 2 小果油茶和越南油茶不同基因密码子的GC含量和ENC值Table 2. GC contents and ENC values of different gene codons of Camellia meiocarpa and C. vietnamensis基因 小果油茶Camellia meiocarpa 越南油茶C. vietnamensis GC1/% GC2/% GC3/% GCall/% ENC GC1/% GC2/% GC3/% GCall/% ENC rps12 51.61 48.39 27.42 44.90 55.31 52.10 50.42 29.41 45.60 45.54 ycf1 43.55 34.68 38.71 30.70 49.34 45.38 33.61 38.66 30.60 49.16 rps7 50.00 45.16 20.16 40.40 45.81 51.26 45.38 20.17 41.70 53.41 ndhB 37.10 30.65 35.48 37.40 50.52 36.97 30.25 36.13 37.00 46.46 ycf2 40.32 31.45 37.90 37.70 53.35 39.50 31.09 36.97 38.80 50.59 rpl2 45.97 42.74 30.65 45.00 57.04 38.98 43.22 25.42 39.00 41.33 psbA 48.39 45.16 31.45 43.40 55.78 47.06 45.38 31.93 43.30 55.79 matK 44.35 22.58 22.58 32.80 47.1 44.54 22.69 23.53 32.80 46.71 atpA 58.06 37.10 20.97 41.40 51.02 57.98 36.13 21.85 41.60 60.37 atpF 45.16 36.29 37.90 38.00 43.37 44.54 35.29 38.66 39.90 48.78 atpI 48.39 37.90 27.42 38.70 56.35 47.90 38.66 28.57 38.70 56.35 rps2 41.13 45.97 27.42 39.70 55.68 41.18 47.06 26.05 39.70 55.68 rpoC2 50.00 34.68 32.26 38.70 56.29 50.42 33.61 31.09 38.80 56.17 rpoC1 48.39 38.71 25.00 38.80 50.15 48.74 37.82 26.05 38.80 50.15 rpoB 41.94 31.45 24.19 39.70 56.80 42.86 31.93 23.53 39.70 56.73 psbD 45.97 48.39 30.65 43.50 54.60 45.38 48.74 31.09 43.50 54.60 psbC 57.26 43.55 35.48 44.90 53.61 57.98 42.86 36.97 43.70 43.75 rps14 43.93 49.07 29.63 42.00 50.66 43.56 47.52 31.68 42.00 50.66 psaB 56.45 45.16 28.23 42.20 48.20 57.14 45.38 26.89 42.20 48.14 psaA 50.00 43.55 37.90 43.60 52.35 49.58 44.54 37.82 43.50 52.29 ycf3 45.97 37.10 28.23 40.80 55.61 44.54 36.97 26.89 39.60 61.00 rps4 53.23 40.32 25.81 39.90 48.7 52.1 40.34 26.89 39.90 48.70 ndhJ 50.81 37.10 31.45 41.30 56.28 52.1 37.82 31.93 39.90 51.41 ndhK 40.32 49.19 22.58 38.10 58.39 40.34 49.58 21.01 36.90 47.24 ndhC 45.53 33.87 24.19 36.30 46.95 47.06 33.61 24.37 35.00 45.99 atpE 50.81 38.71 26.61 39.10 47.78 51.26 36.13 26.89 41.70 55.91 atpB 62.10 41.94 29.03 42.00 44.86 62.18 42.86 31.09 43.30 56.14 rbcL 54.84 47.58 27.42 44.20 48.16 54.62 48.74 27.73 44.20 47.96 accD 35.48 33.06 29.84 36.40 46.69 36.13 31.93 31.09 37.40 51.70 ycf4 38.71 42.74 29.03 38.00 46.79 38.66 44.54 30.25 38.00 55.97 cemA 34.68 23.39 37.10 33.50 52.34 34.45 22.69 38.66 33.50 52.34 petA 52.42 40.32 24.19 39.20 48.56 51.26 39.5 25.21 39.20 48.56 rps18 35.29 43.14 26.47 35.00 35.64 35.29 43.69 26.47 35.00 35.64 rpl20 38.98 43.22 25.42 39.00 41.33 38.98 43.22 25.42 39.00 41.33 clpP 50.81 36.29 27.42 40.60 52.48 57.14 37.82 25.21 41.80 54.64 psbB 50.81 47.58 38.71 44.20 47.41 51.26 48.74 40.34 44.00 47.14 petB 48.39 38.71 31.45 42.80 55.46 47.06 38.66 30.25 41.60 60.80 petD 48.39 40.32 30.65 39.50 44.62 51.26 36.13 26.05 41.20 44.37 rpoA 47.58 41.94 27.42 35.00 59.26 47.9 42.02 25.21 34.90 58.49 rps11 53.23 58.06 20.97 45.60 50.97 52.10 50.42 29.41 45.60 45.54 rps8 42.74 41.13 26.61 39.00 54.31 45.38 33.61 38.66 30.00 49.16 rpl14 56.10 36.59 26.02 41.70 55.31 51.26 45.38 20.17 41.70 53.41 rpl16 52.42 53.23 20.16 42.70 44.20 36.97 30.25 36.13 37.00 46.46 rps3 46.77 29.03 25.81 35.00 47.72 39.50 31.09 36.97 38.80 50.59 rpl22 43.55 38.71 22.58 36.70 57.04 52.10 57.98 20.17 45.60 50.97 ndhF 39.52 34.68 23.39 33.20 50.05 40.34 41.18 26.89 38.20 57.94 ccsA 29.84 33.87 24.19 33.80 59.41 56.30 37.82 26.05 41.70 55.31 ndhE 39.22 33.01 24.51 33.30 50.31 52.94 52.94 19.33 42.70 44.20 ndhG 41.13 33.06 25.00 33.80 61.00 46.22 27.73 26.05 34.60 44.18 ndhI 42.74 39.52 29.84 37.40 55.31 43.70 39.50 23.53 36.70 57.04 ndhA 44.35 37.10 21.77 34.00 41.23 40.34 34.45 22.69 31.70 41.66 ndhH 49.19 37.10 22.58 38.30 58.29 28.57 34.45 24.37 33.80 59.38 对小果油茶与越南油茶叶绿体基因组密码子不同位置上GC含量和ENC值进行关联性分析,结果表明:小果油茶密码子中GCall与GC1、GC2极显著相关,GC1与GC2极显著相关;越南油茶密码子中GCall与GC1、GC2极显著相关,GC1与GC2极显著相关,GC2与GC3显著相关,表明两物种叶绿体基因密码子第1、2位上的碱基组成相似(图1)。
![]() 图 1 密码子不同位置上GC占比及ENC值的关联性分析**表示极显著相关(P≤0.01),*表示显著相关(P≤0.05)。Figure 1. Correlation analysis of GC proportion and ENC value at different positions of codons
图 1 密码子不同位置上GC占比及ENC值的关联性分析**表示极显著相关(P≤0.01),*表示显著相关(P≤0.05)。Figure 1. Correlation analysis of GC proportion and ENC value at different positions of codons将RSCU值 > 1的密码子作为高频密码子。RSCU分析(表3)显示:小果油茶有28种高频密码子,其中以U或A结尾的分别有12、13个,以G或C结尾的分别有3、0个;越南油茶有30种高频密码子,其中以U或A结尾的分别有14、13个,以G或C结尾的分别有3、0个。因此两物种密码子都偏好以A或U结尾。
表 3 小果油茶和越南油茶不同基因密码子RSCU分析Table 3. RSCU analysis of different gene codons of Camellia meiocarpa and C. vietnamensis氨基酸 小果油茶 Camellia meiocarpa 越南油茶 C. vietnamensis 密码子 数目 RSCU 密码子 数目 RSCU 苯丙氨酸 Phe UUU 795 1.17 UUU 815 1.19 UUC 553 0.83 UUC 520 0.81 亮氨酸 Leu UUA 520 1.51 UUA 535 1.59 UUG 558 1.46 UUG 493 1.44 CUU 478 1.12 CUU 439 1.11 CUC 217 0.52 CUC 214 0.54 CUA 340 0.84 CUA 314 0.86 CUG 254 0.56 CUG 230 0.47 异亮氨酸 Ile AUU 720 1.27 AUU 732 1.33 AUC 436 0.73 AUC 422 0.75 AUA 581 0.99 AUA 548 0.92 蛋氨酸 Met AUG 513 0.96 AUG 463 0.92 缬氨酸 Val GUU 367 1.32 GUU 366 1.44 GUC 172 0.75 GUC 181 0.7 GUA 336 1.21 GUA 312 1.11 GUG 204 0.71 GUG 174 0.69 丝氨酸 Ser UCU 362 1.2 UCU 405 1.04 UCC 250 0.84 UCC 264 0.85 UCA 405 1.65 UCA 422 1.49 UCG 184 0.87 UCG 200 0.81 脯氨酸 Pro CCU 230 1.06 CCU 248 1.16 CCC 146 0.85 CCC 126 0.65 CCA 239 1.19 CCA 265 1.3 CCG 143 0.91 CCG 145 0.88 苏氨酸 Thr ACU 235 1.01 ACU 233 1 ACC 169 0.8 ACC 174 0.73 ACA 297 1.41 ACA 299 1.02 ACG 133 0.79 ACG 133 0.89 丙氨酸 Ala GCU 196 1.1 GCU 233 1.17 GCC 109 0.77 GCC 117 0.99 GCA 188 1.11 GCA 192 1.04 GCG 89 0.87 GCG 76 0.79 酪氨酸 Tyr UAU 595 1.28 UAU 622 1.32 UAC 319 0.68 UAC 313 0.68 终止密码子 TER UAA 403 1.18 UAA 407 1.06 UAG 340 0.91 UAG 365 0.96 组氨酸 His CAU 288 1.26 CAU 315 1.18 CAC 154 0.66 CAC 161 0.66 谷氨酰胺 Gln CAA 446 1.31 CAA 430 1.37 CAG 249 0.69 CAG 215 0.63 天冬酰胺 Asn AAU 636 1.32 AAU 575 1.25 AAC 313 0.68 AAC 322 0.71 赖氨酸 Lys AAA 831 1.25 AAA 824 1.36 AAG 459 0.75 AAG 437 0.64 天冬氨酸 Asp GAU 447 1.31 GAU 451 1.43 GAC 166 0.66 GAC 152 0.57 谷氨酸 Glu GAA 592 1.37 GAA 612 1.33 GAG 280 0.63 GAG 277 0.67 半胱氨酸 Cys UGU 235 0.99 UGU 227 1.04 UGC 232 0.86 UGC 235 0.84 终止密码子 TER UGA 336 0.92 UGA 407 0.98 色氨酸 Trp UGG 421 0.96 UGG 428 0.94 精氨酸 Arg CGU 152 0.7 CGU 149 0.86 CGC 93 0.45 CGC 83 0.4 CGA 214 1.11 CGA 224 1.08 CGG 143 0.74 CGG 147 0.73 丝氨酸 Ser AGU 205 0.85 AGU 227 1.01 AGC 186 0.59 AGC 164 0.57 精氨酸 Arg AGA 383 1.79 AGA 406 1.69 AGG 263 1.81 AGG 251 1.34 甘氨酸 Gly GGU 237 0.9 GGU 249 0.88 GGC 164 0.54 GGC 161 0.56 GGA 333 1.16 GGA 404 1.28 GGG 300 1.33 GGG 284 1.28 注:加粗的表示高频密码子(RSCU > 1)。 2.2 中性绘图分析
中性绘图分析(图2)显示:小果油茶和越南油茶的GC12的取值范围分别为29.0% ~ 55.7%、28.6% ~ 55.0%,GC3的取值范围分别为20.1% ~ 38.7%、19.3% ~ 40.3%;小果油茶和越南油茶的回归曲线斜率分别为-0.137 86和-0.136 17,GC12与GC3呈负相关关系,基因大量分布在坐标轴对角线上方,说明两物种叶绿体基因组密码子使用偏好性受自然选择影响较大。
![]() 图 2 叶绿体基因密码子中性绘图GC12.GC1和GC2的平均值;GC3.基因中所有密码子第3位碱基G + C含量(除蛋氨酸、色氨酸和终止密码子外)。Figure 2. Codon neutrality mapping of chloroplast genes
图 2 叶绿体基因密码子中性绘图GC12.GC1和GC2的平均值;GC3.基因中所有密码子第3位碱基G + C含量(除蛋氨酸、色氨酸和终止密码子外)。Figure 2. Codon neutrality mapping of chloroplast genes2.3 ENC-plot分析
ENC-plot分析(图3)显示:小果油茶和越南油茶基因总体分布均较为分散,较多基因距离标准曲线较远且分布于标准曲线下方,表明两物种的密码子使用偏好性受自然选择压力的影响较大。通过计算ENC比值,进一步量化ENC检测值与ENC标准值之间的差异。结果(表4)显示:−0.05 ~ 0.05区间内,小果油茶有14个基因,约占26.9%;越南油茶有18个基因,约占34.6%,这部分基因ENC检测值与ENC标准值较为接近,而小果油茶(73.1%)和越南油茶(65.4%)大部分基因ENC检测值与ENC标准值差异比较大,进一步表明两个物种的密码子使用偏好性主要受自然选择影响。
![]() 图 3 叶绿体基因密码子ENC-plot分析ENC. 有效密码子数;GC3. 基因中所有密码子第3位碱基GC含量。Figure 3. ENC-plot analysis of chloroplast gene codons表 4 ENC比值的频率分布Table 4. Frequency distribution of ENC ratios
图 3 叶绿体基因密码子ENC-plot分析ENC. 有效密码子数;GC3. 基因中所有密码子第3位碱基GC含量。Figure 3. ENC-plot analysis of chloroplast gene codons表 4 ENC比值的频率分布Table 4. Frequency distribution of ENC ratios小果油茶 Camellia meiocarpa 越南油茶 C. vietnamensis 组段 组中值 组数 频率/% 组段 组中值 组数 频率/% −0.15 ~ −0.05 −0.1 2 3.8 −0.15-−0.05 −0.1 4 7.7 −0.05 ~ 0.05 0 14 26.9 −0.05-0.05 0 18 34.6 0.05 ~ 0.15 0.1 27 51.9 0.05-0.15 0.1 21 40.4 0.15 ~ 0.25 0.2 6 11.5 0.15-0.25 0.2 6 11.5 0.25 ~ 0.35 0.3 3 5.8 0.25-0.35 0.3 3 5.8 合计 52 100 合计 52 100 2.4 PR2-plot分析
PR2-plot分析结果(图4)显示:小果油茶和越南油茶的密码子基因大部分分布在右上角,密码子第3位碱基嘧啶碱(A 和 G)的使用频率高于嘌呤碱(T和C),这表明两物种密码子第3位碱基分布不均匀,可以推测自然选择影响了小果油茶和越南油茶叶绿体基因的密码子使用偏好性。
![]() 图 4 叶绿体基因密码子PR2-plot分析A3、T3、G3、C3代表基因中所有密码子第3位碱基A、T、G、C的含量。Figure 4. Analysis of codon PR2-plot of chloroplast genes
图 4 叶绿体基因密码子PR2-plot分析A3、T3、G3、C3代表基因中所有密码子第3位碱基A、T、G、C的含量。Figure 4. Analysis of codon PR2-plot of chloroplast genes2.5 最优密码子分析
在小果油茶中选择5个高表达基因(ycf4、accD、rps7、atpB、petD)和5个低表达基因(ndhG、ccsA、rpoA、ndhK、ndhH),在越南油茶中选择5个高表达基因(psbC、ndhA、rpl2、rpl20、rps18)和5个低表达基因(ycf3、petB、atpA、ndhH、rpoA)分别进行分析,构建两物种基因的高低表达库(表5)。以RSCU > 1且△RSCU ≥ 0.08为标准筛选最优密码子,其中小果油茶有23个最优密码子,分别为UUU、UUA、UUG、CUU、GUU、GUA、UCU、UCA、CCA、ACU、ACA、GCU、GCA、UAU、UAA、CAU、CAA、AAU、AAA、GAU、GAA、CGA、AGA,以U结尾的有10个,以A结尾的有12个,以G结尾的有1个,以C结尾的有0个。越南油茶有11个最优密码子,分别为UUU、AUU、GUU、UCA、CCU、ACU、GCU、UAA、CAU、AAU、UGU,以U结尾的有9个,以A结尾的有2个,以G、C结尾的有0个。小果油茶和越南油茶的最优密码子均偏好以A或U结尾,这与前面RSCU分析的结果(表3)相一致。
表 5 小果油茶和越南油茶叶绿体基因组的最优密码子分析Table 5. Optimal codons analysis in chloroplast genome of Camellia meiocarpa and C. vietnamensis氨基酸 密码子 小果油茶Camellia meiocarpa 密码子 越南油茶C. vietnamensis 高表达基因 低表达基因 △RSCU 高表达基因 低表达基因 △RSCU 数目 RSCU 数目 RSCU 数目 RSCU 数目 RSCU Phe UUU* 46 1.32 75 1.18 0.14 UUU* 75 1.28 19 1.15 0.13 UUC 26 0.68 46 0.82 −0.14 UUC 40 1.04 20 1.22 −0.18 Leu UUA*** 50 2.12 51 1.47 0.65 UUA 56 1.13 27 1.30 −0.18 UUG* 40 1.43 40 1.21 0.22 UUG 35 0.78 25 0.83 −0.05 CUU** 35 1.24 27 0.81 0.43 CUU 15 0.60 55 0.93 −0.33 CUC 11 0.32 25 0.68 −0.36 CUC 15 1.20 34 1.13 0.07 CUA 22 0.60 33 1.02 −0.42 CUA 13 0.60 59 0.87 −0.27 CUG 10 0.29 25 0.80 −0.51 CUG 8 0.40 49 1.07 −0.67 Ile AUU 60 1.46 72 1.63 −0.17 AUU* 14 1.20 31 0.98 0.22 AUC 38 0.65 26 0.59 0.06 AUC 9 0.60 48 0.95 −0.35 AUA* 46 0.89 37 0.78 0.10 AUA 3 1.00 28 1.00 0.00 Met AUG 45 1.00 45 1.00 0.00 AUG* 12 1.20 46 1.07 0.13 Val GUU* 44 1.39 29 1.23 0.15 GUU* 50 0.86 13 0.64 0.22 GUC 9 0.36 9 0.52 −0.16 GUC 19 0.79 35 0.78 0.01 GUA* 54 1.62 30 1.52 0.10 GUA 9 1.08 83 1.05 0.03 GUG 29 0.63 17 0.72 −0.10 GUG 42 1.05 13 1.03 0.02 Ser UCU* 22 1.31 10 1.14 0.17 UCU 34 1.32 11 1.37 −0.05 UCC*** 16 1.49 2 0.40 1.09 UCC 41 0.83 5 1.44 −0.61 UCA** 16 1.41 19 1.04 0.37 UCA* 32 1.01 19 0.84 0.17 UCG 9 0.51 6 0.93 −0.43 UCG** 34 1.12 6 0.66 0.46 Pro CCU 23 0.91 6 1.24 −0.33 CCU* 16 0.91 5 0.80 0.11 CCC 15 0.92 13 1.25 −0.33 CCC*** 5 2.00 23 1.01 0.99 CCA*** 18 1.31 3 0.80 0.51 CCA 4 0.80 17 0.91 −0.11 CCG 16 0.86 8 0.86 0.01 CCG 6 0.68 8 1.12 −0.44 Thr ACU*** 28 1.61 5 0.95 0.66 ACU* 5 1.13 23 1.00 0.13 ACC 9 0.32 14 1.24 −0.92 ACC 8 1.07 14 1.13 −0.06 ACA*** 36 1.87 8 0.96 0.91 ACA 3 0.80 13 1.05 −0.25 ACG 5 0.19 13 1.03 −0.84 ACG* 7 1.08 8 0.83 0.25 Ala GCU** 30 1.10 12 0.80 0.30 GCU** 22 1.13 5 0.80 0.33 GCC 18 0.94 18 1.30 −0.36 GCC 20 0.80 17 1.17 −0.37 GCA* 29 1.23 16 1.02 0.22 GCA 18 1.03 31 1.02 0.01 GCG 15 0.73 6 1.12 −0.39 GCG 21 0.93 12 1.20 −0.27 Tyr UAU** 33 1.73 48 1.33 0.40 UAU 25 0.68 15 1.09 −0.42 UAC 7 0.27 51 1.12 −0.85 UAC* 43 1.09 20 1.00 0.09 TER UAA* 15 1.44 61 1.29 0.14 UAA* 22 1.11 24 0.97 0.14 UAG 7 0.13 51 1.12 −0.99 UAG 26 0.92 15 1.04 −0.12 His CAU** 12 1.70 29 1.27 0.43 CAU* 36 1.13 80 0.87 0.27 CAC 4 0.30 12 0.73 −0.43 CAC*** 14 1.25 19 0.69 0.55 Gln CAA** 33 1.59 47 1.27 0.32 CAA 9 0.90 31 0.91 −0.01 CAG 14 0.41 29 0.73 −0.32 CAG 19 0.87 43 1.13 −0.27 Asn AAU* 41 1.55 38 1.34 0.21 AAU* 15 1.20 62 1.03 0.17 AAC 17 0.45 18 0.66 −0.21 AAC* 18 1.11 43 1.01 0.11 Lys AAA** 67 1.53 45 1.22 0.32 AAA 9 0.89 33 0.99 −0.11 AAG 20 0.47 27 0.78 −0.32 AAG 1 0.60 35 0.98 −0.38 Asp GAU** 32 1.80 14 1.40 0.40 GAU 21 0.77 17 1.11 −0.34 GAC 8 0.20 9 0.60 −0.40 GAC* 40 1.12 33 1.03 0.10 Glu GAA* 51 1.44 27 1.16 0.28 GAA 28 0.98 45 1.03 −0.04 GAG 16 0.56 25 0.84 −0.28 GAG 16 0.83 18 0.89 −0.06 Cys UGU*** 8 1.22 6 0.69 0.53 UGU*** 30 1.66 6 0.67 0.99 UGC 4 0.38 10 1.31 −0.93 UGC 59 1.19 12 1.22 −0.03 TER UGA*** 15 1.44 25 0.58 0.85 UGA 20 0.72 27 1.10 −0.37 Trp UGG 21 0.80 29 1.00 −0.20 UGG 11 0.53 7 0.65 −0.12 Arg CGU*** 26 2.05 3 0.44 1.61 CGU** 32 1.44 19 1.03 0.42 CGC 4 0.22 8 0.92 −0.70 CGC 3 0.43 28 1.53 −1.10 CGA** 20 1.48 8 1.06 0.41 CGA 6 1.03 18 1.03 0.00 CGG 2 0.10 10 1.13 −1.03 CGG 19 1.11 8 1.04 0.07 Ser AGU** 15 0.98 8 0.52 0.46 AGU 7 0.80 30 0.94 −0.14 AGC 6 0.30 9 0.74 −0.43 AGC 16 1.24 11 1.17 0.08 Arg AGA* 27 1.74 10 1.53 0.21 AGA 4 0.87 11 0.91 −0.04 AGG 7 0.42 8 0.92 −0.50 AGG 3 0.21 22 0.99 −0.77 Gly GGU*** 43 1.54 10 0.95 0.59 GGU 11 0.66 16 0.88 −0.23 GGC 5 0.21 7 0.46 −0.25 GGC* 29 0.87 18 0.73 0.14 GGA 33 1.22 16 1.18 0.04 GGA 23 0.55 29 0.75 −0.20 GGG 28 1.03 3.8 1.41 −0.37 GGG 25 1.20 22 1.29 −0.09 注:*表示0.08 ≤ △RSCU < 0.3,**表示0.3 ≤ △RSCU < 0.5,***表示△RSCU ≥ 0.5,加粗表示最优密码子。 3. 讨 论
叶绿体基因组具有母系遗传的特点,即遗传信息通过母本传递给后代,其序列信息可以揭示物种的起源、进化和亲缘关系,具有进化速率慢、序列较为保守、易受到选择压力影响等特性[34]。叶绿体基因组的密码子偏好性可能受到自然选择、基因突变和基因表达水平等多种因素的影响[35],研究叶绿体基因组的密码子偏好性不仅有助于深入了解植物的遗传特性和进化历程,还能提高基因表达效率,推动基因工程研究与应用,揭示植物适应性与环境响应机制,促进植物资源保护与利用[36],以及拓展生物信息学应用,对植物科学研究和农业生产实践具有重要意义。
研究叶绿体基因组密码子偏好性的过程中,最直观体现偏好性的是密码子使用的频率和碱基的分布,GC3常用于衡量密码子偏好性[37]。本研究小果油茶与越南油茶的GC3含量为28.64%和28.59%,该结果与普通核桃(Juglans regia)、深纹核桃(Juglans sigillate)[31]等相近。前人研究[38−40]表明,单子叶植物偏好G/C结尾,双子叶植物偏好A/T(U)结尾,小果油茶与越南油茶的研究结果符合双子叶植物的这一特性,由此推出,同类物种在进化的过程中遵循着相似的规律。中性绘图结果表明,小果油茶与越南油茶的回归系数分别为−
0.13786 和−0.13617 ,接近于0,表明两物种的密码子偏好性更多受到自然选择的影响,该结果与王鹏良等[11]在普通油茶和郝丙青等[12]在香花油茶中的相关结果一致,这一结果也证实了叶绿体基因组具有进化速率慢、序列较为保守、易受到选择压力影响的特性。结合ENC-plot与PR2-plot分析的结果同样发现,小果油茶与越南油茶叶绿体基因偏好性主要受自然选择的影响,该结果与北沙参(Glehnia littoralis)[41]、葫芦巴(Trigonella foenum-graecum)[42]、甜高粱(Sorghum bicolor)[43]的研究结果一致,与大部分双子叶植物的研究结果保持一致。近年来,国家重视粮油安全,大力发展木本油料作物,油茶作为明星树种,得以大力推广,但油茶病害、虫害日益严重。因此,培育抗病良种有益于油茶提质增产,研究油茶叶绿体基因组可为培育抗病良种助力,进而提高油茶抗逆性。本文对小果油茶和越南油茶叶绿体基因组密码子偏好性进行分析,发现两个物种在碱基组成分析、中性绘图、ENC-plot分析、PR2-plot分析和相关性分析等方面都呈现出极强的相似性,也进一步说明小果油茶与越南油茶存在一定的亲缘关系。该研究结果为未来实现小果油茶与越南油茶优良性状的结合提供了理论依据,通过优化目标基因的密码子,可显著提升基因表达效率,从而为培育更具市场竞争力的优良油茶品种奠定坚实基础。鉴于资源和研究时间的限制,本文聚焦于叶绿体基因组这一关键领域,进行了深入的分析与研究。此外,叶绿体基因组的母系遗传特性使其在某些育种策略中具有独特的优势。通过研究叶绿体基因组的密码子偏好性,能够更深入地挖掘这一特性在育种中的应用潜力。与此同时,植物细胞核基因组在育种过程中也起着至关重要的作用。在未来的研究中,我们将对叶绿体基因组和核基因组展开综合分析,以获取更全面且深入的遗传信息,为育种研究提供更有力的支持。
4. 结 论
小果油茶和越南油茶叶绿体基因组密码子碱基使用偏好性不强,其偏好性主要受到自然选择的影响。本研究在小果油茶中筛选到23个最优密码子,越南油茶中有11个最优密码子。研究成果为进一步探索油茶基因组研究和良种培育提供了坚实的理论依据和数据支撑。
-
![]()
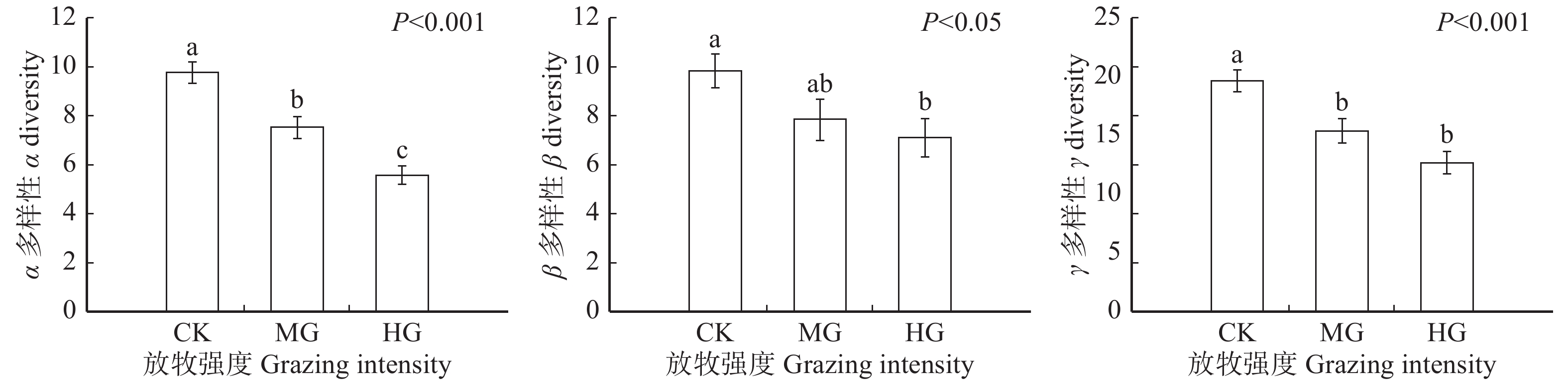
图 1 科尔沁沙地植物群落α、β、γ多样性对放牧干扰的响应
CK为围封;MG为中度放牧;HG为重度放牧。CK stands for grazing exclusion; MG stands for moderate grazing; HG stands for heavy grazing.
Figure 1. Responses of α, β and γ diversity to grazing disturbance in the Horqin Sandy Land
![]()
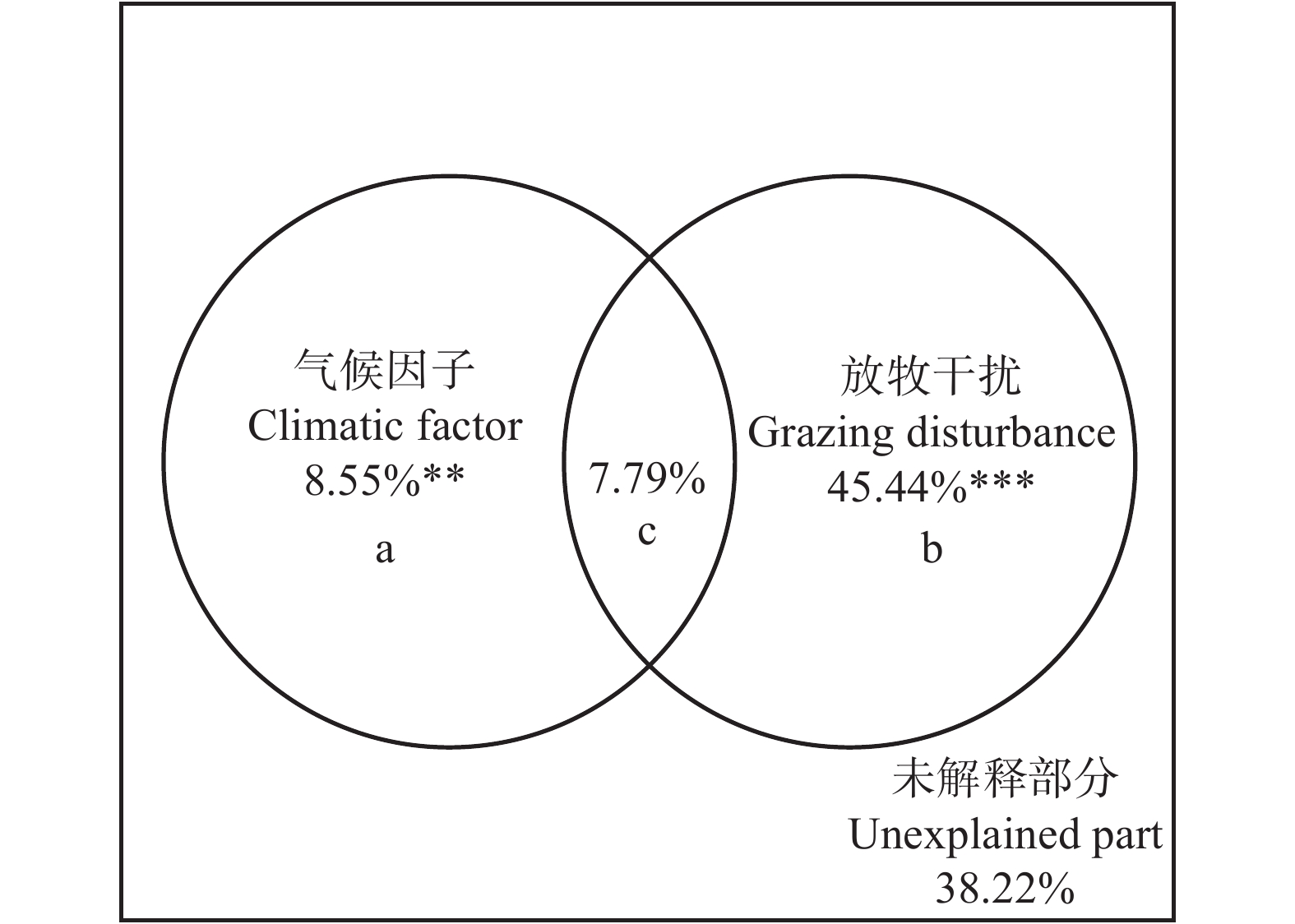
图 2 放牧干扰和环境因子对科尔沁沙地植物群落α多样性的解释
*** P < 0.001;** P < 0.01。a为气候因子的单独解释率;b为放牧干扰的单独解释率;c为气候因子与放牧干扰的共同解释率。下同。*** means P < 0.001; ** means P < 0.01. a, the independent explanation rate of climatic factors; b, the independent explanation rate of grazing disturbance; c, the co-explanation rate of climatic factors and grazing disturbance.The same below.
Figure 2. Explanation of grazing disturbance and environmental factors to α diversity of plant communities in the Horqin Sandy Land
![]()
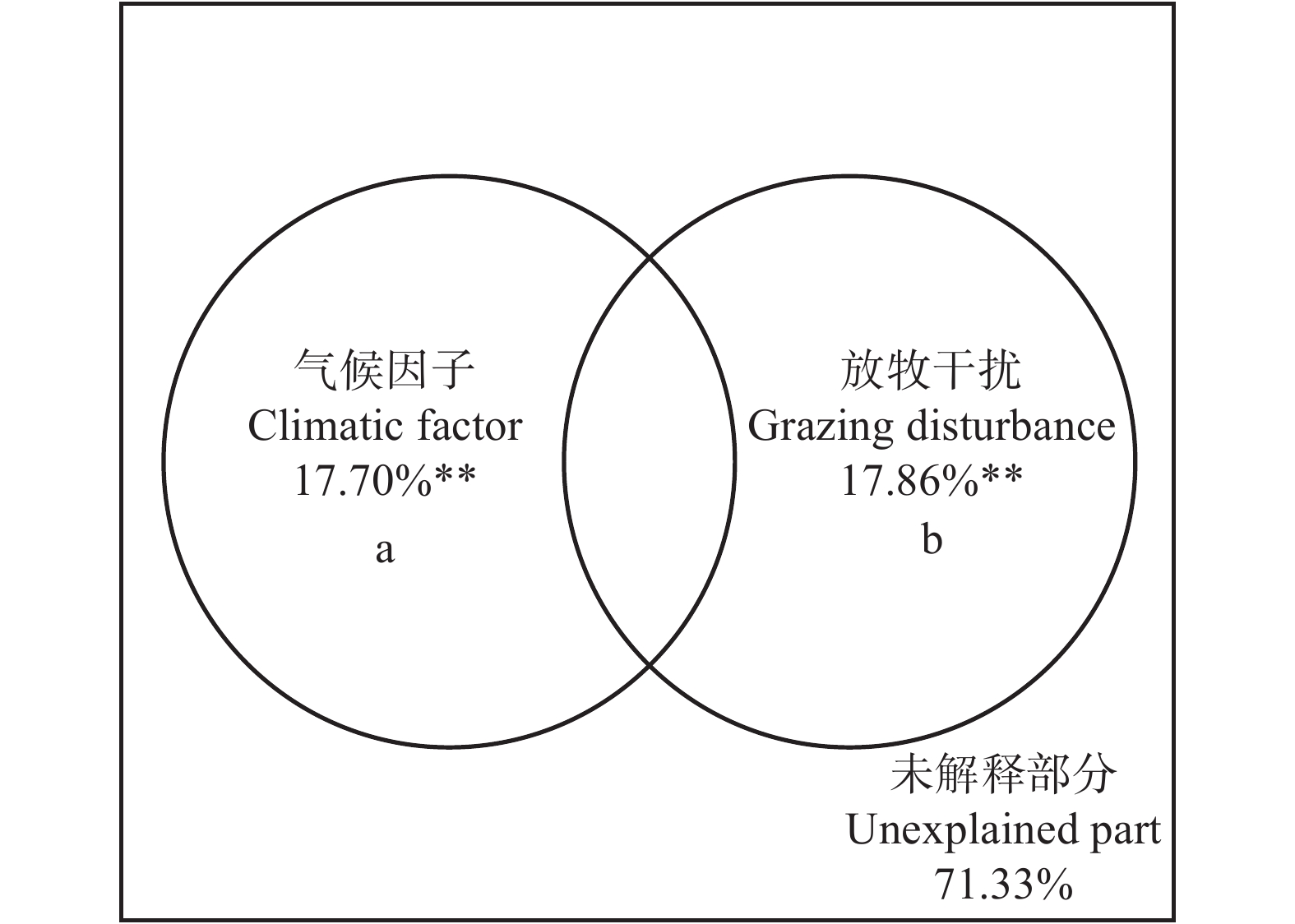
图 3 放牧干扰和环境因子对科尔沁沙地植物群落β多样性的解释
Figure 3. Explanation of grazing disturbance and climatic factors to β diversity of plant communities in the Horqin Sandy Land
![]()
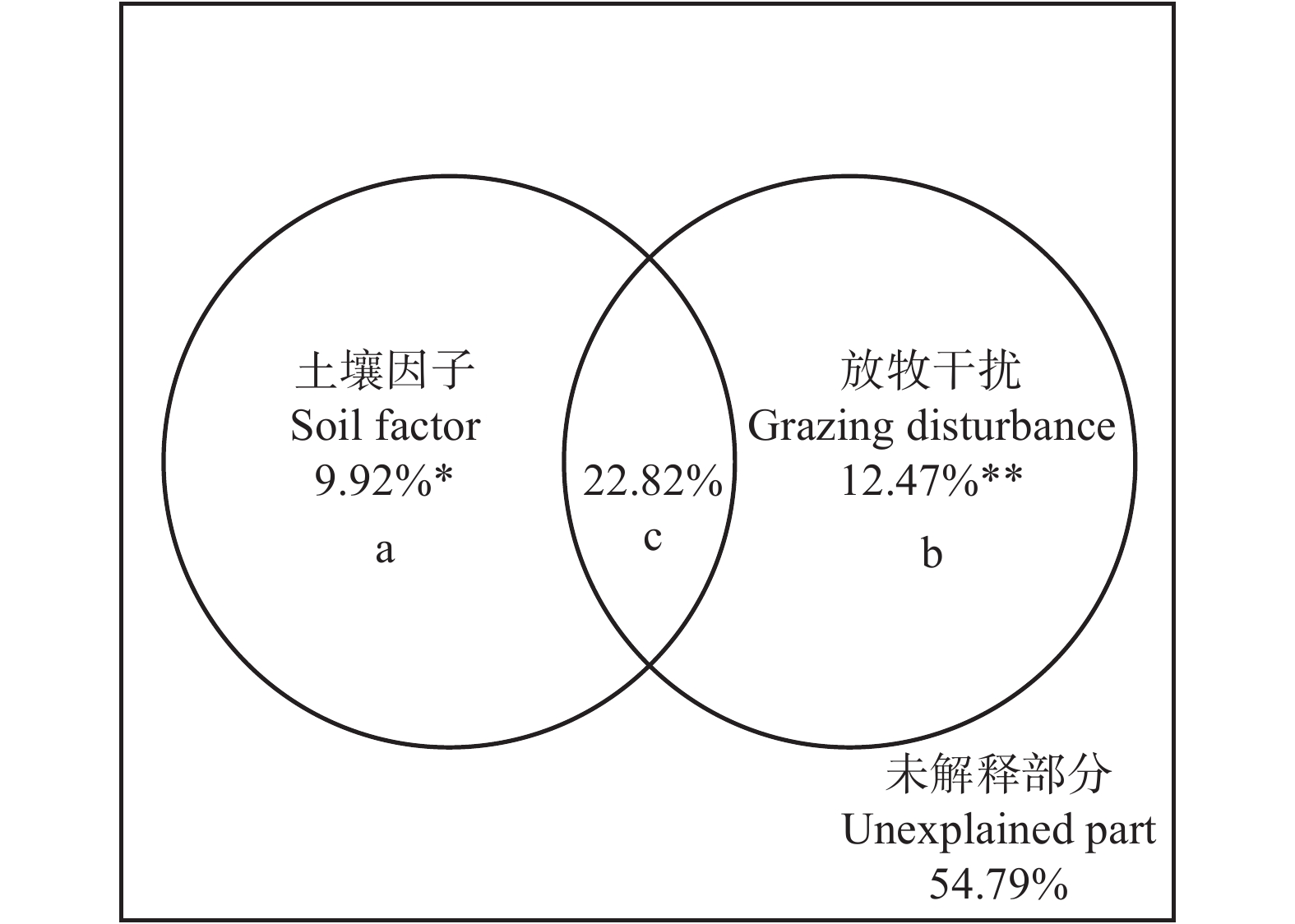
图 4 放牧干扰和环境因子对科尔沁沙地植物群落γ多样性的解释
** P < 0.01;* P < 0.05。a为土壤因子的单独解释率;b为放牧干扰的单独解释率;c为土壤因子与放牧干扰的共同解释率。** means P < 0.01; * means P < 0.05. a, the independent explanation rate of soil factors; b, the independent explanation rate of grazing disturbance; c, the co-explanation rate of soil factors and grazing disturbance.
Figure 4. Explanation of grazing disturbance and environmental factors to γ diversity of plant communities in the Horqin Sandy Land
表 1 科尔沁沙地植物物种组成(分布频率 > 15%)
Table 1 Plant species composition in the Horqin Sandy Land (distribution frequency > 15%)
种名
Species name科
Family属
Genus生活型
Life form分布频率
Distribution frequency/%小叶锦鸡儿
Caragana microphylla豆科
Leguminosae锦鸡儿属
Caragana灌木
Shrub62 狗尾草
Setaria viridis禾本科
Gramineae狗尾草属
Setaria一年生草本
Annual herb61 地梢瓜
Cynanchum thesioides萝藦科
Asclepiadaceae鹅绒藤属
Cynanchum半灌木
Subshrub58 兴安虫实
Corispermum chinganicum藜科
Chenopodiaceae虫实属
Corispermum一年生草本
Annual herb51 胡枝子
Lespedeza bicolor豆科
Leguminosae胡枝子属
Lespedeza小灌木
Dwarf shrub49 盐蒿
Artemisia halodendron菊科
Compositae蒿属
Artemisia小灌木
Dwarf shrub42 蒺藜
Tribulus terrester蒺藜科
Zygophyllaceae蒺藜属
Tribulus一年生草本
Annual herb37 止血马唐
Digitaria ischaemum禾本科
Gramineae马唐属
Digitaria一年生草本
Annual herb30 雾冰藜
Bassia dasyphylla藜科
Chenopodiaceae雾冰藜属
Bassia一年生草本
Annual herb30 尖头叶藜
Chenopodium acuminatum藜科
Chenopodiaceae藜属
Chenopodium一年生草本
Annual herb26 猪毛菜
Salsola collina藜科
Chenopodiaceae猪毛菜属
Salsola一年生草本
Annual herb26 山竹岩黄耆
Hedysarum fruticosum豆科
Leguminosae岩黄耆属
Hedysarum半灌木或小半灌木
Subshrub or small subshrub22 沙蓬
Agriophyllum squarrosum藜科
Chenopodiaceae沙蓬属
Agriophyllum一年生草本
Annual herb20 虎尾草
Chloris virgata禾本科
Gramineae虎尾草属
Chloris一年生草本
Annual herb19 画眉草
Eragrostis pilosa禾本科
Gramineae画眉草属
Eragrostis一年生草本
Annual herb19 大果榆
Ulmus macrocarpa榆科
Ulmaceae榆属
Ulmus乔木
Tree18 光梗蒺藜草
Cenchrus calyculatus禾本科
Gramineae蒺藜草属
Cenchrus一年生草本
Annual herb17 黑沙蒿
Artemisia ordosica菊科
Compositae蒿属
Artemisia小灌木
Dwarf shrub17 地锦
Euphorbia humifusa大戟科
Euphorbiaceae大戟属
Euphorbia一年生草本
Annual herb16  下载: 导出CSV
下载: 导出CSV
表 2 放牧干扰、环境因子与科尔沁沙地植物群落α、β、γ多样性的相关性分析
Table 2 Correlation analysis among grazing disturbance, environmental factors, and α, β, γ diversity of plant communities in the Horqin Sandy Land
项目
Itemα多样性
α diversityβ多样性
β diversityγ多样性
γ diversityGI 0.532 3(−)*** 0.109 7(−)* 0.352 8(−)*** MAP 0.143 3(US)* 0.107 9(+)* PET 0.100 1(−)* MAT MTWM 0.192 0(−)** MTCM 0.126 3(HS)* 0.108 1(−)* MDR 0.163 3(+)** SOC 0.239 4(+)*** 0.173 4(+)** STN 0.348 5(HS)*** 0.221 5(+)*** STP 0.187 9(+)** pH 注:*** P < 0.001;** P < 0.01;* P < 0.05;空白代表无显著的相关性;−表示负相关关系;+表示正相关关系;HS代表先增后减的二次曲线关系;US代表先减后增的二次曲线关系。GI为放牧强度;MAP为年均降水量;PET为潜在蒸散量;MAT为年均温度;MTWM为最热月均温;MTCM为最冷月均温;MDR为气温日较差;SOC为土壤有机碳;STN为土壤全氮;STP为土壤全磷。Notes: *** means P < 0.001; ** means P < 0.01; * means P < 0.05. The blank means no significant correlation. − stands for a negative correlation; + stands for a positive correlation; HS stands for a hump-shaped relationship of increasing first and then decreasing; US stands for a U-shaped relationship of decreasing first and then increasing. GI, grazing intensity; MAP, mean annual precipitation; PET, potential evapotranspiration; MAT, mean annual temperature; MTWM, mean temperature of the warmest month; MTCM, mean temperature of the coldest month; MDR, mean diurnal range; SOC, soil organic carbon; STN, soil total nitrogen; STP, soil total phosphorus.
下载: 导出CSV
-
[1] 马克平. 生物多样性科学的热点问题[J]. 生物多样性, 2016, 24(1):1−2. doi: 10.17520/biods.2016029 Ma K P. Hot topics for biodiversity science[J]. Biodiversity Science, 2016, 24(1): 1−2. doi: 10.17520/biods.2016029
[2] 孟庆兰. 农牧交错区植被生产力对降水波动的响应: 以科尔沁地区为例[D]. 烟台: 鲁东大学, 2017. Meng Q L. The relationship between vegetation production and rainfall fluctuation in Horqin area[D]. Yantai: Ludong University, 2017.
[3] 左小安, 赵学勇, 赵哈林, 等. 科尔沁沙地草地退化过程中的物种组成及功能多样性变化特征[J]. 水土保持学报, 2006, 20(1):181−185. doi: 10.3321/j.issn:1009-2242.2006.01.044 Zuo X A, Zhao X Y, Zhao H L, et al. Changes on species composition and function diversity in degraded process of grassland in Horqin Sand Land[J]. Journal of Soil and Water Conservation, 2006, 20(1): 181−185. doi: 10.3321/j.issn:1009-2242.2006.01.044
[4] Austrheim G, Eriksson O. Plant species diversity and grazing in the Scandinavian Mountains: patterns and processes at different spatial scales[J]. Ecography, 2001, 24: 683−695. doi: 10.1034/j.1600-0587.2001.240607.x
[5] 唐志尧, 方精云. 植物物种多样性的垂直分布格局[J]. 生物多样性, 2004, 12(1):20−28. doi: 10.3321/j.issn:1005-0094.2004.01.004 Tang Z Y, Fang J Y. A review on the elevational patterns of plant species diversity[J]. Biodiversity Science, 2004, 12(1): 20−28. doi: 10.3321/j.issn:1005-0094.2004.01.004
[6] Kraft N J B, Comita L S, Chase J M, et al. Disentangling the drivers of β diversity along latitudinal and elevational gradients[J]. Science, 2011, 333: 1755−1758. doi: 10.1126/science.1208584
[7] 王兴, 宋乃平, 杨新国, 等. 放牧扰动下草地植物多样性对土壤因子的响应[J]. 草业学报, 2013, 22(5):27−36. doi: 10.11686/cyxb20130504 Wang X, Song N P, Yang X G, et al. The reponse of grassland plant diversity to soil factors under grazing disturbance[J]. Acta Prataculturae Sinica, 2013, 22(5): 27−36. doi: 10.11686/cyxb20130504
[8] 刘文亭, 卫智军, 吕世杰, 等. 放牧对短花针茅荒漠草原植物多样性的影响[J]. 生态学报, 2017, 37(10):3394−3402. Liu W T, Wei Z J, Lü S J, et al. The impacts of grazing on plant diversity in Stipa breviflora desert grassland[J]. Acta Ecologica Sinica, 2017, 37(10): 3394−3402.
[9] 董全民, 赵新全, 马玉寿, 等. 牦牛放牧强度对小嵩草草甸两季轮牧草场植物群落数量特征的影响[J]. 生态学杂志, 2011, 30(10):2233−2239. Dong Q M, Zhao X Q, Ma Y S, et al. Effects of yak grazing intensity on quantitative characteristics of plant community in a two-seasonal rotation pasture in Kobresia parva meadow[J]. Chinese Journal of Ecology, 2011, 30(10): 2233−2239.
[10] 张继义, 赵哈林, 张铜会, 等. 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态[J]. 植物生态学报, 2004, 28(1):86−92. doi: 10.3321/j.issn:1005-264X.2004.01.013 Zhang J Y, Zhao H L, Zhang T H, et al. Dynamics of species diversity of communities in restoration processes in Horqin Sandy Land[J]. Acta Phytoecologica Sinica, 2004, 28(1): 86−92. doi: 10.3321/j.issn:1005-264X.2004.01.013
[11] 李玉霖, 孟庆涛, 赵学勇, 等. 科尔沁沙地流动沙丘植被恢复过程中群落组成及植物多样性演变特征[J]. 草业学报, 2007, 16(6):54−61. doi: 10.3321/j.issn:1004-5759.2007.06.008 Li Y L, Meng Q T, Zhao X Y, et al. Characteristics of species composition and plant diversity in the process of vegetation restoration on moving dunes in the Kerqin Sandy Land[J]. Acta Prataculturae Sinica, 2007, 16(6): 54−61. doi: 10.3321/j.issn:1004-5759.2007.06.008
[12] 赵丽娅, 钟韩珊, 赵美玉, 等. 围封和放牧对科尔沁沙地群落物种多样性与地上生物量的影响[J]. 生态环境学报, 2018, 27(10):1783−1790. Zhao L Y, Zhong H S, Zhao M Y, et al. Effect of enclosure and grazed management on aboveground biomass and species diversity in sandy grasslands of Horqin Sandy Land, eastern Inner Mongolia, China[J]. Ecology and Environmental Sciences, 2018, 27(10): 1783−1790.
[13] 吕朋, 左小安, 张婧, 等. 放牧强度对科尔沁沙地沙质草地植被的影响[J]. 中国沙漠, 2016, 36(1):34−39. doi: 10.7522/j.issn.1000-694X.2015.00197 Lü P, Zuo X A, Zhang J, et al. Effects of grazing intensity on vegetation in sandy grassland of Horqin[J]. Journal of Desert Research, 2016, 36(1): 34−39. doi: 10.7522/j.issn.1000-694X.2015.00197
[14] 赵哈林, 赵学勇, 张铜会, 等. 科尔沁沙地沙漠化过程及其恢复机理[M]. 北京: 海洋出版社, 2009. Zhao H L, Zhao X Y, Zhang T H , et al. Desertification processes and its restoration mechanisms in the Horqin Sand Land[M]. Beijing: Ocean Press, 2009.
[15] 左小安, 赵学勇, 张铜会, 等. 科尔沁沙地榆树疏林草地物种多样性及乔木种群空间格局[J]. 干旱区资源与环境, 2005, 19(4):63−68. doi: 10.3969/j.issn.1003-7578.2005.04.013 Zuo X A, Zhao X Y, Zhang T H, et al. Species diversity and arbor population distribution pattern of Ulmus pumila L. scattered grassland of Horqin Sand[J]. Journal of Arid Land Resources and Environment, 2005, 19(4): 63−68. doi: 10.3969/j.issn.1003-7578.2005.04.013
[16] 任鸿昌, 吕永龙, 杨萍, 等. 科尔沁沙地土地沙漠化的历史与现状[J]. 中国沙漠, 2004, 24(5):544−547. doi: 10.3321/j.issn:1000-694X.2004.05.005 Ren H C, Lü Y L, Yang P, et al. History and present status of desertification in Horqin Sandy Land region[J]. Journal of Desert Research, 2004, 24(5): 544−547. doi: 10.3321/j.issn:1000-694X.2004.05.005
[17] 曹文梅. 科尔沁沙地植被群落结构及其空间格局与生境因子响应关系研究[D]. 呼和浩特: 内蒙古农业大学, 2017. Cao W M. Research for the response relationships of habitat factors to community structure and spatial pattern of vegetation in the Horqin Sandy Land[D]. Huhhot: Inner Mongolia Agricultural University, 2017.
[18] 曹成有, 蒋德明, 朱丽辉, 等. 科尔沁沙地草甸草场退化的原因与植物多样性变化[J]. 草业学报, 2006, 15(3):18−26. doi: 10.3321/j.issn:1004-5759.2006.03.003 Cao C Y, Jiang D M, Zhu L H, et al. Degradation and diversity changes of meadow grassland in Keerqin Sandy Land[J]. Acta Prataculturae Sinica, 2006, 15(3): 18−26. doi: 10.3321/j.issn:1004-5759.2006.03.003
[19] 王健铭, 王文娟, 李景文, 等. 中国西北荒漠区植物物种丰富度分布格局及其环境解释[J]. 生物多样性, 2017, 25(11):1192−1201. doi: 10.17520/biods.2017149 Wang J M, Wang W J, Li J W, et al. Biogeographic patterns and environmental interpretation of plant species richness in desert regions of Northwest China[J]. Biodiversity Science, 2017, 25(11): 1192−1201. doi: 10.17520/biods.2017149
[20] 龙婷, 王健铭, 李景文, 等. 青藏高原北部戈壁区植物多样性及其环境解释[J]. 北京林业大学学报, 2017, 39(12):17−24. Long T, Wang J M, Li J W, et al. Plant diversity and its environmental explaination in gobi district of northern Qinghai-Tibet Plateau, northwestern China[J]. Journal of Beijing Forestry University, 2017, 39(12): 17−24.
[21] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6):533−548. doi: 10.3724/SP.J.1003.2009.09253 Fang J Y, Wang X P, Shen Z H, et al. Methods and protocols for plant community inventory[J]. Biodiversity Science, 2009, 17(6): 533−548. doi: 10.3724/SP.J.1003.2009.09253
[22] Wang Z H, Fang J Y, Tang Z Y, et al. Relative role of contemporary environment versus history in shaping diversity patterns of China’s woody plants[J]. Ecography, 2011, 34: 1−10. doi: 10.1111/j.1600-0587.2010.06453.x
[23] Hawkins B A, Porter E E. Water-energy balance and the geographic pattern of species richness of western Palearctic butterflies[J]. Ecological Entomology, 2003, 28: 678−686. doi: 10.1111/j.1365-2311.2003.00551.x
[24] Burnham K P, Anderson D R. Model selection and multimodel inference[M]. New York: Springer, 2002.
[25] Borcard D, Gillet F, Legendre P. Numerical ecology with R[M]. New York: Springer Science & Business Media, 2011.
[26] Zhang Q, Hou X Y, Li F Y H, et al. Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia Grassland[J/OL]. PLoS One, 2014, 9(3): e93518 (2014−03−27) [2018−05−10]. https://doi.org/10.1371/journal.pone.0093518.
[27] 姚鸿云, 李小雁, 郭娜, 等. 多年放牧对不同类型草原植被及土壤碳同位素的影响[J]. 应用生态学报, 2019, 30(2):553−562. Yao H Y, Li X Y, Guo N, et al. Effects of long-term grazing on carbon isotope composition in plants and soils of different grasslands[J]. Chinese Journal of Applied Ecology, 2019, 30(2): 553−562.
[28] 马少薇, 郭建英, 李锦荣, 等. 放牧强度对短花针茅群落特征及冠层截留的影响[J]. 中国草地学报, 2016, 38(5):66−70. Ma S W, Guo J Y, Li J R, et al. Effect of grazing intensity on Stipa breviflora communities and canopy interception[J]. Chinese Journal of Grassland, 2016, 38(5): 66−70.
[29] 牛钰杰, 杨思维, 王贵珍, 等. 放牧强度对高寒草甸土壤理化性状和植物功能群的影响[J]. 生态学报, 2018, 38(14):5006−5016. Niu Y J, Yang S W, Wang G Z, et al. Effects of grazing disturbance on soil properties and plant functional groups and their relationships in an alpine meadow on the Tibetan Plateau, China[J]. Acta Ecologica Sinica, 2018, 38(14): 5006−5016.
[30] Augustine D J, Derner J D, Milchunas D, et al. Grazing moderates increases in C3 grass abundance over seven decades across a soil texture gradient in shortgrass steppe[J]. Journal of Vegetation Science, 2017, 28: 562−572.
[31] 杨婧, 褚鹏飞, 陈迪马, 等. 放牧对内蒙古典型草原α、β和γ多样性的影响机制[J]. 植物生态学报, 2014, 38(2):188−200. doi: 10.3724/SP.J.1258.2014.00017 Yang J, Chu P F, Chen D M, et al. Mechanisms underlying the impacts of grazing on plant α, β and γ diversity in a typical steppe of the Inner Mongolia Gassland[J]. Chinese Journal of Plant Ecology, 2014, 38(2): 188−200. doi: 10.3724/SP.J.1258.2014.00017
[32] Zhang R Y, Wang Z W, Han G D, et al. Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the desert steppe, northern China[J]. Agriculture, Ecosystems and Environment, 2018, 265: 73−83. doi: 10.1016/j.agee.2018.05.014
[33] 王寅, 王键铭, 崔盼杰, 等. 科尔沁沙地植物物种丰富度格局及其与环境的关系[J]. 植物科学学报, 2018, 36(6):794−803. doi: 10.11913/PSJ.2095-0837.2018.60794 Wang Y, Wang J M, Cui P J, et al. Biogeographical patterns and environmental interpretation of plant species richness in the Horqin Sandy Lands[J]. Plant Science Journal, 2018, 36(6): 794−803. doi: 10.11913/PSJ.2095-0837.2018.60794
[34] 韩梦琪, 王忠武, 靳宇曦, 等. 短花针茅荒漠草原物种多样性及生产力对长期不同放牧强度的响应[J]. 西北植物学报, 2017, 37(11):2273−2281. doi: 10.7606/j.issn.1000-4025.2017.11.2273 Han M Q, Wang Z W, Jin Y X, et al. Response of species diversity and productivity to long-term grazing in the Stipa braviflora desert steppe[J]. Acta Bot Boreal Occident Sin, 2017, 37(11): 2273−2281. doi: 10.7606/j.issn.1000-4025.2017.11.2273
[35] Deng L, Sweeney S, Shangguan Z P. Grassland responses to grazing disturbance: plant diversity changes with grazing intensity in a desert steppe[J]. Grass and Forage Science, 2013, 69: 524−533.
[36] 杨婧. 放牧对典型草原生态系统服务功能影响的研究[D]. 呼和浩特: 内蒙古农业大学, 2013. Yang J. Study on ecosystem service in typical steppe under different grazing intensities[D]. Huhhot: Inner Mongolia Agricultural University, 2013.
[37] 赵丽娅, 张晓雨, 熊炳桥, 等. 围封和放牧对科尔沁沙质草地植被和土壤的影响[J]. 生态环境学报, 2017, 26(6):971−977. Zhao L Y, Zhang X Y, Xiong B Q, et al. Influence of fencing and grazing on the soil and standing vegetation changes in Horqin Sandy Grassland[J]. Ecology and Environmental Sciences, 2017, 26(6): 971−977.
[38] 赵哈林, 大黑俊哉, 李玉霖, 等. 人类放牧活动与气候变化对科尔沁沙质草地植物多样性的影响[J]. 草业学报, 2008, 17(5):1−8. doi: 10.3321/j.issn:1004-5759.2008.05.001 Zhao H L, Okuro T, Li Y L, et al. Effects of human activities and climate changes on plant diversity in Horqin Sandy Grassland, Inner Mongolia[J]. Acta Prataculturae Sinica, 2008, 17(5): 1−8. doi: 10.3321/j.issn:1004-5759.2008.05.001
[39] 左小安, 赵学勇, 赵哈林, 等. 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系[J]. 环境科学, 2007, 28(5):945−951. doi: 10.3321/j.issn:0250-3301.2007.05.003 Zuo X A, Zhao X Y, Zhao H L, et al. Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin Sand Land[J]. Environmental Science, 2007, 28(5): 945−951. doi: 10.3321/j.issn:0250-3301.2007.05.003




计量
- 文章访问数: 3227
- HTML全文浏览量: 1014
- PDF下载量: 163
