
Dynamics of chlorophyll fluorescence parameters under drought condition for three desert shrub species
-
摘要:目的 研究荒漠植物生理特征对自然环境波动下的响应过程可以更好地认识和预测气候变化对荒漠生态系统的影响。方法 本文研究了宁夏盐池油蒿、沙柳和杨柴3种荒漠灌木优势物种在春季干旱环境下光系统II(PSII)最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSII)和非光化学淬灭(NPQ)对环境因子的响应,旨在探讨荒漠灌木在干旱环境下PSII的光合生理状态以及其保护调节机制。结果 结果表明:在水分胁迫下(土壤含水量 < 0.08 m3/m3),3种荒漠灌木Fv/Fm、ΦPSII显著降低,NPQ显著增加(P < 0.01),沙柳Fv/Fm显著低于油蒿并且显著高于杨柴(P < 0.01),杨柴ΦPSII显著低于油蒿和沙柳(P < 0.01)。3种荒漠灌木ΦPSII和NPQ受光合有效辐射(PAR)的影响呈现出明显的日变化,且PAR、空气温度(Ta)、相对湿度 (RH)和饱和水汽压差(VPD)对ΦPSII和NPQ均有影响。研究发现3种荒漠灌木PSII表现出不同受胁程度,其通过自身调节机制增加热耗散比例来抵御干旱下的环境胁迫。油蒿Fv/Fm能通过自身调节恢复到适宜状态值(约0.75),认为其反应中心损伤具有可恢复性。杨柴热耗散的光保护响应机制的敏感性较高,认为其受环境波动的影响较大。结论 综上得出油蒿可以作为干旱区植被恢复的首选物种,沙柳其次,杨柴不建议考虑。可见由叶绿素荧光参数变化表征PSII反应中心的活性,可以充分体现荒漠灌木对环境波动的响应策略,可以作为探究干旱半干旱区荒漠植物对环境变化响应的重要手段。Abstract:Objective Research on the response process of physiological parameters of desert plants to natural environment fluctuations can better understand and predict the impact of climate change on desert ecosystems.Method We conducted in situ field measurements of chlorophyll fluorescence for three desert shrub species (Artemisia ordosica, Salix psammophila and Hedysarum mongolicum) to explore the responses of photosystem II (PSII) maximum quantum yield (Fv / Fm), photochemical efficiency (ΦPSII) and non-photochemical quenching (NPQ) to environmental factors, in order to understand the PSII photosynthetic physiology and its photoprotection mechanism of desert shrubs in arid environment.Result Under water stress (soil water content < 0.08 m3/m3), Fv / Fm and Φ PSII of three kinds of desert shrubs were reduced and NPQ increased (P < 0.01). The Fv / Fm of S. psammophila was lower than that of A. ordosica and higher than H. mongolicum (P < 0.01). The ΦPSII of H. mongolicum was lower than both A. ordosica and S. psammophila (P < 0.01). The ΦPSII and NPQ for three typical desert shrubs presented a clear daily variation with photosynthetic active radiation (PAR), and also were affected by air temperature (Ta), relative humidity (RH) and vapor pressure difference (VPD). Three shrub species resisted drought stress through heat dissipation mechanism, but had different degrees. The Fv / Fm of A. ordosica can be restored to an appropriate value of about 0.75 through self-regulation of heat dissipation, and its reaction center damage was recoverable. H. mongolicum was greatly affected by drought stress, and its photoprotective response of heat dissipation was sensitive.Conclusion A. ordosica can be considered as the first choice for vegetation restoration in arid areas, S. psammophila is the second and H. mongolicum is not recommended. The activity of PSII reaction center is characterized by changes in chlorophyll fluorescence parameters, which can fully reflect the response strategy of desert shrubs to environmental fluctuations, and can be used as an important means to explore the response of desert plants in arid and semi-arid areas to environmental changes.
-
Keywords:
- arid and semiarid ecosystem /
- chlorophyll fluorescence /
- drought /
- desert shrub
-
-
![]()
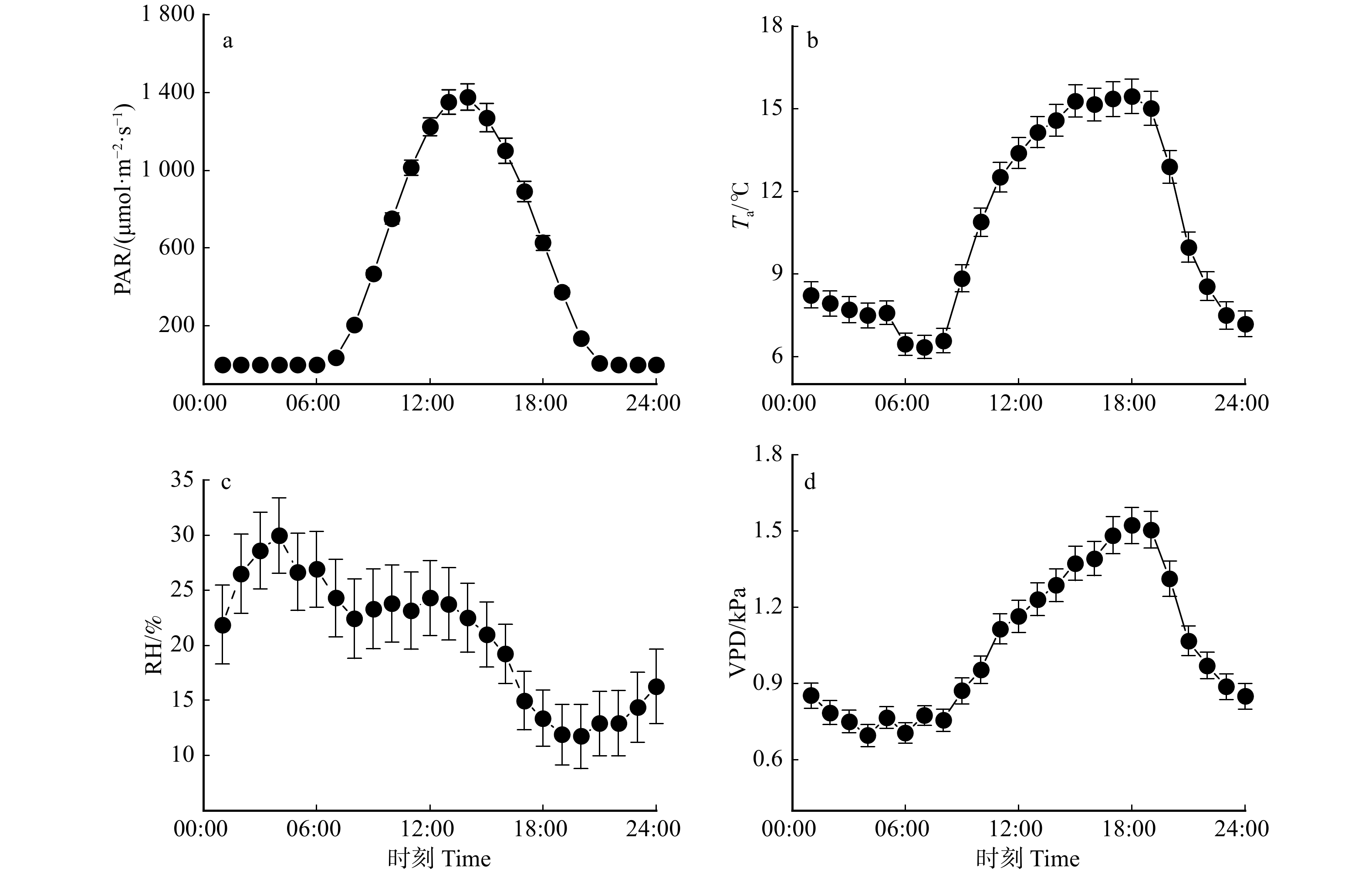
图 1 环境因子的动态变化
SWC10 cm. 10 cm 土壤含水量; SWC30 cm. 30 cm 土壤含水量; SWC70 cm. 70 cm 土壤含水量。SWC10 cm, soil water content at 10 cm depth;SWC30 cm, soil water content at 30 cm depth;SWC70 cm, soil water content at 70 cm depth.
Figure 1. Dynamic variations in environmental factors
![]()
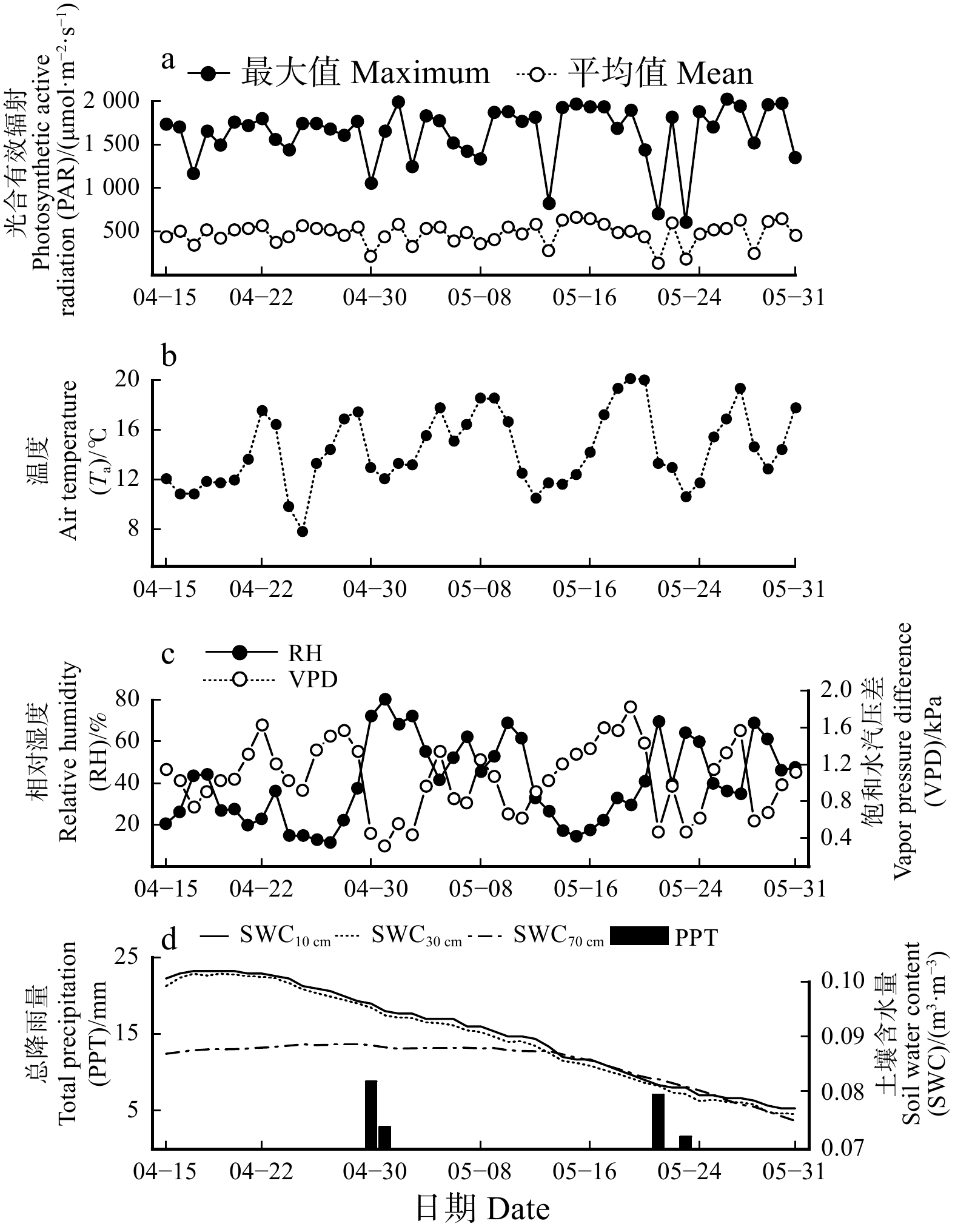
图 2 环境因子的日动态
数据为按小时整理后求得的小时平均值(± 标准误)。下同。All data are averaged into hourly mean values (± standard error) . The same below.
Figure 2. Diurnal variations in environmental factors
![]()
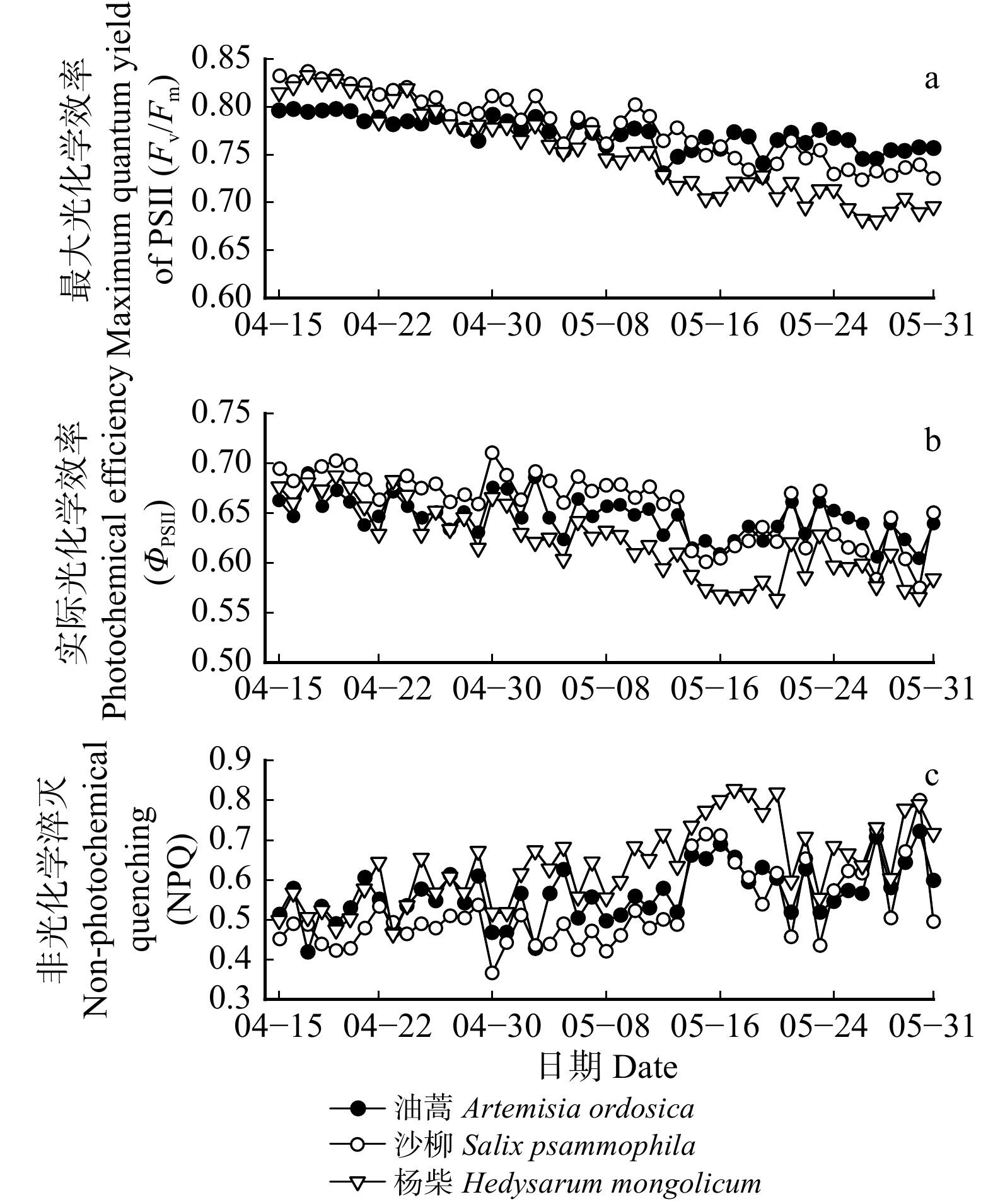
图 3 叶绿素荧光参数的动态变化
Fv/Fm为夜晚数据的平均值、ΦPSII和NPQ为昼夜平均值。Night means are shown for Fv/Fm , daily means are shown for ΦPSII and NPQ.
Figure 3. Dynamic variations in chlorophyll fluorescence parameters
![]()
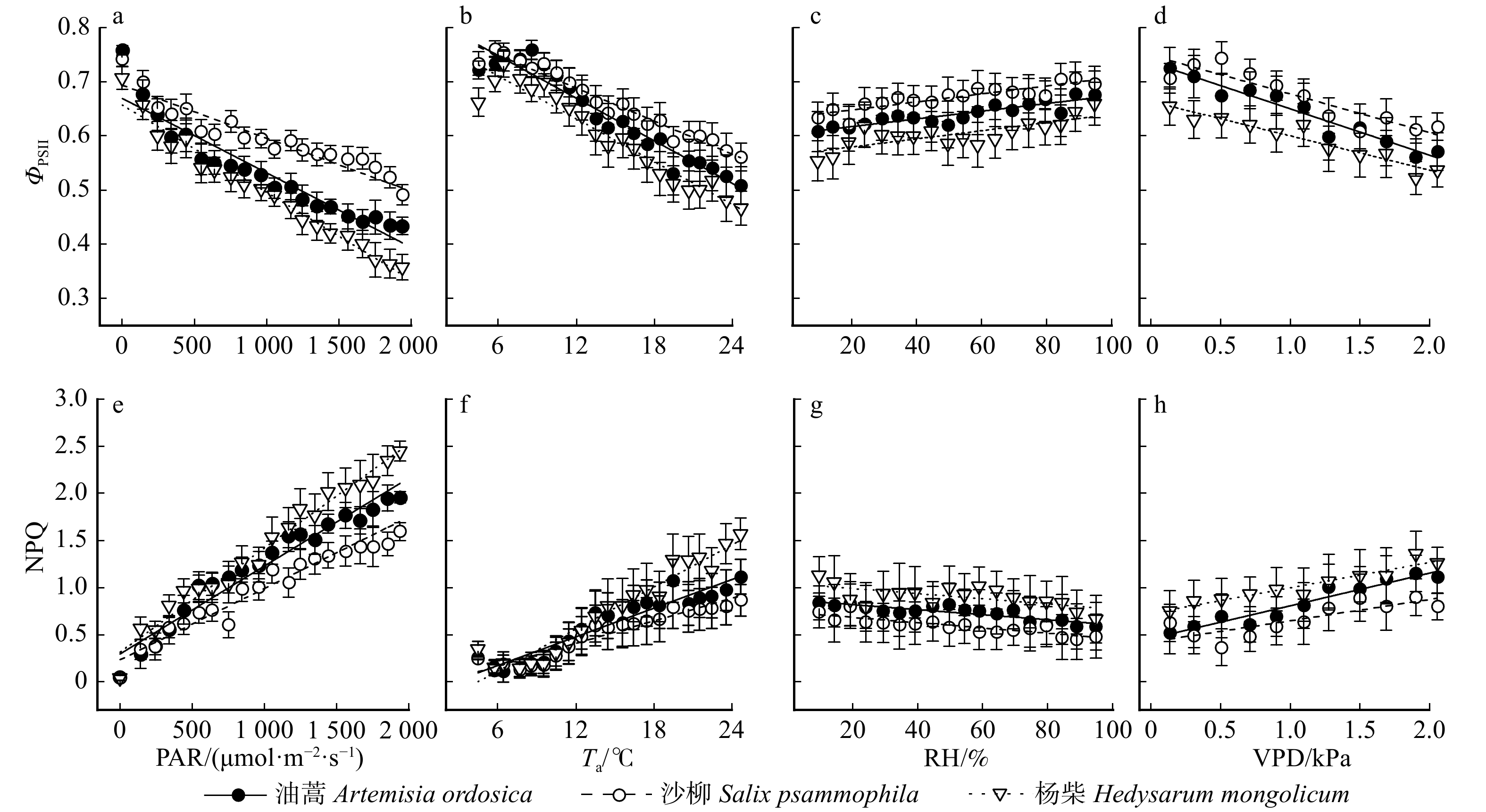
图 5 3种荒漠灌木叶绿素荧光参数对环境因子的敏感性
去除夜晚数据,将每小时的ΦPSII和NPQ按PAR间隔100 μmol/(m2·s)、Ta间隔1°C、RH间隔5%、VPD 间隔0.2 kPa 合并,数据为平均值 ± 标准误。Removing the night data, hourly ΦPSII and NPQ are bin-averaged into 100 μmol/(m2·s) PAR(a, e), 1°C Ta(b, f), 5% RH(c, g) and 0.2 kPa VPD(d, h) intervals, respectively. Data are mean ± standard error.
Figure 5. Responses of chlorophyll fluorescence parameters to environmental factors for three desert shrub species
表 1 3种荒漠灌木叶绿素荧光参数随环境因子的变化幅度
Table 1 Changing range of chlorophyll fluorescence parameters with environmental factors for three desert shrub species
叶绿素荧光参数
Chlorophyll fluorescence parameter物种 Species PAR Ta RH VPD ΦPSII 油蒿 Artemisia ordosica 0.61 0.36 0.11 0.26 沙柳 Salix psammophila 0.48 0.28 0.13 0.20 杨柴 Hedysarum mongolicum 0.70 0.41 0.18 0.22 NPQ 油蒿 Artemisia ordosica 1.58 1.66 0.35 0.75 沙柳 Salix psammophila 1.84 1.39 0.58 0.79 杨柴 Hedysarum mongolicum 1.70 1.88 0.52 0.60  下载: 导出CSV
下载: 导出CSV
-
[1] 张煜星. 中国荒漠化气候类型的分布[J]. 干旱区研究, 1998, 15(2):46−50. Zhang Y X. Distribution of climate type of Chinese desertification[J]. Arid Zone Research, 1998, 15(2): 46−50.
[2] 林年丰, 汤洁. 中国干旱半干旱区的环境演变与荒漠化的成因[J]. 地理科学, 2001, 21(1):24−29. doi: 10.3969/j.issn.1000-0690.2001.01.005 Lin N F, Tang J. Study on the environmental evolution and the causes of desertification in arid and semiarid regions in China[J]. Scientia Geographica Sinica, 2001, 21(1): 24−29. doi: 10.3969/j.issn.1000-0690.2001.01.005
[3] Barros V, Stocker T F. Managing the risks of extreme events and disasters to advance climate change adaptation: special report of the Intergovernmental Panel on Climate Change[J]. Journal of Clinical Endocrinology & Metabolism, 2012, 18(6): 586−599.
[4] Huang J, Yu H, Dai A, et al. Drylands face potential threat under 2 degrees C global warming target[J]. Nature Climate Change, 2017, 7(6): 417−422. doi: 10.1038/nclimate3275
[5] Li B, Chen Y, Shi X, et al. Temperature and precipitation changes in different environments in the arid region of northwest China[J]. Theoretical and Applied Climatology, 2013, 112(3−4): 589−596. doi: 10.1007/s00704-012-0753-4
[6] Huang J, Yu H, Guan X, et al. Accelerated dryland expansion under climate change[J]. Nature Climate Change, 2016, 6(2): 166−171. doi: 10.1038/nclimate2837
[7] Juan W Y, Cai R, Yun T, et al. Photosynthetic gas-exchange and PSⅡ photochemical acclimation to drought in a native and non-native xerophytic species (<italic>Artemisia ordosica</italic> and <italic>Salix psammophila</italic>)[J]. Ecological Indicators, 2018, 94: 130−138. doi: 10.1016/j.ecolind.2018.06.040
[8] 种培芳, 李毅, 苏世平. 荒漠植物红砂叶绿素荧光参数日变化及其与环境因子的关系[J]. 中国沙漠, 2010, 30(3):539−545. Chong P F, Li Y, Su S P. Diurnal change in chlorophyllⅡfluorescence parameters of desert plant <italic>Reaumuria soongorica</italic> and its relationship with environmental factors[J]. Journal of Desert Research, 2010, 30(3): 539−545.
[9] 陈凤丽, 靳正忠, 李生宇, 等. 高温对花花柴(<italic>Karelinia caspica</italic>)光系统Ⅱ的影响[J]. 中国沙漠, 2013, 33(5):1371−1376. doi: 10.7522/j.issn.1000-694X.2013.00201 Chen F L, Jin Z Z, Li S Y, et al. Effects of heat stress on photosystem Ⅱ in <italic>Karelinia caspica</italic>[J]. Journal of Desert Research, 2013, 33(5): 1371−1376. doi: 10.7522/j.issn.1000-694X.2013.00201
[10] Chambers J C, Bradley B A, Brown C S, et al. Resilience to stress and disturbance, and resistance to <italic>Bromus tectorum</italic> L. invasion in cold desert shrublands of western North America[J]. Ecosystems, 2014, 17: 360−375. doi: 10.1007/s10021-013-9725-5
[11] Grant O M, Tronina L, García-Plazaola J L, et al. Resilience of a semi-deciduous shrub, <italic>Cistus salvifolius</italic>, to severe summer drought and heat stress[J]. Functional Plant Biology, 2014, 42(2): 219−228.
[12] 吴雅娟, 查天山, 贾昕, 等. 油蒿 (<italic>Artemisia ordosica</italic>) 光化学量子效率和非光化学淬灭的动态及其影响因子[J]. 生态学杂志, 2015, 34(2):319−325. Wu Y J, Zha T S, Jia X, et al. Temporal variation and controlling factors of photochemical efficiency and non-photochemical quenching in <italic>Artemisia ordosica</italic>[J]. Chinese Journal of Ecology, 2015, 34(2): 319−325.
[13] 张继伟, 赵昕, 陈国雄, 等. 盐胁迫下荒漠植物柠条和油蒿的离子吸收及分配特征[J]. 干旱区资源与环境, 2016, 30(3):68−73. Zhang J W, Zhao X, Chen G X, et al. Ion absorption and distribution in <italic>Caragana korshinskii</italic> and <italic>Artemisia ordosica</italic> seedlings under different NaCl stress[J]. Journal of Arid Land Resources and Environment, 2016, 30(3): 68−73.
[14] 王飞, 刘世增, 康才周, 等. 干旱胁迫对沙地云杉光合、叶绿素荧光特性的影响[J]. 干旱区资源与环境, 2017, 31(1):142−147. Wang F, Liu S Z, Kang C Z, et al. Effects of drought stress on photosynthesis and chlorophyll fluorescence characteristics of <italic>Picea mongolica</italic>[J]. Journal of Arid Land Resources and Environment, 2017, 31(1): 142−147.
[15] 张明艳, 贾昕, 查天山, 等. 油蒿 (<italic>Artemisia ordosica</italic>) 光系统Ⅱ光化学效率对去除降雨的响应[J]. 中国沙漠, 2017, 37(3):475−482. doi: 10.7522/j.issn.1000-694X.2016.00021 Zhang M Y, Jia X, Zha T S, et al. PSⅡ photochemical efficiency of <italic>Artemisia ordosica</italic> in response to rainfall exclusion[J]. Journal of Desert Research, 2017, 37(3): 475−482. doi: 10.7522/j.issn.1000-694X.2016.00021
[16] 张景波, 张金鑫, 卢琦, 等. 乌兰布和沙漠油蒿叶片PSⅡ叶绿素荧光动力学参数及其光响应曲线动态[J]. 草业科学, 2019, 36(3):713−719. Zhang J B, Zhang J X, Lu Q, et al. Dynamic changes of leaf parameters of PSⅡ fluorescence kinetics and fast photosynthetic response curves in <italic>Artemisia ordosica</italic>[J]. Pratacultural Science, 2019, 36(3): 713−719.
[17] 朴世龙, 张新平, 陈安平, 等. 极端气候事件对陆地生态系统碳循环的影响[J]. 中国科学:地球科学, 2019, 49(9):1321−1334. doi: 10.1360/N072018-00316 Piao S L, Zhang X P, Chen A P, et al. The impacts of climate extremes on the terrestrial carbon cycle: a review[J]. Scientia Sinica Terrae, 2019, 49(9): 1321−1334. doi: 10.1360/N072018-00316
[18] 任才, 贾昕, 吴雅娟, 等. 油蒿光系统Ⅱ光化学参数在生长季早期对降雪事件的响应[J]. 北京林业大学学报, 2019, 41(12):119−127. doi: 10.12171/j.1000-1522.20190058 Ren C, Jia X, Wu Y J, et al. Responses of PSⅡ photochemical parameter to a snowfall event in early growing season in <italic>Artemisia ordosica</italic>[J]. Journal of Beijing Forestry University, 2019, 41(12): 119−127. doi: 10.12171/j.1000-1522.20190058
[19] 杨洪晓, 张金屯, 吴波, 等. 油蒿对半干旱区沙地生境的适应及其生态作用[J]. 北京师范大学学报(自然科学版), 2004, 40(5):681−690. Yang H X, Zhang J T, Wu B, et al. Adaptation of <italic>Artemisia ordosica</italic> to temperature arid sandy land and its roles in habitat shift[J]. Journal of Beijing Normal University (Natural Science), 2004, 40(5): 681−690.
[20] 曹生奎, 冯起, 司建华, 等. 植物水分利用效率研究方法综述[J]. 中国沙漠, 2009, 29(5):853−858. Cao S K, Feng Q, Si J H, et al. Summary on research methods of water use efficiency in plant[J]. Journal of Desert Research, 2009, 29(5): 853−858.
[21] 王燕凌, 刘君, 李文兵, 等. 塔里木河下游刚毛柽柳光合作用、蒸腾作用及水分利用效率特性研究[J]. 新疆农业科学, 2015, 52(2):292−299. Wang Y L, Liu J, Li W B, et al. Study on characteristics in photosynthesis, transpiration and water use efficiency of <italic>Tamarix hispida</italic> Willd. in the lower reaches of the Tarim River[J]. Xinjiang Agricultural Sciences, 2015, 52(2): 292−299.
[22] Massacci A, Nabiev S M, Pietrosanti L, et al. Response of the photosynthetic apparatus of cotton (<italic>Gossypium hirsutum</italic>) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging[J]. Plant Physiology and Biochemistry, 2008, 46(2): 189−195. doi: 10.1016/j.plaphy.2007.10.006
[23] 张亚娟, 谢忠奎, 赵学勇, 等. 水分胁迫对东方百合光合特性、叶绿素荧光参数及干物质积累的影响[J]. 中国沙漠, 2011, 31(4):884−888. Zhang Y J, Xie Z K, Zhao X Y, et al. Effects of water stress on photosynthetic characteristics, chlorophyll fluorescence, and dry matter of <italic>Oriental lilies</italic>[J]. Journal of Desert Research, 2011, 31(4): 884−888.
[24] Xu W Z, Deng X P, Xu B C. Photosynthetic activity and efficiency of <italic>Bothriochloa ischaemum</italic> and <italic>Lespedeza davurica</italic> in mixtures across growth periods under water stress[J]. Acta Physiologiae Plantarum, 2014, 36(4): 1033−1044. doi: 10.1007/s11738-013-1481-9
[25] 贾虎森, 韩亚琴. 高等植物光合作用的光抑制研究进展[J]. 植物学通报, 2000, 17(3):218−224. Jia H S, Han Y Q. Advances in studies on photoinhibition in photosynthesis of higher plants[J]. Chinese Bulletin of Botany, 2000, 17(3): 218−224.
[26] 石莎, 冯金朝, 邹学勇. 腾格里沙漠南缘2种沙地灌木植物的光合特征[J]. 云南大学学报(自然科学版), 2007, 29(5):519−524. doi: 10.3321/j.issn:0258-7971.2007.05.019 Shi S, Feng J Z, Zou X Y. Photosynthetic characteristics of two desert shrubs in south of Tengger Desert[J]. Journal of Yunnan University (Natural Sciences Edition), 2007, 29(5): 519−524. doi: 10.3321/j.issn:0258-7971.2007.05.019
[27] 韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性[J]. 生态学报, 2010, 30(15):4019−4026. Han G, Zhao Z. Light response characteristics of photosynthesis of four xerophilous shrubs under different soil moistures[J]. Acta Ecologica Sinica, 2010, 30(15): 4019−4026.
[28] Doupis G, Bertaki M, Psarras G, et al. Water relations, physiological behavior and antioxidant defence mechanism of olive plants subjected to different irrigation regimes[J]. Scientia Horticulturae, 2013, 153(3): 150−156.
[29] 何炎红, 白玉娥, 王海燕, 等. 光胁迫对沙冬青叶绿素荧光特征和光呼吸的影响[J]. 西北农业学报, 2015, 24(10):124−130. doi: 10.7606/j.issn.1004-1389.2015.10.018 He Y H, Bai Y E, Wang H Y, et al. Effect of light stress on chlorophyll fluorescence and photorespiration of <italic>Ammopiptanthus mongolicus</italic>[J]. Acta Agriculturae Boreali-Occidentalis Sinica, 2015, 24(10): 124−130. doi: 10.7606/j.issn.1004-1389.2015.10.018
[30] 姜闯道, 高辉远, 邹琦, 等. 叶角、光呼吸和热耗散协同作用减轻大豆幼叶光抑制[J]. 生态学报, 2005, 25(2):319−325. doi: 10.3321/j.issn:1000-0933.2005.02.021 Jiang C D, Gao H Y, Zou Q, et al. The co-operation of leaf orientation, photorespiration and thermal dissipation alleviate photoinhibition in young leaves of soybean plants[J]. Acta Ecologica Sinica, 2005, 25(2): 319−325. doi: 10.3321/j.issn:1000-0933.2005.02.021
[31] Öquist G, Chow W S, Anderson J M. Photoinhibition of photosynthesis represents a mechanism for the long-term regulation of photosystem Ⅱ[J]. Planta, 1992, 186(3): 450−460.
[32] Ruban A V, Murchie E H. Assessing the photoprotective effectiveness of non-photochemical chlorophyll fluorescence quenching: a new approach[J]. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 2012, 1817(7): 977−982. doi: 10.1016/j.bbabio.2012.03.026
[33] Ruban A V. Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage[J]. Plant Physiology, 2016, 170(4): 1903−1916. doi: 10.1104/pp.15.01935
[34] 阮成江, 李代琼. 黄土丘陵区沙棘气孔导度及其影响因子[J]. 西北植物学报, 2001, 21(6):30−36. Ruan C J, Li D Q. Stomatal conductance and influence factors of seabuckthorn in loess hilly region[J]. Acta Botanica Boreali-Occidentalia Sinica, 2001, 21(6): 30−36.
[35] Campos H, Trejo C, Peña-Valdivia C B, et al. Stomatal and non-stomatal limitations of bell pepper (<italic>Capsicum annuum</italic> L <italic></italic>.) plants under water stress and re-watering: delayed restoration of photosynthesis during recovery[J]. Environmental & Experimental Botany, 2014, 98(1): 56−64.
[36] 许大全. 光系统Ⅱ反应中心的可逆失活及其生理意义[J]. 植物生理学通讯, 1999, 35(4):273−276. Xu D Q. Reversible inactivation of photosystem Ⅱ reaction centers and its physiological ignificance[J]. Plant Physiology Communications, 1999, 35(4): 273−276.
[37] Porcar-Castell A, Pfündel E, Korhonen J F J, et al. A new monitoring PAM fluorometer (MONI-PAM) to study the short-and long-term acclimation of photosystem Ⅱ in field conditions[J]. Photosynthesis Research, 2008, 96(2): 173−179. doi: 10.1007/s11120-008-9292-3
[38] Kalaji H M, Jajoo A, Oukarroum A, et al. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions[J]. Acta Physiologiae Plantarum, 2016, 38: 102−112. doi: 10.1007/s11738-016-2113-y
[39] 靳虎甲, 王继和, 李毅, 等. 油蒿生态学研究综述[J]. 西北林学院学报, 2009, 24(4):62−66. Jin H J, Wang J H, Li Y, et al. Summary of <italic>Artemisia ordosica</italic> ecology studies[J]. Journal of Northwest Forestry University, 2009, 24(4): 62−66.
[40] 马全林, 郑庆中, 贾举杰, 等. 乌兰布和沙漠沙蒿与油蒿群落的物种组成与数量特征[J]. 生态学报, 2012, 3(11):3423−3431. Ma Q L, Zheng Q Z, Jia J J, et al. Quantitative characteristics and species composition of <italic>Artemisia sphaerocephala</italic> and <italic>A. ordosica</italic> communities in the Ulanbuh Desert[J]. Acta Ecologica Sinica, 2012, 3(11): 3423−3431.
[41] Jia X, Zha T S, Wu B, et al. Biophysical controls on net ecosystem CO<sub>2</sub> exchange over a semiarid shrubland in northwest China[J]. Biogeosciences Discussions, 2014, 11(3): 4679−4693.
[42] Schansker G, Van Rensen J J S. Performance of active photosystem Ⅱ centers in photoinhibited pea leaves[J]. Photosynthesis Research, 1999, 62(2): 175−184.
[43] 刘晓晴, 常宗强, 马亚丽, 等. 胡杨(<italic>Populus euphratica</italic>)异形叶叶绿素荧光动力学[J]. 中国沙漠, 2014, 34(3):704−711. doi: 10.7522/j.issn.1000-694X.2014.00007 Liu X Q, Chang Z Q, Ma Y L, et al. Characteristics of the fast chlorophyll fluorescence induction kinetics of heteromorphic leaves in <italic>Populus euphratica</italic>[J]. Journal of Desert Research, 2014, 34(3): 704−711. doi: 10.7522/j.issn.1000-694X.2014.00007
[44] 师生波, 张怀刚, 师瑞, 等. 青藏高原春小麦叶片光合作用的光抑制及PSⅡ反应中心光化学效率的恢复分析[J]. 植物生态学报, 2014, 38(4):375−386. doi: 10.3724/SP.J.1258.2014.00034 Shi S B, Zhang H G, Shi R, et al. Assessment of photosynthetic photo-inhibition and recovery of PSⅡ photochemical efficiency in leaves of wheat varieties in Qinghai-Xizang Plateau[J]. Chinese Journal of Plant Ecology, 2014, 38(4): 375−386. doi: 10.3724/SP.J.1258.2014.00034
[45] 郝建卿, 吕娜, 杨扬, 等. 内蒙乌海胡杨异形叶水分及叶绿素荧光参数的比较[J]. 北京林业大学学报, 2010, 32(5):41−44. Hao J Q, Lü N, Yang Y, et al. Comparative study of chlorophyll fluorescence parameters and water physiological characters of heteromorphic leaves for <italic>Populus euphratica</italic>[J]. Journal of Beijing Forestry University, 2010, 32(5): 41−44.
[46] 韩婧, 吴益, 赵琳,等. 光周期对促成栽培芍药生长开花和叶绿素荧光动力学影响[J]. 北京林业大学学报, 2015, 37(9):62−69. Han J, Wu Y, Zhao L, et al. Effects of photoperiod on the growth, flowering and chlorophyll fluorescence kinetics of forced <italic>Paeonia lactiflora</italic>[J]. Journal of Beijing Forestry University, 2015, 37(9): 62−69.
-
期刊类型引用(1)
1. 徐媛,陈锦玲,陈玉梅,李璐璐,李惠敏,秦新民. 干旱胁迫下花生转录组与miRNA测序及相关基因的表达. 贵州农业科学. 2021(01): 1-9 .  百度学术
百度学术
其他类型引用(3)

计量
- 文章访问数: 2164
- HTML全文浏览量: 628
- PDF下载量: 82
- 被引次数: 4
