
Construction of the height to crown base mixed model for Korean pine
-
摘要:目的 基于帽儿山红松人工林63块样地2 972株红松数据,利用非线性混合模型构建红松枝下高模型,为进一步研究生长与收获模型提供理论依据。方法 本文首先使用8个常用的枝下高模型,选出最优基础模型;其次,研究林分变量或单木变量对枝下高的影响,建立含林分变量的枝下高模型;最终在基础模型和含林分变量模型的基础上,考虑样地效应对红松枝下高的影响,构建红松枝下高基础混合效应模型和广义混合效应模型。模型用4种抽样方式(随机抽取、抽取最大树、抽取最小树、抽取平均树)和8种样本大小(1 ~ 8株树)对基础混合效应模型和广义混合效应模型进行抽样检验。结果 Logistic模型拟合精度好,符合生物学意义,且模型形式简单,选为最优基础模型。除树高、胸径以外,大于对象木断面积之和、优势木高和冠幅与枝下高有显著相关性,加入后明显提升模型的拟合精度。枝下高广义混合效应模型的拟合效果要优于其他模型。模型检验结果表明:当应用基础混合效应模型预测时,建议抽取胸径最小的4个样本;当应用广义混合效应模型预测时,建议随机抽取4个样本。结论 枝下高广义混合效应模型在拟合效果和预测精度方面优于其他3种模型,建议将此模型作为人工红松枝下高模型。当应用广义混合效应模型预测时,建议随机抽取4个样本。Abstract:Objective Based on the data of 2 972 Korean pine trees in 63 sample plots of Korean pine plantation in Maor Mountain of northeastern China, a nonlinear mixed model was used to construct the height to crown base model of Korean pine, which provided a theoretical basis for further research on growth and yield model.Method Firstly, the optimal basic model was selected from eight commonly used models of height to crown base. Secondly, the influence of stand variables or individual tree variables on height to crown base was studied, and a generalized model was established. Finally, on the basis of the basic model and the generalized model, considering the effects of sample plot on the height to crown base of Korean pine, the basic mixed effect model and generalized mixed effect model of height to crown base for Korean pine were constructed. Four alternatives of height to crown base (HCB) sampling designs (the randomly selected trees and selecting the maximum, medium-size and minimum trees) and eight sample sizes (1−8 trees) were studied for sampling correction of basic mixed effect model and generalized mixed effect model respectively.Result Logistic model had good fitting accuracy and biological significance, and its form was simple, so it was chosen as the the optimal basic model. In addition to tree height (H) and DBH, there was a significant correlation between the basal area sum larger than subject tree (BAL), dominant height (HD), crown width (CW) and HCB, and the fitting accuracy of the model was improved obviously. The fitting effect of height to crown base generalized mixed effect model was better than that of other models. The model validation showed that when the basic mixed effect model was used to predict, it was recommended to select four samples with the smallest DBH, and when the generalized mixed effect model was used to predict, it was recommended to randomly select four samples.Conclusion The generalized mixed effect model is superior to the other three models in fitting effect and prediction accuracy. It is recommended that this model can be used as the height to crown base model for Korean pine. When applying generalized mixed effect model prediction, it is recommended that four samples can be randomly selected.
-
Keywords:
- Korean pine /
- nonlinear mixed model /
- height to crown base /
- plantation /
- mixed model calibration
-
多倍化是高等植物基因组进化的显著特征之一,大约有75%的被子植物和95%的蕨类植物在进化过程中发生过多倍化事件[1]。许多主要作物,包括小麦(Triticum aestivum,2n = 6x = 42)、土豆(Solanum tuberosum,2n = 4x = 48)、玉米(Zea mays,2n = 4x = 20)、咖啡豆(Coffea arabica,2n = 4x = 44)等,经过漫长的进化过程,最终生存下来的都是多倍体植株[2]。多倍体由于基因剂量效应,重复基因的表达情况也发生变化,导致植物在生长发育、形态生理、遗传适应性及对环境胁迫的耐性等方面与亲本二倍体有很大差异[3-5]。例如,相比二倍体,四倍体柑橘(Citrus limonia)叶片肥厚,茎干加粗,栅栏组织和海绵组织增长,表皮细胞体积增大,在高盐碱的逆境环境下,表现出更强的抗性[6-7];四倍体毛泡桐(Paulownia tomentosa)叶片面积、叶绿素含量、可溶性糖含量、可溶性蛋白质含量、净光合速率均大于二倍体,耐盐、耐寒、耐旱能力更强[8]。植物加倍后产生的新表型往往具有实际应用价值,如四倍体葡萄(Vitis vinifera)[9]、三倍体柑橘[10]、三倍体毛白杨(Populus tomentosa)[11]等许多品种都具有优于二倍体的性状,已经广泛应用于农林生产实践。
枣(Ziziphus jujuba)是鼠李科(Rhamnaceae)枣属(Ziziphus)植物,原产于中国,栽培历史悠久,枣果富含维生素和人体必需的氨基酸及多种微量元素[12],具有重要的营养和保健价值。酸枣(Ziziphus jujuba Mill. var. spinosa)作为枣的变种,果小、味酸,果仁可入药,其果实常用于制作果脯、果汁等,具有重要的经济价值[13]。此外,酸枣根系发达,耐寒、耐旱、耐贫瘠能力强[14],是重要的生态经济树种。近年来,枣树多倍体育种工作取得了显著的成果。Gu等[15]通过离体加倍获得了四倍体冬枣,相比二倍体,四倍体叶片变大变厚、颜色变深,保卫细胞内叶绿体增多,芽、茎变粗,花变大,花期延迟。刘孟军等[16]经秋水仙素诱变获得四倍体鲜食枣品种‘辰光’,具有果实大,果皮薄,果肉酥脆,早果速丰,枣头节间变短,叶缘锯齿增大,气孔密度减小,保卫细胞增大等特征。但目前尚未在分子水平上对这些变异特征进行深度探讨。
在前期工作中,本实验室以酸枣组培苗为试材进行秋水仙素离体诱导,成功获得了酸枣四倍体种质,并对二倍体和四倍体酸枣进行了表型特征的初步对比[17]。本研究以实验室原有的酸枣二倍体和四倍体组培苗为材料,对二者的生理指标进行测定,同时开展RNA-seq技术的转录组学研究,分析酸枣多倍体化导致的基因表达变化。通过该研究,有利于进一步了解基因组复制后分子水平的变异,深入理解酸枣四倍体性状变异,为今后关键调控基因的克隆及基因工程育种等工作提供参考。
1. 材料与方法
1.1 试验材料
以北京林业大学林木遗传育种国家工程实验室已有的酸枣二倍体及其四倍体组培苗为试验材料。组培室内温度维持在26 ℃,光周期为16 h光照/8 h黑暗。选取继代60 d的生长健壮的二倍体和四倍体酸枣组培苗,转接到生根培养基上,生根后30 d进行生理指标的测定及转录组测序。
分别随机选取二倍体和四倍体组培苗各5株,采集叶片测定叶绿素含量及叶片相对含水量,重复3次。同时,二倍体和四倍体组培苗分别随机选取5个植株,采集植物的叶片和茎,重复3次,取样后迅速放入液氮中磨成粉末,置于− 80 ℃冰箱保存,用于可溶性糖、可溶性蛋白含量测定以及转录组测序。
1.2 生理指标的测定
1.2.1 测定方法
用SPAD-502Plus仪测定叶绿素含量;用饱和称重法测定植物的叶片相对含水量[18];采用蒽酮比色法测定可溶性总糖含量[19];采用考马斯亮蓝染色法测定可溶性蛋白质含量[20]。
1.2.2 数据分析
使用Excel 2010进行数据录入及处理,用SPSS软件对数据进行独立样本t检验(P < 0.05)。
1.3 RNA提取
使用上海天根生化有限公司(Qiagen China, Shanghai, China)生产的RNeasy plant Mini kits试剂盒提取RNA,提取过程按说明书中的操作流程进行。用天根公司生产的RNase-Free DNaseSet对提取的RNA进行纯化。用琼脂糖电泳对提取出来的RNA完整性进行检测,紫外灯下观察是否具有清晰的条带。在进行转录组测序之前,对提取的RNA质量进一步评估,计算A260/A280是否介于1.8 ~ 2.0之间,28S/18S是否介于1.6 ~ 2.0之间,再分别用NanoDrop 2000分析仪(DE, USA)和Agilent 2100生物芯片分析系统检测RNA浓度和完整性,RNA检测合格之后用于后续分析。
1.4 转录组测序
委托北京百迈客生物科技有限公司(Biomarker Technology, Beijing, China)用Illunima HiSeq 2000测序系统进行cDNA文库构建及转录组测序(RNA-seq)。获得raw data后,去除测序接头及引物序列,并将低质量的序列过滤掉(Q30 ≥ 93.96%),得到clean data。利用HISAT2软件(http://ccb.jhu.edu/software/hisat2/index.shtml)将clean data比对到枣树参考基因组(https://www.ncbi.nlm.nih.gov/genome/15586?genome_assembly_id=219393),得到mapped data,进行插入片段长度检验、随机性检验等文库质量评估。
采用NCBI(https://www.ncbi.nlm.nih.gov/)、GO Ontology(http://geneontology.org/)、KEGG(https://www.kegg.jp/)、KOG(http://www.ncbi.nlm.nih.gov/KOG/)、Pfam(https://pfam.xfam.org/)、Swiss-Prot(https://www.uniprot.org/)数据库对基因组功能进行注释。基于所选参考基因组序列,使用StringTie(https://ccb.jhu.edu/software/stringtie/index.shtml)软件对mapped reads进行拼接,并与原有的基因组注释信息进行比较,寻找原来未被注释的转录区,过滤掉编码过短肽链(少于50个氨基酸残基)或只包含单个外显子的序列,发掘该物种的新基因,从而补充和完善原有的基因组注释信息。
1.5 差异基因的鉴定及生物信息学分析
应用DESeq2软件(http://www.bioconductor.org/packages/release/bioc/html/DESeq.htm)进行样品组间的差异表达分析,将差异倍数(FC) > 1.5且错误发现率(FDR) < 0.05作为筛选差异表达基因(DEG)的标准。对筛选出的DEG进行GO Ontology和KEGG Pathway分类,并应用超几何检验对注释到的不同的GO(P < 0.05)和KEGG条目(P < 0.05)进行富集分析。用FPKM值(Fragments per kilobase of transcript per million fragments mapped)作为衡量基因表达水平的指标,R语言包绘制表达量聚类热图。
2. 结果与分析
2.1 二倍体及其同源四倍体酸枣生理指标对比
植物染色体加倍后,细胞核内的遗传物质相应增加,一些营养物质或其他成分含量显著提高。由表1可知,四倍体酸枣可溶性糖、可溶性蛋白质、叶绿素含量显著高于二倍体,可溶性糖含量是二倍体的2倍;可溶性蛋白质比二倍体高50.1%;叶绿素含量比二倍体高27.2%。但是,四倍体植株叶片相对含水量与二倍体没有显著差异。
表 1 二倍体和四倍体酸枣不同生理指标的比较Table 1. Comparison of different physiological index between diploid and its autotetraploid sour jujube样品
Sample可溶性糖
Soluble sugar可溶性蛋白
Soluble protein叶绿素
Chlorophyll叶片相对含水量
Leaf relative water content二倍体酸枣
Diploid sour jujube25.03 ± 4.73a 4.13 ± 0.63a 29.17 ± 3.02a 0.52 ± 0.01a 四倍体酸枣
Autotetraploid sour jujube41.15 ± 4.45b 6.20 ± 0.62b 37.10 ± 2.82b 0.57 ± 0.01a 注:表内数值表示方式为“平均值 ± 标准差”,同列不同字母表示差异显著(P < 0.05)。Notes: values in table were “mean ± SD”, different letters in the same column represent significant differences (P < 0.05). 2.2 测序结果统计
为进一步理解二倍体酸枣加倍后变异的分子机制,对二倍体和四倍体酸枣共6个样本进行RNA-seq,获得平均7.92 Gb clean reads,Q30均在94%以上,将这些数据比对到枣树基因组,比对效率在82%以上,表明测序结果良好(表2)。
表 2 测序结果统计Table 2. Statistics of sequencing results样品 Sample Clean reads Clean bases Q30/% GC 含量 GC content/% 比对率 Mapped ratio/% D1 25 635 963 7 620 864 926 94.57 45.34 83.31 D2 27 771 165 8 268 560 788 94.43 45.1 83.67 D3 29 668 511 8 501 884 148 94.62 45.41 83.38 T1 27 572 679 7 659 547 606 94.21 45.19 82.92 T2 27 039 014 7 346 405 010 94.52 45.46 83.27 T3 29 816 038 8 096 301 012 94.53 45.32 82.52 注:D1、D2、D3分别为二倍体酸枣的3个重复;T1、T2、T3分别为四倍体酸枣的3个重复。Notes: D1, D2 and D3 are three biological duplicates of diploid sour jujube; T1, T2 and T3 are three biological duplicates of autotetraploid sour jujube. 通过对原有的枣树基因组进行补充和完善,共发掘2 536个新基因。将这些新基因与GO、KEGG、KOG、Pfam、Swiss-Prot、Nr数据库进行序列比对,获得新基因的注释信息(图1)。
2.3 二倍体及其同源四倍体酸枣差异表达基因分析
对二倍体和四倍体酸枣的差异表达基因进行分析。结果表明,二倍体与四倍体酸枣共有1 329个基因差异表达(FC > 1.5, FDR < 0.05),其中四倍体酸枣有636个基因表达量高于二倍体,表达量倍数介于1.5 ~ 18.2之间;693个基因表达量低于二倍体,表达量倍数介于1.5 ~ 11.8之间(图2)。
![]() 图 2 二倍体与四倍体酸枣差异表达基因统计红色的点代表四倍体相对于二倍体表达量上调的基因,绿色的点代表表达量下调的基因,黑色的点代表没有差异的基因;FC代表差异倍数;FDR代表错误发现率。The red point indicates up-regulated genes in autotetraploid than that in diploid, green point indicates down-regulated genes and black point indicates unchanged genes; FC represents fold change; FDR represents false discovery rate.Figure 2. Statistics of differentially expressed genes in diploid and autotetraploid sour jujube
图 2 二倍体与四倍体酸枣差异表达基因统计红色的点代表四倍体相对于二倍体表达量上调的基因,绿色的点代表表达量下调的基因,黑色的点代表没有差异的基因;FC代表差异倍数;FDR代表错误发现率。The red point indicates up-regulated genes in autotetraploid than that in diploid, green point indicates down-regulated genes and black point indicates unchanged genes; FC represents fold change; FDR represents false discovery rate.Figure 2. Statistics of differentially expressed genes in diploid and autotetraploid sour jujube2.3.1 差异表达基因GO功能分类与富集分析
为了分析二倍体与四倍体差异基因的生物学功能,分别从生物学过程、细胞组分和分子功能3个方面对注释到GO Ontology数据库中的1 064个差异基因进行功能分类(表3)。在生物学过程分类中,参与细胞过程、单一生物过程和代谢过程的差异基因数目最多,分别为873、851和829个,其次是胁迫应答过程,为753个;在细胞组分分类中,细胞部位和细胞差异基因最多,分别为878和864个,其次是细胞器为714个。在分子功能分类中,参与催化活性的差异基因最多,为655个,其次是参与结合功能的差异基因为620个。
表 3 二倍体与四倍体酸枣差异表达基因的GO分类及富集分析Table 3. GO classification and enrichment analysis of differentially expressed genes in diploid and autotetraploid sour jujubeGO一级分类
GO classify 1GO ID GO二级分类
GO classify 2所有基因
All gene差异基因
Different gene富集P值
Enriched P value生物学过程
Biological processGO:0009987 细胞过程 Cellular process 15 527 873 0.725 1 GO:0008152 代谢过程 Metabolic process 14 536 829 0.983 6 GO:0044699 单一生物过程 Single-organism process 14 286 851 0.004 2 GO:0050896 胁迫应答 Response to stimulus 11 883 753 0.000 0 GO:0065007 生物调控 Biological regulation 9 802 603 0.021 6 GO:0032502 发育过程 Developmental process 8 154 492 0.003 7 GO:0071840 细胞组分或生物合成
Cellular component organization or biogenesis7 166 386 0.373 7 GO:0051179 定位 Localization 6 533 378 0.054 4 GO:0032501 多细胞生物过程 Multicellular organismal process 6 469 372 0.090 1 GO:0000003 生殖 Reproduction 5 573 331 0.070 8 GO:0022414 生殖过程 Reproductive process 5 536 332 0.065 2 GO:0051704 多生物过程 Multi-organism process 4 643 249 0.002 4 GO:0023052 信号传导 Signaling 3 681 237 0.184 7 GO:0040007 生长 Growth 2 639 142 0.016 5 GO:0002376 免疫系统过程 Immune system process 2 446 131 0.001 1 GO:0098754 解毒 Detoxification 550 49 GO:0048511 节律过程 Rhythmic process 294 26 0.064 5 GO:0022610 生物附着 Biological adhesion 134 7 0.413 8 GO:0040011 运动 Locomotion 73 4 0.295 6 GO:0001906 细胞杀伤 Cell killing 15 2 0.556 6 细胞组分
Cellular componentGO:0044464 细胞部位 Cell part 16 088 878 0.000 0 GO:0005623 细胞 Cell 15 666 864 0.713 8 GO:0043226 细胞器 Organelle 12 958 714 0.659 2 GO:0016020 细胞膜 Membrane 8 588 489 0.740 6 GO:0044422 细胞器部位 Organelle part 5 560 302 0.938 5 GO:0044425 细胞膜部位 Membrane part 2 828 163 0.948 7 GO:0032991 高分子复合物 Macromolecular complex 2 285 81 0.999 8 GO:0030054 细胞连接点 Cell junction 2 259 154 0.438 3 GO:0005576 胞外区 Extracellular region 2 091 173 0.000 2 GO:0031974 膜封闭腔 Membrane-enclosed lumen 563 21 0.284 9 GO:0099080 超分子复合物 Supramolecular complex 154 14 GO:0044421 胞外区部位 Extracellular region part 132 11 0.428 4 GO:0009295 拟核 Nucleoid 45 4 0.058 4 分子功能
Molecular functionGO:0005488 结合 Binding 11 517 620 0.983 3 GO:0003824 催化活性 Catalytic activity 10 913 655 0.587 2 GO:0005215 转运活性 Transporter activity 1 496 91 0.801 3 GO:0001071 核酸结合转录因子活性
Nucleic acid binding transcription factor activity1 062 76 0.327 1 GO:0004871 信号传导活性 Signal transducer activity 782 23 0.216 1 GO:0060089 分子传感器活动 Molecular transducer activity 627 21 0.216 1 GO:0005198 结构分子活动 Structural molecule activity 437 16 0.914 3 GO:0009055 电子载体活动 Electron carrier activity 350 27 0.040 0 GO:0098772 分子功能调节剂 Molecular function regulator 285 23 GO:0016209 抗氧化活性 Antioxidant activity 179 15 0.489 6 GO:0000988 转录因子活性,蛋白结合
Transcription factor activity, protein binding78 3 0.763 7 GO:0045735 营养库活动 Nutrient reservoir activity 45 1 0.154 2 利用KS检验,以富集P值小于0.05作为评价标准,在二倍体与四倍体差异表达基因中满足此条件的GO条目即为GO功能显著富集。在生物过程分类中显著富集了包括胁迫应答过程、生物调控、发育过程、生长过程和免疫系统过程等7个条目;在细胞组分类别中显著富集的GO条目有2个,包括细胞部位和胞外区组成;在分子功能类别中,显著富集的条目只有1个,为电子载体活动。结果说明,二倍体与四倍体酸枣在生长发育、细胞代谢过程、逆境胁迫抗性等方面存在差异。
2.3.2 差异表达基因KEGG Pathway富集分析
二倍体与四倍体酸枣共有244个差异表达基因注释到特定的KEGG 通路中,这些通路可以分为细胞过程、环境信息加工、遗传信息加工、代谢和生物系统5类,其中在代谢中注释到的差异基因数目最多,占差异基因总注释数目的73.8%(图3)。有13条通路在KEGG Pathway中显著富集(P < 0.05),(图4),主要包括氨基酸代谢、碳水化合物代谢、信号传导、能量代谢等。碳水化合物代谢通路中,参与淀粉和蔗糖代谢途径的差异表达基因数目最多,占注释的差异表达基因总数的10.66%;信号传导通路中,参与植物激素信号转导途径的差异基因有18个,占注释的差异表达基因总数的7.38%;在氨基酸代谢通路中,丙氨酸,天门冬氨酸和谷氨酸代谢、精氨酸和脯氨酸代谢以及酪氨酸代谢3个途径差异基因数目分别占注释的差异基因总数的2.5%、3.3%和3.7%;在能量代谢通路中,光合天线蛋白途径中注释的差异基因占注释差异基因总数的1.2%。此外,相比二倍体植株,四倍体植株中表达量增加的差异基因显著富集在淀粉与蔗糖代谢(P < 0.000 9)、植物激素信号传导(P < 0.004 0)等通路中。以上结果表明,二倍体与四倍体酸枣在碳水化合物和氨基酸代谢、信号传递以及能量代谢途径中存在差异,其中,与淀粉和蔗糖代谢、植物激素传导途径相关的基因在四倍体中显著上调。
![]() 图 3 二倍体与四倍体酸枣差异表达基因KEGG分类Figure 3. KEGG classification of differentially expressed genes in diploid and autotetraploid sour jujube
图 3 二倍体与四倍体酸枣差异表达基因KEGG分类Figure 3. KEGG classification of differentially expressed genes in diploid and autotetraploid sour jujube![]() 图 4 二倍体与四倍体酸枣差异表达基因显著富集的KEGG通路Figure 4. KEGG pathway with significantly enriched differentially expressed genes in diploid and autotetraploid sour jujube
图 4 二倍体与四倍体酸枣差异表达基因显著富集的KEGG通路Figure 4. KEGG pathway with significantly enriched differentially expressed genes in diploid and autotetraploid sour jujube2.3.3 差异基因表达量分析
为进一步解析二倍体及其同源四倍体可溶性物质的差异,对与糖和蛋白质转运、代谢相关的16个基因进行差异表达分析(图5)。结果发现,与糖代谢相关的关键基因,如编码β-呋喃果糖苷酶的INV(CCG007195)基因、编码β-葡萄糖苷酶的BGLU47(CCG000062)基因、编码果胶酯酶的PME35(CCG011493)基因、编码葡糖醛酸异构酶的GAE6(CCG027883)基因、编码α-海藻糖磷酸合成酶的TPS9(CCG011925)基因和编码蔗糖磷酸合成酶的SPS2(CCG011942)基因等均上调表达,差异表达倍数介于1.5 ~ 2.5之间。与氨基酸转运和代谢相关的基因,例如编码丝氨酸乙酰转移酶的SAT1(CCG028964)基因、编码精氨琥珀酸合酶(NewGene3586)和磷酸甘油酸脱氢酶PGDH3(CCG002072)等的基因在四倍体酸枣中上调表达,表达量为二倍体的1.7 ~ 2.0倍。
![]() 图 5 二倍体与四倍体差异基因表达量热图D1、D2、D3分别为二倍体酸枣的3个生物学重复;T1,T2,T3分别为四倍体酸枣的3个生物学重复。方块中不同的颜色表示基于FPKM值的基因表达水平;每行的数据分别进行标准化;蓝色表示在四倍体中表达量下调,红色表示在四倍体中表达量上调。D1, D2 and D3 are the three duplicates of diploid sour jujube; T1, T2 and T3 were the three duplicates of autotetraploid sour jujube. Different colors indicate different levels of gene expression based on the FPKM values; the data in each row were normalized and compared, separately; blue indicates down-regulation in autotetraploid, and red indicates up-regulation.Figure 5. Heat map of differentially expressed genes in the diploid and autotetraploid sour jujube
图 5 二倍体与四倍体差异基因表达量热图D1、D2、D3分别为二倍体酸枣的3个生物学重复;T1,T2,T3分别为四倍体酸枣的3个生物学重复。方块中不同的颜色表示基于FPKM值的基因表达水平;每行的数据分别进行标准化;蓝色表示在四倍体中表达量下调,红色表示在四倍体中表达量上调。D1, D2 and D3 are the three duplicates of diploid sour jujube; T1, T2 and T3 were the three duplicates of autotetraploid sour jujube. Different colors indicate different levels of gene expression based on the FPKM values; the data in each row were normalized and compared, separately; blue indicates down-regulation in autotetraploid, and red indicates up-regulation.Figure 5. Heat map of differentially expressed genes in the diploid and autotetraploid sour jujube对与内源激素相关的23个差异基因表达量进行比较分析(图5),结果发现,在二倍体植株中,编码油菜素内酯生物合成相关蛋白(CYP)的基因(CCG015397、CCG007876、CCG007877)的表达量是四倍体的1.8 ~ 2.1倍;编码乙烯不敏感蛋白的基因EIN2(CCG001434)的表达量是四倍体的1.5倍。在四倍体植株中,与诱导、响应和转运生长素相关的基因,如ARG7(Addgene2586)、ARF9(CCG010227)、GH3.6(CCG025779)、IAA26(CCG025671、CCG025726)、LAX3(CCG001581)、X15(CCG013493)的表达量是二倍体的1.5 ~ 2.1倍。说明酸枣四倍体在表型上呈现的变异特征可能与这些关键基因的差异表达有关。
2.4 二倍体及其同源四倍体酸枣的差异转录因子分析
转录因子参与了许多生物学进程,在调控下游功能基因方面起重要作用。为了解酸枣加倍后的转录因子变异,对二倍体和四倍体酸枣进行了转录因子功能预测,发现与转录因子相关的差异基因共158个(FC > 1.5, FDR < 0.05),包含14个编码转录抑制子的基因,58个蛋白激酶相关基因和86个编码转录因子的基因,共编码35种转录因子(图6)。编码AP2/ERF-ERF转录因子的基因数目最多,其次为HB-HD-ZIP、LOB和MYB转录因子。四倍体植株中表达量高于二倍体的编码转录因子的基因有102个,属于28种转录因子,其中,编码MYB转录因子的基因最多。
![]() 图 6 编码转录因子的差异基因数目统计Figure 6. Statistics of the number of differentially expressed genes encoding transcription factors
图 6 编码转录因子的差异基因数目统计Figure 6. Statistics of the number of differentially expressed genes encoding transcription factors3. 讨 论
3.1 二倍体与四倍体生理特征差异
四倍体通常较二倍体具有更大的器官、细胞、更多的次生代谢物含量[21],这使得四倍体成为植物育种专家增加植物产量的重要手段之一。本研究中发现四倍体酸枣的叶绿素含量、可溶性糖和可溶性蛋白含量均显著高于二倍体。这与白桦(Betula platyphylla)[22]和刺槐(Robinia pseudoacacia)[23]基因组加倍后植株的生理表现相同。这些生理指标的差异可能是四倍体植株形态异于二倍体的重要原因之一,叶绿素含量高可能导致四倍体叶色更深,而可溶性糖和可溶性蛋白含量高为植株提供更多的能源物质,从而导致四倍体叶片更大、茎更粗。进一步说明,叶片细胞中的叶绿素含量、可溶性糖含量和可溶性蛋白质含量均可作为初步鉴定四倍体酸枣的辅助指标。但是,研究中发现叶片相对含水量在二倍体与四倍体酸枣中没有显著差异,这与四倍体连翘(Forsythia suspensa)[24]、四倍体甜瓜(Cucumis melo)[25]的研究结果表现一致。
3.2 二倍体与四倍体差异基因分析
植物多倍化后引起表型及生理的变异,这主要是由于基因表达改变引起的。大量研究发现植物中基因表达发生微小的变化都可能引起表型的变异[26]。高通量测序技术已经成为多倍体植物优良性状解析的常用策略[27],本研究中我们利用RNA-seq技术对酸枣二倍体和四倍体进行转录组学分析,测序数据分析结果表明测序结果良好。基因差异表达是四倍体酸枣变异的分子基础。因此,我们进一步对二倍体与四倍体酸枣的差异表达基因进行分析,二者共有1 329个基因差异表达,通过GO Ontology分析表明在胁迫应答、生物调控、发育、生长过程和免疫系统7个过程中差异基因富集,说明四倍体在生长发育、抗性方面与二倍体存在差异。KEGG通路分析发现差异基因显著富集在碳水化合物和氨基酸代谢、信号传递以及能量代谢方面,其中在淀粉与蔗糖代谢、植物激素传导通路中显著上调表达。这些差异基因在四倍体中的表达量升高可能参与了可溶性糖和内源生长素的积累过程,为四倍体提供更多能源物质,从而增强植株生长势。这一结果与杨树三倍体研究中发现的主要参与碳水化合物代谢、细胞生长的基因上调使其在生长表现上优于二倍体的结果相同[28]。
3.3 二倍体与四倍体差异基因功能分析
酸枣经基因组加倍后,表现为合成更多的可溶性糖和可溶性蛋白,这可能与众多关键基因在四倍体中的差异表达有关。前人研究表明,GAE6催化拟南芥半乳糖醛酸的合成,促进果胶多糖的形成[29];SPS控制光合产物向蔗糖和淀粉的分配,在蔗糖的形成过程中起正向调节作用[30]。在本研究中GAE6和SPS2在四倍体酸枣中上调表达,可能是导致四倍体植株体内可溶性糖含量增加的关键基因。此外,BGLU47、TPS9和SPS2编码糖代谢过程中的关键酶,可以通过调节植物体内渗透胁迫进而增强植物承受逆境胁迫的能力[31]。本研究对这些关键调控基因分析发现,此类基因在四倍体中具有较高的表达量,推测四倍体酸枣可能具有较强的渗透调节能力,具体性状还需要进一步的实验研究。植物磷酸丝氨酸途径在植物发育和次生代谢产物的生物合成中具有重要作用,PGDH3编码催化这一途径的PGDH酶,与色氨酸的生物合成呈正相关[32]。本研究中四倍体内PGDH3表达量显著高于二倍体,可能是导致四倍体酸枣体内可溶性蛋白含量较高的原因之一。
植物生长发育与内源激素的调节有密不可分的联系,内源激素的水平会显著影响细胞分化、糖合成与代谢等生理过程[33]。青杨(Populus cathayana)三倍体中参与生长素、细胞分裂素、油菜素内酯等激素合成与转导的基因相比二倍体显著上调表达导致三倍体生长势增强[34];桑树(Morus alba)四倍体与二倍体的差异基因参与了生长素和乙烯等激素的合成及传导过程,使四倍体较二倍体具有更大的生长优势[35]。在本研究中,四倍体与二倍体有18个差异基因显著富集在植物激素信号传导通路中,其中12个基因在四倍体中表达量较高,包括与生长素诱导相关的基因ARG7(SAUR家族)、GH3.6(GH3家族)、X15(SAUR家族),生长素响应基因IAA26(AUX/IAA家族),编码转运蛋白的基因LAX3(AUX1家族)以及生长素应答因子ARF9(ARF家族)等(图7),这些基因不仅正调控IAA的生物合成,还参与了激素的信号转导过程[36-38]。6个基因在四倍体中表达量较低,包括细胞色素P450单加氧酶(CYP)和乙烯不敏感蛋白(EIN)等,CYP参与甾醇催化形成油菜素甾醇的过程,从而促进叶片伸展和茎的伸长生长[39];EIN2是正调控乙烯信号传导通路的中心元件,与器官的衰老、成熟有关[40-41]。因此,编码内源激素的差异基因表达量的变化可能是导致四倍体生长表型变异的重要原因之一,对此还需要进一步实验验证。
![]() 图 7 与内源生长素相关的表达通路方块内的红色背景代表基因在四倍体中上调表达,蓝色背景代表基因在四倍体中既有上调表达又有下调表达。The red background in the square indicates up-regulated expression of genes in autotetraploid than that of diploid, while the blue background indicates up-regulated and down-regulated expressed genes.Figure 7. Expression pathway related to endogenous auxin
图 7 与内源生长素相关的表达通路方块内的红色背景代表基因在四倍体中上调表达,蓝色背景代表基因在四倍体中既有上调表达又有下调表达。The red background in the square indicates up-regulated expression of genes in autotetraploid than that of diploid, while the blue background indicates up-regulated and down-regulated expressed genes.Figure 7. Expression pathway related to endogenous auxin3.4 二倍体与四倍体转录因子差异分析
转录因子通过与下游基因相互作用,激活或抑制其他基因的转录,或者与其他转录因子互作调节基因的表达,从而引起一系列的应答反应,改变植物性状[42]。MYB转录因子广泛参与了植物的生长发育与形态建成[43]、次生代谢[44]和环境胁迫应答[45]等过程。在本研究中,我们分析二倍体和四倍体酸枣的差异转录因子,发现四倍体植株中编码MYB转录因子的基因在所有转录因子类型中上调表达数目最多。这一结果与菘蓝(Isatis indigotica)四倍体研究中发现的MYB在根、茎、叶中表达量高于二倍体的结果相一致[46],因此,四倍体在表型和生理方面的变异特征可能与编码MYB转录因子的基因上调表达有关。此外,MYB转录因子的差异表达还导致了菘蓝四倍体抗性增强,说明酸枣四倍体在对环境胁迫耐受性方面可能高于二倍体。因此,今后应该进一步对酸枣四倍体的抗性进行研究。
多倍体植物在基因组复制后含有新的基因调控网络,完整的RNA-seq数据对于理解新的调控网络至关重要。本研究在检测生理指标的基础上,对酸枣不同倍性植株进行转录组测序并结合性状分析了差异表达基因的功能,为深入理解酸枣四倍体的变异性状提供参考,通过进一步的研究筛选候选基因,为枣树关键调控基因的克隆及基因工程育种提供分子基础。
-
![]()
图 1 基本模型(A)、广义模型(B)、基础混合效应模型(C)和广义混合效应模型(D)的残差分布图
Figure 1. Residual plots of basic model (A), generalized model (B), basic mixed effect model (C), and generalized mixed effect model (D)
![]()
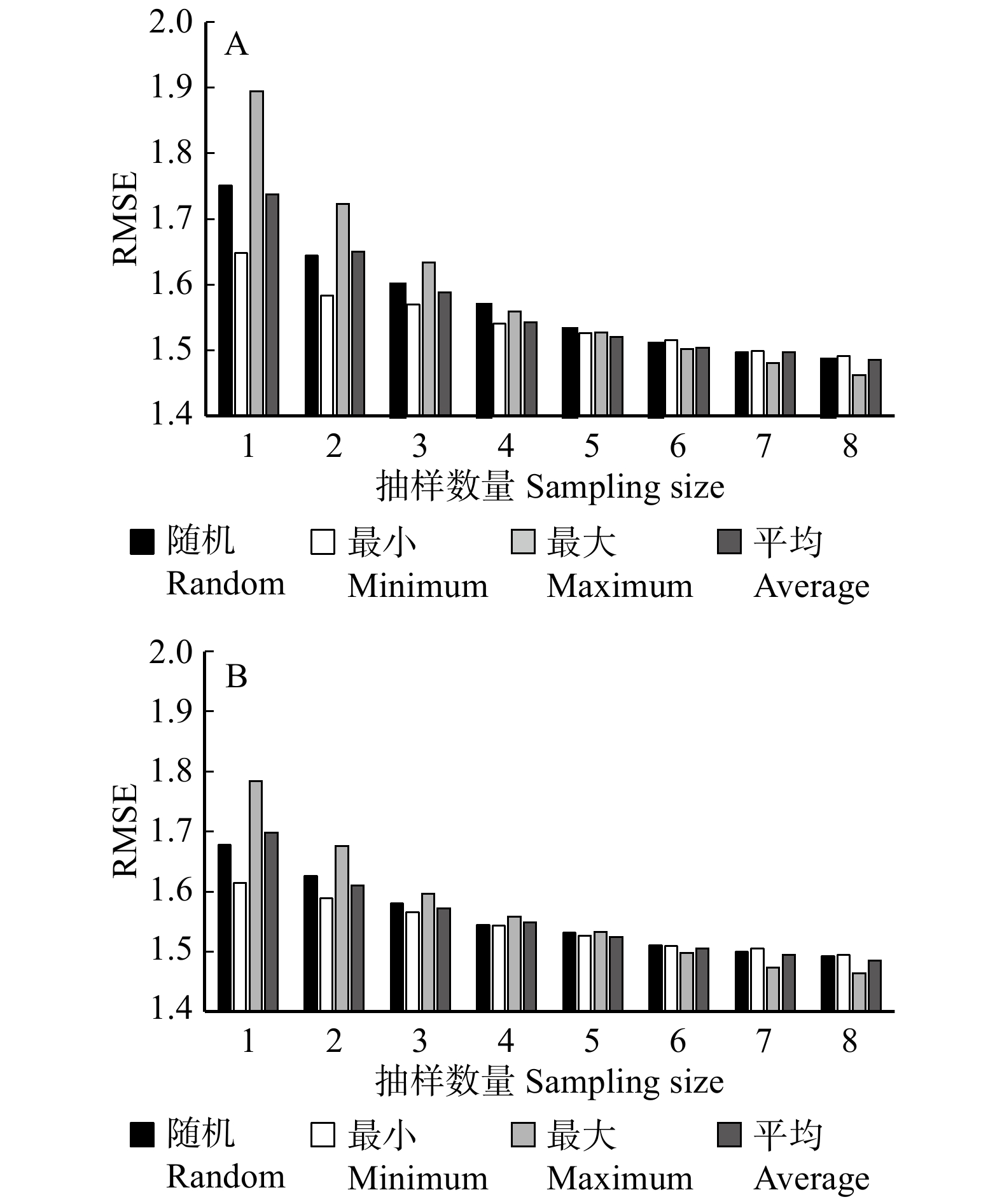
图 2 不同抽样设计下基础混合效应模型(A)和广义混合效应模型(B)的预测精度
Figure 2. Prediction accuracy of basic mixed effect model (A) and generalized mixed effect model (B) based on different sampling designs
表 1 红松人工林建模数据和检验数据基本统计量
Table 1 Statistics of fitting data and validation data of Korean pine plantation
数据类型
Data type变量
Variable均值
Mean最小值
Minimum最大值
Maximum标准差
Standard
deviation变异系数
Coefficient of
variation建模数据
Fitting data枝下高 Height to crown base (HCB)/m 5.27 1.10 13.20 2.49 47.24 树高 Tree height (H)/m 10.86 3.30 19.60 3.16 29.12 胸径 DBH (D)/cm 14.78 5.00 36.50 5.99 40.53 高径比 Height-diameter ratio (RHD) 0.78 0.43 2.68 0.17 22.05 大于对象木的断面积和/(m2·hm−2)
Basal area sum larger than subject tree (BAL)/(m2·ha−1)1.01 0.00 2.56 0.50 49.54 冠幅 Crown width (CW)/m 1.66 0.33 4.30 0.44 26.46 优势木高 Dominant height (HD)/m 17.88 14.90 20.62 1.23 6.86 检验数据
Validation data枝下高 HCB/m 3.62 0.80 12.10 1.89 52.25 树高 H/m 9.32 3.20 17.90 2.78 29.81 胸径 DBH (D)/cm 11.72 5.00 28.20 4.57 38.98 高径比 RHD 0.83 0.50 1.59 0.16 19.13 大于对象木的断面积和/(m2·hm−2)
BAL/(m2·ha−1)0.87 0.00 1.40 0.31 35.45 冠幅 CW/m 1.70 0.78 3.83 0.40 23.73 优势木高 HD/m 17.93 16.86 19.47 0.81 4.54  下载: 导出CSV
下载: 导出CSV
表 2 候选基础枝下高模型
Table 2 Model of HCB candidate
模型
Model模型表达式
Model expression参考文献
Reference(1) HCB=H/(1+exp(X)) [31] (2) HCB=H/√(1+exp(X)) [14] (3) HCB=H/6√(1+exp(X)) [14] (4) HCB=H(1−exp(X)) [29] (5) HCB=H(a+exp(X)) [30] (6) HCB=H(1−aexp(X2) [31] (7) HCB=H/c√(1+aexp(X)) [14] (8) HCB=H(1−aexp(Xc)) [14] 注: a、c为模型参数; X 为关于林木大小、竞争因子、立地条件的函数,在表中特指X = b0 + b1D,b0、b1为模型参数。下同。Notes: a and c refer to model parameters; X is the function about tree size competition index and site condition, here in the table, X = b0 + b1D, b0, b1 are model parameters. The same below.
下载: 导出CSV
表 3 候选模型评价指标和拟合参数
Table 3 Evaluation indices and fitting parameters of candidate model
模型
Model参数
Parameter参数估计值
Parameter estimate拟合优度
Goodness-of-fit statisticsR2 RMSE (1) b0 0.396 4 0.579 5 1.614 8 b1 −0.020 0 (2) b0 1.633 6 0.577 9 1.617 7 b1 −0.026 9 (3) b0 5.085 8 0.577 7 1.618 3 b1 −0.050 0 (4) a 0.561 9 0.578 5 1.616 7 b0 0.039 0 b1 −0.008 4 (5) a 不收敛
No convergenceb0 b1 (6) a 不收敛
No convergenceb0 b1 (7) a 不收敛
No convergenceb0 b1 c (8) a 不收敛
No convergenceb0 b1 c
下载: 导出CSV
表 4 不同预测变量组合模型拟合结果
Table 4 Model fitting results based on different prediction variables
变量组合 Variable combination R2 MAE RMSE — 0.567 0 1.284 0 1.638 6 大于对象木的断面积和−优势木高 BAL-HD 0.683 9 1.087 8 1.400 1 大于对象木的断面积和−林分密度 BAL-N 0.671 1 1.113 2 1.428 1 大于对象木的断面积和−冠幅 BAL-CW 0.687 4 1.080 3 1.392 3 大于对象木的断面积和−优势木高−冠幅 BAL-HD-CW 0.700 0 1.050 2 1.363 8 大于对象木的断面积和−高径比−优势木高 BAL-RHD-HD 0.690 0 1.070 5 1.386 5 大于对象木的断面积和−优势木高−林分密度 BAL-HD-N 0.690 0 1.088 7 1.386 5 注:—表示除胸径外,无林分变量添加。Notes: — indicates the models without other stand variables except D.
下载: 导出CSV
表 5 广义非线性混合模型评价指标
Table 5 Evaluation indices of generalized nonlinear mixed model
随机效应参数
Random effect parameterAIC BIC LOGLIK LRT P值
P value— 8 167.35 8 207.88 −4 076.67 b0 7 987.10 8 027.63 −3 985.55 182.24 < 0.000 1 b0,d2 7 969.87 8 021.98 −3 975.93 19.24 < 0.000 1 b0,b1,d2 7 815.16 7 890.44 −3 894.58 162.7 < 0.000 1 注:—表示无随机参数。Note:— indicates the models without random parameters.
下载: 导出CSV
表 6 模型参数估计和模型拟合统计量
Table 6 Parameter estimates and fitting statistics for each model
项目
Item参数
Parameter不含随机效应模型 Model without random effect 含随机效应模型 Model with random effect 基础模型
Base model广义模型
Generalized model基础混合效应模型
Base mixed effect model广义混合效应模型
Generalized mixed effect model固定参数
Fixed parameterb0 0.396 4 −1.201 6 0.020 2 −1.739 8 b1 0.020 0 −0.043 9 0.010 5 −0.024 7 c1 0.093 9 0.112 8 d1 −0.363 3 −0.276 2 d2 0.383 7 0.285 9 方差组成
Composition of varianceσ2b0 0.416 6 0.135 4 σ2b1 0.000 2 0.000 3 σ2d2 0.054 6 σb0b1 −0.008 9 −0.000 7 σb0d2 −0.031 9 σb1d2 −0.003 0 拟合统计量
Fitting statisticsR2 0.578 1 0.699 5 0.766 7 0.775 2 MAE 1.284 0 1.051 8 0.926 2 0.905 5 RMSE 1.617 5 1.364 9 1.202 6 1.180 5 注: σ2b0 、σ2b1 、σ2d2 分别为b0 、b1 、d2 的方差;σb0b1 、σb0d2 、σb1d2 分别为b0与b1、b0与d2、b1与d2的协方差;MAE为平均绝对误差,RMSE为均方根误差。Notes:σ2b0 ,σ2b1 ,σ2d2 are the variance ofb0 ,b1 ,d2 , respectively;σb0b1 ,σb0d2 ,σb1d2 are the covariance of b0 and b1, b0 and d2, b1 and d2, respectively; MAE is mean absolute error, RMSE is root mean square error.
下载: 导出CSV
表 7 4种抽样方式RMSE对比
Table 7 Comparison of RMSE of four sampling methods
样本数
Sample size含随机效应基础模型 Basic model with random effect 含随机效应广义模型 Generalized model with random effect 随机抽取
Random sampling最小值
Minimum value最大值
Maximum value平均值
Average value随机抽取
Random sampling最小值
Minimum value最大值
Maximum value平均值
Average value1 1.751 2 1.648 7 1.894 7 1.737 9 1.635 1 1.691 9 1.704 4 1.684 3 2 1.644 4 1.583 0 1.722 5 1.650 1 1.588 8 1.624 7 1.636 6 1.636 4 3 1.600 6 1.569 5 1.633 5 1.588 3 1.550 6 1.580 8 1.569 2 1.572 5 4 1.569 0 1.540 6 1.558 9 1.542 5 1.524 3 1.533 7 1.537 5 1.545 0 5 1.532 2 1.526 4 1.527 6 1.520 1 1.513 2 1.514 9 1.514 3 1.511 9 6 1.510 4 1.515 4 1.502 3 1.503 7 1.493 3 1.482 3 1.490 7 1.485 2 7 1.497 5 1.499 5 1.481 0 1.496 9 1.479 3 1.466 0 1.471 8 1.472 7 8 1.485 5 1.491 0 1.462 0 1.485 3 1.468 1 1.448 2 1.450 7 1.446 5
下载: 导出CSV
-
[1] Hasenauer H, Monserud R. A crown ratio model for Austrian forests[J]. Forest Ecology and Management, 1996, 84(1/3): 49−60.
[2] Sharma R P, Bílek L, Vacek, Z, et al. Modelling crown width-diameter relationship for Scots pine in the central Europe[J]. Trees, 2017, 31(6): 1875−1889. doi: 10.1007/s00468-017-1593-8
[3] Kuprevicius A, Auty D, Achim A, et al. Quantifying the influence of live crown ratio on the mechanical properties of clear wood[J]. Forestry: An International Journal of Forest Research, 2013, 86(3): 361−369. doi: 10.1093/forestry/cpt006
[4] Monserud R A, Sterba H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria[J]. Forest Ecology and Management, 1996, 80(1/3): 57−80.
[5] 吕乐, 董利虎, 李凤日. 黑龙江省东部地区天然椴树单木冠幅预测模型[J]. 东北林业大学学报, 2019, 47(7):37−42. Lü L, Dong L H, Li F R. Individual tree crown width prediction models for natural Tilia tuan in eastern Heilongjiang Province[J]. Journal of Northeast Forestry University, 2019, 47(7): 37−42.
[6] Sharma R P, Vacek Z, Vacek S. Individual tree crown width models for Norway spruce and European beech in Czech Republic[J]. Forest Ecology and Management, 2016, 366: 208−220. doi: 10.1016/j.foreco.2016.01.040
[7] Fu L Y, Sharma R P, Hao K J, et al. A generalized interregional nonlinear mixed-effects crown width model for Prince Rupprecht larch in northern China[J]. Forest Ecology and Management, 2017, 389: 364−373. doi: 10.1016/j.foreco.2016.12.034
[8] Crecente-Campo F, Álvarez-González J G, Castedo-Dorado F, et al. Development of crown profile models for Pinus pinaster Ait. and Pinus sylvestris L. in northwestern Spain[J]. Forestry, 2013, 86(4): 481−491. doi: 10.1093/forestry/cpt019
[9] Ritson P, Sochacki S. Measurement and prediction of biomass and carbon content of Pinus pinaster trees in farm forestry plantations, southwestern Australia[J]. Forest Ecology and Management, 2003, 175(1/3): 103−117.
[10] Fu L Y, Zhang H R, Sharma R P, et al. A generalized nonlinear mixed-effects height to crown base model for Mongolian oak in northeast China[J]. Forest Ecology and Management, 2017, 384: 34−43. doi: 10.1016/j.foreco.2016.09.012
[11] McRoberts R, Hahn J T, Hefty G J, et al. Variation in forest inventory field measurements[J]. Canadian Journal of Forest Research, 1994, 24(9): 1766−1770. doi: 10.1139/x94-228
[12] Temesgen H, Lemay V, Mitchell S J. Tree crown ratio models for multi-species and multi-layered stands of southeastern British Columbia[J]. The Forestry Chronicle, 2005, 81(1): 133−141. doi: 10.5558/tfc81133-1
[13] Ritchie M W, Hann D W. Equations for predicting height to crown base for fourteen tree species in southwest Oregon[R]. Corvallis: Oregon State University, 1987.
[14] Rijal B, Weiskittel A R, Kershaw J A. Development of regional height to diameter equations for 15 tree species in the North American Acadian Region[J]. Forestry: An International Journal of Forest Research, 2012, 85(3): 379−390. doi: 10.1093/forestry/cps036
[15] Yang Y Q, Huang S M. Effects of competition and climate variables on modelling height to live crown for three boreal tree species in Alberta, Canada[J]. European Journal of Forest Research, 2018, 137(2): 153−167. doi: 10.1007/s10342-017-1095-7
[16] 段光爽, 李学东, 冯岩, 等. 基于广义非线性混合效应的华北落叶松天然次生林枝下高模型[J]. 南京林业大学学报(自然科学版), 2018, 42(2):170−176. Duan G S, Li X D, Feng Y, et al. Generalized nonlinear mixed-effects crown base height model of Larix principis-rupprechtii natural secondary forests[J]. Journal of Nanjing Forestry University (Natural Science Edition), 2018, 42(2): 170−176.
[17] Sharma R P, Vacek Z, Vacek S, et al. Modelling individual tree height to crown base of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.)[J]. PLoS ONE, 2017, 12(10): e0186394. doi: 10.1371/journal.pone.0186394
[18] Calama R, Montero G. Interregional nonlinear height-diameter model with random coefficients for stone pine in Spain[J]. Canadian Journal of Forest Research, 2004, 34(1): 150−163. doi: 10.1139/x03-199
[19] Yang Y Q, Huang S M, Meng S X, et al. A multilevel individual tree basal area increment model for aspen in boreal mixedwood stands[J]. Canadian Journal of Forest Research, 2009, 39(11): 2203−2214. doi: 10.1139/X09-123
[20] Temesgen H, Monleon V J, Hann D W. Analysis and comparison of nonlinear tree height prediction strategies for Douglas-fir forests[J]. Canadian Journal of Forest Research, 2008, 38(3): 553−565. doi: 10.1139/X07-104
[21] Crecente C F, Tomé M, Soares P, et al. A generalized nonlinear mixed-effects height-diameter model for Eucalyptus globulus L. in northwestern Spain[J]. Forest Ecology and Management, 2010, 259(5): 943−952. doi: 10.1016/j.foreco.2009.11.036
[22] Sharma R P, Breidenbach J. Modeling height-diameter relationships for Norway spruce, Scots pine, and downy birch using Norwegian national forest inventory data[J]. Forest Science and Technology, 2015, 11(1): 44−53. doi: 10.1080/21580103.2014.957354
[23] 段光爽, 李学东, 冯岩, 等. 华北落叶松天然次生林树高曲线的混合效应模型[J]. 南京林业大学学报(自然科学版), 2018, 42(2):163−169. Duan G S, Li X D, Feng Y, et al. Developing a height-diameter relationship model with mixed random effects for Larix principis-rupprechtii natural secondary forests[J]. Journal of Nanjing Forestry University (Natural Science Edition), 2018, 42(2): 163−169.
[24] 李婉婷, 姜立春, 万道印. 基于混合效应的兴安落叶松树高与胸径关系模拟[J]. 植物研究, 2014, 34(3):343−348. Li W T, Jiang L C, Wan D Y. Simulation of height-diameter relationships for Larix gmelinii based on mixed effects[J]. Bulletin of Botanical Research, 2014, 34(3): 343−348.
[25] 雷相东, 李永慈, 向玮. 基于混合模型的单木断面积生长模型[J]. 林业科学, 2009, 45(1):74−80. Lei X D, Li Y C, Xiang W. Individual basal area growth model using multi-level linear mixed model with repeated measures[J]. Scientia Silvae Sinicae, 2009, 45(1): 74−80.
[26] 李春明. 基于两层次线性混合效应模型的杉木林单木胸径生长量模型[J]. 林业科学, 2012, 48(3):66−73. Li C M. Individual tree diameter increment model for Chinese fir plantation based on two-level linear mixed effects models[J]. Scientia Silvae Sinicae, 2012, 48(3): 66−73.
[27] Calama R, Montero G. Multilevel linear mixed model for tree diameter increment in stone pine (Pinus pinea): a calibrating approach[J]. Sliva Fennica, 2005, 39(1): 394.
[28] 李想, 董利虎, 李凤日. 基于联立方程组的人工樟子松枝下高模型构建[J]. 北京林业大学学报, 2018, 40(6):9−18. Li X, Dong L H, Li F R. Building height to crown base models for Mongolian pine plantation based on simultaneous equations in Heilongjiang Province of northeastern China[J]. Journal of Beijing Forestry University, 2018, 40(6): 9−18.
[29] Wykoff W R, Crookston N L, Stage A R. User’s guide to the stand prognosis model[R].Ogden: Forest Service, United States Department of Agriculture, 1982.
[30] Popoola F S, Adesoye P O. Crown ratio models for Tectona grandis (Linn. f) stands in Osho Forest Reserve, Oyo State, Nigeria[J]. Journal of Forest & Environmental Science, 2012, 28(2): 63−67.
[31] Walters D K, Hann D W. Taper equations for six conifer species in southwest Oregon[M]. Corvallis: Oregon State University, 1986..
[32] 韩斐斐, 姜立春. 基于树干不同高度直径的落叶松立木材积方程[J]. 东北林业大学学报, 2017, 45(4):65−69. Han F F, Jiang L C. Tree volume function based on diameter at different relative heights of Dahurian larch[J]. Journal of Northeast Forestry University, 2017, 45(4): 65−69.
-
期刊类型引用(8)
1. 刘丽,王永康,苏万龙,赵爱玲,任海燕,薛晓芳,石美娟,李登科. 中国酸枣种质资源收集保存、鉴定评价及筛选利用进展. 西北农业学报. 2024(02): 191-200 .  百度学术
百度学术
2. Meng Li,Chenxing Zhang,Lu Hou,Xinru Liu,Hanqing Zhao,Xiaoming Pang,Wenhao Bo,Yingyue Li. Differences in leaf cuticular wax induced by whole-genome duplication in autotetraploid sour jujube. Horticultural Plant Journal. 2024(01): 66-76 . 必应学术
3. 杨亚杰,李昱樱,申状状,陈天,荣二花,吴玉香. 草棉不同倍性材料叶片转录组差异表达分析. 作物学报. 2022(11): 2733-2748 . 百度学术
4. 宋文龙,葛卉,刘兴菊,梁海永. 金叶榆二倍体及其同源四倍体的生理特征与转录组差异分析. 河北农业大学学报. 2022(04): 86-92 . 百度学术
5. 张秀珍,宋文龙,梁海永,王志彬. 四倍体白榆叶片形态特征和光合特性研究. 林业科技. 2022(06): 1-4 . 百度学术
6. 张子璇,谭明谱,王将,张成才,罗丽娜,向增旭. 茅苍术二倍体和同源四倍体叶片转录组比较. 植物资源与环境学报. 2021(04): 41-49 . 百度学术
7. 高真,张成才,罗丽娜,张子璇,王将,向增旭. 茅苍术二倍体及其同源四倍体的生理特征与转录组差异分析. 南京农业大学学报. 2020(06): 1024-1032 . 百度学术
8. 张成才,高真,罗丽娜,梁晖辉,向增旭. 霍山石斛同源四倍体与二倍体活性成分及转录组比较分析. 中国中药杂志. 2020(23): 5669-5676 . 百度学术
其他类型引用(14)







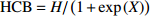
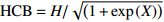
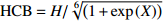
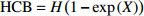
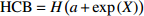
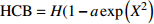
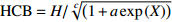
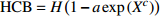


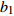
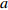
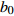
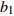

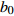
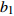
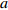
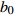


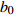
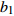
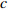
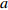

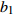

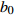

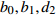
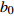



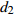
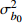
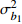
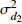


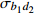


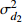
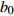
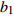
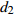
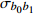
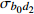
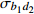

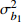
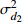
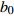



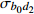
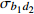

计量
- 文章访问数: 2099
- HTML全文浏览量: 801
- PDF下载量: 72
- 被引次数: 22
