
Global potential suitable area and ecological characteristics of Moringa oleifera
-
摘要:目的 预测辣木全球潜在适生区,并分析不同适生区的生态特征。探究不同适生区与辣木原产地的气候差异,为辣木引种和栽培管理措施提供参考。方法 通过国内外植物标本馆和相关研究文献得到辣木在全球的500个分布点,运用Maxent模型预测辣木在全球的潜在适生区,并基于不同时间尺度的环境因子和相关统计学方法分析不同适生区的生态特征。结果 辣木广泛分布于南北半球的热带、亚热带地区,北半球主要集中于南亚、东南亚、非洲西部地区。南半球主要位于非洲南部、南美洲。影响其地理分布的主要是6个环境变量,其中3个极端温度指标(最冷季度平均温度(bio11)、最冷月份最低温度(bio6)、最暖季度平均温度(bio10));一个温度变异幅度指标(温度变化方差(bio4));一个水热同步指标(最湿季度平均温度(bio8));以及一个极端水分条件(最湿季度降水量(bio16))。结论 辣木在全球范围内主要分布在北纬32°到南纬29°的区域,温度是影响辣木地理分布的主要环境因子。Abstract:Objective This paper aims to predict the global potential distributing area and analyze the ecological features of different suitable areas for introduction and cultivation management measures of Moringa oleifera.Method We predicted the potential distribution area by Maxent according to the 500 distribution points of M. oleifera, and analyzed the ecological features based on the environmental variables in different time scales and corresponding statistical method.Result M. oleifera was widely distributed in tropical and subtropical areas. In the northern hemisphere, it is mainly distributed in South Asia, Southeast Asia and West Africa. In the southern hemisphere, it is largely located in South Africa, South America. The variables affecting its geographical distribution were six climate factors, including three extreme temperature indicators (mean temperature of the coldest quarter (bio11), min. temperature of the coldest month (bio6) and mean temperature of the warmest quarter (bio10)); SD of temperature seasonality (bio4) indicating the temperature variation amplitude; mean temperature of the wettest quarter (bio8) denoting the hydrothermal synchronization; precipitation of the wettest quarter (bio16) representing the extreme moisture condition.Conclusion The potential distribution areas for M. oleifera were mainly located from north of 32°N to south of 29°S. Temperature is the main affecting environmental factor for M. oleifera distribution.
-
辣木(Moringa oleifera )是生长在热带及亚热带地区的一种高营养、多功效的天然绿色植物,具有降血压、降血脂、降血糖、抑制肿瘤等功效,被誉为“生命之树”[1-2]。辣木籽能够榨取食用油,亦称多油辣木,在印度传统医学的药用历史长达上千年之久,其根、茎、叶、花、种子等多个部位均含有多种生物活性物质,具有抗乙肝病毒、降血糖、抗氧化等生物活性[3-4]。另外,辣木在畜禽饲料、水质净化、工业生产等领域也有广泛应用[5-7]。因此,辣木是一种集食、油、药、饲料等为一体的新资源树种。
辣木喜温、喜光、耐旱、耐贫瘠、忌积涝,能耐受轻微低温和40 ℃以上的高温[8]。辣木较强的抗逆性和气候耐受性使得其能够适应不同的生态条件,在温度25 ~ 35 ℃左右,降水量300 ~ 3 000 mm的地方均适宜辣木生长[9-11]。辣木原产地位于印度北部的喜马拉雅山南麓地区,现广布于亚洲、非洲、南美洲等热带、亚热带地区,主要集中分布在印度、菲律宾、马来西亚、斯里兰卡、古巴、尼日利亚、墨西哥、巴西、加纳等热带国家,其中印度是目前最大的辣木产品出口国。在我国,辣木主要种植在云南、福建、广东和海南等地区。
近年来,国内外学者对辣木的研究多在于化学成分提取,药理活性及安全性,相关产品的开发,引种栽培技术,病虫害防治等方面,对辣木在全球潜在适生区的研究较少。辣木作为多用途经济树种,具有很高的经济价值和生态价值,有良好的发展前景。虽然辣木在全球许多地方都有引种,但是还没有从生态地理角度探究辣木种植的最佳区域以及新的适宜发展区。同时有研究表明,我国一些地区种植的辣木营养成分与原产地有一定差异性[12-14],为探究不同产地辣木成分及生长差异与各个产地气候等生态因子的关系,本文尝试从不同时间尺度的气候因子层面,探讨辣木不同适生区的气候差异,为不同地区的辣木引种或栽培管理措施提供参考。
1. 数据来源与方法
1.1 物种分布数据与环境图层数据
1.1.1 辣木分布数据
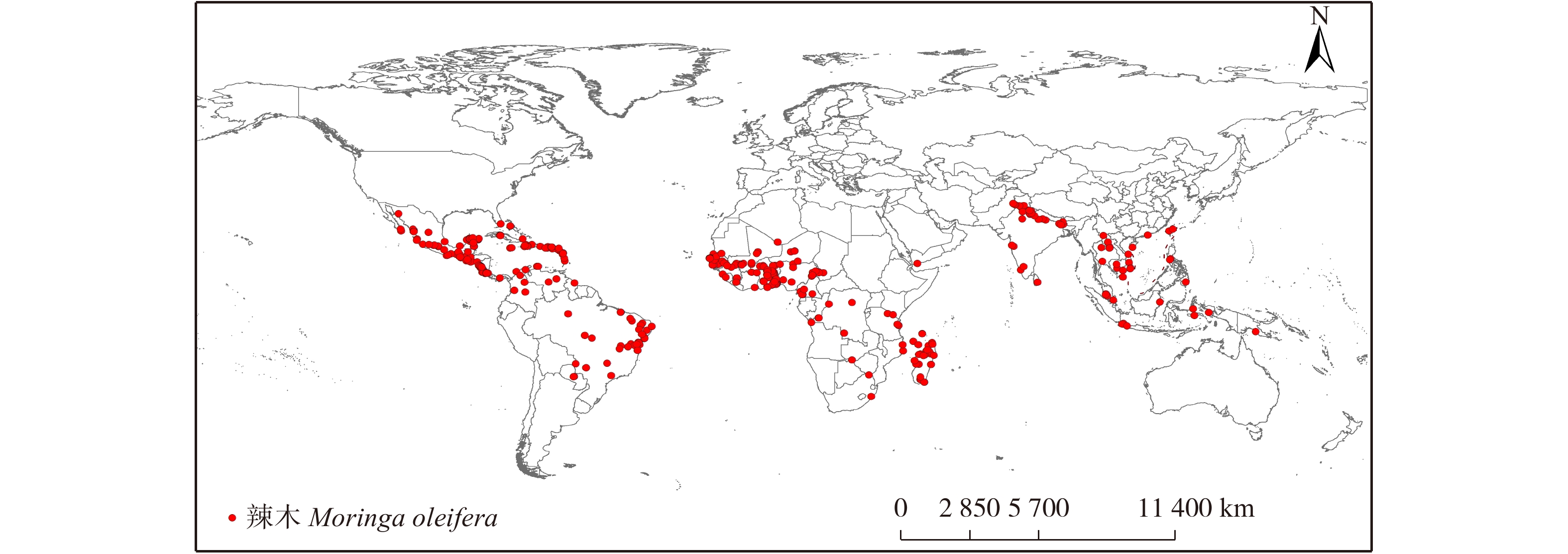
根据中国数字植物标本馆(CVH,http://www.cvh.org.cn/),全球生物多样性信息网络(GBIF,http://www.gbif.org/),中国科学院植物标本馆(http://www.nhpe.org/)等植物标本馆资源共享平台以及文献检索得到辣木的分布数据。由于所获取的分布点来源复杂多样,为保证所得辣木样本点信息准确且符合物种实际生境,基于以下方式对其进行核实和修订:保留拉丁名正确和经纬度详细的点,信息不全以及重复点均删除;有具体地址但无经纬度的点均以最小可查的行政单位为准在谷歌地球上查询,最后共获得505个辣木分布的有效记录(图1)。
1.1.2 环境变量数据
本文所采用的环境变量来源于世界气候数据库(http://www.worldclim.org/),为1970—2000年监测数据的平均值,空间分辨率为2.5°。分为两大类环境变量,第一类是综合气候因子,包括与温度及降水相关的19个生物气候变量[15],这19个气候因子作为综合气候变量,消除了南北半球在季节上的错移,可以用于预测物种的潜在适生区;第二类是月均气候因子(月均温、月降水),可以更精细分析和比较不同地区的气候差异。中国行政区划矢量图(1∶4 000 000)下载自国家基础地理信息系统(http://nfgis.nsdi.gov.cn/),作为本研究中绘图的地图数据。
1.1.3 相关软件来源
MAXENT3.3.3k软件(http://www.cs.princeton.edu/~schapire/maxent.)用于最大熵模型预测。生物地理信息系统平台ArcGIS 10.1来源于ESRI公司,主要进行数据的格式转换和图层的重分类。
1.2 方 法
1.2.1 环境变量选择
由于19个综合环境变量之间存在一定的相关性,故需要在运用Maxent模型进行适生区预测前对环境变量进行共线性诊断。首先将19个综合环境因子导入Maxent进行运算,去除贡献率为0的综合环境因子;其次选取剩余的环境因子在SPSS中进行Spearman相关性分析,对于相关系数 ≥ 0.8的两个变量,选择贡献率较大的综合环境因子,将最终筛选出的11个综合环境因子(表1)代入Maxent进行运算分析。
表 1 环境变量Table 1. Environmental variables反应指标 Response indicator 环境变量 Environmental variable 平均温度及其变异幅度
Average temperature and its variation range温度变化方差
SD of temperature seasonality (bio4)极端温度影响
Extreme temperature effect最冷月份最低温度、最暖季度平均温度、最冷季度平均温度
Min. temperature of the coldest month (bio6), mean temperature of the warmest quarter (bio10), mean temperature of the coldest quarter (bio11)降雨量及其季节性分布
Rainfall and its seasonal distribution雨量变化方差
Variance of rainfall variation (bio15)极端水分条件
Extreme moisture condition最湿季度降水量、最干季度降水量
Precipitation of the wettest quarter (bio16), precipitation of the driest quarter (bio17)水热是否同步
Hydrothermal synchronization or not最湿季度平均温度、最暖季度平均降水量、最冷季度平均降水量
Mean temperature of the wettest quarter (bio8), mean precipitation of the warmest quarter (bio18), mean precipitation of the coldest quarter (bio19)温差特点
Temperature difference characteristics昼夜温差月均值
Monthly mean temperature difference between day and night (bio2)月均变化
Monthly average change1—12月月均温度,1—12月月均降水量
Average monthly temperature and monthly average precipitation from January to December1.2.2 Maxent模型参数设置
将辣木现实分布点的数据(.csv)和环境变量数据导入MAXENT3.3中,随机选取75%的分布点用于建立模型,其余25%的分布点用于模型验证,选择利用Jackknife来检测各个环境变量在预测辣木分布区中的效用,并对各生境因子进行精确性分析,具体就是依次剔除1个环境变量,利用剩余变量重建模型。并再次利用所剔除变量和所有变量分别生成模型。以此来检验各环境变量在生成该物种潜在分布区时的重要性。其他参数均为模型的默认值,结果以Logistic格式输出。
1.2.3 适生区划分
将Maxent模型输出结果加载到ArcGIS中进行进一步分析,输出格式为ASCII的栅格图层,并与全球地图叠加。Maxent模型的预测结果在ArcGIS中以0 ~ 0.944的概率范围表示,即以存在概率值来表示物种分布区的适宜性。将Maxent模型结果生成的栅格数据按照人工分级法进行重分类,根据其适宜性指数对模型预测的辣木结果进行等级划分。
1.2.4 生态特征分析
通过RISGIM工作空间,提取辣木不同产地的综合气候因子以及更为细化的月均气候参数,运用统计学方法进行分析,并对结果进行可视化表达。
1.2.5 预测结果评价
Maxent自定义设置ROC(receive operating characteristic)评价曲线及AUC值对预测结果进行精度评测。受试者工作曲线下的面积即为AUC值,其不受阈值影响,数值范围为0 ~ 1,是目前认可度较高的诊断试验评价指标。AUC值在0.5 ~ 0.6,表明模型模拟效果为失败;0.7 ~ 0.8表明模拟效果一般;0.8 ~ 0.9表明模拟效果好;0.9 ~ 1.0表明模拟效果非常好[16]。
2. 结果与分析
2.1 Maxent模型预测结果评价
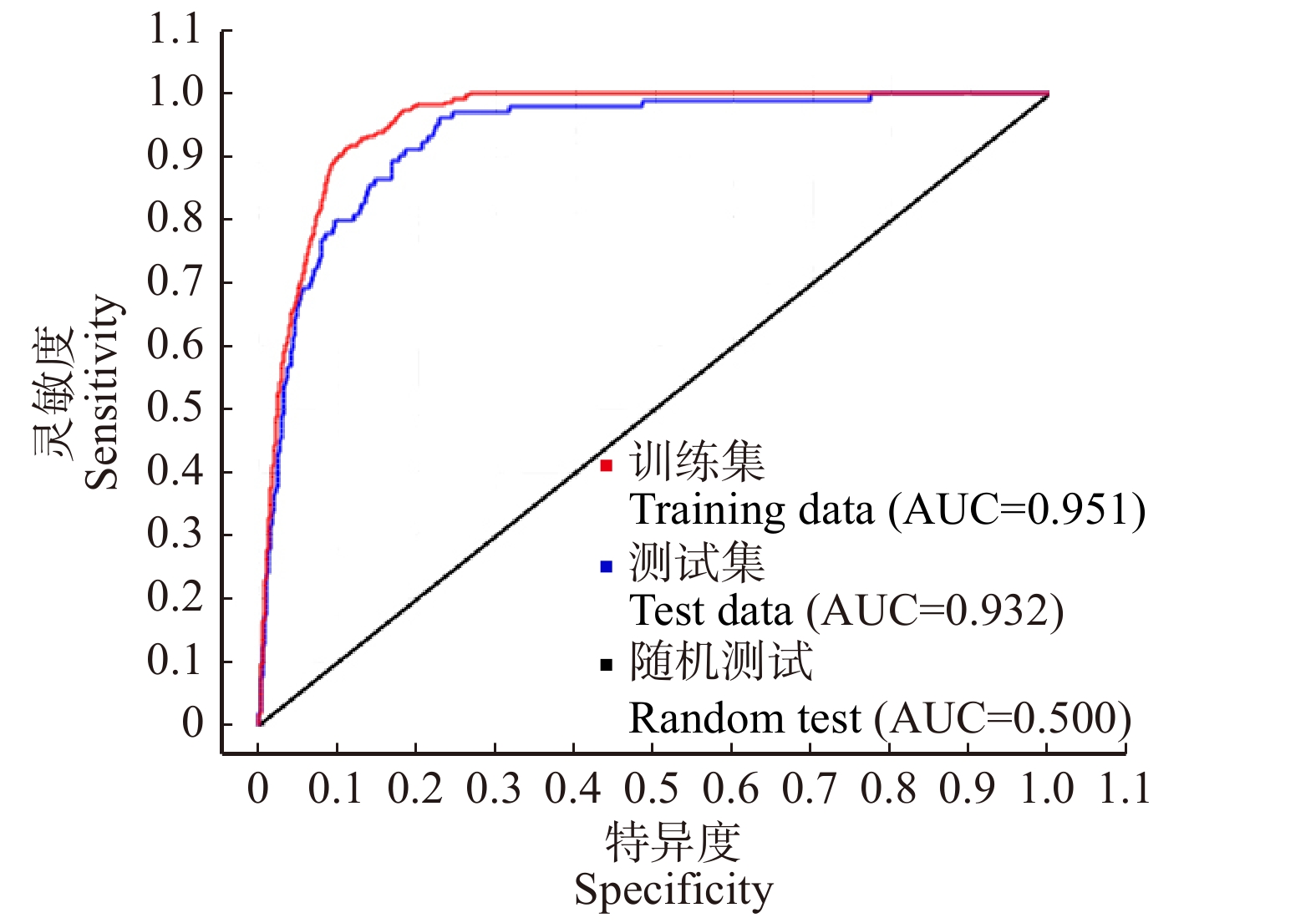
应用ROC曲线法对Maxent模型预测的辣木全球适生区结果(图2)进行精度检验,得到训练集AUC值为0.951;测试集AUC值为0.932;说明模型预测效果佳,预测结果有较高的精确度和可信度。
2.2 因子贡献率及适宜值范围
模型运用刀切法通过多次迭代运算,检验11个环境变量对辣木适生区的影响权重。图中红色条带表示用所有环境变量建立模型,绿色条带表示的是除该变量以外的其他各变量组合的贡献,蓝色条带表示单个变量的贡献。图3结果显示模型自动生成的各环境变量的贡献值概况。模型分析结果显示总贡献率达到91.7%的环境变量共6个因子,其中5个是温度因子,温度因子的累积贡献率为79.2%;另1个是降水因子,累积贡献率为12.5%,皆为影响辣木适生区的重要生态因子。这6个因子贡献率由大到小排列为:最冷季度平均温度(bio11)、最冷月份最低温(bio6)、最湿季度降水量(bio16)、温度变化方差(bio4)、最暖季度平均温度(bio10)、最湿季度平均温度(bio8)。
![]() 图 3 不同环境变量训练得分的Jackknife检验Figure 3. Jackknife test on training score of different environmental variables
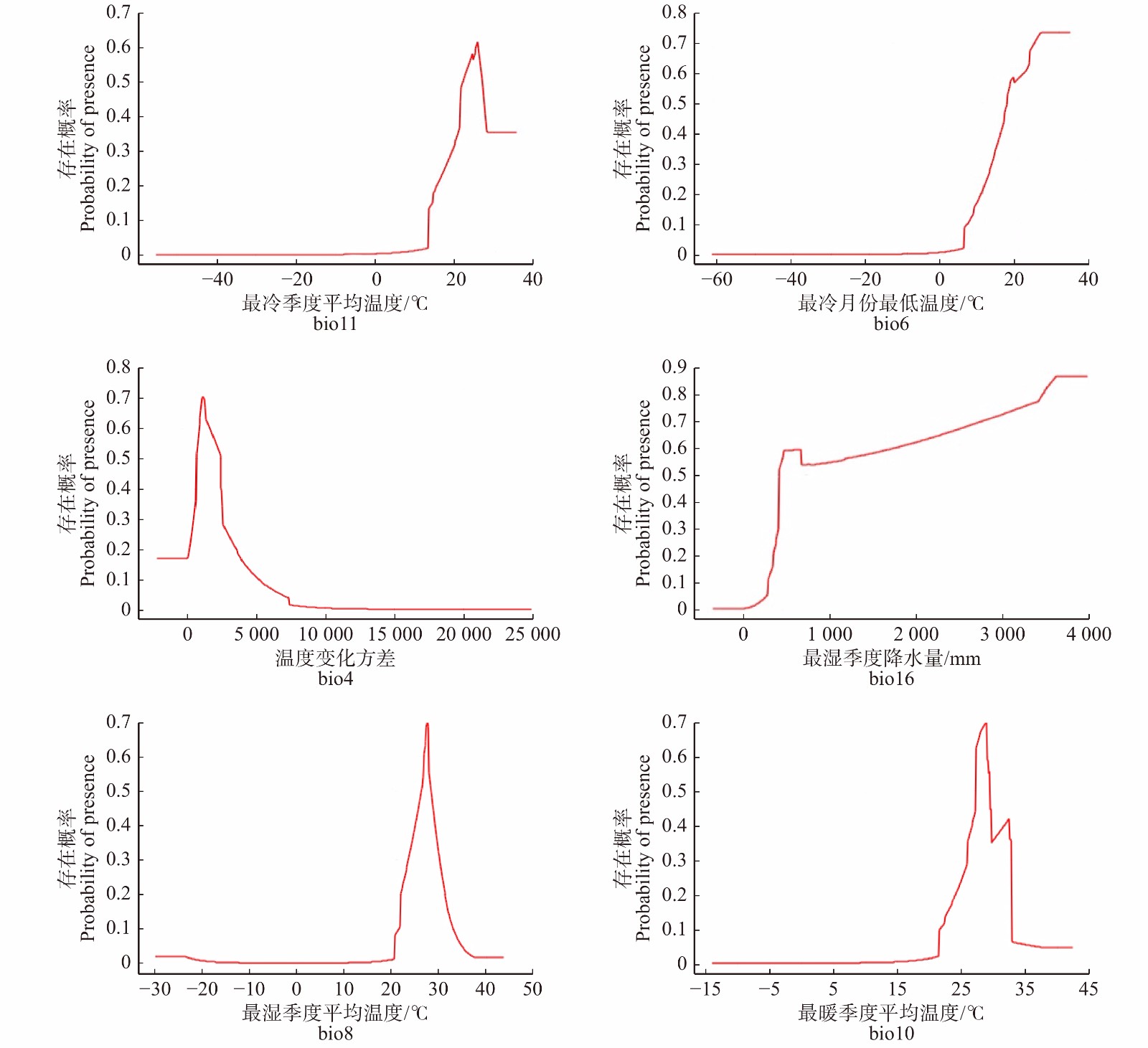
图 3 不同环境变量训练得分的Jackknife检验Figure 3. Jackknife test on training score of different environmental variables物种存在概率−环境影响因子响应曲线中(图4),一般认为存在概率 > 0.5(高适生区)所对应的环境变量被视为是该物种的环境变量适宜值范围,存在概率最高点对应环境变量的值则为最适宜值。本文选取了贡献率大于5%的6个综合环境变量,对其进行环境适宜值估算。辣木潜在适生区的最冷季度平均温度(bio11)适宜范围为21.9 ~ 27.1 ℃,最适宜值为25.7 ℃;最冷月份最低温(bio6)适宜范围为15.5 ~ 25 ℃,最适宜值为25 ℃;最湿季度降水量(bio16)适宜范围为409 ~ 3 590 mm,最适宜值为3 590 mm;温度变化方差(bio4)适宜范围为7.14 ~ 25.71,最适宜值为11.42;最湿季度平均温度(bio8)适宜值范围为25.8 ~ 28.5 ℃,最适宜值为27.7 ℃;最暖季度平均温度(bio10)适宜值范围为27.0 ~ 29.3 ℃,最适宜值为28.5 ℃。
2.3 不同适生区生态特征差异
为探索中国适生区与辣木原产适生区的气候差异性,本文从不同时间尺度的气候因子,系统分析云南、广西、广东、海南、台湾等地区与辣木原产地喜马拉雅南麓的气候特征差异。
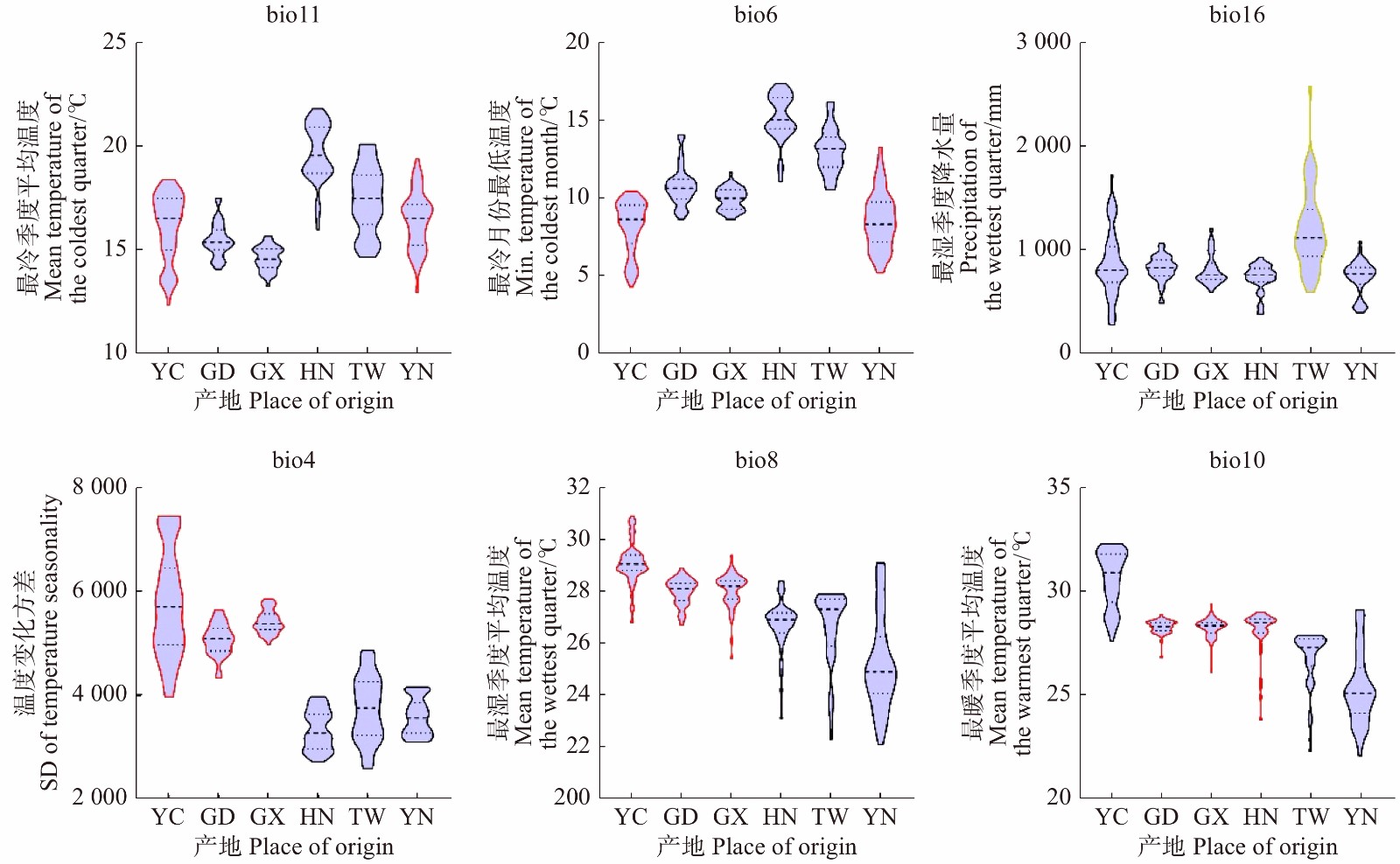
在综合气候因子尺度下(图5),分别对比原产地区与其他不同适生区的生态特征可知:
![]() 图 5 辣木原产地和中国分布区主要综合气候因子YC. 原产地区;GD. 广东;GX. 广西;HN. 海南;TW. 台湾;YN. 云南。下同。YC, original area; GD, Guangdong; GX, Guangxi; HN, Hainan; TW, Taiwan; YN, Yunnan. The same below.Figure 5. Main comprehensive climatic factors of M. oleifera in original area and the distributing area in China
图 5 辣木原产地和中国分布区主要综合气候因子YC. 原产地区;GD. 广东;GX. 广西;HN. 海南;TW. 台湾;YN. 云南。下同。YC, original area; GD, Guangdong; GX, Guangxi; HN, Hainan; TW, Taiwan; YN, Yunnan. The same below.Figure 5. Main comprehensive climatic factors of M. oleifera in original area and the distributing area in China(1)原产地与云南地区。两地区最冷季度平均温度(bio11)、最冷月份最低温(bio6)、最湿季度降水量(bio16)在参数分布上基本一致。最冷季度平均温(bio11)参数在原产地集中分布于15.1 ~ 17.5 ℃,在云南地区集中分布在15.3 ~ 17.2 ℃;最冷月份最低温(bio6)参数在原产地集中于7.2 ~ 9.5 ℃,在云南地区集中于7.2 ~ 9.7 ℃。最湿季度降水量(bio16)在原产地集中于679 ~ 1 030 mm,在云南地区集中于683 ~ 830 mm。
(2)原产地与广东、广西适生区。三地区温度变化方差(bio4)、最湿季度平均温度(bio8)参数分布相近。温度变化方差(bio4)在原产地区集中于4 977 ~ 6 428,在广东地区集中于4 861 ~ 5 279,在广西地区主要分布于5 274.25 ~ 5 559.5;最湿季度平均温度(bio8)在原产地区主要是28.8 ~ 29.4 ℃,广东地区为27.7 ~ 28.3 ℃,广西地区范围为27.7 ~ 28.4 ℃。
(3)原产地与海南、台湾地区。原产地区与海南地区的最湿季度降水量(bio16)在参数分布上较为一致,原产地区集中分布在679 ~ 1 030 mm,海南地区集中分布在701 ~ 817 mm。台湾地区没有任何一个综合因子参数与原产地区相近。
基于月均气候因子数据更精细比较不同地区的气候差异,分析结果如图6。
![]() 图 6 辣木原产区和中国分布区月均气候因子Figure 6. Monthly mean climatic factors of M. oleifera in original area and the distributing area in China
图 6 辣木原产区和中国分布区月均气候因子Figure 6. Monthly mean climatic factors of M. oleifera in original area and the distributing area in China(1)从月降水因子可以看出,云南产区和台湾产区与原产地喜马拉雅山南麓地区的降水趋势比较一致,都是在7、8月份达到最高,但在月均数值上云南更接近印度原产区,而台湾地区月均降水比原产区高出100 mm左右;此外,只有7、8两月原产地区和云南地区的月均降水量达到200 mm以上。
(2)从月均温度因子可以看出,原产地喜马拉雅山南麓地区、海南产区以及云南产区的月均温度变化趋势比较一致,都是在5、6月份达到最高,但数值上,海南更接近于原产地区,而云南地区月均温度比原产地区低5 ℃左右。
2.4 辣木全球潜在分布预测
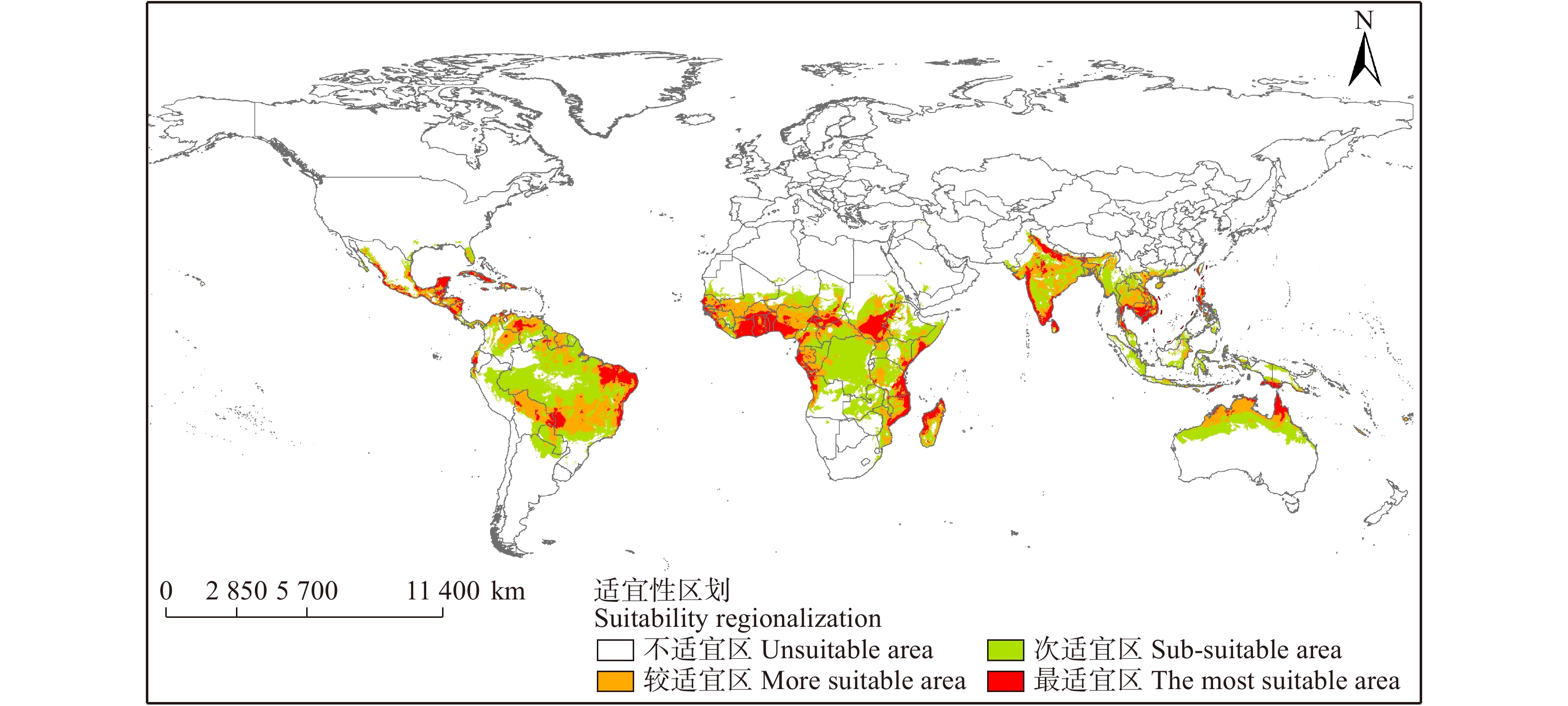
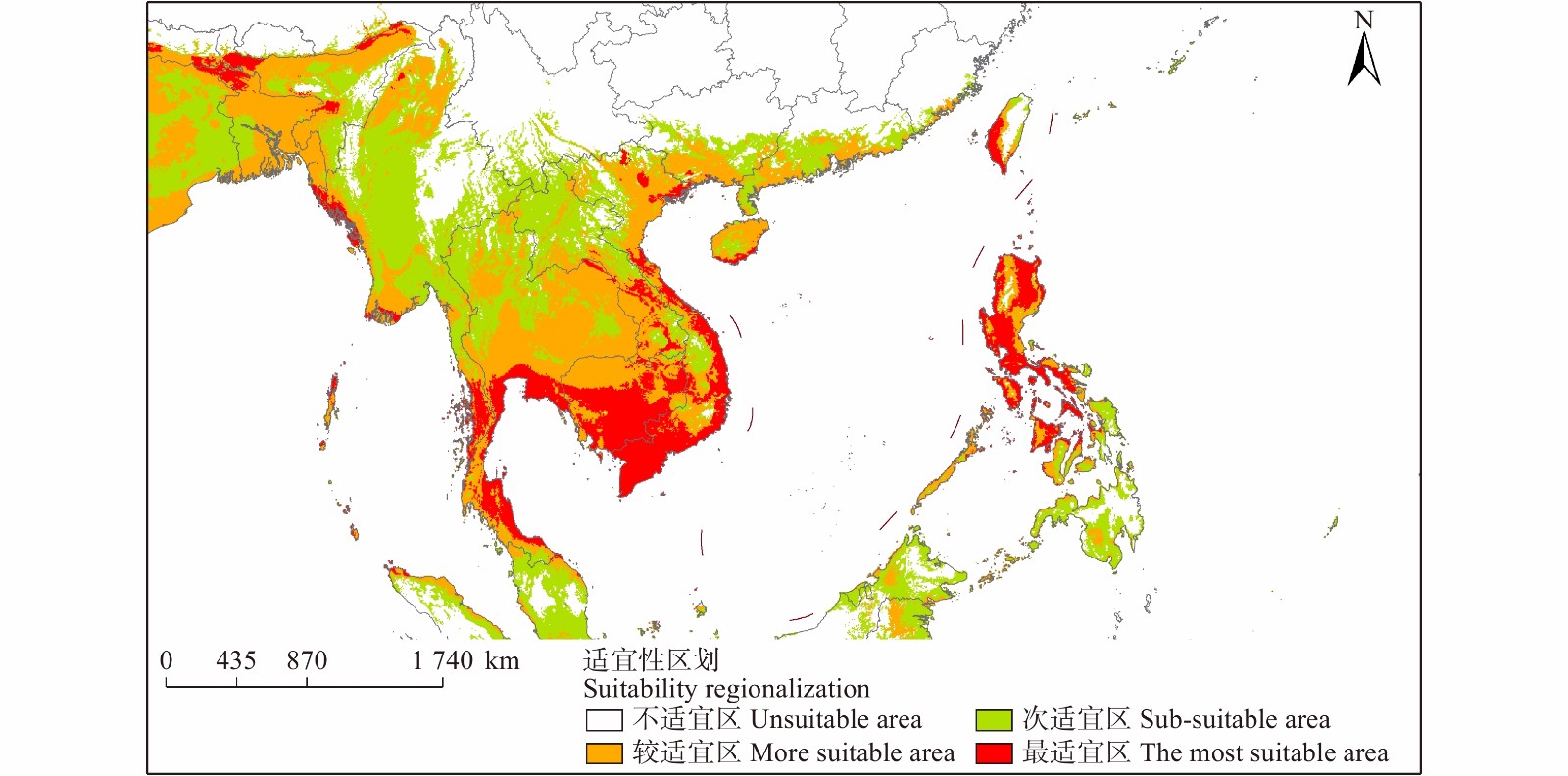
Maxent模型预测结果说明(图7、8),辣木全球潜在适生区域主要分布于南半球S29°至北半球N32°之间的热带、亚热带气候带。在北半球主要集中于南亚、东南亚、非洲西部地区。在南半球主要位于非洲南部、南美洲。
![]() 图 8 辣木中国潜在分布区局部图Figure 8. Partial graph of potential distribution area of M. oleifera in China
图 8 辣木中国潜在分布区局部图Figure 8. Partial graph of potential distribution area of M. oleifera in China亚洲地区最适宜区主要分布在印度、斯里兰卡、菲律宾、越南、柬埔寨;较适宜区主要分布在泰国、老挝、孟加拉国以及中国南部小部分区域;次适宜区主要分布在新加坡、印度尼西亚、文莱等国家。非洲地区最适宜区主要分布在加纳、贝宁、尼日利亚等;较适宜区主要分布在马里、乍得、加蓬等;次适宜区主要分布在刚果、索马里、坦桑尼亚等国家。北美洲地区最适宜区主要分布在古巴、危地马拉等;较适宜区主要分布在尼加拉瓜、洪都拉斯;次适宜区主要分布在美国南端佛罗里达州。南美洲地区最适宜区主要分布在巴西;较适宜区主要分布在委内瑞拉、玻利维亚、哥伦比亚等;次适宜区主要分布在巴西、巴拉圭、秘鲁等国家。大洋洲地区各等级适宜区均分布在在澳大利亚北部。
3. 讨 论
3.1 影响辣木适生区的环境因素
辣木作为一种热带亚热带植物,适宜于热带以及南亚热带地区种植,而不耐寒性是制约其向中亚热带地区引种栽培的最大障碍[17]。多项研究表明,在辣木的整个生长周期内,年平均极端低温、最冷季度最低温、最冷月最低温等低温环境是限制辣木生长的主要因素[18]。此外,有研究人员曾在北方地区种植辣木,裸地种植生长缓慢,不能正常开花结果,冬季需在暖棚才能正常越冬,因此低温是影响辣木在北方地区引种育苗的首要因素[19]。
本文研究结果表明,影响辣木地理分布的环境因素主要是最冷季度平均温(bio11)和最冷月份最低温(bio6),说明低温对辣木空间分布的重要性,与其他学者的研究结果一致。辣木性喜温喜光,耐干旱耐贫瘠,对土壤和降水量要求不严格,因此,在引种栽培时,首先要考量的是上述气候条件。有文献表明,适宜辣木生长的地区一年中的最冷月最低温 ≥ 10 ℃,年平均极端气温 ≥ 0 ℃,并且一年中气温高于10 ℃的时间要达到300 d左右[20]。冬季辣木生长缓慢,并且当温度低于10 ℃时植株会出现冻害甚至死亡[21-23]。
其次,本文研究表明,降水因子作为影响辣木生长的次要因素,在辣木的整个生长发育期间也起着一定的作用。研究结果显示,最湿季度降水量(bio16)适宜范围为409 ~ 3 590 mm,年平均降水量(bio12)适宜范围为833 ~ 2 166 mm。这与相关文献中所记载的辣木所能承受的年均降雨量范围约在500 ~ 3 000 mm[24-26]的记录相吻合。另高温高湿极易导致病虫害频发,持续降雨还会加剧辣木根茎腐烂,因此引种辣木时,还要考察地区的降水情况。
总之,辣木对极端低温的响应最为敏感,其次是降水量。因此,在选择引种栽培区时,温度是辣木在相宜的环境条件下正常生长、开花和结果的基本条件。
3.2 辣木原产地与中国产地气候差异
为比较不同地区与辣木原产地的气候差异,本文基于不同时间尺度的气候因子,从整个物候期角度精细研究了不同地区的气候差异,为不同地区的引种和栽培管理提供数据支撑。
研究结果表明,按照气候因子贡献率大小,最冷季度最低温、最冷月份最低温和最湿季度降雨量这3个综合气候因子是影响辣木地理分布的最主要因子。而云南地区的这3个因子与辣木原产地(喜马拉雅山南麓)基本一致,说明云南地区的温度和降水在季度尺度上都比较适合辣木的生长。同时,云南产区的月均温度变化趋势与辣木原产地也较为相似,都是在5、6月份达到最高。虽然云南地区月均温度比原产地区均低5 ℃左右,但高温持续时间比原产地区要长一个月左右,两地区间月均温度的不同动态变化可能是不同地区辣木品质产生差异的气候原因。同时,从月均降水量比较,云南产区与原产地喜马拉雅山南麓地区的降水趋势比较一致,都是在7、8月份达到最高,但在月均数值上云南稍少于印度产区,云南地区蒸发量少。总而言之,辣木在云南适生区的水热气候条件与原产地的水热气候条件最为相近,但由于云南属于气候区域差异和垂直变化十分明显的高山地带,因此,云南省的潜在适生区面积比较分散,在云南地区要科学选择和比较合适区域进行辣木的生产和栽培。
另外,广东、广西的温度变化方差(bio4)和最湿季度平均温度(bio8)与原产地分布相近,说明这3个地区的温度变异幅度和水热变化基本同步。但在月均降水量方面,广东、广西降水量明显高于原产地,栽培管理上可根据情况注意排涝。
3.3 辣木全球潜在适生区格局
3.3.1 辣木的全球潜在适生区
辣木原产于喜马拉雅山脉南麓以下的印度、尼泊尔、孟加拉国部分地区。现广泛种植于全球的热带及亚热带地区。模型预测结果显示辣木的全球潜在适生区集中在亚洲南部,非洲中部以及南美洲北部地区。
位于亚洲的印度作为目前最大的辣木产品出口国,其辣木产量位居世界前列,模型预测结果也显示印度整个国家基本都属于辣木潜在适生区。印度位于南亚季风区,全境大部分属于热带季风气候,终年高温,降水集中,来自印度洋含有大量水汽的海风导致此时期降水量大。冬季受喜马拉雅山脉屏障影响,无寒流或冷高压南下的影响,故辣木能正常越冬生长。
模型预测结果显示,非洲地区是辣木适生区覆盖最多的地区,其中最适生区主要分布在西非。非洲西部干热,沿海多雨,气候和植被有明显的纬度地带性。该地辣木适生区的主要气候类型为热带草原气候和热带雨林气候。几内亚、贝宁、加纳等适生区面对几内亚湾,洋流向北,因此终年潮湿,气温也比同纬度的其他西非国家高,季节变化明显,降水量很大。
另外,预测结果还显示在北美洲的潜在适生区集中在危地马拉、古巴等地。南美洲的潜在适生区主要在巴西东部以及委内瑞拉西北部。这些国家皆地处热带,潜在适生区大部分为热带雨林气候,终年高温多雨。大洋洲的澳大利亚北部也存在部分辣木潜在适生区。澳大利亚跨两个气候带,北部靠近赤道,属于热带,属热带草原气候和热带雨林气候。
3.3.2 辣木的中国分布
由于喜马拉雅山、帕米尔高原等的阻隔,中国并没有辣木的自然分布,现有的辣木资源均是从国外引进的。我国主要种植在广东、广西、海南、云南等地。模型预测结果显示,辣木在我国的适生分布区集中在我国南部的海南、广西、广东部分区域,云南也有小部分适生区存在。
植物引种的成功率受很多因素影响,但区系地理因素和气候因素起着重要作用[27]。有文献表明,云南是我国辣木种植面积最大省,适生区分布地处低纬度地带,受北部湾东南暖湿气流和孟加拉湾西南暖湿气流的影响大,高温多雨,属典型的南亚热带气候类型。气候区域差异十分明显,热带、亚热带区域分为湿热区和干热区两类[28],北部有高山阻挡寒潮南下,导致冬季气温比中国同纬度其他地方高,故辣木能够正常越冬。海南岛全境都是辣木适宜引种区,海南地处热带北缘,属热带季风气候,终年温暖,年平均气温约22 ~ 27 ℃,即使是最冷月份的温度也能达到17 ~ 24 ℃,雨量充沛,年降水量约1 000 ~ 2 600 mm。广西位于我国南部,属于亚热带季风气候,该地区拥有充足的光照,降雨量大,与辣木适宜生长环境的气候吻合,年平均气温21.6 ℃,自然条件有利于辣木生长[29]。广东地区则是华夏植物区系的重要组成部分[30-31],其邻近地区中印半岛、马来西亚、喜马拉雅与印度北部都是华夏植物区系的后裔,两者具有相近的植物区系,而辣木也恰好源于印度北部的喜马拉雅山南麓地区。这可能是辣木作为热带季风气候植物也较为适宜在广州地区种植的原因。目前,辣木在我国的引种地主要分布在广西南宁、百色,云南景洪,广东、海南等长江以南各省区,与文中预测分析的辣木在中国的潜在适生区分布结果较为一致,但是由于本文所选环境变量种类并不全面,以及模型拟合过程中的误差,本研究预测结果可能和辣木实际生长地区有一些差异,还需后续进一步考察分析。
-
![]()
图 3 不同环境变量训练得分的Jackknife检验
Figure 3. Jackknife test on training score of different environmental variables
![]()
图 5 辣木原产地和中国分布区主要综合气候因子
YC. 原产地区;GD. 广东;GX. 广西;HN. 海南;TW. 台湾;YN. 云南。下同。YC, original area; GD, Guangdong; GX, Guangxi; HN, Hainan; TW, Taiwan; YN, Yunnan. The same below.
Figure 5. Main comprehensive climatic factors of M. oleifera in original area and the distributing area in China
![]()
图 6 辣木原产区和中国分布区月均气候因子
Figure 6. Monthly mean climatic factors of M. oleifera in original area and the distributing area in China
![]()
图 8 辣木中国潜在分布区局部图
Figure 8. Partial graph of potential distribution area of M. oleifera in China
表 1 环境变量
Table 1 Environmental variables
反应指标 Response indicator 环境变量 Environmental variable 平均温度及其变异幅度
Average temperature and its variation range温度变化方差
SD of temperature seasonality (bio4)极端温度影响
Extreme temperature effect最冷月份最低温度、最暖季度平均温度、最冷季度平均温度
Min. temperature of the coldest month (bio6), mean temperature of the warmest quarter (bio10), mean temperature of the coldest quarter (bio11)降雨量及其季节性分布
Rainfall and its seasonal distribution雨量变化方差
Variance of rainfall variation (bio15)极端水分条件
Extreme moisture condition最湿季度降水量、最干季度降水量
Precipitation of the wettest quarter (bio16), precipitation of the driest quarter (bio17)水热是否同步
Hydrothermal synchronization or not最湿季度平均温度、最暖季度平均降水量、最冷季度平均降水量
Mean temperature of the wettest quarter (bio8), mean precipitation of the warmest quarter (bio18), mean precipitation of the coldest quarter (bio19)温差特点
Temperature difference characteristics昼夜温差月均值
Monthly mean temperature difference between day and night (bio2)月均变化
Monthly average change1—12月月均温度,1—12月月均降水量
Average monthly temperature and monthly average precipitation from January to December 下载: 导出CSV
下载: 导出CSV
-
[1] 肖正春, 张广伦. 神奇的辣木与人类生活[J]. 中国野生植物资源, 2016, 35(1):64−66. Xiao Z C, Zhang G L. The magic moringa and human life[J]. Chinese Wild Plant Resources, 2016, 35(1): 64−66.
[2] 沙子珺, 刘英华, 杨滨, 等. 辣木传统应用的文献考证[J]. 中国中药杂志, 2020, 45(12):2800−2807. Sha Z J, Liu Y H, Yang B, et al. Textual research on traditional application of Moringa[J]. China Journal of Chinese Materia Medica, 2020, 45(12): 2800−2807.
[3] Paikra B K, Dhongade H K J, Gidwani B. Phytochemistry and pharmacology of Moringa oleifera Lam.[J]. Journal of Pharmacopuncture, 2017, 20(3): 194−200. doi: 10.3831/KPI.2017.20.022
[4] Olurishe C, Kwanashie H, Zezi A, et al. Chronic administration of ethanol leaf extract of Moringa oleifera Lam. (Moringaceae) may compromise glycaemic efficacy of sitagliptin with no significant effect in retinopathy in a diabetic rat model[J]. Journal of Ethnopharmacology, 2016, 194: 895−903. doi: 10.1016/j.jep.2016.10.065
[5] 吴頔, 蔡志华, 魏烨昕, 等. 辣木作为新型植物性蛋白质饲料的研究进展[J]. 动物营养学报, 2013, 25(3):63−71. Wu D, Cai Z H, Wei Y X, et al. Research progress of Moringa oleifera Lam. as a new plant protein feed[J]. Chinese Journal of Animal Nutrition, 2013, 25(3): 63−71.
[6] 白旭华, 黎小清, 伍英. 辣木天然絮凝剂提取工艺研究初报[J]. 热带农业科技, 2013, 36(3):26−31. Bai X H, Li X Q, Wu Y. Preliminary report on extraction technology of bio-flocculant from Moringa oleifera[J]. Topical Agricultural Science&Technology, 2013, 36(3): 26−31.
[7] 段琼芬, 马李一, 余建兴, 等. 辣木油抗紫外线性能研究[J]. 食品科学, 2008, 29(9):118−121. Duan Q F, Ma L Y, Yu J X, et al. Study on anti-ultraviolet properties of Moringa oleifera Oil[J]. Food Science, 2008, 29(9): 118−121.
[8] 罗会英, 韩学琴, 邓红山, 等. 不同种源辣木生长差异性分析[J]. 热带农业科学, 2019, 39(3):16−20. Luo H Y, Han X Q, Deng H S, et al. Growth difference of Moringa oleifera Lam. from different provenances[J]. Chinese Journal of Tropical Agriculture, 2019, 39(3): 16−20.
[9] 李东, 赵一鹤. 辣木研究现状及发展趋势[J]. 西部林业科学, 2011, 47(5):32−38. Li D, Zhao Y H. Research status and development trend of Moringa[J]. Journal of West China Forestry Science, 2011, 47(5): 32−38.
[10] Muhl Q E, Toit E S D, Robbertse P J. Moringa oleifera (Horseradish tree) leaf adaptation to temperature regimes[J]. International Journal of Agriculture & Biology, 2011, 13(6): 1021−1024.
[11] Silva N, Mendes-Bonato A B, Sales J G C, et al. Meiotic behavior and pollen viability in Moringa oleifera (Moringaceae) cultivated in southern Brazil[J]. Genetics Molecular Research, 2011, 10(3): 1728−1732. doi: 10.4238/vol10-3gmr1490
[12] 苏瑶, 赵一鹤, 冯武, 等. 云南引种辣木籽营养成分分析与评价[J]. 西部林业科学, 2015, 44(4):142−145, 149. Su Y, Zhao Y H, Feng W, et al. Analysis and evaluation on nutritional components of Moringa oleifera in Yunnan[J]. Journal of West China Forestry Science, 2015, 44(4): 142−145, 149.
[13] 曾明莹, 傅咏梅, 张蜀, 等. 不同产地辣木叶提取物的含量测定及指纹图谱相似性评价[J]. 云南农业大学学报(自然科学), 2018, 33(3):485−493. Zeng M Y, Fu Y M, Zhang S, et al. Simultaneous determination of 4 flavonoids and chlorogenic acid of extract from Moringa oleiferam Lam. leaves and its UPLC fingerprint analysis[J]. Journal of Yunnan Agricultural University (Natural Science), 2018, 33(3): 485−493.
[14] 杜丽娟, 严红梅, 王丽, 等. 云南省不同地区辣木各器官中总硒及硒形态含量的比较研究[J]. 食品安全质量检测学报, 2018, 9(18):4929−4934. Du L J, Yan H M, Wang L, et al. Comparison of total selenium and selenium forms content in moringa organs in different regions of Yunnan Province[J]. Journal of Food Safety and Quality, 2018, 9(18): 4929−4934.
[15] 张琴, 张东方, 吴明丽, 等. 基于生态位模型预测天麻全球潜在适生区[J]. 植物生态学报, 2017, 41(7):770−778. doi: 10.17521/cjpe.2016.0380 Zhang Q, Zhang D F, Wu M L, et al. Predicting the global areas for potential distribution of Gastrodia elata based on ecological niche models[J]. Chinese Journal of Plant Ecology, 2017, 41(7): 770−778. doi: 10.17521/cjpe.2016.0380
[16] 谢彩香, 张琴, 白光宇. 木本能源植物文冠果的生态特征及区划[J]. 植物科学学报, 2018, 36(2):229−236. Xie C X, Zhang Q, Bai G Y. Ecological characteristics and regionalization of Xanthoceras sorbifolia Bunge, a woody energy plant[J]. Plant Science Journal, 2018, 36(2): 229−236.
[17] 林宗铿, 张天翔, 杨俊杰. 应用Logistic方程确定辣木的抗寒性[J]. 福建农业学报, 2018, 33(5):512−515. Lin Z K, Zhang T X, Yang J J. Low-temperature tolerance of Moringa spp. determined by a Logistic model[J]. Fujian Journal of Agricultural Science, 2018, 33(5): 512−515.
[18] Thurber M, Fahey J. Adoption of Moringa oleifera to combat under-nutrition viewed through the lens of the “Diffusion of Innovations” theory[J]. Ecology of Food & Nutrition, 2009, 48(3): 212−225.
[19] 王书瑞, 魏殿文, 姚琴, 等. 北方地区辣木育苗影响因子分析[J]. 国土与自然资源研究, 2017(6):65−68. Wang S R, Wei D W, Yao Q, et al. Analysis of seedling environmental factors on Moringa oleifera in the North of China[J]. Territory & Natural Resource Study, 2017(6): 65−68.
[20] 周利平, 李永, 姜绍红, 等. 普洱市辣木引种生境适应性初步研究[J]. 林业调查规划, 2015, 40(5):147−152. Zhou L P, Li Y, Jiang S H, et al. A preliminary study on the eco-adaptability of the introduction of Moringa oleifera in Pu’er City[J]. Forest Inventory and Planning, 2015, 40(5): 147−152.
[21] 彭兴民, 郑益兴, 段琼芬, 等. 印度传统辣木引种栽培研究[J]. 热带亚热带植物学报, 2008, 16(6):579−585. Peng X M, Zheng Y X, Duan Q F, et al. Studies on introduction and cultivation of Moringa oliferia[J]. Journal of Tropical and Subtropical Botany, 2008, 16(6): 579−585.
[22] 盘李军, 刘小金. 辣木的栽培及开发利用研究进展[J]. 广东林业科技, 2010, 26(3):71−77. Pan L J, Liu X J. Research overview on cultivation and utilization of Moringa oleifera Lam.[J]. Guangdong Forestry Science and Technology, 2010, 26(3): 71−77.
[23] 庄文晶, 祝秋萍, 陈福梓, 等. 辣木寒冻害指标初探[C]// 第35届中国气象学会年会S6应对气候变化、低碳发展与生态文明建设. 安徽: 中国气象学会, 2018: 622−625. Zhuang W J, Zhu Q P, Chen F Z, et al. Preliminary study on freezing damage index of Moringa oleifera Lam.[C]// Proceedings of the 35th Annual Meeting of Chinese Meteorological Society S6 on Climate Change, Low-Carbon Development and Ecological Civilization Construction. Anhui: Chinese Meteorological Society, 2018: 622−625.
[24] 陈江平. 辣木的种植技术[J]. 农业与技术, 2017, 37(3):73−74. Chen J P. Cultivation technology of Moringa oleifera Lam.[J]. Agriculture and Technology, 2017, 37(3): 73−74.
[25] 邓红山, 金杰, 罗会英, 等. 辣木在干热河谷地区不同品种种子质量表现[J]. 中国热带农业, 2019(4):46−48. Deng H S, Jin J, Luo H Y, et al. Seed quality performance of Moringa oleifera in different varieties in hot and dry area[J]. China Tropical Agriculture, 2019(4): 46−48.
[26] Leone A, Spada A, Battezzati A, et al. Cultivation, genetic, ethnopharmacology, phytochemistry and pharmacology of Moringa oleifera leaves: an overview[J]. International Journal of Molecular Sciences, 2015, 16(6): 12791−12835.
[27] Woodward F I. Climate and plant distribution[M]. Cambridge: Cambridge University Press, 1987: 62−107.
[28] 廖承飞, 李贵华, 韩学琴, 等. 云南辣木产业发展的SWOT分析及对策[J]. 中国热带农业, 2016(2):13−16. Liao C F, Li G H, Han X Q, et al. SWOT analysis and countermeasures of Moringa oleifera Lam. industry development in Yunnan[J]. Industry Development, 2016(2): 13−16.
[29] 梁潘霞, 刘永贤, 沙国新, 等. 广西辣木产业发展现状及富硒辣木发展前景展望[J]. 热带农业科学, 2017, 37(8):88−92. Liang P X, Liu Y X, Sha G X, et al. Development status and prospect of selenium-rich Moringa in Guangxi[J]. Chinese Journal of Tropical Agriculture, 2017, 37(8): 88−92.
[30] 张宏达. 广东植物区系的特点[J]. 中山大学学报(自然科学版), 1962(1):1−34. Zhang H D. Tropical flora of Guangdong[J]. Acta Scientiarum Naturalium Universitatis Sunyatseni (Natural Science), 1962(1): 1−34.
[31] 李许文, 刘文, 陈红锋, 等. 广州适宜的植物引种来源地与气候区选择研究[J]. 中国园林, 2016, 32(4):96−100. Li X W, Liu W, Chen H F, et al. A study on the selection of appropriate source regions and climate zones for Guangzhou to introduce plants[J]. Chinese Landscape Architecture, 2016, 32(4): 96−100.
-
期刊类型引用(3)
1. 刘索名,王君杰,燕云飞,姜立春. 小兴安岭兴安落叶松人工林冠幅模型构建. 北京林业大学学报. 2023(05): 79-87 .  本站查看
本站查看
2. 李应涛,刘时良,孙海龙,王卫霞,向玮. 云冷杉针阔混交林单木枝下高和冠幅模型构建. 森林与环境学报. 2022(03): 289-296 . 百度学术
3. 佟艺玟,陈东升,冯健,高慧淋. 基于线性分位数混合效应的辽东山区红松冠幅模型. 应用生态学报. 2022(09): 2321-2330 . 百度学术
其他类型引用(1)


计量
- 文章访问数: 1504
- HTML全文浏览量: 533
- PDF下载量: 88
- 被引次数: 4
