
Leaf water absorption and canopy rainfall interception of twenty-one plant species in Beijing
-
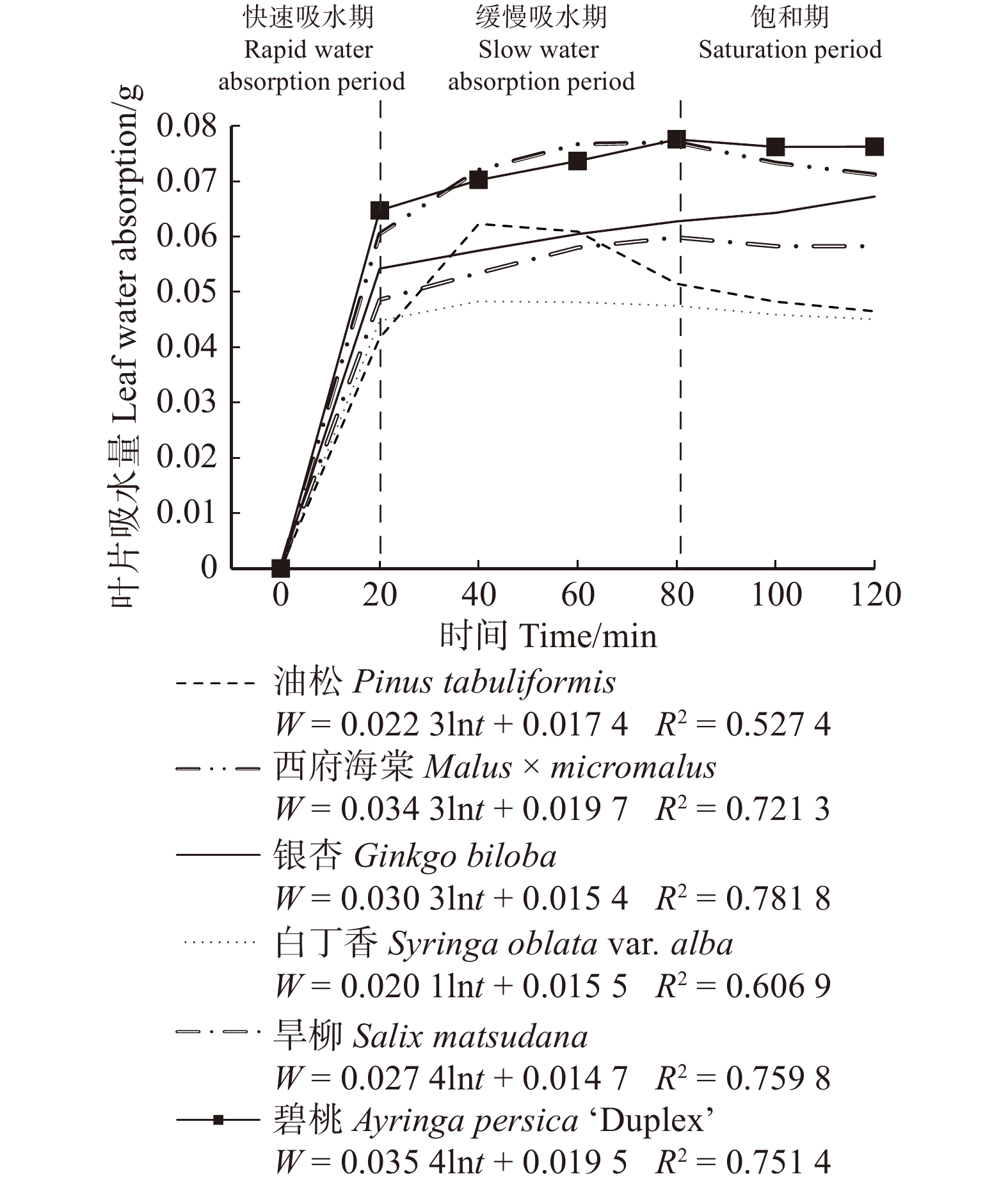
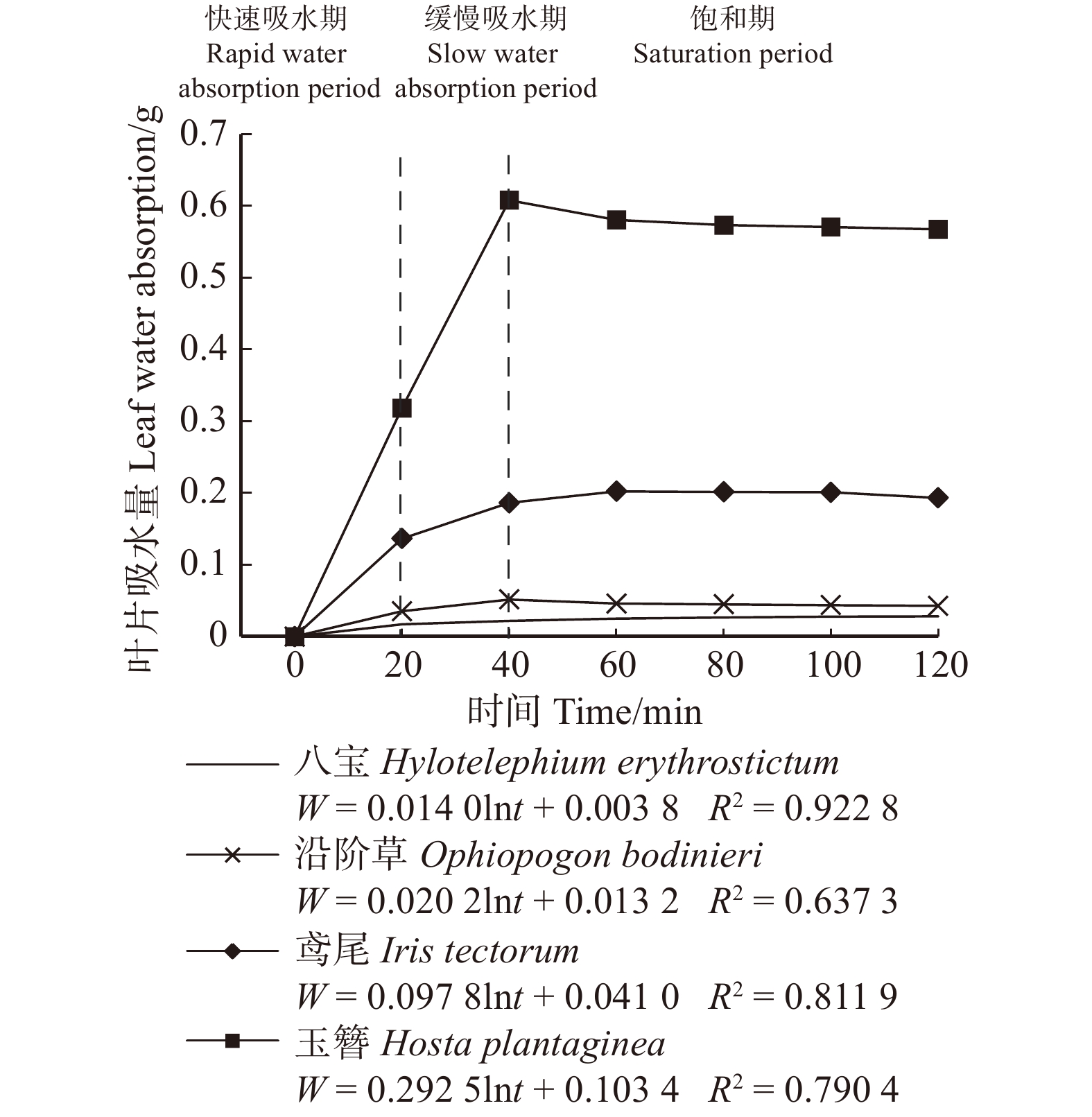
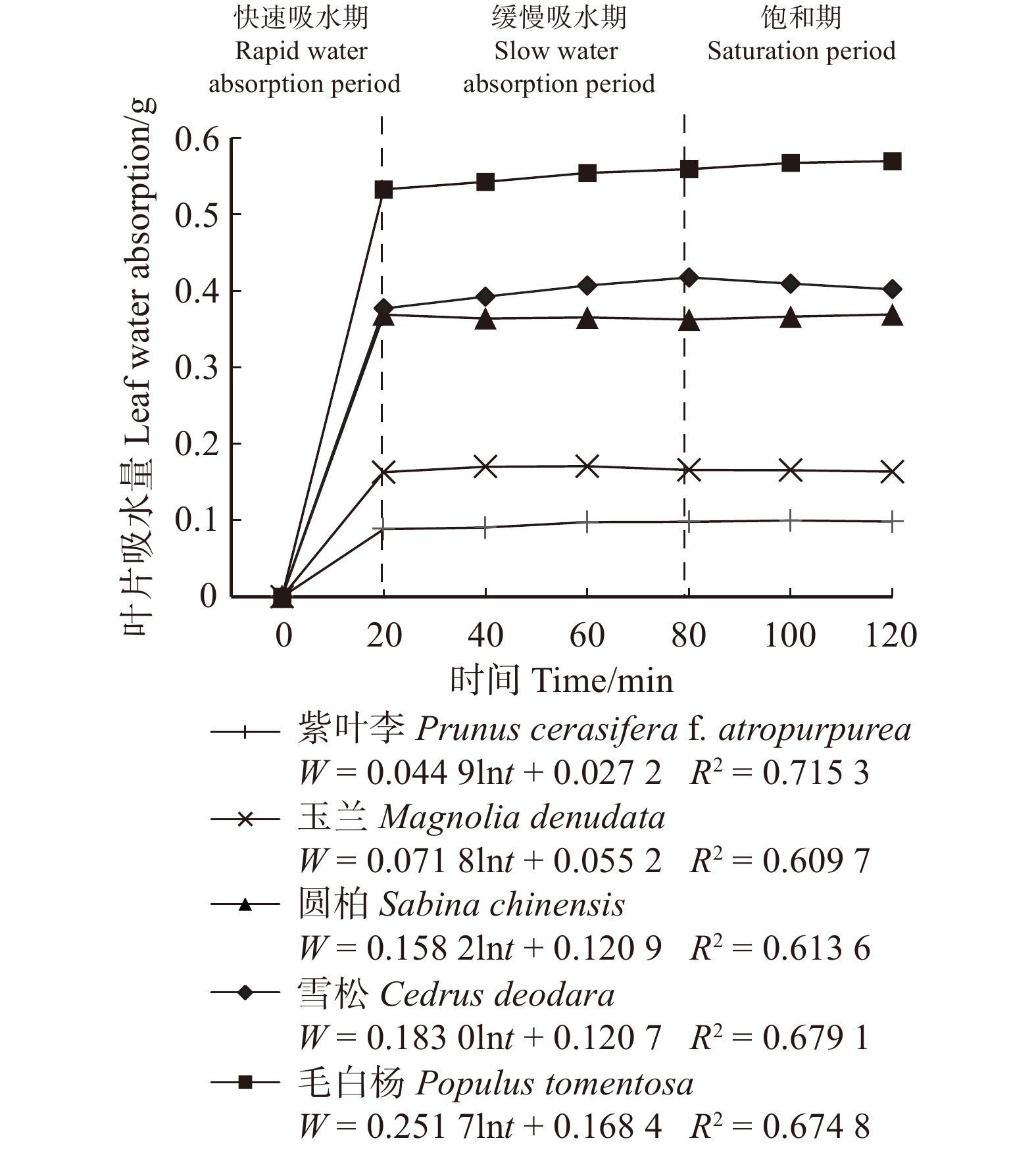
摘要:目的 植物冠层截留雨水对削减雨水径流起到重要作用,而植物叶片吸水性能与植物冠层截留雨水的能力息息相关。方法 为了评估植物冠层对雨水的截留能力,通过浸水实验、实地测量、航片分析等方法,对北京市21种不同类型植物叶片吸水量、冠层截留量和冠层截留体积进行计算。结果 研究表明:(1)吸水量与浸水时间呈对数函数关系;(2)植物叶片吸水过程可划分为3个阶段,乔木和灌木的叶片吸水过程类似,20 min时叶片吸水量均值分别为0.17 g和0.05 g,80 min时叶片吸水量均值分别为0.18 g和0.06 g,120 min时叶片吸水量均值分别为0.18 g和0.06 g;草本植物叶片20 min时叶片吸水量均值为0.13 g,40 min时叶片吸水量均值为0.27 g,120 min时叶片吸水量均值为0.21 g;(3)11种乔木的冠层雨水截留量在71.30 ~ 738.72 g/m2之间,6种灌木的冠层雨水截留量在41.79 ~ 275.28 g/m2之间,4种草本的冠层雨水截留量在57.82 ~ 217.49 g/m2之间,常绿针叶植物冠层截留量明显高于落叶阔叶植物;(4)部分灌木和草本植物冠层截留量比部分乔木冠层截留量高,如黄杨、鸢尾;(5)植物冠层截留体积与冠层覆盖面积、植物冠层截留量呈正相关关系,常绿针叶乔木+常绿灌木+草本搭配组合的冠层截留体积最大;(6)雪松+黄杨+鸢尾群落冠层的多年平均降雨截留量占总降雨量的比例为11.57%,对小降雨事件有显著截留效果。结论 不同植物种类及配置对城市绿地雨水截留能力有影响,本研究可为构建高截留能力植物群落及海绵城市绿地建设提供参考。Abstract:Objective The rainfall interception by plant canopy plays an important role in reducing stormwater runoff, and the water absorption performance of plant leaves is closely related to the ability of plant canopy to intercept rainfall.Method In order to evaluate the interception ability of plant canopy, the leaf water absorption, canopy interception quantity and canopy interception volume of 21 plant species in Beijing were calculated by water immersion experiment, field measurement and aerial image analysis.Result Research showed that: (1) the relationship between water absorption and immersion time was logarithmic; (2) the water absorption process of plant leaves can be divided into three stages. The water absorption process of trees and shrubs was similar. The mean water absorption of leaves at 20 min was 0.17 and 0.05 g, respectively; the mean water absorption of leaves at 80 min was 0.18 and 0.06 g, respectively; and the mean water absorption of leaves at 120 min was 0.18 and 0.06 g, respectively. The average water absorption of herb leaves was 0.13 g at 20 min, 0.27 g at 40 min, and 0.21 g at 120 min; (3) the canopy rainfall interception of 11 arbor species was between 71.30 and 738.72 g/m2, and the canopy rainfall interception of 6 shrub species was 41.79−275.28 g/m2, the canopy rainfall interception of 4 herb species ranged from 57.82 to 217.49 g/m2, and the canopy interception of evergreen conifers was significantly higher than that of deciduous broadleaved plants; (4) canopy interception of some shrubs and herbs was higher than that of some arbors, such as Buxus sinica and Iris tectorum;
(5) the plant canopy interception volume was positively correlated with canopy cover area and plant canopy interception quantity, the evergreen coniferous tree + evergreen shrub + herb combination had the largest canopy interception volume; (6) the ratio of average annual rainfall interception of the Cedrus deodara +Buxus sinica + Iris tectorum community canopy to the total rainfall was 11.57%, which had a significant interception effect on small rainfall events. Conclusion Different plant types and configurations have an impact on urban green space rainfall interception capacity. This study can provide reference for the construction of high retention capacity plant communities and sponge city green space construction.-
Keywords:
- immersion method /
- leaf water absorption /
- canopy interception /
- leaf area index /
- Beijing City
-
-
![]()
图 5 21种植物冠层截留量排序
Figure 5. Sequence of canopy interception quantity of 21 plant species
![]()
图 6 13种植物冠层截留体积排序
Figure 6. Sequence of canopy interception volume of 13 plant species
![]()
图 7 10组典型植物群落冠层截留体积
Figure 7. Canopy interception volume of 10 typical plant communities
![]()
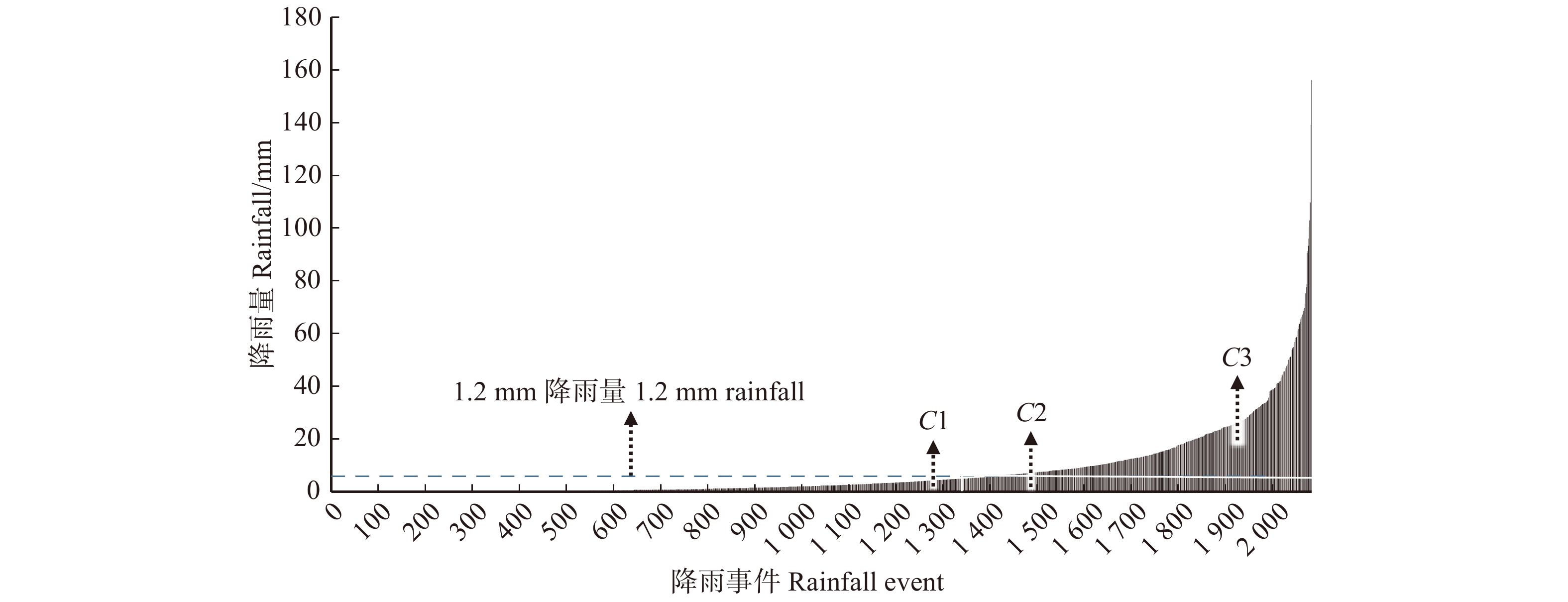
图 8 北京市降雨事件与降雨量关系曲线示意图(1982—2011年)
Figure 8. Schematic diagram of the relationship between rainfall events and rainfall in Beijing (1982−2011)
![]()
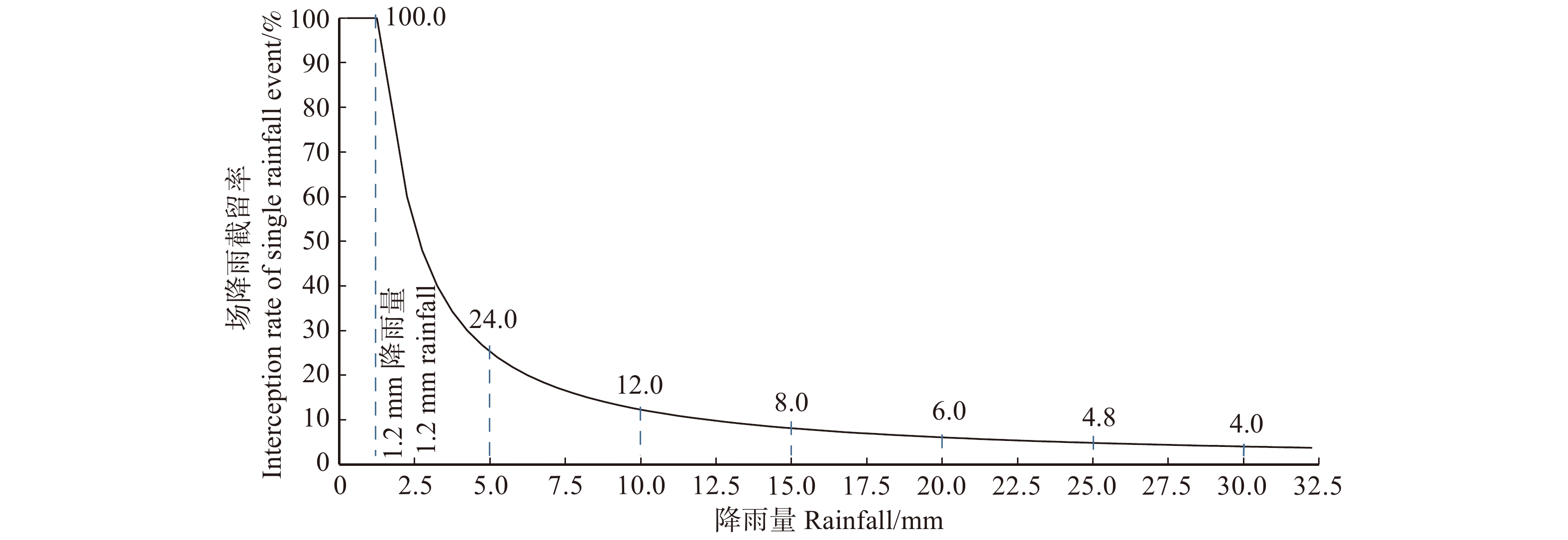
图 9 植物群落的场降雨截留率与降雨量的关系示意图
Figure 9. Schematic diagram of the relationship between interception rate of single rainfall event and rainfall
![]()
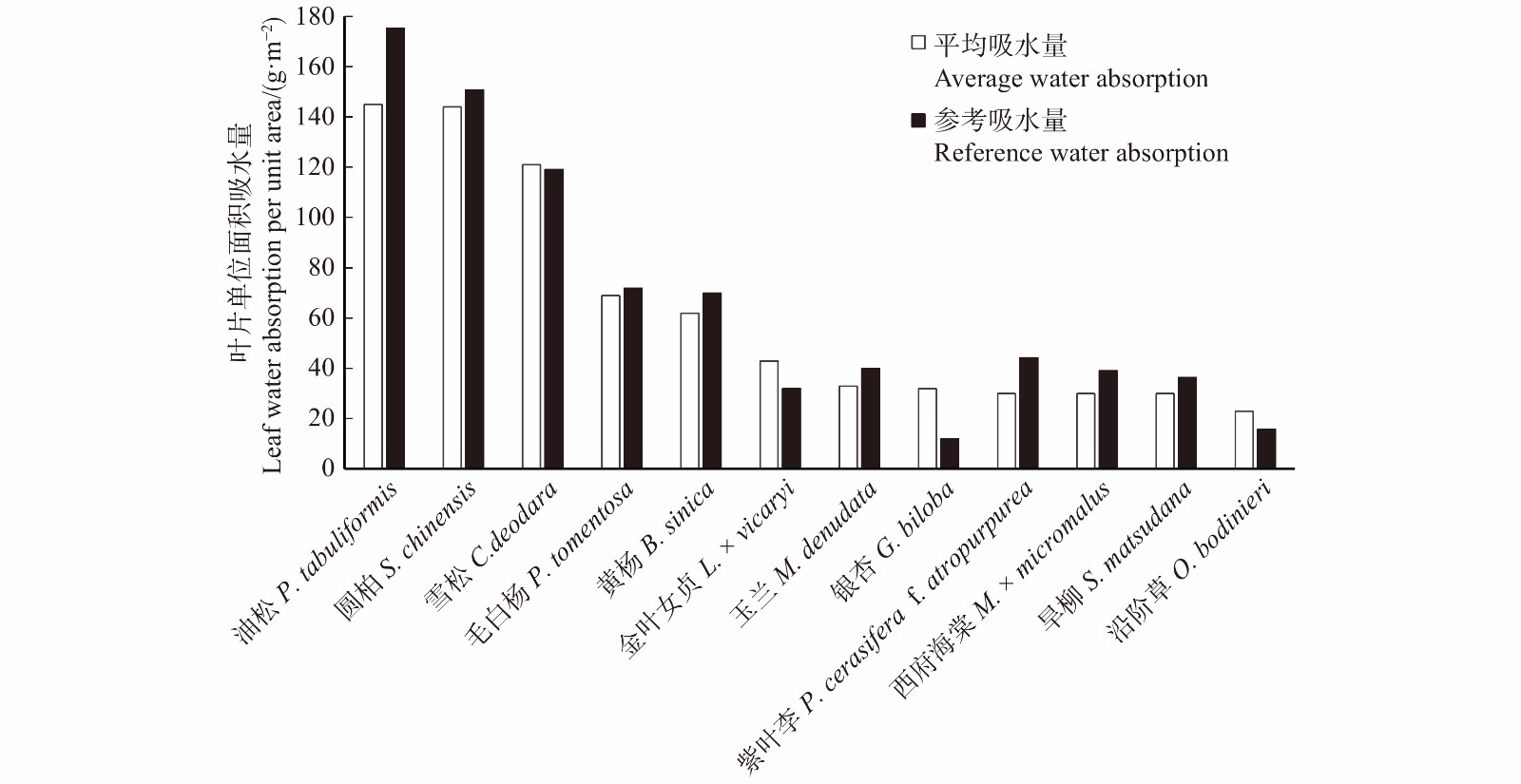
图 10 本研究叶片单位面积吸水量与相关研究比较
Figure 10. Leaf water absorption per unit area compared with the related study
表 1 试验区基础数据
Table 1 Basic data in experimental area
研究区域
Experimental area绿地性质
Classification of green space建成年代
Year of completion占地面积
Site area/m2建筑面积
Construction area/m2绿化覆盖率
Green coverage/%乔木冠层
覆盖面积
Arbor canopy
cover area/m2灌木冠层
覆盖面积
Shrub canopy
cover area/m2草本冠层
覆盖面积
Herb canopy
cover area/m2恭王府
Prince Kung’s Mansion公园绿地
Public park1785 61 120 32 260 46 23 054 5 060 6 466 中央音乐学院
Central Conservatory of Music附属绿地
Attached green space1940 54 446 87 485 23 9 767 3 130 3 882 北京口腔医院
Beijing Stomatological Hospital附属绿地
Attached green space1945 23 000 28 659 20 3 680 874 1 150 前门大街
Qianmen Street附属绿地
Attached green space1550 22 300 − 22 4 464 294 147 菊儿胡同
Juer Hutong附属绿地
Attached green space1985 14 600 18 666 24 3 188 280 70 荣宁园小区
Rongningyuan Community附属绿地
Attached green space2001 18 500 80 300 15 1 942 888 1 137 注:− 代表无建筑面积。Note: − indicates no construction area.  下载: 导出CSV
下载: 导出CSV
表 2 21种植物冠层截留量
Table 2 Canopy interception of 21 plant species
类型
Class类别
Type种名
Species name叶面积指数
Leaf area index叶片单位面积
吸水量
Leaf water absorption
per unit area/(g·m−2)单株植物冠层
覆盖面积
Individual plant
canopy coverage/m2植物冠层
截留量
Plant canopy interception/(g·m−2)乔木 Arbor 常绿针叶
Evergreen needle圆柏
Sabina chinensis5.13 144 20.25 738.72 雪松
Cedrus deodara5.84 121 58.50 706.64 油松
Pinus tabuliformis4.72 145 64.00 684.40 落叶阔叶
Deciduous broadleaf毛白杨
Populus tomentosa3.77 69 45.50 260.13 玉兰
Magnolia denudata4.55 33 19.78 150.15 银杏
Ginkgo biloba3.50 32 63.08 112.00 碧桃
Amygdalus persica ‘Duplex’3.16 34 22.50 107.44 西府海棠
Malus × micromalus3.50 30 28.08 105.00 白丁香
Syringa oblata var. alba2.78 31 55.76 86.18 紫叶李
Prunus cerasifera f. atropurpurea2.50 30 21.20 75.00 旱柳
Salix matsudana3.10 23 48.28 71.30 灌木 Shrub 常绿
Evergreen黄杨
Buxus sinica4.44 62 − 275.28 金叶女贞
Ligustrum × vicaryi4.16 43 − 178.88 落叶
Deciduous连翘
Forsythia suspensa1.90 48 8.40 91.20 榆叶梅
Amygdalus triloba2.10 36 9.80 75.60 月季
Rosa chinensis2.57 27 − 69.39 紫叶小檗
Berberis thunbergii ‘Atropurpurea’1.99 21 − 41.79 草本 Herb 鸢尾
Iris tectorum5.83 37 − 217.49 玉簪
Hosta plantaginea3.35 42 − 142.80 沿阶草
Ophiopogon bodinieri4.76 30 − 139.92 八宝
Hylotelephium erythrostictum4.13 14 − 57.82 注:− 表示植物常为丛生或者簇生种植模式,不进行单株冠幅面积测定。Notes: − indicates that plants are often planted in clusters, and the canopy area of a single plant is not measured.
下载: 导出CSV
-
[1] 王佳, 王思思, 车伍, 等. 雨水花园植物的选择与设计[J]. 北方园艺, 2012(19):77−81. Wang J, Wang S S, Che W, et al. Plant selection and design of rain garden[J]. Northern Horticulture, 2012(19): 77−81.
[2] 李俊清. 森林生态学[M]. 北京: 高等教育出版社, 2006: 100−103. Li J Q. Forest ecology[M]. Beijing: Higher Education Press, 2006: 100−103.
[3] 李晶晶, 白岗栓, 张蕊. 陕北丘陵沟壑区常见树种叶片的吸水性能[J]. 中国水土保持科学, 2013, 11(1):99−102. Li J J, Bai G S, Zhang R. Water absorption of common trees leaves in loess hilly and gully region of Northern Shaanxi[J]. Science of Soil and Water Conservation, 2013, 11(1): 99−102.
[4] Huang J Y, Black T A, Jassal R S, et al. Modelling rainfall interception by urban trees[J]. Canadian Water Resources Journal / Revue canadienne des ressources hydriques, 2017, 42(4): 336−348. doi: 10.1080/07011784.2017.1375865
[5] Toba T, Ohta T. Factors affecting rainfall interception determined by a forest simulator and numerical model[J]. Hydrological Processes, 2008, 22(14): 2634−2643. doi: 10.1002/hyp.6859
[6] 郭胜男, 林萍, 吴荣, 等. 昆明市园林植物树冠截留降雨及其影响因素研究[J]. 广东农业科学, 2014, 41(23):47−51. Guo S N, Lin P, Wu R, et al. Study on canopy interception rainfall of garden plant in Kunming and its influencing factors[J]. Guangdong Agricultural Sciences, 2014, 41(23): 47−51.
[7] 李想, 王亚明, 孟晨, 等. 基于幼树模拟降雨实验的树冠动态截留模型[J]. 北京林业大学学报, 2018, 40(4):43−50. Li X, Wang Y M, Meng C, et al. A dynamic crown interception model based on simulated rainfall experiments of small trees[J]. Journal of Beijing Forestry University, 2018, 40(4): 43−50.
[8] Návar J. Fitting rainfall interception models to forest ecosystems of Mexico[J]. Journal of Hydrology, 2017, 548: 458−470. doi: 10.1016/j.jhydrol.2017.03.025
[9] Linhoss A C, Siegert C M. A comparison of five forest interception models using global sensitivity and uncertainty analysis[J]. Journal of Hydrology, 2016, 538: 109−116. doi: 10.1016/j.jhydrol.2016.04.011
[10] Návar J. Modeling rainfall interception components of forests: extending drip equations[J]. Agricultural and Forest Meteorology, 2019, 279: 107704. doi: 10.1016/j.agrformet.2019.107704
[11] Junqueira J J A, De Mello C R, De Mello J M, et al. Rainfall partitioning measurement and rainfall interception modelling in a tropical semi-deciduous Atlantic forest remnant[J]. Agricultural and Forest Meteorology, 2019, 275: 170−183. doi: 10.1016/j.agrformet.2019.05.016
[12] Fan J L, Oestergaard K T, Guyot A, et al. Measuring and modeling rainfall interception losses by a native Banksia woodland and an exotic pine plantation in subtropical coastal Australia[J]. Journal of Hydrology, 2014, 515: 156−165. doi: 10.1016/j.jhydrol.2014.04.066
[13] Fathizadeh O, Hosseini S M, Zimmermann A, et al. Estimating linkages between forest structural variables and rainfall interception parameters in semi-arid deciduous oak forest stands[J]. Science of the Total Environment, 2017, 601−602: 1824−1837. doi: 10.1016/j.scitotenv.2017.05.233
[14] Stringham T K, Snyder K A, Snyder D K, et al. Rainfall interception by single leaf Piñon and Utah Juniper: implications for stand-level effective precipitation[J]. Rangeland Ecology & Management, 2018, 71(3): 327−335.
[15] Ghimire C P, Bruijnzeel L A, Lubczynski M W, et al. Measurement and modeling of rainfall interception by two differently aged secondary forests in upland eastern Madagascar[J]. Journal of Hydrology, 2017, 545: 212−225. doi: 10.1016/j.jhydrol.2016.10.032
[16] Zabret K, Šraj M. Rainfall interception by urban trees and their impact on potential surface runoff[J]. Clean: Soil, Air, Water, 2019, 47(8): 1800327. doi: 10.1002/clen.201800327
[17] 车生泉, 于冰沁, 严魏. 海绵城市研究与应用: 以上海城乡绿地建设为例[M]. 上海: 上海交通大学出版社, 2015: 177−178. Che S Q, Yu B Q, Yan W. Research and practices for sponge city: taking examples of Shanghai urban and rural green space[M]. Shanghai: Jiaotong University Press, 2015: 177−178.
[18] 王文, 诸葛绪霞, 周炫. 植物截留观测方法综述[J]. 河海大学学报(自然科学版), 2010, 38(5):495−504. Wang W, Zhuge X X, Zhou X. Methods for plant interception measurement[J]. Journal of Hohai University (Natural Sciences), 2010, 38(5): 495−504.
[19] 余开亮. 亚高寒草甸不同退化程度冠层截留容量及其与冠层特征的关系[D]. 兰州: 兰州大学, 2012. Yu K L. Canopy rainfall storage capacity related to canopy properties along sub-alpine meadow degradation gradient[D]. Lanzhou: Lanzhou University, 2012.
[20] Garcia-Estringana P, Alonso-Blnso-Estr, Alegre J. Water storage capacity, stemflow and water funneling in Mediterranean shrubs[J]. Journal of Hydrology, 2010, 389(3/4): 363−372.
[21] 申晓瑜. 北京常见园林植物叶面积指数模型研究[D]. 北京: 北京林业大学, 2007. Shen X Y. Research on the model of leaf area index of common landscape plants in Beijing[M]. Beijing: Beijing Forestry University, 2007.
[22] 张毅川. 海绵城市导向下绿地典型下垫面的雨水特征及优化[D]. 武汉: 武汉大学, 2017. Zhang Y C. Rainwater characteristics and optimazition of typical inderlying surface of green space under sponge city: taking Xinxiang City as an example[D]. Wuhan: Wuhan University, 2017.
[23] 于璐, 苏德荣, 刘艺杉. 3种草坪草叶片的水分吸收特性研究[J]. 北京林业大学学报, 2013, 35(3):97−101. Yu L, Su D R, Liu Y S. Characters of leaf water absorption for three turfgrasses[J]. Journal of Beijing Forestry University, 2013, 35(3): 97−101.
[24] 康伟健, 刘东焕, 赵世伟, 等. 东北玉簪和白玉簪叶片水分利用效率的差异及原因分析[J]. 植物生理学报, 2017, 53(4):641−648. Kang W J, Liu D H, Zhao S W, et al. Causes of differences in water use efficiency between Hosta ensata and H. plantaginea leaves[J]. Plant Physiology Journal, 2017, 53(4): 641−648.
[25] 王立新, 李山山. 八宝景天叶片表面润湿性测试与疏水机理分析[J]. 河北科技大学学报, 2018, 39(1):1−8. Wang L X, Li S S. Wettability measurement and hydrophobicity mechanism analysis of leaf surface of Hylotelephium erythrostictum[J]. Journal of Hebei University of Science and Technology, 2018, 39(1): 1−8.
[26] 牟金磊. 北京市设计暴雨雨型分析[D]. 兰州: 兰州交通大学, 2011. Mou J L. Design storm pattern analysis in the city of Beijing[D]. Lanzhou: Lanzhou Jiaotong University, 2011.
[27] 游宇, 车伍, 张伟, 等. 8种园林乔木林冠对雨水截留作用的研究[J]. 中国给水排水, 2018, 34(9):121−127. You Y, Che W, Zhang W, et al. Effect of rainfall interception by eight species of garden arbors[J]. China Water & Wastewater, 2018, 34(9): 121−127.
[28] 陈然, 侯凯翔, 徐众, 等. 徐州市园林植物的冠层雨水截留能力分析[J]. 绿色科技, 2018(3):1−4. Chen R, Hou K X, Xu Z, et al. Analysis of canopy rainwater interception capacity of garden plants in Xuzhou City[J]. Journal of Green Science and Technology, 2018(3): 1−4.
[29] 刘艳丽, 王全九, 杨婷, 等. 不同植物截留特征的比较研究[J]. 水土保持学报, 2015, 29(3):172−177. Liu Y L, Wang Q J, Yang T, et al. Study on interception characteristics of different plants[J]. Journal of Soil and Water Conservation, 2015, 29(3): 172−177.
[30] 樊才睿, 李畅游, 贾克力, 等. 不同放牧制度下呼伦湖流域草原植被冠层截留[J]. 生态学报, 2015, 35(14):4716−4724. Fan C R, Li C Y, Jia K L, et al. Grass canopy interception of Hulun watershed under different grazing systems[J]. Acta Ecologica Sinica, 2015, 35(14): 4716−4724.
[31] ASCE. Design and construction of urban stormwater management systems[EB/OL]. (1992−10−12)[2019−09−18]. https://www.waterboards.ca.gov/losangeles/water_issues/programs/stormwater/municipal/AdminRecordOrderNoR4_2012_0175/2001LAMS4_AR/Items%20532%20to%20553_reduced.pdf.
[32] 白伟岚, 高亦珂, 方翠莲, 等. 一种复层景观草本植物群落的构建方法: 201410108836.3[P]. 2014−06−25. Bai W L, Gao Y K, Fang C L, et al. Method for constructing multi-level landscape herb community: 201410108836.3[P]. 2014−06−25.
[33] 鲍仁强. 树木在老城区海绵化改造中的应用策略研究[D]. 北京: 北京建筑大学, 2019. Bao R Q. Research on the application of urban trees in sponge city construction of old city[D]. Beijing: Beijing University of Civil Engineering and Architecture, 2019.
-
期刊类型引用(4)
1. 张苗苗,罗于洋,王树森,张丽娜,马成功,于胜利,王景圆. 内蒙古旺业甸华北落叶松人工林空间结构分析及其优化. 西北林学院学报. 2024(01): 81-87+107 .  百度学术
百度学术
2. 荆媛,魏爽,史文辉,马梓贺,王德宇,戎可. 天然次生林中小斑啄木鸟的取食偏好. 野生动物学报. 2024(01): 84-94 . 百度学术
3. 孙宇,刘盛,田佳歆,程福山,赵士博,王诗俊. 基于空间结构优化的长白落叶松人工林分间伐模型构建. 中南林业科技大学学报. 2023(01): 72-83 . 百度学术
4. 刘鑫,黄浪,卿东升,李建军. 基于Voronoi空间单元的林分空间结构智能优化研究. 林业资源管理. 2023(04): 27-35 . 百度学术
其他类型引用(2)




计量
- 文章访问数: 1968
- HTML全文浏览量: 775
- PDF下载量: 131
- 被引次数: 6
