
Inversion exploration on forest aboveground biomass of optical and SAR data supported by parameter optimization method
-
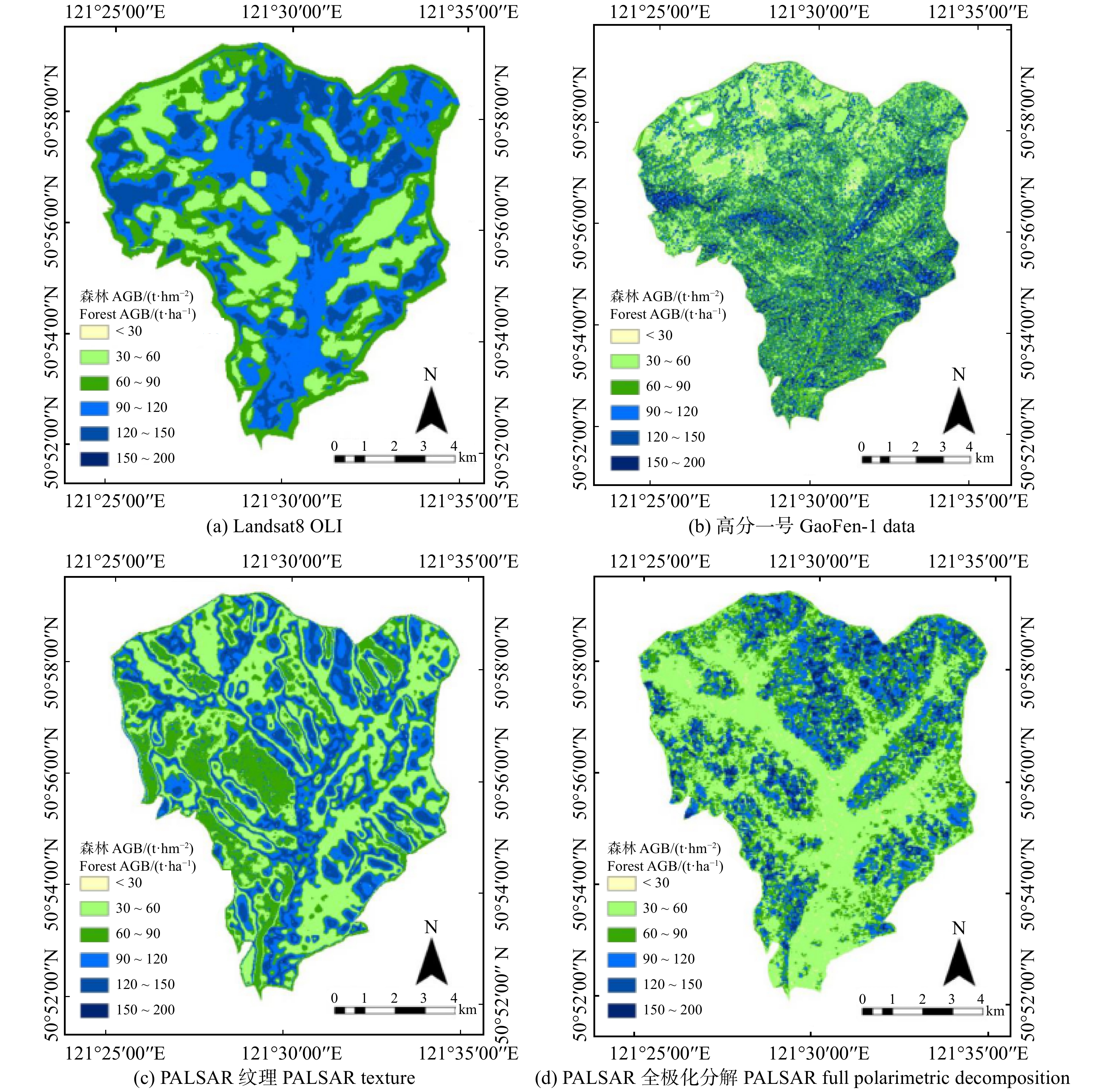
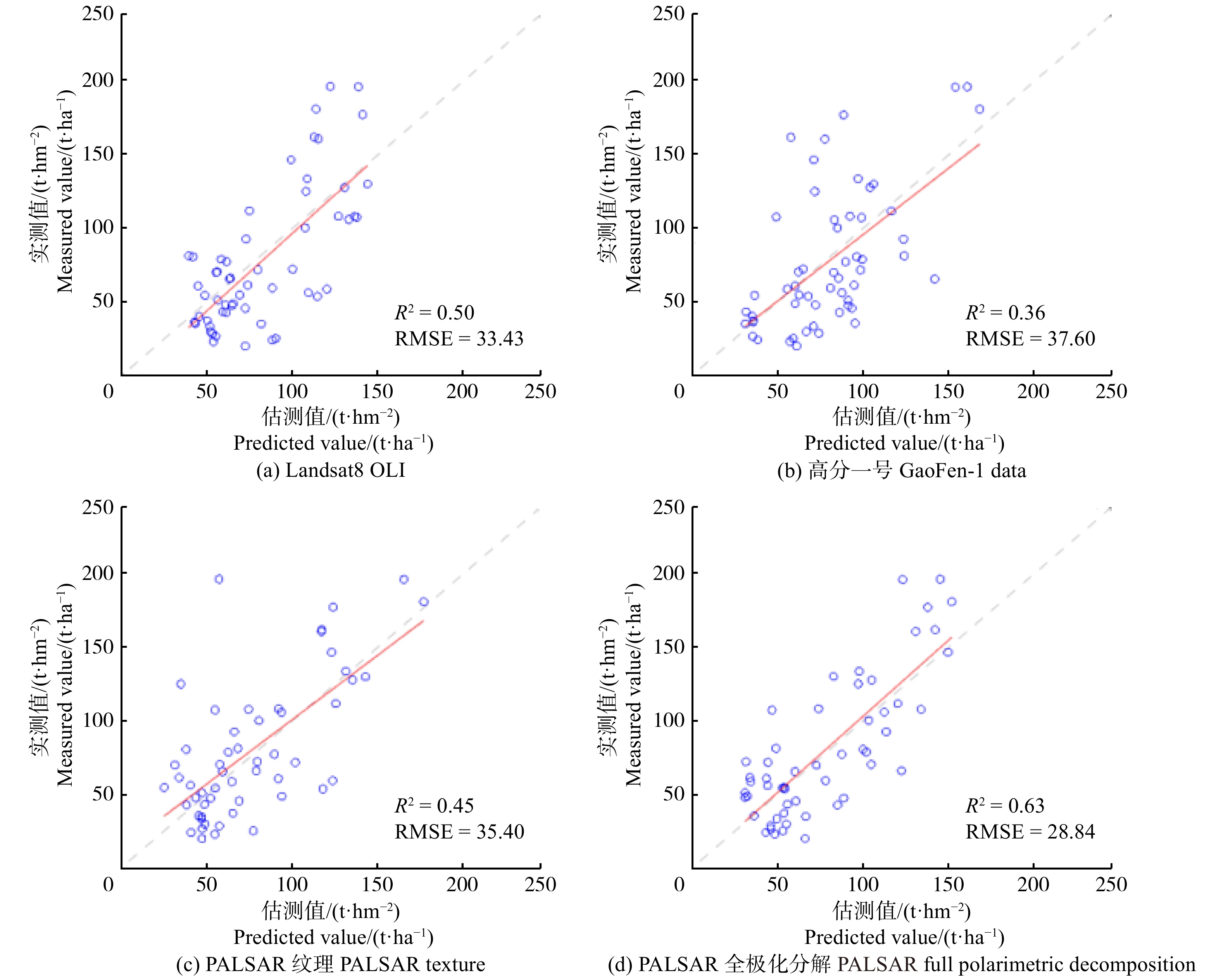
摘要:目的 森林是整个陆地碳循环系统中最大的有机碳贮库,准确地估测森林地上生物量影响着全球碳源与碳储量的分析与评价。本文旨在评价利用Landsat8 OLI、高分一号光学数据、ALOS-1 PALSAR-1SAR 3组不同源遥感数据估测森林AGB的潜力,进而剖析光学数据和SAR数据在估测森林AGB方面的差异。方法 首先对Landsat8 OLI、高分一号光学数据、ALOS-1 PALSAR-1SAR数据分别提取波段比值、植被指数、纹理信息,对ALOS-1 PALSAR-1SAR数据同时提取极化分解信息;然后,利用随机森林算法对不同数据提取的特征参数进行重要性排序,选择排序靠前的特征进行建模;最后,利用KNN-FIFS算法分析不同特征组合,对4组数据建立4个模型估测森林AGB,并使用留一交叉验证法对4个模型估测森林AGB值进行精度评价。结果 使用植被因子、波段比值、纹理因子、极化分解信息4种特征参数分别对3组数据进行建模估测森林AGB,基于Landsat8 OLI数据反演森林AGB的精度评价结果为R2 = 0.50,RMSE = 33.34 t/hm2;基于高分一号数据估测精度为R2 = 0.36,RMSE = 37.60 t/hm2;基于PALSAR纹理特征估测精度为R2 = 0.45,RMSE = 35.40 t/hm2;基于PALSAR全极化分解信息估测精度为R2 = 0.63,RMSE = 28.84 t/hm2。结论 参数提取方法相同时,即基于植被因子、波段比值、纹理信息3种特征参数估测森林AGB,其光学数据和SAR数据的反演潜力基本一致;参数提取方法不同时,即SAR数据加入极化分解信息估测森林AGB,与光学数据相比,SAR数据对森林AGB的反演潜力较好。Abstract:Objective Forest is the largest storage of organic carbon in the whole terrestrial carbon cycle system. Accurate estimation of forest aboveground biomass (AGB) is essential for global carbon storage analysis and estimation. This paper aims to explore the potential of optical and synthetic aperture radar (SAR) data for forest AGB inversion. In this study, Landsat8 OLI, GF-1 data were selected as optical data and advanced land observing satellite (ALOS)-1 phased array type L-band synthetic aperture radar (PALSAR)-1 data was selected as SAR data.Method Firstly, band ratio parameters, vegetation index parameters, and texture information were extracted from Landsat8OLI, gaofen-1 optical data and ALOS-1 PALSAR-1 SAR data, respectively. While polarization decomposition information was also extracted from ALOS-1 PALSAR-1 SAR data. Then these parameters extracted from different remote sensing data were sorted according to their importance by random forest (RF) algorithm. Finally, fast iterative feature selection method for k-nearest neighbor (KNN-FIFS) algorithm was used to analyze different feature combinations, and four models were constructed to estimate forest AGB and a cross-validation method was applied for result validation.Result These remote sensing data were modeled to estimate forest AGB using four characteristic parameters: vegetation factor, band ratio, texture factor and polarization decomposition information. For parameters extracted from vegetation factor, band ratio, texture factor, Landsat8 OLI data performed best than GF-1 and PALSAR-1 data with R2 = 0.50, RMSE = 33.34 t/ha. For GF-1 data, R2 was 0.36, RMSE was 37.60 t/ha, R2 was 0.45, RMSE was 35.40 t/ha for PALSAR-1 data. However, for parameters extracted from polarization decomposition, PALSAR-1data showed better performance with R2 = 0.63 and RMSE = 28.84 t/ha.Conclusion When the parameter extraction methods are the same, the forest AGB inversion potentials of the optical and SAR data are similar. However, when the parameter extraction method is different, for example, using polarization decomposition to extract parameters, the SAR data show better performance for forest AGB inversion.
-
Keywords:
- forest biomass /
- feature selection /
- KNN-FIFS
-
在当前全球气候不断变暖的背景下,中国气候发展趋势变化明显,其升温速度远远超过了同期的全球平均水平[1];作为由中纬度向高纬度过渡的地区,东北地区在过去50年中平均温度每10年增加0.3 ℃[2]。长白山位于我国东北温带季风气候区,其森林面积大,生境条件多样,且受到人为活动干扰少,是研究气候变化对森林生态系统影响的理想区域[3],近年来该地区生态环境受气候变化的影响被广泛关注[2]。
树木的生长和发育与气候因子紧密相关,它可以对环境变化和极端气候产生长期的响应并将其记录在年轮宽度信息之中[4-5]。树木年轮由于具有定年准确、分辨率高、连续性强、分布广泛、易于采样等特点[6],已成为研究气候变化对树木生长的影响和重建过去气候变化的关键资源[7-8]。近年来,已有大量研究运用树木年轮学从不同角度探讨长白山森林生长对全球气候变化的响应。例如:高露双等[9]通过研究阔叶红松(Pinus koraiensis) 林建群树种的生长−气候关系指出,不同树种的径向生长与气候因子之间的关系有所不同;王守乐等[10]研究了同一生境下不同树种对季节性气候的响应及其动态关系;于健等[11]研究了长白山地区群落交错带不同树种的径向生长对气候变暖的响应,但大部分研究仍集中于阔叶红松林中共存的两个树种与气候因子之间的关系,还缺乏对于云冷杉针阔混交林中多个共存树种径向生长对气候变化响应的研究。
汪清地区是长白山云冷杉针阔混交林的重要分布区,已有众多学者对该地区云冷杉针阔混交林进行研究,包括林分结构[12]、进界模型[13]、林下更新[14]等多个方面。为了解该地区云冷杉针阔混交林中不同树种对气候因子的响应特征及升温后气候因子对不同树种生长的影响。本研究以吉林汪清金沟岭林场云冷杉针阔混交林为研究对象,对臭冷杉(Abies nephrolepis)、鱼鳞云杉(Picea jezoensis)和红松3种常见树种进行采样,建立树木年轮宽度标准年表,分析其对气候因子的响应特征,探讨不同树种径向生长与气候因子的关系及升温对其生长的影响,为该地区树木的保护提供依据。
1. 研究区概况与研究方法
1.1 研究区概况
研究区位于吉林省延边汪清林业局金沟岭林场(43°17′ ~ 43°25′N、130°05′ ~ 130°20′E),属长白山系老爷岭山脉雪岭支脉,经营面积16 286 hm2。全场四面环山,以低山丘陵地貌为主,海拔500 ~ 1 200 m,坡度多在 5° ~ 25°。林区属温带大陆性季风气候,年均温4 ℃左右,年降水量600 ~ 700 mm。林区属低山灰化土灰棕壤区,母岩为玄武岩。研究区内的主要树种有臭冷杉、鱼鳞云杉、红松、红皮云杉(Picea koraiensis)、白桦(Betula platyphylla)、落叶松(Larix gmelinii)、枫桦(Betula costata)、紫椴(Tilia amurensis)、花楷槭(Acer ukurunduense)、色木槭(Acer mono)、青楷槭(Acer tegmentosum)、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、黄檗(Phellodendron amurense)等;草本植物以苔草(Carex tristachya)和禾本科(Gramineae)为主。
1.2 样品的采集与处理
2019年8月在典型云冷杉天然林,海拔710 m,西北坡向,按照径级选取生长良好的臭冷杉、鱼鳞云杉和红松作为取样目标,利用口径为5.15 mm的树木生长锥在胸高(1.3 m)位置钻取年轮样芯(尽可能通过髓芯)。为减少对树木的损害,每株树仅钻取一根样芯。臭冷杉、鱼鳞云杉和红松分别钻取了65、25和55根样芯。将采集的样芯进行编号并记录每株树的生境信息。将样品带回实验室后,对年轮样芯进行风干和打磨等处理。打磨时用200目、400目、600目的砂纸依次打磨,直到年轮界限在显微镜下清晰可见。
1.3 建立年表
1.3.1 年表的制作方法
利用LINTABTM6.0年轮宽度测量仪(测量精度为0.001 mm)及配套的TSAP软件测量年轮宽度。利用 COFECHA 程序[15]对定年和测量结果进行检验,剔除与主序列相关性差以及难以交叉定年的样芯,将保留的样芯用于建立年表。为消除其他非气候因素导致的生长趋势,利用R语言“dplr”包中的ModNegExp方法对树轮宽度序列进行去趋势和标准化处理[16],建立3个树种标准年表。
![]() 图 3 3种常见树种的标准年表和样本量Figure 3. Standard chronology and sample size of three common tree species
图 3 3种常见树种的标准年表和样本量Figure 3. Standard chronology and sample size of three common tree species1.3.2 标准年表统计参数
年轮宽度指数、信噪比、样本一阶自相关系数、样本间相关系数、平均敏感度、树木年轮的标准差、样本总体代表性等见文献。
(1)年轮宽度指数。在建立树木年轮宽度年表时,将树木年轮宽度序列用统计学方法进行曲线拟合,得到树木生长的期望值(Yi),树木年轮实际宽度值(Wi)与期望值(Yi)的比值为年轮宽度指数(Ii)。
Ii=WiYi (1) (2)信噪比。信噪比(signal-to-noise ratio,RSN)用于表示样本中所包含环境信息多少的统计量,其值为树木年表中气候信息与非气候噪声之比,若数值越大,则年表中包含的气候信息越多。
RSN=N¯rbt1−¯rbt (2) 式中:N代表样本数,
¯rbt 代表不同样芯间的平均相关系数。(3)平均敏感度。平均敏感度(mean sensitivity,SM)用于衡量年轮宽度的逐年变化,是无量纲值,其值越高,树木生长对气候变化越敏感,受到气候因子的限制作用越强。
SM=1n−1n−1∑i=1|2(xi+1−xi)xi+1+xi| (3) 式中:xi和xi + 1分别是第i个和第i + 1个年轮的宽度值;n为该树轮样本的年轮总数。
(4)样本总体代表性。样本总体代表性(expressed population signal,SEP)表示采集的树轮样本能够代表采样点森林总体生长趋势的百分比值,是决定采样是否成功的一项指标,其值越大,建立的树轮年表越具有理论年表的特征,采样工作越成功。
SEP=¯rbt¯rbt+(1−¯rbt)/N (4) 样本总体代表性的阈值通常设定为0.85[17]。
1.4 气象数据
从中国气象科学数据共享网(http://cdc.cma.gov.cn/home.do)获取距离采样点最近的汪清气象站(43°18′N、129°47′E,海拔244.8 m)的气象资料,时间为60年(1958—2018 年)。气象要素包含逐月的最低气温、最高气温、平均气温和总降水量。根据气象站内的降水量和温度数据使用R软件的“SPEI”包来计算标准化降水蒸散指数(standardized precipitation evapotranspiration index,SPEI)。利用 Mann-Kendall(M-K)方法[18]对年平均气温进行突变分析。
1.5 统计分析
利用Pearson相关分析分别对臭冷杉、鱼鳞云杉及红松的径向生长对逐月气候因子的响应进行分析。考虑到前一年气候的条件可能对树木当年径向生长存在影响[19],故选用上一年6月至当年9月的气候数据用于分析。为研究升温突变(1985年)后树木生长与气候关系的变化,对1985年升温前后主要气候因子与臭冷杉、鱼鳞云杉、红松径向生长的关系进行比较。利用滑动相关分析探讨径向生长与气候因子的动态关系,滑动窗口为32年。利用R软件计算各年表的参数,所有相关分析利用DendroClim2002程序[20]进行,使用Origin 2019软件绘制图表。
2. 结果与分析
2.1 气候因子变化特征分析
该区域具有雨热同期的规律,最低平均气温出现在1月(约−18 ℃),最高平均气温在7月(约6 ℃)。降水集中在5—9月,最大降水量在8月,10 月份开始降水逐渐减少,12月份降水量最低(图1)。
![]() 图 1 研究区1958—2018年气象数据T.平均气温;Tmin.最低气温;Tmax.最高气温;SPEI.标准化降水蒸散指数。T, mean temperature; Tmin, minimum temperature; Tmax, maximum temperature; SPEI, standardized precipitation evapotranspiration index.Figure 1. Meteorological data of research area from 1958 to 2018
图 1 研究区1958—2018年气象数据T.平均气温;Tmin.最低气温;Tmax.最高气温;SPEI.标准化降水蒸散指数。T, mean temperature; Tmin, minimum temperature; Tmax, maximum temperature; SPEI, standardized precipitation evapotranspiration index.Figure 1. Meteorological data of research area from 1958 to 2018Mann-Kendall的检验结果为1978年后正常的统计值时间序列UFk > 0,表明近40年平均气温呈增加趋势,且1985年为年平均气温升高的突变点(图2)。1958—1985年年均气温为4 ℃,1986—2018年年均气温为4.65 ℃,比突变前增加了0.65 ℃。1958—1985年年均降水量为44.58 mm,1986—2018年年均降水量为47.51 mm,增温突变后的年均降水量大于突变前。1958—1985年期间,平均气温以0.01 ℃/a的速率上升,但变化趋势不显著,平均最低气温以0.04 ℃/a的速率显著上升,平均最高气温变化趋势不显著,降水量以0.44 mm/a的速率显著下降,标准化蒸散指数以0.01的速率显著下降。1986—2018年期间平均最低气温以0.03 ℃/a的速率显著下降,平均最高气温以0.04 ℃/a的速率显著上升,平均气温、降水量和标准化蒸散指数变化均不显著。总体而言,汪清地区在1958—2018年期间高温变高,低温变低,平均气温升高,降水量及干旱情况不变。
![]() 图 2 1958—2018年年均气温 Mann-Kendall检验结果UFk.正常的统计值时间序列 ;UBk.逆序的统计值时间序列。UFk, normal statistical time sequence; UBk, inverted statistical time sequence.Figure 2. Mann-Kendall test results of average annual temperature from 1958 to 2018
图 2 1958—2018年年均气温 Mann-Kendall检验结果UFk.正常的统计值时间序列 ;UBk.逆序的统计值时间序列。UFk, normal statistical time sequence; UBk, inverted statistical time sequence.Figure 2. Mann-Kendall test results of average annual temperature from 1958 to 20182.2 标准年表的统计特征
臭冷杉、鱼鳞云杉和红松年表整体变化趋势差异显著,特别是在1985年前后(图3)。1985年前臭冷杉年轮指数有明显下降趋势(−0.03 a−1,R2 = 0.77),鱼鳞云杉和红松年轮指数下降趋势较弱;1985年后臭冷杉年轮指数出现显著上升趋势(0.05 a−1,R2 = 0.89),鱼鳞云杉次之(0.04 a−1,R2 = 0.81),红松年轮指数上升趋势较弱。
如表1所示,3个树种年表序列长度的平均值为110年,其中红松年表的时间跨度最长,达120年(1898—2018年),臭冷杉、鱼鳞云杉年表时间跨度分别为118年(1900—2018年)、92年(1926—2018年)。平均敏感度(SM)为0.16 ~ 0.27,信噪比(RSN)为6.14 ~ 19.98,臭冷杉的值均为最大。3个树种的平均敏感度、信噪比等指标均较高,说明所建立的年表包含较多的气候信息,对气候变化敏感,适用于气候因子分析。一阶自相关系数为0.81 ~ 0.86,说明汪清地区臭冷杉、鱼鳞云杉、红松年轮受前一年气候因子影响较大。样本总体代表性(SEP)高达0.86 ~ 0.92,均超过了可接受的临界阈值,表明样本包含的信号基本代表当地的总体特征。3个树种相比,虽然臭冷杉标准年表的一阶自相关系数低于鱼鳞云杉,但其平均敏感度、标准差、信噪比、样本总体代表性均高于另外两个树种,说明臭冷杉更适用于气候研究。总体上,各项年表特征均表明,臭冷杉、鱼鳞云杉和红松年表均适用于树木年轮气候学分析。
表 1 标准年表的统计特征及公共区间分析Table 1. Statistical characteristics and common interval analysis of the standard chronology统计特征 Statistic characteristics 臭冷杉 Abies nephrolepis 鱼鳞云杉 Picea jezoensis 红松 Pinus koraiensis 时间跨度 Time span 1900—2018 1926—2018 1898—2018 平均值 Mean value 0.93 0.96 0.95 平均敏感度 Mean sensitivity (SM) 0.27 0.16 0.22 标准差 Standard deviation (SD) 0.40 0.38 0.30 一阶自相关系数 First-order autocorrelation coefficient (AC) 0.81 0.86 0.81 公共区间 Common interval 1969—2011 1969—2011 1959—2013 信噪比 Signal-to-noise ratio (RSN) 19.98 6.14 11.21 样本间相关系数 Correlation coefficient between samples (Rbar) 0.40 0.31 0.32 样本总体代表性 Sample population representativeness (SEP) 0.95 0.86 0.92 2.3 不同树种径向生长对气候因子的响应特征
由图4、5可知,臭冷杉、鱼鳞云杉和红松径向生长均与上年生长季末期(9月)平均气温及上年、当年7月最低气温呈显著正相关(P < 0.05);鱼鳞云杉和红松与降水、标准化降水蒸散指数(SPEI)无显著性关系(P > 0.05);臭冷杉和红松与生长季前期(4—5月)最高气温呈显著正相关(P < 0.05)。此外臭冷杉径向生长与当年7月和上年7月平均气温呈显著正相关(P < 0.05),与当年5月和上年11月降水、SPEI呈显著正相关(P < 0.05);鱼鳞云杉与上年6月平均气温呈显著正相关(P < 0.05),与当年5月最高气温呈显著正相关(P < 0.05)。这表明臭冷杉年表较鱼鳞云杉和红松年表对气候因子响应更加敏感。臭冷杉径向生长受到气温和降水的制约,而鱼鳞云杉和红松径向生长的主要限制因子为温度。
![]() 图 4 标准年表与月最低气温、平均气温和最高气温的相关系数p.上年;c.当年。下同。p, previous year; c, current year. The same below.Figure 4. Correlation coefficients of the standard chronology with monthly minimum temperature, mean temperature and maximum temperature
图 4 标准年表与月最低气温、平均气温和最高气温的相关系数p.上年;c.当年。下同。p, previous year; c, current year. The same below.Figure 4. Correlation coefficients of the standard chronology with monthly minimum temperature, mean temperature and maximum temperature![]() 图 5 标准年表与月降水量、SPEI的相关系数Figure 5. Correlation coefficients of standard chronology with monthly precipitation and SPEI
图 5 标准年表与月降水量、SPEI的相关系数Figure 5. Correlation coefficients of standard chronology with monthly precipitation and SPEI2.4 不同树种径向生长对升温的响应差异
由于汪清地区的气候因子(气温和降水量)在1985年前后具有明显差别(图1、2),因此分两个时间段(1985—1985年和1986—2018年)分别评估温度和降水量对3个树种生长的影响。由图6可知,1958—1985年,臭冷杉和红松与上年9月、当年6月降水量呈显著正相关(P < 0.05),鱼鳞云杉与上年8、9月降水量呈显著正相关(P < 0.05),各树种与平均气温均未达到显著性水平(P > 0.05)。1986—2018年,臭冷杉径向生长与当年4月和7月平均气温呈显著正相关(P < 0.05),鱼鳞云杉径向生长与平均气温均未达到显著水平(P > 0.05),红松径向生长与当年4月平均气温呈显著正相关(P < 0.05),各树种与降水量均未达到显著性水平(P > 0.05)。由此可见,升温突变前后臭冷杉、鱼鳞云杉和红松径向生长与气候因子相关性均发生改变,主要表现为升温突变后径向生长与气温相关性增强但是与降水量相关性减弱。
![]() 图 6 升温突变前后3种常见树种标准年表与气候因子的相关分析*表示的显著相关(P < 0.05)。* means significant correlation (P < 0.05).Figure 6. Correlation analysis between the standard chronology of three common tree species and climate factors before and after the abrupt warming change
图 6 升温突变前后3种常见树种标准年表与气候因子的相关分析*表示的显著相关(P < 0.05)。* means significant correlation (P < 0.05).Figure 6. Correlation analysis between the standard chronology of three common tree species and climate factors before and after the abrupt warming change2.5 不同树种径向生长−气候关系的时间稳定性
由图7可知,上年6月和当年4、6月平均气温对臭冷杉径向生长的影响具有一定的长期稳定性。臭冷杉标准年表与上年9月、当年9月平均气温由显著负相关转为显著正相关,与当年6、7月平均气温由不显著相关转为显著正相关,说明夏季气温对臭冷杉生长的影响逐渐增强,秋季气温对臭冷杉径向生长由抑制作用转为促进作用。气温对红松的影响与臭冷杉和鱼鳞云杉相比,抑制作用较少。当年7月降水量与臭冷杉由显著负相关转为不显著相关,而与鱼鳞云杉和红松由不显著相关转为显著正相关。当年9月降水量对红松由促进作用变为抑制作用。上年6、7、12月和当年6、7、9月降水量对臭冷杉径向生长具有较明显的抑制作用,尤其是上年6月,显著相关关系持续时间长达11年(2005—2016)。
![]() 图 7 3种常见树种与气候因子的滑动相关分析Figure 7. Moving correlation analysis of three common tree species and climate factors
图 7 3种常见树种与气候因子的滑动相关分析Figure 7. Moving correlation analysis of three common tree species and climate factors3. 讨 论
3.1 臭冷杉、鱼鳞云杉和红松径向生长−气候关系差异
研究结果表明:臭冷杉、鱼鳞云杉和红松径向生长与均与上年9月平均气温呈显著正相关,这主要因为树木的生长集中在6—9月,上年9月为上个生长季的末期,而当上个生长季结束时较高的温度有利于延长生长季,保证树木正常的代谢活动,使树木能够储存更多的营养物质以供下一个生长季开始时树木的生长,从而表现出促进作用[21]。3个树种径向生长均与当年7月最低气温呈显著正相关,这可能是因为研究区在7、8月降水集中,水分充足,有利于增强植物的蒸腾作用,促进叶片气孔开放和CO2吸收,在水分充足的条件下,生长季的温度升高有助于提高酶的活性,增加细胞生产速率,进而促进树木径向生长[22]。以上结果表明同一区域不同树种径向生长对气候的响应具有一定的相似性,与杨婧雯等[23]的研究结果一致,这可以解释为树木对区域气候信号的共同响应[24]。
本研究中,臭冷杉径向生长与生长季前期降水有较强相关性,生长季前期(当年5月)降水量的增加,不仅可以有效补充土壤水分,缓解生长季温度过高造成的水分不足,还可以促进细胞运输营养物质和合成蛋白质,从而促进宽轮的形成[25]。树木需要通过根系从土壤吸收足够的水分来维持自身生理活动,作为喜湿、耐荫、浅根性树种,臭冷杉的生长很容易受到水分胁迫[24],因此臭冷杉生长受温度和降水的共同影响。鱼鳞云杉径向生长与5月最高气温均呈显著正相关,可能是因为5月温度的升高不仅可以促进地面积雪的融化,温暖土壤和茎分生组织,使土壤早日解冻,还可以提高光合作用,增加细胞生产速率,进而促进早期树木的生长[26-27]。红松径向生长与当年4月最高气温呈显著正相关,春季树木结束休眠转入生长季,较高的生长季前期温度可以促进生长季提前开始,从而延长生长季,而适宜的温度可以促进树木形成层活动,因此生长季前期足够的热量有利于树木形成宽轮[24]。该地区不同树种的生长与气候因子之间的关系存在一定差异可能是受树种遗传特性的影响[28]。此外,韩艳刚等[29]对长白山关于径向生长−气候关系已发表的文献总结的结果也显示长白山地区不同树种对气候因子的响应有所不同,在川西高原[30]、大兴安岭[23]等地的研究同样表明不同树种对气候因子的响应有差异。
3.2 气候变暖对不同树种径向生长的影响
本研究结果显示:升温突变(1985年)后,臭冷杉、鱼鳞云杉和红松径向生长与气温的正相关性增加,即升温突变后3个树种径向生长对气温的敏感性增加,且其径向生长具有与气温升高相一致的趋势;这与Andreu等[24]研究发现的树木生长随着气温升高对气候的敏感性增加的结果一致,而与王守乐等[10]研究显示的树木径向对气候的敏感性降低的结果不同。目前,已有很多研究提出气候变暖对树木径向生长影响。树木的径向生长随着气候变暖可能出现加快[31]、降低[32]、及变化不明显[33]等多种情况。这也验证了气候变暖后树木径向生长对气候因子的响应存在差异。
此外,结果显示,气温升高有利于3个树种的径向生长,这与于健等[11]在位于1 125 m海拔的研究地得出的气候变暖抑制鱼鳞云杉生长的研究结果不同。通常情况下,海拔可以改变树木的生长速度,进而影响树木生长与气候的关系。Zhu等[34]研究表明鱼鳞云杉对气候变暖的响应存在海拔差异,快速升温加速了高海拔鱼鳞云杉的径向生长,而减少了低海拔鱼鳞云杉的径向生长。但是本研究在海拔低于于健等[11]研究的情况下气候变暖仍然促进树木径向生长,这可能是由于气温的阈值效应,树木生长之所以会出现对气候因子响应的敏感性变化主要是因为该效应,气温的升高满足了树木生长的下限阈值,而逐渐突破了树木生长所需要的上限阈值[35]。当气温高于树木生长的上限阈值后,会使其蒸发、蒸腾作用加剧,从而导致土壤水分有效性降低,此时水分可能变成限制因素,出现干旱胁迫现象[31]。目前升温可能未超过臭冷杉、鱼鳞云杉和红松径向生长的临界温度,且由于汪清地区降水量充足(图1),故升温并未对该地区云冷杉林造成干旱胁迫,进而表现出对其径向生长的促进作用。李波[13]在2017年对汪清地区云冷杉林的研究中同样发现该地区年均温促进树木的径向生长。即使目前汪清地区树木生长并未受到气温升高的抑制作用,但是随着未来气候的持续变暖,该地区树木生长对气候的响应仍需持续关注。
4. 结 论
本研究表明,相比较而言,臭冷杉标准年表统计参数值高于鱼鳞云杉和红松,其对气候变化的响应更敏感,更适用于树轮气候学的研究。长白山汪清地区臭冷杉、鱼鳞云杉和红松径向生长对于特定气候因子的响应有一致性,上年生长季末期(9月)平均气温及上年、当年7月最低气温促进3个树种径向生长。同时,该地区树木径向生长−气候因子之间的关系也有着显著的差异,臭冷杉生长受气温和降水的制约,鱼鳞云杉和红松生长的主要限制因子为温度。这可能主要是因为树种的生理特征以及种间遗传因素存在差异。1985年升温前树木径向生长有显著下降趋势,升温后则出现上升趋势。升温突变后,臭冷杉、鱼鳞云杉和红松与气温和降水的相关性变化存在明显差异,气温对树木径向生长的促进作用增加而降水的促进作用减弱。总体而言,目前气温的升高促进汪清地区云冷杉林常见树种的径向生长。
-
表 1 ALOS PALSAR-1 影像主要参数
Table 1 Main parameters of ALOS PALSAR-1 images
成像时间
Acquisition time极化方式
Polarization way入射角
Incidence angle/(°)波长
Wavelength/m距离向分辨率
Range resolution/m方位向分辨率
Azimuth resolution/m卫星飞行轨道
Satellite orbit2011−04−12 HH, HV, VH, VV 23.799 0.236 9.368 3.554 升轨 Rail lifting 注:HH表示发射和接收的都为水平极化的电磁波;HV表示发射的电磁波为水平极化,接收的电磁波为垂直极化;VH表示发射的电磁波为垂直极化,接收的电磁波为水平极化;VV表示发射和接收的都为垂直极化的电磁波。Notes: HH, transmitting and receiving both with horizontal polarization; HV, transmitting with horizontal polarization and receiving with vertical polarization; VH, transmitting with vertical polarization and receiving with horizontal polarization; VV, transmitting and receiving both with vertical polarization.  下载: 导出CSV
下载: 导出CSV
表 2 3种数据源重要参数
Table 2 Major parameters of three data sources
数据源 Data source 重要参数 Major parameter OLI DVI, MEgree, PVI, CONSWIR1, VARSWIR1 GF HOMONIR, DISNIR, ENNIR, CONNIR, CORNIR PALSAR纹理 PALSAR texture VHME, HHME, VVME, VARVH, DISHV PALSAR全极化分解特征 PALSAR full polarimetric decomposition characteristics Yamaguchi4_vo1_dB, Yamaguchi4_vo1, Yamaguchi4_h1x_dB,
Yamaguchi4_Odd_dB, T22注:OLI为Landsat-8 OLI;GF为高分一号数据;DVI为差值植被指数;MEgree为绿光波段的均值纹理信息;PVI为正交植被指数;CONSWIR1、VARSWIR1分别为短波红外波段的对比度纹理信息、方差纹理信息;HOMONIR、DISNIR、ENNIR、CONNIR、CORNIR分别为近红外波段的均匀性纹理信息、相异性纹理信息、熵纹理信息、对比度纹理信息、相关性纹理信息;VHME、HHME、VVME分别为VH、HH、VV极化通道信息的均值纹理特征;VARVH为VH极化通道信息的方差纹理信息;DISHV为HV极化通道信息的相异性纹理信息;Yamaguchi4_vo1_dB、Yamaguchi4_vo1、Yamaguchi4_h1x_dB、Yamaguchi4_Odd_dB分别为Yamaguchi4分量分解中体散射分量分贝化结果、体散射分量、螺旋散射分量分贝化结果、表面散射分量分贝化结果;T22为相干矩阵中的一个元素。Notes: OLI is the Landsat-8 OLI data; GF is the Gaofen-1 data; DVI is the differential vegetation index; MEgree is the mean texture of green band; PVI is the perpendicular vegetation index; CONSWIR1 and VARSWIR1 are the contrast texture and variance texture of the short-wave infrared band, respectively; HOMONIR, DISNIR, ENNIR, CONNIR, CORNIR are the homogeneity texture, dissimilarity texture, entropy texture, contrast texture, correlation texture of the near-infrared band, respectively; VHME, HHME, VVME are the mean texture of polarization channel of VH, HH, VV, respectively; VARVH is the variance texture of VH channel; DISHV is the dissimilarity texture of the HV channel; Yamaguchi4_vo1_dB, Yamaguchi4_vo1, Yamaguchi4_h1x_dB, Yamaguchi4_Odd_dB are the decibel results of volume scattering component, volume scattering component, helix scattering component and surface scattering component in the decomposition of Yamaguchi4 component decompositions, respectively; T22 is an element in the coherence matrix.
下载: 导出CSV
表 3 特征组合
Table 3 Feature combination
数据源
Data sourceK 窗口大小
Size of the window特征组合
Combination of feature vectorOLI 8 11 × 11 DVI, VARSWIR1, CONgreen, MEgree GF 3 3 × 3 HOMONIR, NIR, CONNIR PALSAR纹理 PALSAR texture 9 7 × 7 HHME, VVME PALSAR全极化分解特征
PALSAR full polarimetric decomposition characteristics3 7 × 7 alpha, T23_imag, Freeman_Odd, anisotropy, T33, T12_real 注:DVI为差值植被指数;VARSWIR为短波红外波段swir1的方差纹理信息;CONgree、MEgree为绿光波段的对比度纹理信息、均值纹理信息;HOMONIR、NIR、CONNIR为近红外波段的均匀性纹理信息、光谱信息、对比度纹理信息;HHME为HH极化通道信息的均值纹理特征;VVME为VV极化通道信息的均值纹理特征;alpha为H-A- ¯α 分解中的平均散射角;T23_imag为相干矩阵中T23分量的虚部;Freeman_Odd为Freeman三分量分解的表面散射分量;anisotropy为H-A-¯α 分解中的极化各向异质性;T33为相干矩阵中的一个分量;T12_real为相干矩阵中T12分量的实部。Notes: DVI is the differential vegetation index; VARSWIR1 is the variance texture of the short-wave infrared band; CONgree and MEgree are the contrast texture and mean texture of green band, respectively; HOMONIR, NIR and CONNIR are the homogeneity texture, spectral information and the contrast texture of the near-infrared band, respectively; HHME and VVME are the mean texture of polarization channel of HH and VV, respectively; alpha is the average scattering angle of H-A-¯α decomposition; T23_imag is the imaginary part of the T23 component in the coherence matrix; Freeman_Odd is the surface scattering component of Freeman-Durden 3 components decomposition; anisotropy is the polarimetric anisotropy of the H-A-¯α decomposition; T33 is a component of the component matrix; T12_real is the real part of the T12 component of the coherence matrix.
下载: 导出CSV
-
[1] 陶玉华. 森林生态系统碳储量研究的意义及国内外研究进展[J]. 现代农业科技, 2012(9):205, 212. Tao Y H. Research significance of carbon storage of forest ecosystem and research progress at home and abroad[J]. Modern Agricultural Sciences and Technology, 2012(9): 205, 212.
[2] 杨洪晓, 吴波, 张金屯, 等. 森林生态系统的固碳功能和碳储量研究进展[J]. 北京师范大学学报(自然科学版), 2005, 41(2):172−177. doi: 10.3321/j.issn:0476-0301.2005.02.018. Yang H X, Wu B, Zhang J T, et al. Progress of research into carbon fixation and storage of forest ecosystems[J]. Journal of Beijing Normal University (Natural Science), 2005, 41(2): 172−177. doi: 10.3321/j.issn:0476-0301.2005.02.018.
[3] Lu D S, Chen Q, Wang G X, et al. A survey of remote sensing-based aboveground biomass estimation methods in forest ecosystems[J]. International Journal of Digital Earth, 2014, 9(1): 63−105.
[4] 韩宗涛, 江洪, 王威, 等. 基于多源遥感的森林地上生物量KNN-FIFS估测[J]. 林业科学, 2018, 54(9):70−79. doi: 10.11707/j.1001-7488.20180909. Han Z T, Jiang H, Wang W, et al. Forest above-ground biomass estimation using KNN-FIFS method based on multi-source remote sensing data[J]. Scientia Silvae Sinicae, 2018, 54(9): 70−79. doi: 10.11707/j.1001-7488.20180909.
[5] 李春梅. 基于多源数据的森林地上生物量估测方法研究[D]. 昆明: 西南林业大学, 2016. Li C M. A study of the retrieval methods of forest above ground biomass based on multi-source data[D]. Kunming: Southwest Forestry University, 2016.
[6] 李春梅, 张王菲, 李增元, 等. 基于多源数据的根河实验区生物量反演研究[J]. 北京林业大学学报, 2016, 38(3):64−72. Li C M, Zhang W F, Li Z Y, et al. Retrieval of forest above-ground biomass using multi-source data in Genhe, Inner Mongolia[J]. Journal of Beijing Forestry University, 2016, 38(3): 64−72.
[7] 庞勇, 李增元, 谭炳香, 等. 点云密度对机载激光雷达林分高度反演的影响[J]. 林业科学研究, 2008, 21(增刊): 14−19. Pang Y, Li Z Y, Tan B X, et al. The effects of airborne LiDAR point density on forest height estimation[J]. Forest Research, 2008, 21(Suppl.): 14−19.
[8] 刘清旺, 李增元, 陈尔学, 等. 机载LIDAR点云数据估测单株木生物量[J]. 高技术通讯, 2010, 20(7):765−770. doi: 10.3772/j.issn.1002-0470.2010.07.019 Liu Q W, Li Z Y, Chen E X, et al. Estimating biomass of individual trees using point cloud data of airborne[J]. Chinese High Technology Letters, 2010, 20(7): 765−770. doi: 10.3772/j.issn.1002-0470.2010.07.019
[9] 穆喜云, 张秋良, 刘清旺, 等. 基于激光雷达的大兴安岭典型森林生物量制图技术研究[J]. 遥感技术与应用, 2015, 30(2):220−225. Mu X Y, Zhang Q L, Liu Q W, et al. A study on typical forest biomass mapping technology of Great Khingan using airborne laser scanner data[J]. Remote Sensing Technology and Application, 2015, 30(2): 220−225.
[10] 陈尔学. 合成孔径雷达森林生物量估测研究进展[J]. 世界林业研究, 1999, 12(6):18−23. doi: 10.3969/j.issn.1001-4241.1999.06.004 Chen E X. Development of forest biomass estimation using SAR data[J]. World Forestry Research, 1999, 12(6): 18−23. doi: 10.3969/j.issn.1001-4241.1999.06.004
[11] 张王菲, 陈尔学, 李增元, 等. 干涉、极化干涉SAR技术森林高度估测算法研究进展[J]. 遥感技术与应用, 2017, 32(6):983−997. Zhang W F, Chen E X, Li Z Y, et al. Development of forest height estimation using InSAR/PolInSAR technology[J]. Remote Sensing Technology and Application, 2017, 32(6): 983−997.
[12] Pu L, Zhang X L, Shi J, et al. Adaptive filtering for 3D SAR data based on dynamic gaussian threshold[C]// Proceedings of the 6th Asia-Pacific Conference on Synthetic Aperture Radar (APSAR). Xiamen: IEEE, 2019.
[13] 张王菲. 星载SAR遥感反演中地形辐射校正的关键技术研究[D]. 昆明: 昆明理工大学, 2011. Zhang W F. Study on the key techniques of terrain radiometric correction in spaceborne SAR remote sensing inversion[D]. Kunming: Kunming University of Science and Technology, 2011.
[14] 杨浩. 基于时间序列全极化与简缩极化SAR的作物定量监测研究[D]. 北京: 中国林业科学研究院, 2015. Yang H. Study on quantitative crop monitoring by time series of fully polarimetric and compact polarimetric Sar imagery[D]. Beijing: Chinese Academy of Forestry, 2015.
[15] 陈传国. 东北主要林木生物量手册[M]. 北京: 中国林业出版社, 1989. Chen C G. Manual of main forest biomass in Northeast China[M]. Beijing: China Forestry Publishing House, 1989.
[16] Haralick R M. Statistical and structural approaches to texture[J]. Proceedings of the IEEE, 1979, 67(5): 786−804. doi: 10.1109/PROC.1979.11328
[17] Tuominen S, Pekkarinen A. Performance of different spectral and textural aerial photograph features in multi-source forest inventory[J]. Remote Sensing of Environment, 2005, 94(2): 256−268. doi: 10.1016/j.rse.2004.10.001
[18] Chica-Olmo M, Abarca-Hernández F. Computing geostatistical image texture for remotely sensed data classification[J]. Computers & Geosciences, 2000, 26(4): 373−383.
[19] 张腊梅, 段宝龙, 邹斌. 极化SAR图像目标分解方法的研究进展[J]. 电子与信息学报, 2016, 38(12):3289−3297. Zhang L M, Duan B L, Zou B. Research development on target decomposition method of polarimetric SAR image[J]. Journal of Electronics and Information Technology, 2016, 38(12): 3289−3297.
[20] Breiman L. Random forests[D]. Berkeley: University of California, 2001: 5−26
[21] 李欣海. 随机森林模型在分类与回归分析中的应用[J]. 应用昆虫学报, 2013, 54(4):1190−1197. doi: 10.7679/j.issn.2095-1353.2013.163 Li X H. Using “random forest” for classification and regression[J]. Chinese Journal of Applied Entomology, 2013, 54(4): 1190−1197. doi: 10.7679/j.issn.2095-1353.2013.163
[22] 苏瑞雪, 汤玉奇. 光学-极化SAR影像特征融合与分类[J]. 测绘与空间地理信息, 2019(6):51−55. doi: 10.3969/j.issn.1672-5867.2019.06.014 Su R X, Tang Y Q. Feature fusion and classification of optical-PolSAR images[J]. Geomatics & Spatial Information Technology, 2019(6): 51−55. doi: 10.3969/j.issn.1672-5867.2019.06.014
[23] 范亚雄. 星载X-波段干涉SAR森林高度估测方法研究[D]. 北京: 中国林业科学研究院, 2019. Fan Y X. Study on forest height estimation method of spaceborne X- band interferometric SAR[D]. Beijing: Chinese Academy of Forestry, 2019.
-
期刊类型引用(3)
1. 刘索名,王君杰,燕云飞,姜立春. 小兴安岭兴安落叶松人工林冠幅模型构建. 北京林业大学学报. 2023(05): 79-87 .  本站查看
本站查看
2. 李应涛,刘时良,孙海龙,王卫霞,向玮. 云冷杉针阔混交林单木枝下高和冠幅模型构建. 森林与环境学报. 2022(03): 289-296 . 百度学术
3. 佟艺玟,陈东升,冯健,高慧淋. 基于线性分位数混合效应的辽东山区红松冠幅模型. 应用生态学报. 2022(09): 2321-2330 . 百度学术
其他类型引用(1)
















计量
- 文章访问数: 1411
- HTML全文浏览量: 430
- PDF下载量: 102
- 被引次数: 4
