
Fine root anatomical and morphological traits of three temperate liana species in northeastern China
-
摘要:目的 研究温带木质藤本植物细根的功能性状及其潜在吸收能力。方法 选取中国东北温带3种代表性木质藤本植物,即缠绕类藤本狗枣猕猴桃和北五味子、卷须攀援类藤本山葡萄为材料,研究了1 ~ 5级细根的解剖、形态、分支结构和吸收根组成。结果 随着根序的增加,狗枣猕猴桃和北五味子的皮层厚度有增加趋势,而山葡萄则呈降低趋势。3种藤本植物的中柱直径均随根序增加而显著增粗,但是皮层厚度与中柱直径比值显著下降。皮层的存留状态显示,所有树种吸收根的比例均随根序增加而明显下降,1级根是典型的吸收根。从1级根至5级根,3种藤本植物比根长显著降低,直径显著增粗,组织密度增加,但是分支比仅在山葡萄中呈现单调的降低趋势。树种对各根系性状有显著的影响,但是与根序的影响存在交互作用。与其他2个缠绕类藤本植物相比,卷须攀援类藤本山葡萄中柱直径较大,皮层厚度与中柱直径比值较低,而2、3级根的分支比明显更高。与本地区其他类群植物相比,3种藤本植物1级根的平均直径较粗、比根长较低,而平均组织密度介于草本蕨类和其他木本植物之间。结论 细根解剖和形态性状在3种藤本植物间存在明显差异,但是这些性状随根序变化的规律与其他木本植物较一致。与本地区其他类群植物相比,藤本植物1级根形态特征表现出了较为独立的特点。Abstract:Objective The objective was to investigate the functional traits and potential absorbing capacity in fine roots of liana (woody climbing plants) in temperate forests.Method We sampled three typical temperate liana species in northeastern China, including two twining climbers (Actinidia kolomikta and Schisandra chinensis) and one tendril climber (Vitis amurensis). The anatomical, morphological, and branching traits, as well as the composition of absorptive roots were examined for the grade 1−5 fine roots in the three species.Result The results showed that the cortical thickness of Actinidia kolomikta and Schisandra chinensis increased by ascending branch order, while that of Vitis amurensis decreased. The stele diameter of all three liana species significantly increased with increasing branch order, but the ratio of cortical thickness to stele diameter significantly reduced. In term of the status of cortex appearance, the proportion of roots with absorbing function apparently decreased with ascending branch order across all three species, with the grade 1 fine roots as typical absorptive roots. From grade 1 to 5, specific root length significantly decreased, but root diameter and tissue density increased. The decline in root branching ratio associated with increasing branch order was only found in V. amurensis. Those root functional traits in liana were significantly influenced by tree species, but showing significant interaction with branch orders. Compared with two twining climbers, tendril climber of V. amurensis had wider stele, lower ratio of cortical thickness to stele diameter, and much higher branching ratio to the grade 2 and grade 3 roots. In comparison to other woody plants and herbaceous ferns at the same study site, mean root diameter of three liana species was thicker but mean specific root length was lower, with intermediate mean tissue root density.Conclusion Although the interspecific differences in root anatomical and morphological traits exist in three temperate liana species, the variation patterns of those root traits associated with increasing branch order are consistent with other woody plants. The morphological traits of the grade 1 roots in these lianas display distinctive characteristics compared with other plant taxa in this region.
-
Keywords:
- root order /
- woody liana /
- fine root /
- functional trait /
- resource acquisition
-
六盘山位于黄土高原西北部,不仅是干旱区−半干旱区的过渡区,也是黄土区−土石山区的过渡区,具有特殊的地理位置。华北落叶松是六盘山区的主要造林树种,在木材生产和水土保持方面发挥着重要的作用[1],但由于该地区气候梯度和年际波动大,降水分布不均,大面积造林虽然增加了植被覆盖率,却使区域生态耗水增加,河川径流减少[2],林水矛盾日益突出。森林蒸散是土壤−植物−大气系统(SPAC)中水热传输和能量转换的重要环节之一[3-4],是植被对干旱等环境条件响应的关键过程[5-6],其中,林分蒸腾是森林蒸散的最主要组分[7],约占 40% ~ 70%[8]。因此,精确推算和预测林分蒸腾及其变化规律,揭示其与环境因子之间的关系,将有助于深入理解林水关系,并为当地植被恢复和林水管理提供理论指导。
以边材面积为空间纯量将树干液流进行尺度上推计算整株和林分蒸腾量的方法已经被广泛应用[9-10]。但不同时间尺度下,影响树干液流的生态因子会发生显著变化,进而影响林木/林分蒸腾的变化。目前,对不同时间尺度林木/林分蒸腾对环境因子的响应已开展了一些研究,如张静等[11]对苹果林的蒸腾研究发现,空气温度和饱和水汽压差均是日、月尺度下苹果林蒸腾的主要因子。李振华[12]发现,潜在蒸散量可解释华北落叶松林分日蒸腾量变异的81%。Liu等[13]发现林分日蒸腾量受潜在蒸散(PET)、叶面积指数(LAI)和不同深度土壤相对可利用水分(REW)的共同影响。王文杰等[14]发现,随时间尺度的减小,影响树干液流的主要因子将由土壤环境因子(作用于根系的水分吸收)向大气因子如光照和空气湿度(作用于叶片气孔)转变。但这些研究多在日、年尺度下探讨林分蒸腾对单个或多个环境条件的响应,较少关注小时尺度林分蒸腾对环境因子的响应,同时缺乏小时尺度林分蒸腾对多因子响应的耦合模型。事实上,随着全球气候逐渐变暖,导致干旱加剧、短时和极端天气条件频发[15],日尺度上的研究无法揭示树干液流对日内剧烈变化的气象条件的响应,这将影响林分蒸腾的准确估计,限制对林木蒸腾响应环境变化的深入理解,因此有必要开展日内尺度的林分蒸腾的研究。因此,本研究以宁夏六盘山香水河小流域华北落叶松(Larix principis-rupprechtii)为研究对象,利用热扩散液流探针(SF-L)连续监测2018年生长季(5—10月)样树的树干液流,同步观测气象条件和土壤含水量,并拟合多因子影响的蒸腾模型,以期揭示短时间(小时)尺度内蒸腾对环境因子的响应规律,为当地林水管理提供理论依据。
1. 研究区概况
研究地点位于宁夏六盘山香水河小流域(106°12′ ~ 106°16′ E、35°27′ ~ 35°33′ N),面积为43.7 km2,海拔为2 070 ~ 2 931 m。该区属于温带半湿润气候,年均气温为5.8 ℃, 年均降水量为618 mm(1981—2010年)。主要土壤类型为灰褐土和高寒草甸土,其剖面主要分为枯落物层、腐殖质层、有机矿质层、淀积层和母质层[16],土壤砾石含量高,平均土层厚度约80 cm[17]。人工纯林占小流域面积的24%,其中华北落叶松约占人工林总面积的90%。该小流域的分水岭沟子流域内华北落叶松均为同年栽植,经营历史相似,坡向均为东南坡向,因此,在该子流域选取一个水平坡长为425.1 m,平均坡度为27.8°的典型华北落叶松坡面,在坡脚处布设了一个长期固定监测样地,样地大小为30 m × 30 m,海拔为2 290 m, 坡度为37.6°。样地林分基本信息及0 ~ 80 cm土壤物理性质见表1。
表 1 研究样地基本信息Table 1. Basic information of the study sample plot平均胸径
Mean DBH/cm平均树高
Mean tree height/m平均冠幅
Mean crown width/m林分密度/(株·hm− 2)
Stand density/
(tree·ha− 1)郁闭度Canopy density 土壤密度
Soil bulk density/
(g·cm− 3)总孔隙度
Total
porosity/%毛管孔隙度
Capillary porosity/%非毛管孔隙度
Noncapillary porosity/%18.71 ± 4.03 16.19 ± 2.34 3.27 ± 0.81 907 0.72 0.94 56.65 40.56 16.09 2. 研究方法
2.1 气象条件监测
在距样地100 m处的林外空旷地安装一台WheatherHawk-232自动气象站(WheatherHawk,美国)记录气象条件,自计频率为5 min。测定的指标包括空气温度(Ta,°C)、空气相对湿度(RH,%)、降雨量(P,mm)、太阳辐射(Rs,W/m2)、风速(Ws,m/s)和风向。饱和水汽压差(vapor pressure deficit,VPD,kPa)由当时气温条件下空气中的饱和水汽压与实际水汽压之差计算而得[18]:
VPD=0.611×(1−RH100)×e[17.502TaTa+240.97] (1) 式中:Ta为空气温度(°C);RH为空气相对湿度(%)。
2.2 土壤水分监测
根系层土壤水分对树木生长有重要作用,林木蒸腾直接受主根系层土壤水分的影响[19],而研究区华北落叶松根系层主要分布在0 ~ 60 cm[20], 故本研究只关注根系层的土壤湿度。土壤湿度随土层深度变化有较大的空间变异[20],且变异程度随土层的增加而逐渐降低,所以需在不同土层深度布设探头,且在表层进行加密观测。鉴于此,在样地内按0 ~ 10 cm、10 ~ 20 cm、20 ~ 40 cm、40 ~ 60 cm土层深度布设土壤湿度、温度传感器(5TE,Decagon,美国),监测不同土层的土壤湿度动态变化规律。传感器与EM50(Em50,Decagon,美国)数据采集器相连,记录频率为 5 min一次。土壤可利用水分(relative extractable water,REW)的计算公式如下:
REW=VSM−VSMmin (2) 式中:VSM为0 ~ 60 cm土层实际土壤含水量的平均值(m3/m3),VSMmin为0 ~ 60 cm土层的最小土壤含水量(m3/m3),VSMmax为0 ~ 60 cm土层的最大土壤含水量(m3/m3)。
2.3 树干液流的测定与计算
在样地内按树干径级(< 14 cm、14 ~ 17 cm、17 ~ 20 cm、> 20 cm)各径级选取1棵生长发育良好,无病虫害的树木做为样树用于监测树干液流,树干液流测定采用四针式热扩散液流探针(SF-L,Ecomatik,德国)。探针安装在树干北侧1.3 m处。安装探针时,首先将老化的树皮去除,将探针(S1、S0、S2、S3)按顺序插入树干内,注意加热探针(S0)应位于上方。探针安装好后,用玻璃胶将探针与树干之间的缝隙密封,用铝箔纸将探针部位的树干包裹,防止雨水及环境热辐射对探针测定数据的影响。探针与数据采集器DL2e(Delta-T Devices,英国)相连,每5 min记录一次数据。
单株液流速率(Ji,g/(m2·s))及平均液流速率
\overline {J_{\rm{s}}} (g/(m2·s))的计算方法为:{J_i} = 119 {\left( {\left( {{d_{{\rm{tmax}}}}/{d_{{\rm{tact}}}}} \right) - 1} \right)^{1.231}} (3) {d_{{\rm{tact}}}} = {T_{1 - 0}} - \left( {{T_{1 - 2}} + {T_{1 - 3}}} \right)/2 (4) \overline {J_{\rm{s}}} = \frac{{\displaystyle\mathop \sum \nolimits_i^n {J_i} \times {A_i}}}{{\displaystyle\mathop \sum \nolimits_{i = 1}^n {A_i}}} (5) 式中:dtmax为无液流时的最大温差(℃),即最大dt值;dtact为实际的dt值(℃);T1−0、T1−2、T1−3分别为探针S1与S0、S2、S3之间的温度差(℃)。n为样树株数,Ai为第i株样树的边材面积(cm2)。
2.4 林分蒸腾量的计算
林分小时蒸腾量(T,mm/h)由样树的液流速率和边面积上推得到:
T = \overline {J_{\rm{s}}} \left( {{\displaystyle\mathop \sum \nolimits_{i = 1}^n {A_i}}} \right) {S}^{-1} \times 60 \times 60 \times {10^{ - 3}} (6) 式中:S为样地面积(m2)。单株树干的边材面积Ai(cm2)由Liu[13]基于边材面积与胸径之间的函数关系拟合得来,见公式:
{A_i} = 0.255 \; 1 {\rm{DB}}{{\rm{H}}^{2.189 \; 2}} (7) 2.5 数据处理
影响树木/林分蒸腾的环境因子众多,且各因素相互影响,很难区分出单个因素的影响。因此,本文使用上外包线法进行单因素分析,并剥离每个因素对蒸腾量的影响,该方法可在一定程度上消除其他影响因素的干扰[21]。同时,假设各单因素对蒸腾量的影响是相互独立的,因此蒸腾量受各因素影响的耦合模型为其对每个单因素响应函数的连乘关系:
T = f\left( {X_1} \right) f\left( {X_2} \right) f\left( {X_3} \right) \cdots f\left( {X_n} \right) (8) 式中:T为林分蒸腾量,
f\left( {X_1} \right) 、f\left( {X_2} \right) 、f\left( {X_3} \right) ···f\left( {X_n} \right) 分别表示林分蒸腾量对单个因素的响应函数,n为因子数。用R2和无量纲的纳什系数(NSE)来评价模型模拟的质量[22]:
{\rm{NSE}} = 1 - \frac{{\displaystyle\mathop \sum \nolimits_i^n {{\left( {{y_i} - {{y'}_i}} \right)}^2}}}{{\displaystyle\mathop \sum \nolimits_i^n {{\left( {{y_i} - \bar y} \right)}^2}}} (9) 式中:yi为实测值;y′i为预测值;
\bar y 为所有实测值的平均值,NES越接近于1,表明模型拟合精度越高。3. 结果与分析
3.1 环境因子的季节变化特征
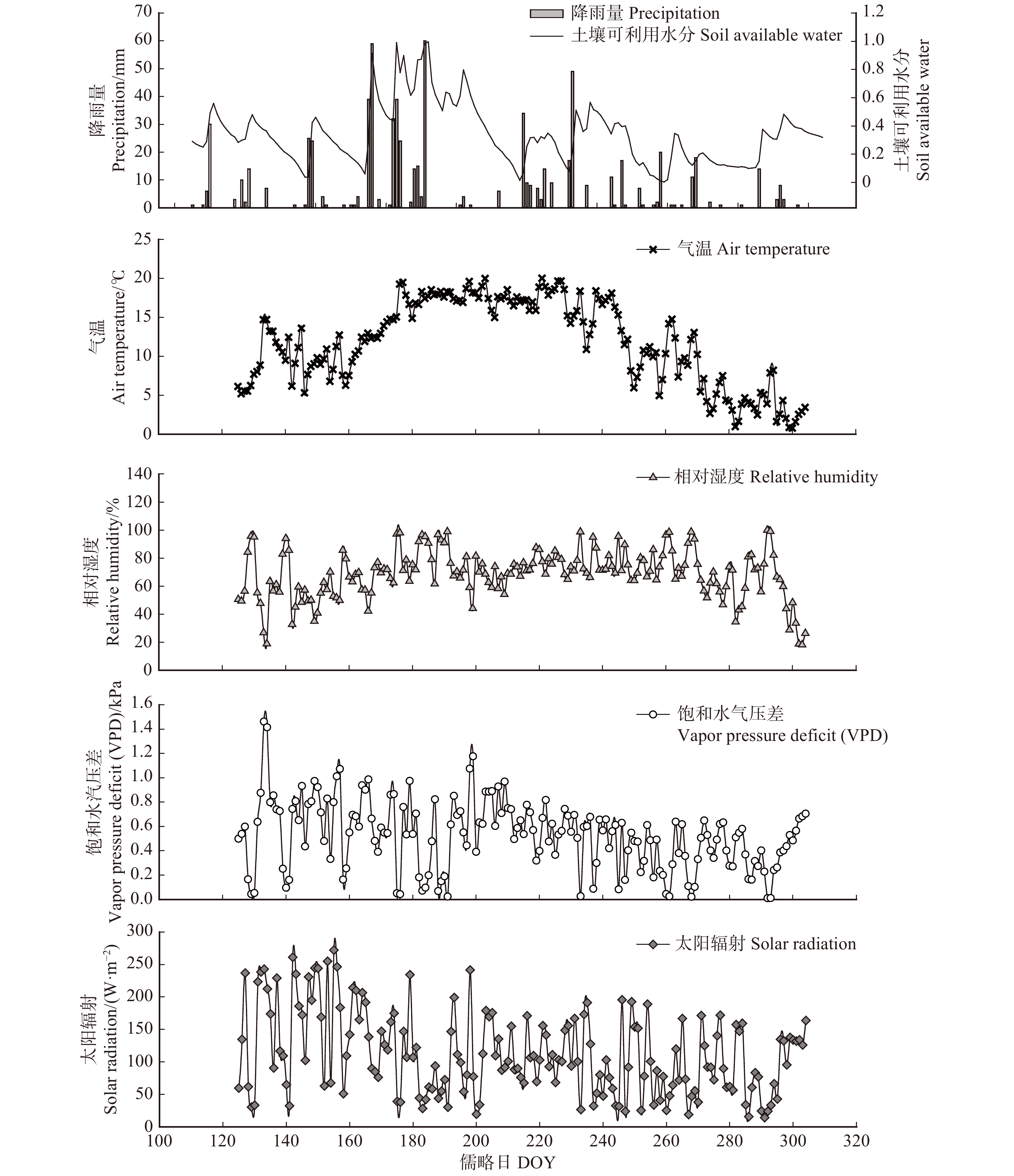
研究期间共观测到降水68场,累积降雨量为728 mm。降雨的季节动态呈单峰型变化格局,生长季初期(5月)和末期(9—10月)降水较少,6—8月降水较多。其中,6—8月总降水量占全年累计降水量的72%,最大的两场降雨分别发生在6月25日(59 mm)和7月10日(60 mm)。观测期间土壤相对可利用水分(REW)的变化范围为0.003 ~ 0.997,平均值为0.337。REW在6月25日至7月26日之间保持较高水平,与该时间段内连续的强降雨事件有关。分析数据发现,降水事件后的第2天,REW会大幅度升高,说明REW对降雨的响应存在一定程度的滞后。日均气温的变化趋势表现为随儒略日(DOY)的增加先升后降,变化范围为0.79 ~ 20.03 ℃,平均值为11.71 ℃,月均值表现为7月最大(17.75 ℃),10月最小(3.72 ℃)。观测期间空气相对湿度呈先增后减的变化趋势,变化范围为18% ~ 100%,平均值为68.3%。饱和水汽压差(VPD)的变化范围为0.01 ~ 1.46 kPa,平均值为0.53 kPa;太阳辐射强度(Rs)的变化范围为14.02 ~ 272.23 W/m2,平均值为113.96 W/m2,二者的月均值均表现为5月最大(0.66 kPa,160.6 W/m2),9月最小(0.36 kPa,87.41 W/m2)。2018年生长季环境因子的变化规律见图1。
![]() 图 1 2018年生长季内环境因子变化Figure 1. Variations of environmental factors in growth season of 2018
图 1 2018年生长季内环境因子变化Figure 1. Variations of environmental factors in growth season of 20183.2 树干液流速率及林分蒸腾的日内变化
华北落叶松叶片在10月份会变黄并逐渐脱落,为减小叶片活性对数据的影响,选择2018年5—9月连续3个晴天(27日,28日,29日)的液流值,分析生长季不同时期华北落叶松树干平均液流速率(
\overline {J_{\rm{s}}} )的日进程。由图2可知,液流速率存在明显的日变化,在一天内呈现单峰或多峰的变化特征,且不同时期树干液流的启动和停止时间不尽相同。5月27日至29日,树干液流于早晨06:50—07:05之间启动,之后迅速增加,至10:15—11:45之间达到峰值后开始下降,直至夜晚22:05—23:00之间达到最低值。6月27日至29日树干液流的启动时间为06:45—07:25之间,达到峰值的时间为10:50—12:25,出现最低值的时间为21:45—23:35。7月液流开始启动和停止的时间分别为03:40—07:00和21:05—23:00;9月液流的启动时间和停止时间分别为07:05—08:20和20:45—23:10。但8月27—29日树干液流的日进程与其他月份不同,存在夜间液流的现象。由于夜间气孔关闭,蒸腾作用停止,此时液流活动主要用于补充白天因强烈的蒸腾而导致的树体内的储水量减少的那部分水分[23-24]。同时,受不同时期土壤水分条件和气候条件的影响,树干液流峰值的大小从5—9月呈逐渐变小的趋势。![]() 图 2 典型晴天树干液流速率的日进程Figure 2. Diurnal variations of sap flux density (
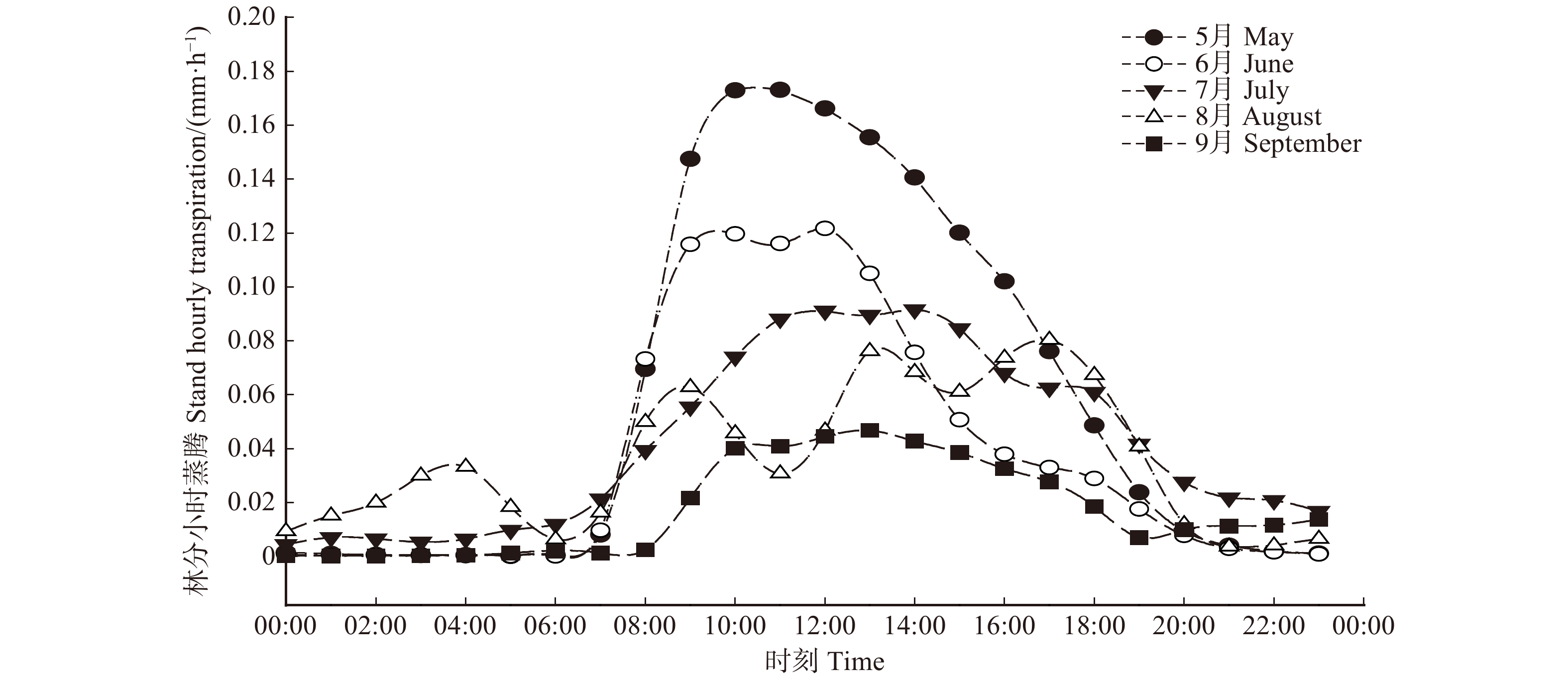
图 2 典型晴天树干液流速率的日进程Figure 2. Diurnal variations of sap flux density (\scriptstyle \overline {J_{\rm{s}}} ) in typical sunny days利用公式6计算得到林分小时蒸腾,将每月连续3天相同时刻的林分小时蒸腾进行平均,结果见图3。由图3可知,林分小时蒸腾的日内变化规律与树干液流速率基本相同,均呈单峰或者多峰的变化。尽管5—8月,林分蒸腾均从早上07:00开始出现,但生长季初期(5月和6月)林分蒸腾增加到最大值的速度要快于其他月份,而9月份林分蒸腾在早上8:00出现,要晚于其他月份。林分小时蒸腾量的峰值(mm/h)表现为5月(0.173)> 6月(0.122)> 7月(0.916)> 8月(0.762)> 9月(0.468)。
![]() 图 3 不同月份内林分小时蒸腾的日内变化特征Figure 3. Diurnal variations of stand hourly transpiration (T) in different months
图 3 不同月份内林分小时蒸腾的日内变化特征Figure 3. Diurnal variations of stand hourly transpiration (T) in different months3.3 林分小时蒸腾对单因素环境条件的响应
在小时尺度上,林分蒸腾的变化主要受VPD[25]、Rs[25]和REW的影响,因此,本文选取Rs、VPD和REW为主要影响因子,基于上外包线分析2018年生长季奇数天林分蒸腾量与Rs、VPD和REW的关系,逐个确定其响应函数。
图4为林分小时蒸腾(T)与Rs的外包线关系,由图可知,林分小时蒸腾对Rs的响应呈二次函数关系,可表示为:
![]() 图 4 林分小时蒸腾对太阳辐射的响应Figure 4. Response of stand hourly transpiration (T) to solar radiation
图 4 林分小时蒸腾对太阳辐射的响应Figure 4. Response of stand hourly transpiration (T) to solar radiation\begin{aligned} & T = - 3.000 \; 0 \times {10^{ - 7}}{R_{\rm{s}}}^2 + 0.000 \; 4{R_{\rm{s}}} + 0.065 \; 4, \\ & {{R^2} = 0.98} \end{aligned} (10) 当Rs较小时,林分蒸腾随Rs的增大而增大,当Rs达到666.7 W/m2后,林分蒸腾随Rs的继续增大而减小。这是因为当Rs较小时,随着Rs的增大,气孔因受到光照的诱导作用而打开;同时空气温度升高,叶面温度也随之升高,为植物的蒸腾提供了能量[26]。而当Rs继续增大时,气孔会因为温度过高而关闭,使得蒸腾量减小。
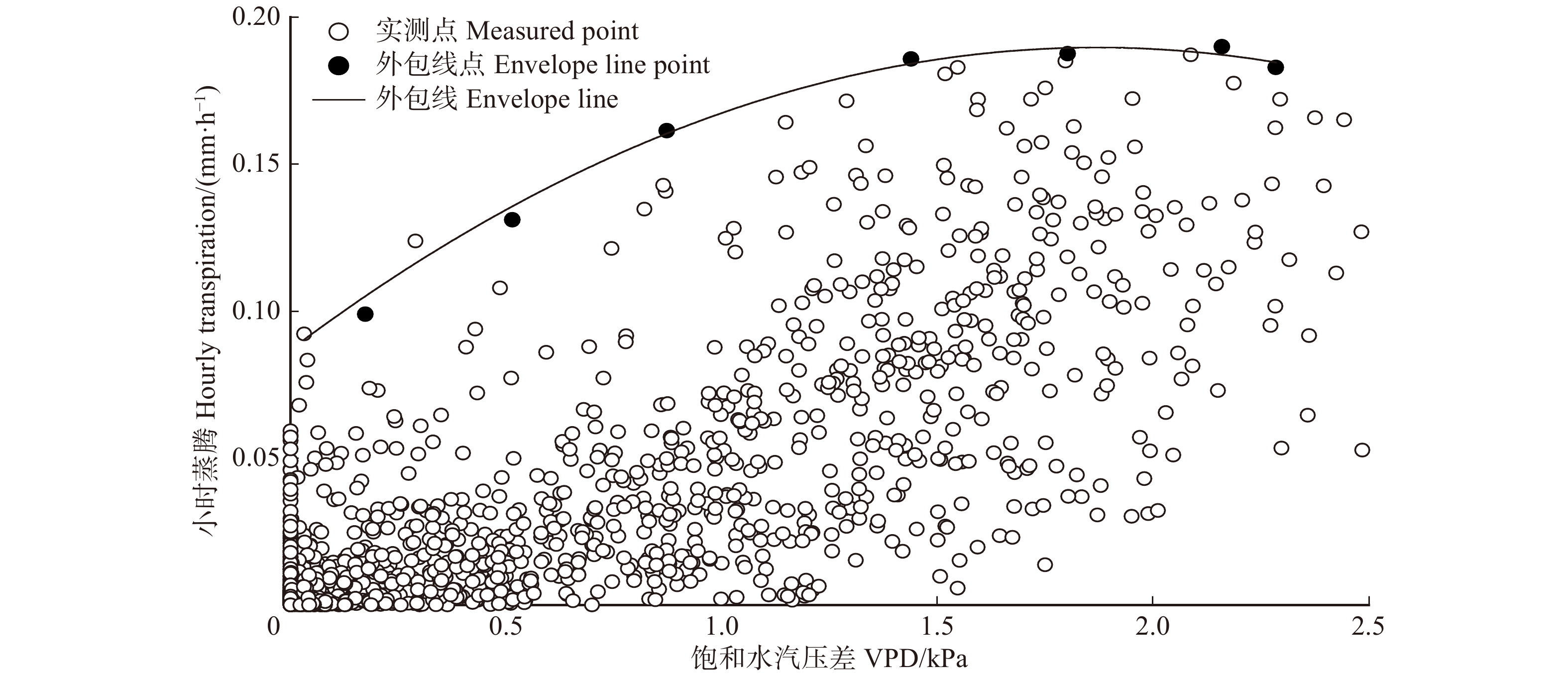
图5为林分小时蒸腾与VPD的外包线关系,由图可知,林分小时蒸腾与VPD的响应呈二次函数关系,其外包线函数关系为:
![]() 图 5 林分小时蒸腾对饱和水汽压差的响应Figure 5. Response of stand hourly transpiration (T) to vapor pressure deficit
图 5 林分小时蒸腾对饱和水汽压差的响应Figure 5. Response of stand hourly transpiration (T) to vapor pressure deficit\begin{aligned} & T = - 0.029 \; 3{\rm{VP}}{{\rm{D}}^2} + 0.109 \; 7{\rm{VPD}} + 0.087 ,\\ & {{R^2} = 0.98} \end{aligned} (11) 当VPD小于1.86 kPa时,林分蒸腾随VPD的增大而增大,当VPD大于1.86 kPa后,林分蒸腾随VPD的增大而减小。这是因为VPD过高时,植物将缩小气孔开度,减少水分过渡耗失,防止叶片水势过低而造成木质部出现灾变性的功能失衡[27],但这将使气孔导度下降,蒸腾减小。
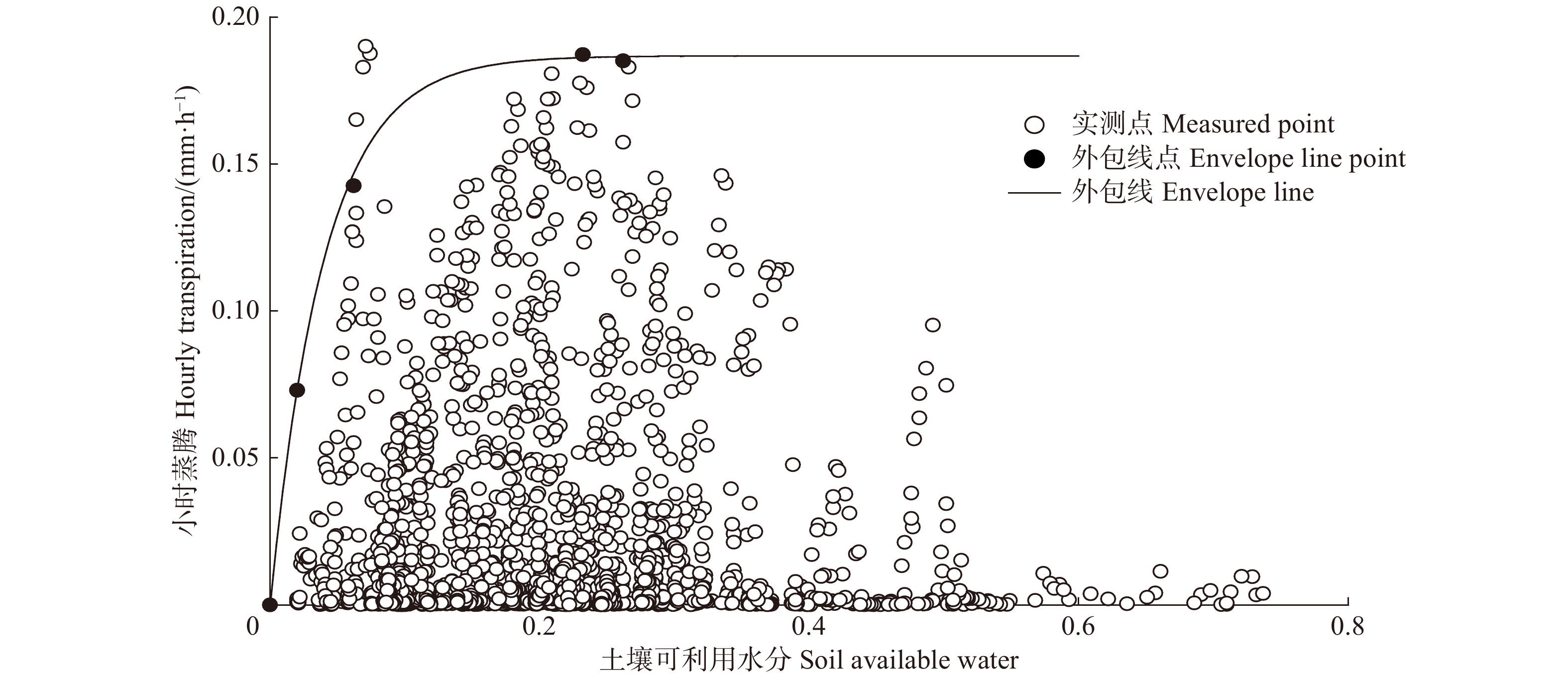
林分小时蒸腾与REW的外包线关系如图6所示。可以看到,林分小时蒸腾与REW的关系符合趋于饱和的指数方程,其外包线关系为:
![]() 图 6 林分小时蒸腾对土壤可利用水分的响应Figure 6. Response of stand hourly transpiration (T) to relative extractable water
图 6 林分小时蒸腾对土壤可利用水分的响应Figure 6. Response of stand hourly transpiration (T) to relative extractable water\!\begin{aligned} T = & - 0.000 \; 2 + 0.187 \; 0 \times \left( {1 - {\rm{exp}}\left( { - 26.875 \; 0{\rm{REW}}} \right)} \right),\\ & {{R^2} = 0.99} \end{aligned}\!\!\!\!\!\!\!\!\!\!\! (12) 林分蒸腾先随REW的增大而快速增加,当REW增加至0.3后,林分蒸腾增加的速度减缓并逐渐趋于稳定。这是因为当REW较小时,叶片水势低,水分由根系向上传递的驱动力弱,水分传输受阻;而随着REW的增加,树木可以吸收的水分和叶片水势增加,蒸腾速率也随之增加[28];当REW增加到某一阈值时,蒸腾量逐渐趋于稳定。
3.4 林分小时蒸腾对多因素的响应及耦合模型
为反映Rs、VPD和REW对林分小时蒸腾的综合影响,基于公式10 ~ 12的函数形式和连乘方程(公式8),得到样地小时蒸腾对Rs、VPD和REW响应的耦合模型基本形式:
T = f\left( {{R_{\rm{s}}}} \right) f\left( {\rm{VPD}} \right) f\left( {\rm{REW}} \right) (13) 进一步利用2018年生长季奇数天的实测数据进行重新拟合模型参数。得到模型表达式为:
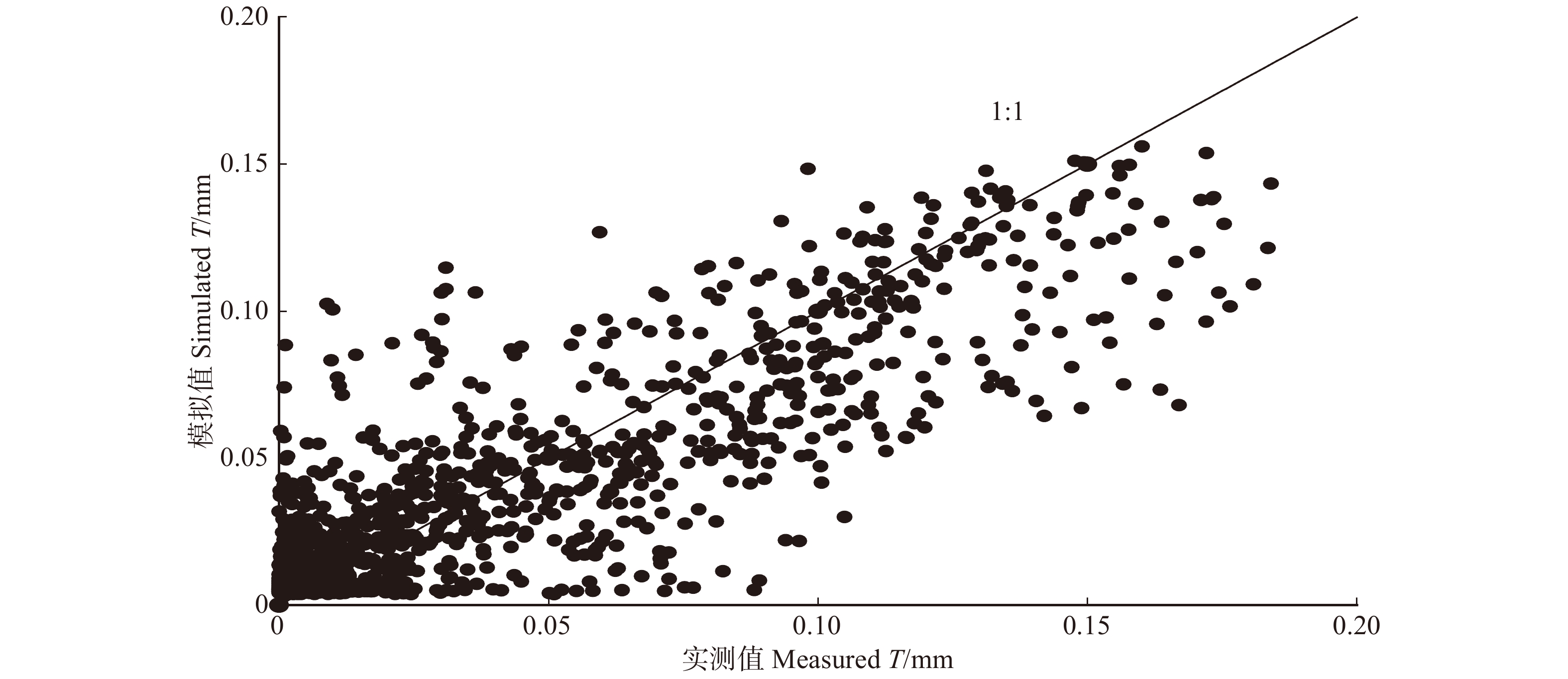
\begin{aligned} T = &\left( { - 6.347 \; 0 \times {{10}^{ - 5}}{R_{\rm{s}}^2} - 0.637 \; 0{R_{\rm{s}}} - 208.734 \; 8} \right) \times \\ &\left( { - 0.003 \; 2{\rm{VP}}{{\rm{D}}^2} + 0.013 \; 8{\rm{VPD}} + 0.001 \; 7} \right) \times \\ & \left( { - 0.008 \; 1 - 0.004 \; 6\left( {1 - {\rm{exp}}\left( { - 12.469 \; 6{\rm{REW}}} \right)} \right)} \right),\\ & {R^2} = 0.74 \end{aligned} (14) 利用该模型(公式14)得到的模拟值(2018年奇数天)与实测值具有较高的吻合性,(R2 = 0.74,NSE = 0.82)。利用未参与模型拟合的偶数天数据对模型进行验证发现(图7),模拟值与实测值的吻合较好(R2 = 0.77,NSE = 0.84),表明该模型能较好的模拟小时尺度林分蒸腾对环境变化的响应。
![]() 图 7 小时尺度林分蒸腾实测值与模拟值对比Figure 7. Comparison between the measured and simulated stand hourly transpiration (T)
图 7 小时尺度林分蒸腾实测值与模拟值对比Figure 7. Comparison between the measured and simulated stand hourly transpiration (T)4. 讨 论
4.1 林分小时蒸腾对单因素环境条件的响应
在小时尺度上,林木/林分蒸腾对Rs和VPD的响应最为敏感,Rs是调节气孔活动的主要环境因子,而VPD是林分蒸腾的主要驱动力[29]。研究表明,蒸腾速率与Rs,VPD显著相关[30-33]。VPD表示空气的干燥程度[34],当VPD较低时,气孔导度与VPD呈正相关;当VPD上升到一定阈值后,二者呈负相关。气孔对VPD的这种响应机制增加了从土壤到叶片这条水流通路上的导水阻力,保证了树木自身用水安全[32]。林分蒸腾响应VPD阈值的大小因树种而异。韩路等[35]对塔里木盆地灰胡杨(Populus pruinosa)的研究发现,蒸腾速率对VPD的响应阈值为2.92 kPa;贾国栋等[36]发现北方土石山区油松(Pinus tabuliformis)和刺槐(Robinia pseudoacacia)林分蒸腾在VPD达到1.1 kPa后降低。Fletcher等[37]发现VPD阈值受到植物耐旱程度的影响,一般在0.8 ~ 2.0 kPa。李新宇等[38]对银杏的蒸腾耗水研究表明,当VPD < 3.2 kPa时,VPD对液流是驱动作用,而VPD > 3.2 kPa时转变为限制作用。本研究中,当VPD于1.86 kPa后,林分蒸腾开始减小,该响应阈值在Fletcher等的研究结果范围之内。Rs影响气温和相对湿度,而VPD是温度和相对湿度的函数,因此,Rs的改变会引起VPD的改变。强太阳辐射使气温升高,空气相对湿度下降而导致VPD增大,引起气孔导度下降从而使林分蒸腾降低,因此,林分蒸腾对Rs的响应也存在一个阈值。朱丽薇等[39]研究发现当VPD大于2 kPa,同时光合有效辐射大于1 000 μmol/ (m2·s)时,荷木冠层气孔导度与水汽压亏缺呈显著负相关,说明辐射与水汽压亏缺共同决定气孔开闭。因此,Rs与VPD对蒸腾的影响是相互作用的。
除气象条件外,土壤水分也是影响树木蒸腾的主要因子。研究发现,樟子松在土壤水分偏低时,对Rs和VPD的敏感程度会降低,此时,REW成为主要的限制因子[40]。REW表征土壤供水能力的大小,林木蒸腾的水分主要来自于植物根系从土壤中吸取的水分,因此,植物是否受到干旱胁迫主要是由土壤水分的可利用性决定,并不完全取决于降雨条件[32]。土壤水分不足时,气孔为了降低干旱造成的风险会关闭[41],导致液流速率下降,蒸腾量降低。Orlandini等[42]通过研究盆栽葡萄(Vitis vinifera)对土壤干旱的响应发现,当REW降低至0.35时,蒸腾速率开始降低。Lagergren 和Lindroth[43]在瑞典中部的针叶混交林内研究发现,树木蒸腾出现明显降低的REW阈值在0.2左右。当土壤水分充足时,叶片水势增加,蒸腾速率表现为先增加后逐渐趋于稳定的变化趋势[12]。有研究表明,林分蒸腾达到稳定时的REW的阈值为0.2 ~ 0.5之间[44]。本研究中,当REW的阈值大于0.3 时,林分小时蒸腾基本保持稳定,该阈值在前人研究的阈值范围之内。
4.2 多因素共同影响下的树木蒸腾
在自然条件下,树木蒸腾受大气蒸腾需求和土壤水分可获得性的共同限制。因此,精确量化各环境因子对蒸腾的影响,并建立多因素影响的林分蒸腾模型非常有必要。利用上外包线法和连乘方程构建多因素对林分蒸腾影响的复合模型,可较好地模拟林分蒸腾[12,44]。但目前研究均集中在日尺度上,对短时间尺度(如小时尺度)上树木蒸腾对多个环境的响应关系及耦合模型还未见报道,这限制了对树木蒸腾响应短时间环境变化的准确理解。本研究在小时尺度上建立了同时反映Rs,VPD和REW影响的华北落叶松林分蒸腾模型,具有较高的拟合精度,可解释小时蒸腾量变异的74%,可为更进一步理解华北落叶松蒸腾对小时尺度上环境变化的响应提供帮助。但本研究建立的耦合模型并未考虑短时间尺度下树干液流对Rs和VPD的响应的时滞效应问题。Zeppel等[45]对澳大利亚桉树(Eucalyptus crebra)和白羽松(Callitris glaucophylla)的研究发现,树干液流对VPD的响应关系会出现顺时针方向的时滞圈;Hong等[6]发现,树干液流速率对Rs的响应在晴天表现为顺时针时滞,而在阴天和雨天表现为逆时针时滞。另外,本研究也未考虑叶面积指数的变化对蒸腾量的影响。已有研究表明,同等辐射强度下,叶面积的大小决定着吸收能量的多少,因叶面积不同导致蒸腾存在差异[46],这均有可能影响本研究构建模型的模拟精度(74%),因此,未来还需继续考虑时滞效应与叶面积指数变化的影响,以提高模型模拟精度。
5. 结 论
(1)林分蒸腾的日内变化呈现单峰或多峰的变化特征,蒸腾量的峰值(mm/h)表现为5月(0.173)> 6月(0.122)> 7月(0.916)> 8月(0.762)> 9月(0.468)。
(2)林分小时蒸腾量对Rs和VPD的响应均符合二次多项式函数,随Rs和VPD的增大,蒸腾量先增后减,达到最高值的Rs和VPD阈值分别为666.7 W/m2和1.86 kPa;而林分小时蒸腾对REW的响应符合趋于饱和的指数关系,随REW的增加,蒸腾量先增加,当REW大于0.3后逐渐趋于稳定。
(3)林分小时蒸腾可用耦合Rs、VPD和REW这3个因素影响的模型进行很好地预测和评估,模型为T = (− 6.347 0 × 10− 5
{R_{\rm{s}}^2} − 0.637 0Rs − 208.734 8) × (− 0.003 2VPD2 + 0.013 8VPD + 0.001 7) × (− 0.008 1 − 0.004 6(1 − exp(− 12.469 6REW)))。 -
![]()
图 1 3种藤本植物1 ~ 5级根典型解剖构造
Figure 1. Typical anatomical structure of the grade 1−5roots in three liana species
![]()
图 2 3种藤本植物1 ~ 5级细根皮层厚度、中柱直径及二者比值的变化
大写字母表示不同树种间差异显著;小写字母表示不同根序间差异显著。下同。Capital letters represent significant differences among different tree species; lowercase letters represent significant differences among varied root orders. The same below
Figure 2. Variations of cortex thickness, stele diameter and their ratios in the grade 1−5 roots in three liana species
![]()
图 3 3种藤本植物1 ~ 5级细根直径、比根长、组织密度和分支比的变化
Figure 3. Variations of the diameter, specific root length, tissue density and branching ratio in grade 1−5 roots in three liana species
表 1 3种藤本植物1 ~ 5级根吸收根的比例
Table 1 Proportion of absorptive fine roots of grade 1−5 in three liana species
% 树种 Tree species 1级根 Grade 1 root 2级根 Grade 2 root 3级根 Grade 3 root 4级根 Grade 4 root 5级根 Grade 5 root 狗枣猕猴桃 Actinidia kolomikta 100 96.43 91.80 45.45 2.94 北五味子 Schisandra chinensis 100 97.22 34.55 6.82 0 山葡萄 Vitis amurensis 100 86.76 7.55 0 0  下载: 导出CSV
下载: 导出CSV
表 2 树种和根序对细根形态、解剖以及分支比的双因素方差分析
Table 2 Two-way (species and root order) factorial ANOVA of tree spscies and root order on root morphological, anatomical characteristics and branching ratios
变异来源
Source of
variation自由度
df皮层厚度
Cortex thickness中柱直径
Stele diameter皮层厚度/中柱直径
Cortex thickness/
stele diameter根直径
Root diameter比根长
Specific root length组织密度
Tissue density自由度
df分支比
Branching ratio树种 Tree species 2 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 0.001 2 < 0.001 根序 Root order 4 0.057 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 3 < 0.001 树种 × 根序
Tree species × root order7 < 0.001 < 0.001 < 0.001 0.004 < 0.001 < 0.001 5 0.001 注:粗体字表示影响显著;山葡萄、北五味子的4、5级根的大部分皮层组织解体,故只对前3级根的皮层厚度、皮层厚度/中柱直径进行了统计分析。Notes: values in bold type indicate significant effects. Only the cortical thickness and cortex thickness/stele diameter of the first three grade roots are analyzed by ANOVA due to most cortex tissue disorganizes in grade 4 and grade 5 roots in Schisandra chinensis and Vitis amurensis.
下载: 导出CSV
表 3 帽儿山地区木质藤本植物与其他植物类群1级根形态特征比较
Table 3 Comparison in grade 1 root morphological characteristics between lianas and other plant taxa in Maoershan Region of northeastern China
生活型
Life form样本量
Sample size平均根直径
Average root diameter/μm根直径范围
Rang of root diameter/μm平均比根长
Average specific root length/(m·g− 1)比根长范围
Rang of specific root length/(m·g− 1)平均组织密度
Average tissue density/(g·cm− 3)组织密度范围
Rang of tissue density/(g·cm− 3)文献来源
Literature source木质藤本
Woody liana3 274.34 ± 20.55 234.30 ~ 327.20 91.52 ± 19.94 42.79 ~ 153.35 0.20 ± 0.02 0.14 ~ 0.31 本研究
This study阔叶树
Broadleaf tree20 240 170 ~ 500 102.9 32.71 ~ 203.83 − − 师伟等, 2008
Shi Wei, etc, 2008阔叶树和针叶树
Broadleaf tree and coniferous tree23 243 168 ~ 520 99.83 33.7 ~ 203.82 0.27 0.14 ~ 0.41 Gu et al, 2014 草本蕨类
Herbaceous fern9 − 187 ~ 468 − 85 ~ 260 − 0.04 ~ 0.12 Dong et al, 2015 注:“−”表示数据未给出。Note: “−” means data are not available.
下载: 导出CSV
-
[1] 刘奇, 吴怀栋, 谭运洪, 等. 西双版纳热带季雨林木质藤本多样性及其攀援方式[J]. 林业科学, 2017, 53(8):1−8. doi: 10.11707/j.1001-7488.20170801 Liu Q, Wu H D, Tan Y H, et al. Liana diversity and its climbing situation on trees in Xishuangbanna tropical seasonal rainforest[J]. Scientia Silvae Sinicae, 2017, 53(8): 1−8. doi: 10.11707/j.1001-7488.20170801
[2] Leicht-Young S A, Pavlovic N B, Frohnapple K J, et al. Liana habitat and host preferences in northern temperate forests[J]. Forest Ecology and Management, 2010, 260(9): 1467−1477. doi: 10.1016/j.foreco.2010.07.045
[3] Pérez-Salicrup D R, Schnitzer S, Putz F E. Community ecology and management of lianas[J]. Forest Ecology and Management, 2004, 190(1): 1−2. doi: 10.1016/j.foreco.2003.10.001
[4] Schnitzer S A, Bongers F. The ecology of lianas and their role in forests[J]. Trends in Ecology & Evolution, 2002, 17(5): 223−230.
[5] 胡亮. 东亚温带藤本植物多样性及其格局[J]. 生物多样性, 2011, 19(5):567−573. doi: 10.3724/SP.J.1003.2011.07064 Hu L. Distribution and diversity of climbing plants in temperate East Asia[J]. Biodiversity Science, 2011, 19(5): 567−573. doi: 10.3724/SP.J.1003.2011.07064
[6] Allen B P, Sharitz R R, Goebel P C. Are lianas increasing in importance in temperate floodplain forests in the southeastern United States?[J]. Forest Ecology and Management, 2007, 242(1): 17−23. doi: 10.1016/j.foreco.2007.01.027
[7] Ladwig L M, Meiner S J. Liana host preference and implications for deciduous forest regeneration[J]. Journal of the Torrey Botanical Society, 2010, 137(1): 103−112. doi: 10.3159/09-RA-041.1
[8] Allen B P, Sharitz R R, Goebel P C. Twelve years post-hurricane liana dynamics in an old-growth southeastern floodplain forest[J]. Forest Ecology and Management, 2005, 218(1−3): 259−269. doi: 10.1016/j.foreco.2005.08.021
[9] Vivek P, Parthasarathy N. Contrasting leaf-trait strategies in dominant liana and tree species of Indian tropical dry evergreen forest[J]. Flora, 2018, 249: 143−149. doi: 10.1016/j.flora.2018.11.002
[10] 丁凌子, 陈亚军, 张教林. 热带雨林木质藤本植物叶片性状及其关联[J]. 植物科学学报, 2014, 32(4):362−370. Ding L Z, Chen Y J, Zhang J L. Leaf traits and their associations among liana species in tropical rainforest[J]. Plant Science Journal, 2014, 32(4): 362−370.
[11] Freschet G T, Valverde-Barrantes O J, Tucker C M, et al. Climate, soil and plant functional types as drivers of global fine-root trait variation[J]. Journal of Ecology, 2017, 105(5): 1182−1196. doi: 10.1111/1365-2745.12769
[12] Ma Z Q, Guo D L, Xu X L, et al. Evolutionary history resolves global organization of root functional traits[J]. Nature, 2018, 555: 94−97. doi: 10.1038/nature25783
[13] Collins C G, Wright S J, Wurzburger N. Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees[J]. Oecologia, 2016, 180(4): 1037−1047. doi: 10.1007/s00442-015-3410-7
[14] Pregitzer K S, Deforest J L, Burton A J, et al. Fine root architecture of nine north American trees[J]. Ecological Monographs, 2002, 72(2): 293−309. doi: 10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2
[15] 纪春艳, 崔大练, 马玉心. 东北地区野生藤本植物区系分析[J]. 中国林副特产, 2007(1):59−63. doi: 10.3969/j.issn.1001-6902.2007.01.029 Ji C Y, Cui D L, Ma Y X. Biotic zonation of the wild climber plants in northeast China[J]. Forest By-product and Speciality in China, 2007(1): 59−63. doi: 10.3969/j.issn.1001-6902.2007.01.029
[16] 王向荣, 王政权, 韩有志, 等. 水曲柳和落叶松不同根序之间细根直径的变异研究[J]. 植物生态学报, 2005, 29(6):871−877. doi: 10.3321/j.issn:1005-264X.2005.06.001 Wang X R, Wang Z Q, Han Y Z, et al. Variations of fine root diameter with root order in Manchurian ash and Dahurian larch plantations[J]. Acta Phytoecologica Sinica, 2005, 29(6): 871−877. doi: 10.3321/j.issn:1005-264X.2005.06.001
[17] 卫星, 刘颖, 陈海波. 黄波罗不同根序的解剖结构及其功能异质性[J]. 植物生态学报, 2008, 32(6):1238−1247. doi: 10.3773/j.issn.1005-264x.2008.06.004 Wei X, Liu Y, Chen H B. Anatomical and functional heterogeneity among different root orders of Phellodendron amurense[J]. Acta Phytoecologica Sinica, 2008, 32(6): 1238−1247. doi: 10.3773/j.issn.1005-264x.2008.06.004
[18] Gu J C, Xu Y, Dong X Y, et al. Root diameter variations explained by anatomy and phylogeny of 50 tropical and temperate tree species[J]. Tree Physiology, 2014, 34(4): 415−425. doi: 10.1093/treephys/tpu019
[19] Bardgett R D, Mommer L, De Vries F T. Going underground: root traits as drivers of ecosystem processes[J]. Trends in Ecology & Evolution, 2014, 29(12): 692−699.
[20] 刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学: 生命科学, 2015, 45(4):325−339. doi: 10.1360/N052014-00244 Liu X J, Ma K P. Plant functional traits concepts, applications and future directions[J]. Scientia Sinica Vitae, 2015, 45(4): 325−339. doi: 10.1360/N052014-00244
[21] Guo D L, Xia M X, Wei X, et al. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J]. New Phytologist, 2008, 180(3): 673−683. doi: 10.1111/j.1469-8137.2008.02573.x
[22] Dong X Y, Wang H F, Gu J C, et al. Root morphology, histology and chemistry of nine fern species (Pteridophyta) in a temperate forest[J]. Plant and Soil, 2015, 393(1−2): 215−227. doi: 10.1007/s11104-015-2484-7
[23] Long Y Q, Kong D L, Chen Z X, et al. Variation of the linkage of root function with root branch order[J/OL]. PLoS One, 2013, 8(2): e57153 (2013−02−25) [2017−04−18]. https//doi.org/10.1371/journal.pone.0057153.
[24] Kong D L, Wang J J, Zeng H, et al. The nutrient absorption-transportation hypothesis: optimizing structural traits in absorptive roots[J]. New Phytologist, 2017, 213(4): 1569−1572. doi: 10.1111/nph.14344
[25] 师伟, 王政权, 郭大立, 等. 帽儿山天然次生林20个阔叶树种细根形态[J]. 植物生态学报, 2008, 32(6):1217−1226. doi: 10.3773/j.issn.1005-264x.2008.06.002 Shi W, Wang Z Q, Guo D L, et al. Fine root morphology of twenty hardwood species in Maoershan natural secondary forest in northeastern China[J]. Acta Phytoecologica Sinica, 2008, 32(6): 1217−1226. doi: 10.3773/j.issn.1005-264x.2008.06.002
[26] Wang Z Q, Guo D L, Wang X R, et al. Fine root architecture, morphology, and biomass of different branch orders of two Chinese temperate tree species[J]. Plant and Soil, 2006, 288(1−2): 155−171. doi: 10.1007/s11104-006-9101-8
[27] 常文静, 郭大立. 中国温带、亚热带和热带森林45个常见树种细根直径变异[J]. 植物生态学报, 2008, 32(6):1248−1257. doi: 10.3773/j.issn.1005-264x.2008.06.005 Chang W J, Guo D L. Variation in root diameter among 45 common tree species in temperate, subtropical and tropical forests in China[J]. Acta Phytoecologica Sinica, 2008, 32(6): 1248−1257. doi: 10.3773/j.issn.1005-264x.2008.06.005
[28] Kou L, Guo D L, Yang H, et al. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China[J]. Plant and Soil, 2015, 391(1−2): 207−218. doi: 10.1007/s11104-015-2420-x
[29] 廖乐平. 亚热带木兰科、樟科10个树种细根形态与功能特性研究及其亲缘关系分析[D]. 长沙: 中南林业科技大学, 2016. Liao L P. Fine root morphology and functional characteristics of 10 species of Magnoliaceae and Camphoraceae in subtropics and their relationship analysis[D]. Changsha: Central South University of Forestry and Technology, 2016.
[30] 许旸, 谷加存, 董雪云, 等. 海南岛4个热带阔叶树种前5级细根的形态、解剖结构和组织碳氮含量[J]. 植物生态学报, 2011, 35(9):955−964. Xu Y, Gu J C, Dong X Y, et al. Fine root morphology, anatomy and tissue nitrogen and carbon contents of the first five orders in four tropical hardwood species in Hainan Island, China[J]. Acta Phytoecologica Sinica, 2011, 35(9): 955−964.
[31] Woods C L, Dewalt S J, Cardelús C L, et al. Fertilization influences the nutrient acquisition strategy of a nomadic vine in a lowland tropical forest understory[J]. Plant and Soil, 2018, 431(1−2): 389−399. doi: 10.1007/s11104-018-3772-9
-
期刊类型引用(3)
1. 蔡家珍. 闽南乡村低山河流型湿地公园植物群落构建. 漳州职业技术学院学报. 2024(04): 103-108 .  百度学术
百度学术
2. 郭雅婷,李运远. 基于生物多样性的城市绿地植物群落构建途径. 风景园林. 2022(01): 59-63 . 百度学术
3. 杨土士,王伟文,常鸣,王雪梅. 北京市潜在风道的数值模拟与综合识别. 地球信息科学学报. 2020(10): 1996-2009 . 百度学术
其他类型引用(3)
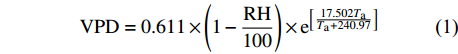
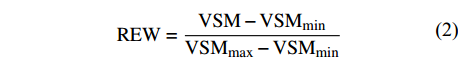
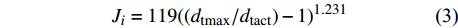
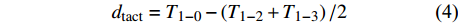
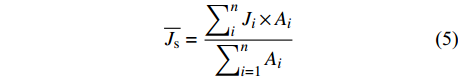
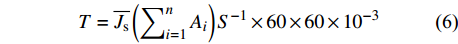
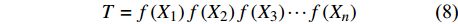
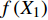
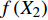
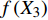
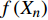
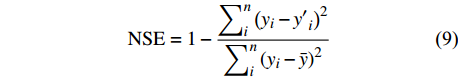
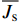

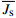

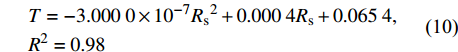
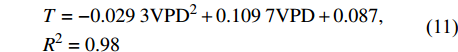
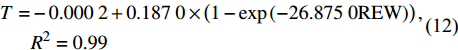
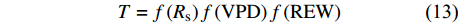




计量
- 文章访问数: 2330
- HTML全文浏览量: 730
- PDF下载量: 103
- 被引次数: 6
