
Reintroducing effects and influencing factors of Taxus cuspidata population
-
摘要:目的 回归是一种濒危物种保护与种群恢复的重要方法。本研究以极小种群物种东北红豆杉为研究对象,选择3种不同的林型以及两组不同苗龄的东北红豆杉幼苗开展野外回归试验,研究适宜东北红豆杉回归林型以及回归过程中幼苗生长及存活的主要影响因素,以期为东北红豆杉回归保护提供科学依据。方法 选择杨桦林、红松云冷杉林和红松紫椴林3种林型作为回归试验地,以1 ~ 2年生和4 ~ 5年生东北红豆杉实生幼苗作为试验材料,移栽后调查土壤因子、地形因子等指标,逐月调查幼苗存活率、苗高、地径、冠幅等指标。对幼苗各生长指标进行差异性显著分析和多重比较,通过灰色关联分析等方法研究回归幼苗生长过程中的影响因子。结果 东北红豆杉回归幼苗当年存活率达86%以上,地径、苗高、冠幅均有所增长,除1 ~ 2年生东北红豆杉幼苗苗高生长量外,各林型中东北红豆杉幼苗生长指标生长量均无显著差异,红松紫椴林的1 ~ 2年生东北红豆杉幼苗苗高生长量显著高于杨桦林和红松云冷杉林。由于越冬期间,4 ~ 5年生东北红豆杉幼苗受到狍啃食,存活率显著下降且长势极差;1 ~ 2年生幼苗受长时间的低温胁迫生长状况也受到一定影响。东北红豆杉回归幼苗的存活和生长受到多种环境因子的影响,关联度最大的是林型,其次是坡向、坡度和郁闭度,土壤化学性质指标的关联度较小。结论 4 ~ 5年生幼苗因受动物啃食而回归效果不良;1 ~ 2年生幼苗适应杨桦林、红松云冷杉林和红松紫椴林下这3种生境,回归最适宜的林型是红松紫椴林;林型、坡向、坡度、光照是影响东北红豆杉回归的主要环境因子。Abstract:Objective Reintroduction is an important method for endangered species conservation and population restoration. Taking Taxus cuspidata, a critically endangered wild plant with extremely small population, as the research object, this study chose three different forest types and two groups of different age T. cuspidata seedlings for field reintroduction tests, studied the reintroducted forest type suitable for T. cuspidate and the main factors influencing seedling growth and survival during reintroduction process. It is hopefully to provide scientific evidence for the conservation of T. cuspidata.Method Betulaplatyphylla-Populus cathayana forest, Pinus koraiensis-spruce-fir forest and P. koraiensis-Tilia amurensis forest were selected as the reintroduction experimental forest types, indexes such as soil factors and topographic factors were investigated, and indexes such as seedling livability, seedling height, basal diameter and crown breadth were investigated monthly. Difference significance analysis and multiple comparison were conducted for each growth index of seedlings, and the influencing factors of reintroducted seedling growth were studied through grey correlation analysis and other methods.Result The very year survival rate of reintroducted seedlings of T. cuspidata was more than 86%. The ground diameter, seedling height and crown width all increased, except for the seedling height growth of T. cuspidata in 1−2 years, the growth indexes of T. cuspidata seedlings of all forest types showed no significant variations. The 1−2 years old T. cuspidata seedlings of P. koraiensis-T. amurensis forest were significantly higher than that of B. platyphylla-P. cathayana forest and P. koraiensis-spruce-fir forest. As it was during the overwintering period, 4−5 years old T. cuspidata seedlings were eaten by roe deer, their survival rate decreased significantly, and the growth rate was very poor; the growth of 1−2 years old seedlings was also affected to some extent by long-term low temperature stress. The survival and growth of reintroducted seedlings of T. cuspidata were affected by diversified environmental factors, among which, the most relevant was forest type, followed by slope aspect, slope degree and canopy density, while the association degree of soil chemical property index was less.Conclusion The effect of reintroduction of 4−5 years old T. cuspidata seedlings is poor due to predation of animals. 1−2 years old seedlings adapt to the three habitats of B. platyphylla-P. cathayana forest, P. koraiensis-spruce-fir forest and P. koraiensis-T. amurensis forest, and the optimum forest type for the seedling reintroduction of 1−2 years old is P. koraiensis-T. amurensis forest. Forest type, slope aspect, slope degree and light are the primary environmental factors influencing the reintroduction of T. cuspidata.
-
Keywords:
- Taxus cuspidata /
- reintroduction /
- forest type /
- seedling age /
- gray correlation analysis /
- influencing factor
-
近年来,人类活动和气候变化对生物多样性的丧失和物种灭绝的影响已经成为生态学家最关心的问题[1]。由于过度开采、分布区缩小、生境恶化等人类的干扰,加上气候变化的影响,全球植物种类正以空前的速度消失[2]。中国是生物多样性受威胁最严重的国家之一,根据数据统计,15% ~ 20%的植物种类受到威胁[3],濒危植物的保护刻不容缓。濒危植物保护主要包括就地保护、迁地保护和野外回归[4]。其中植物回归是珍稀濒危植物的保护及种群恢复的重要策略之一,其保护效果超出了单纯的就地保护和迁地保护,能更有效的对极小种群野生植物进行拯救和保护[5]。
植物的回归是基于迁地保护的基础上, 通过人工繁殖把植物引入到其原来分布的自然或半自然的生境中,以建立具有足够的遗传资源来适应进化改变,可自然维持和更新的新种群[6]。相关研究表明,影响回归成功的因子包括繁殖材料类型、生境等[4],其中适宜生境的选择是植物回归的核心问题[7]。Albrecht等[8]对黄芪(Astragalus propinquus)回归进行研究,结果表明,生境质量和脊椎动物啃食是濒危植物回归长期持续的关键驱动因素。Erin等[9]研究表明,回归植物的建立与生存和生境条件有关。汪越[10]在研究中发现,由于啮齿类动物的啃食导致紫背天葵增强回归失败。李景秀等[11]、谭美等[12]、冯欣欣等[13]、陈芳清等[14]均开展了濒危保护植物的回归试验,取得了一定的成果。在不同生境中建立回归试验种群,并对种群动态持续监测,是解决植物回归适宜生境选择问题最为可靠的途径[15]。
东北红豆杉(Taxus cuspidata)是第三纪孑遗乔木物种、国家I级重点保护植物、极小种群物种[4]。在我国,由于过度砍伐和紫杉醇的提炼与利用[16],东北红豆杉天然种群被人类活动严重干扰和破坏,导致种群稳定性不断下降,适宜分布区减少,其保护形势十分严峻。根据刘彤[17]、李云灵[18]的研究发现,东北红豆杉主要分布在红松(Pinus koraiensis)云(Picea spp.)冷杉(Abies spp.)林中,紫椴(Tilia amurensis)和东北红豆杉之间具有极显著的种间正关联性。周志强等[19]、陈杰等[20]、杨占等[21]对东北红豆杉适宜的坡向、坡度和海拔分布进行研究,认为东北红豆杉最适坡向为阴坡,适宜坡度在15° ~ 20°之间,适宜海拔分布范围在700 ~ 900 m之间。目前有关东北红豆杉群落结构、繁育技术、生境适宜性评价等方面的研究取得了一定的进展[22-25],基本具备了开展东北红豆杉野外回归的条件。现在关于适宜东北红豆杉种群回归林型的研究较少,在回归过程中幼苗生长受到哪些因素的影响也不得而知。本研究以杨(Populus cathayana)桦(Betula platyphylla)林、红松云冷杉林和红松紫椴林3种不同林型作为试验地,以1 ~ 2年生和4 ~ 5年生两种不同苗龄的东北红豆杉实生苗为材料,模拟东北红豆杉幼苗在不同苗龄阶段的生长过程,探究不同林型对东北红豆杉苗木回归存活率和生长指标的影响,以及影响东北红豆杉幼苗生长的主要环境因子,旨在为今后开展东北红豆杉的野外回归提供科学依据。
1. 研究概况和研究方法
1.1 研究区域概况
本研究的回归样地位于吉林省汪清县杜荒子林场,地理位置 130°36′E、43°12′N,属于中温带湿润温凉季风气候区,全年平均气温为5 ℃,极端最高气温37.5 ℃,极端最低气温− 44.1 ℃,年降水量为 450 ~ 750 mm。杜荒子林场大部分山体海拔在1 000 m以下,海拔700 m以下是以蒙古栎(Quercus mongolica)林、杨树和榆树(Ulmus spp.)等杂木林为主的阔叶林,海拔700 ~ 1 000 m是红松阔叶林为主的混交林带,在海拔 800 m以上是红松与云冷杉混交林为主 [26]。
1.2 试验材料
周志强等[19]在穆棱自然保护区发现处于更新层的东北红豆杉较多,演替层的幼苗较少。在杜荒子林场的野外调查过程中发现,在有记录的749株东北红豆杉中,苗高低于50 cm的东北红豆杉幼苗数量为42株,占5.6%。因此选择相同种源的1 ~ 2年生幼苗和4 ~ 5年生幼苗作为试验材料,研究东北红豆杉幼苗在更新层和演替层的生长特征及主要影响因子。
2018年5月从吉林省延边朝鲜族自治州汪清县购得种源相同的1 ~ 2年生东北红豆杉实生幼苗150株,4 ~ 5年生东北红豆杉实生幼苗150株,起苗时间为2018年5月15日。试验材料选择生长健壮、长势一致的幼苗。
1.3 研究方法
1.3.1 试验设计
2018年5月初将1 ~ 2年生东北红豆杉幼苗和4 ~ 5年生东北红豆杉幼苗移栽到3个不同林型的试验点。每个试验点各种植1 ~ 2年生东北红豆杉幼苗50株,株间距30 cm × 30 cm;4 ~ 5年生东北红豆杉幼苗50株,间距3 m × 3 m。
1.3.2 样地选择与设置
根据2017年天然东北红豆杉种群调查,我国东北红豆杉主要分布区内布设的21个20 m × 50 m的临时样方数据发现,有11个样方的林型为红松紫椴林,占52.38%,有6个样方的林型为红松云冷杉林,占28.57%,因此选择红松紫椴林和红松云冷杉林这两种林型作为回归林型。由于人为砍伐红松、云冷杉或紫椴后,该地区演替形成的次生林主要为杨桦林,因此选择在杨桦林这种次生生境中建立回归样地,并与红松云冷杉林、红松紫椴林这两种原生生境对比。
1.3.3 调查与测定方法
苗木栽植后,逐株挂牌编号,测量幼苗的苗高、地径、冠幅、最长侧枝长,统计死亡株数。测量时期为2018年5—9月以及2019年4月和5月,测量时间为每月20—25日。苗高、冠幅、最长侧枝长使用钢卷尺测量,地径使用游标卡尺测量。
冠幅面积=南北冠幅×东西冠幅/2 (1) 样地调查记录各试验样地的海拔、坡度、坡向、郁闭度。
土壤化学取样点采用对角线布设的方法,即在每块试验地对角线上平均布设3个取样点。在每个取样点采集0 ~ 20 cm土层的混合土样,每个土样质量约1 kg。测定指标包括全氮、全磷、有机质及pH值。全氮含量测定使用凯氏定氮法,全磷含量测定使用钼锑抗比色法,有机质含量测定使用重铬酸钾氧化外加热法,pH值测定使用pH酸度计法[27]。
1.4 数据处理及分析
使用Excel 2016整理数据及灰色关联分析,使用SPSS19.0软件中单因素方差分析(one-way ANOVA)模块下的Duncan法对土壤指标和幼苗生长指标进行差异性显著检验和多重分析,并计算生长指标与环境因子的关联系数,各指标增长量也均以2019年5月存活幼苗进行计算。幼苗存活曲线以每个月调查时各林型幼苗存活率进行绘制。作图用SigmaPlot13软件完成。
样地坡向方位角转换:将0 ~ 360°的罗盘测量方位角值转换成坡向指数值。转换式为
T={1−cos[(π/180)(θ−30∘)]}/2, (2) 式中:T为坡向指数;θ 为坡向方位角度转换后的T值,变化范围为0 ~ 1,值越大表示生境条件越趋干热[28]。
林型转化为定量变量:分别对红松云冷杉林、红松紫椴林和杨桦林赋值为1、2、3,数值越大表明阔叶树种所占比例越高,针叶树种所占比例越低。
灰色关联分析:以每个试验地内幼苗各项生长指标平均增长量(4 ~ 5年生幼苗以2018年9月数据计算,排除动物因素的影响,1 ~ 2年生幼苗以2019年5月数据计算)为比较数列,以3个试验地各项生长指标平均增长量之和为参考数列,对其进行无纲化处理后,计算参考数列和比较数列的灰色关联系数,得出关联度并排序。计算公式如下:
εi(k)=min (3) {r}_{i}=\frac{1}{n}\sum\limits _{k=1}^{n}{\mathrm{\varepsilon }}_{i}\left(k\right),\;\;k = 1,2,3, \cdots,n (4) 式中:
{\mathrm{\varepsilon }}_{i}\left(k\right) 为关联系数;k为灰色关联分析的指标;{C}_{0}(k) 为参考数列;{C}_{i}(k) 为比较数列;i为比较数列个数;\; \rho \in \left(\mathrm{0,1}\right) ,本试验中取\; \rho =0.5;n为关联系数的个数,各指标权重相等。2. 结果与分析
2.1 回归幼苗存活状况和生长特征
2.1.1 东北红豆杉幼苗存活状况
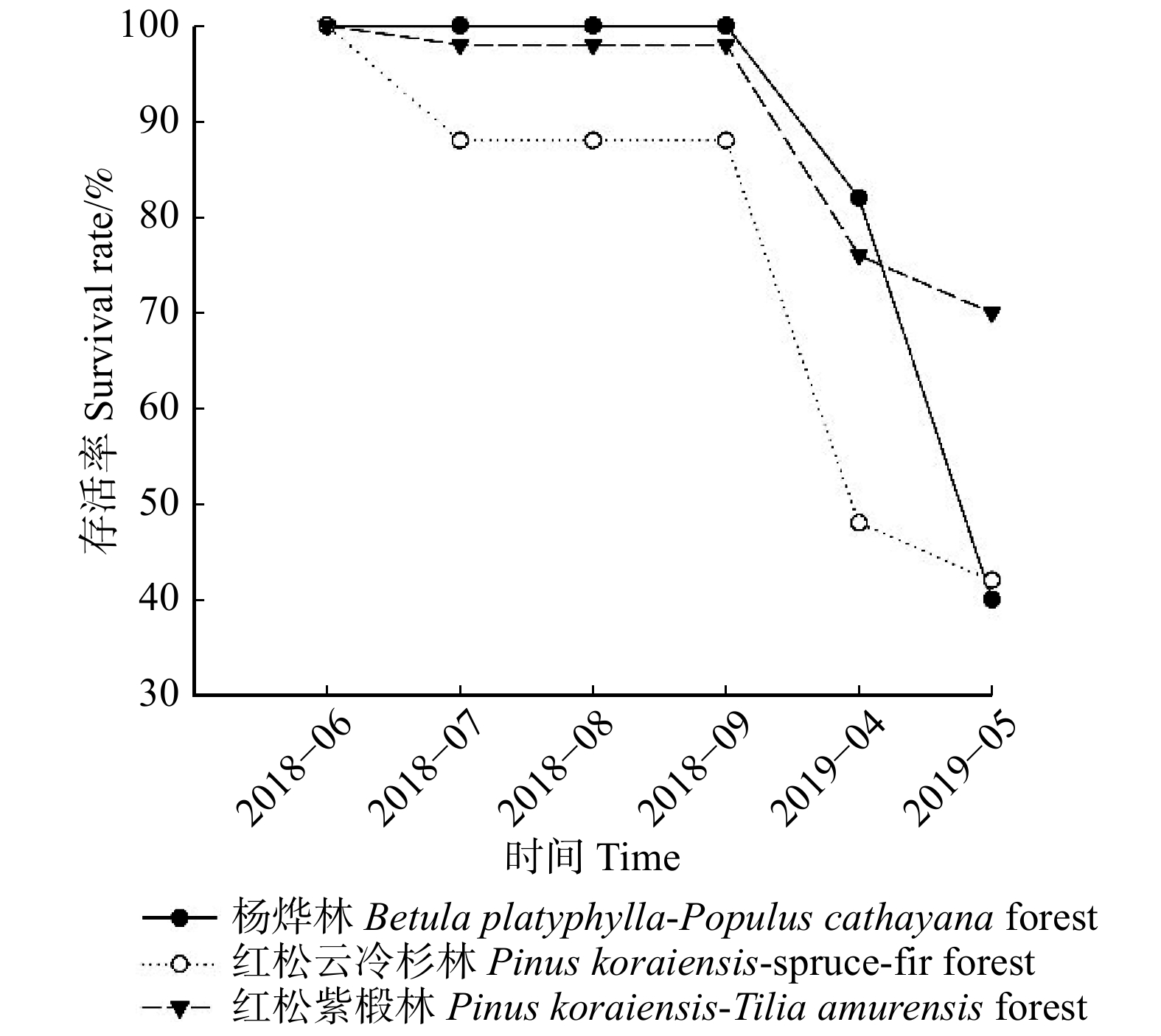
由各林型4 ~ 5年生幼苗存活曲线(图1)可知,4 ~ 5年生东北红豆杉幼苗,除红松云冷杉林幼苗在2018年6—7月期间有较多死亡外,剩下两个林型幼苗成活率在2018年6月至2018年9月期间变化幅度不大,红松云冷杉林在2018年9月至2019年5月期间成活率急剧下降,杨桦林和红松紫椴林幼苗成活率急剧下降的时期为2019年4月至5月。截止2019年5月,各试验样地存活率排序为红松紫椴林(70%) > 红松云冷杉林(42%) > 杨桦林(40%)。
![]() 图 1 各林型4 ~ 5年生幼苗存活曲线Figure 1. Survival rate curves of 4−5 years old seedlings for each forest type
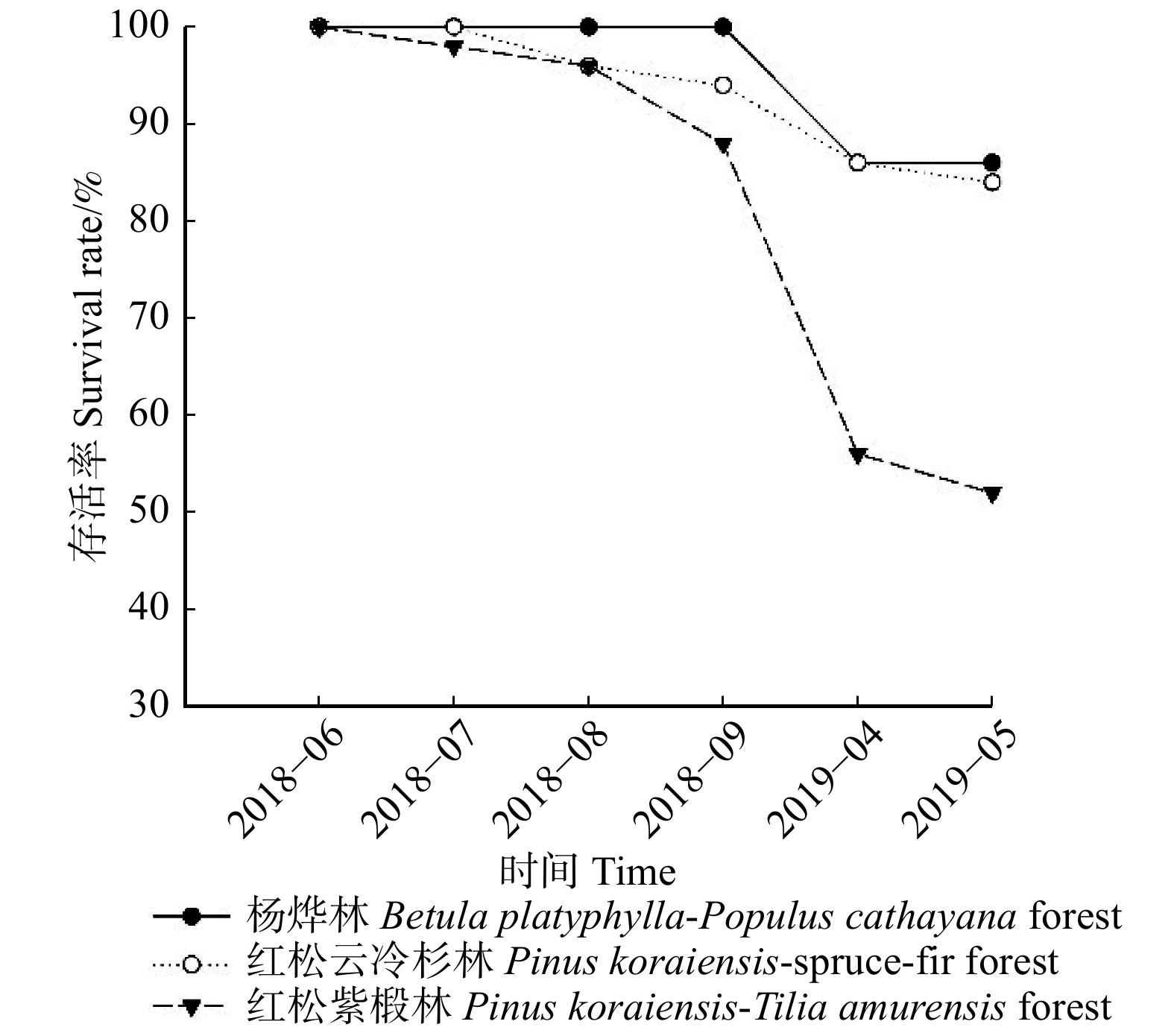
图 1 各林型4 ~ 5年生幼苗存活曲线Figure 1. Survival rate curves of 4−5 years old seedlings for each forest type由各林型1 ~ 2年生幼苗存活曲线(图2)可知,各林型1 ~ 2年东北红豆杉苗木中,红松紫椴林的幼苗存活率变化最大,其幼苗大量死亡的时间为2018年8月至2019年4月,杨桦林、红松云冷杉林的幼苗在2018年9月至2019年5月死亡较多。截止2019年5月,各林型幼苗存活率排序为杨桦林(86%) > 红松云冷杉林(84%) > 红松紫椴林(52%)。
![]() 图 2 各林型1 ~ 2年生幼苗存活率曲线Figure 2. Survival rate curves of 1−2 years old seedlings for each forest type
图 2 各林型1 ~ 2年生幼苗存活率曲线Figure 2. Survival rate curves of 1−2 years old seedlings for each forest type对比图1、2中可以发现,在2018年6月至9月期间,相同林型不同苗龄的幼苗存活率差距不大,在2018年9月以后,杨桦林试验地和红松云冷杉试验地4 ~ 5年生幼苗大量死亡,而1 ~ 2年生幼苗存活率仍在80%以上。红松紫椴林试验地4 ~ 5年生幼苗和1 ~ 2年生幼苗在这一时期均出现大量死亡的症状。
2.1.2 东北红豆杉幼苗生长特征
由表1可知,2018年9月,各林型4 ~ 5年生幼苗地径、苗高、冠幅和最长侧枝长均有一定的生长,地径生长量最大的是红松紫椴林,苗高生长量和冠幅面积生长量最大的是杨桦林,最长侧枝长生长量最大的是红松云冷杉林。各林型1 ~ 2年生幼苗地径、苗高、冠幅也有一定的生长,地径生长量最大的是红松云冷杉林,苗高生长量和冠幅面积生长量最大的是红松紫椴林。
表 1 不同林型幼苗各生长指标生长量Table 1. Growth of each seedling growth index for different forest types时间 Time 苗龄/a
Seedling age/year指标 Index BPF PSF PTF 2018−09 4 ~ 5 地径生长量
Growth of basal diameter/mm0.63 ± 1.32a 0.78 ± 1.10a 1.05 ± 1.15a 苗高生长量
Growth of seedling height/cm2.65 ± 3.23a 2.44 ± 2.84a 1.53 ± 1.80a 冠幅面积生长量
Growth of crown width area/cm266.19 ± 50.87a 55.06 ± 39.95a 63.26 ± 56.97a 最长侧枝长生长量
Growth of the maximum lateral branch length/cm1.28 ± 3.72a 2.72 ± 2.77a 1.11 ± 2.88a 1 ~ 2 地径生长量
Growth of basal diameter/mm0.21 ± 0.20a 0.24 ± 0.31a 0.21 ± 0.27a 苗高生长量
Growth of seedling height/cm0.82 ± 1.43b 1.05 ± 1.09b 1.99 ± 2.02a 冠幅面积生长量
Growth of crown width area/cm22.19 ± 3.97a 1.47 ± 4.36a 2.83 ± 3.75a 2019−05 4 ~ 5 地径生长量
Growth of basal diameter/mm− 0.02 ± 1.11a 0.60 ± 1.99a 0.56 ± 1.03a 苗高生长量
Growth of seedling height/cm− 5.72 ± 5.75b − 1.41 ± 4.35a − 5.40 ± 4.07b 冠幅面积生长量
Growth of crown width area/cm2− 40.45 ± 51.78a − 50.43 ± 41.35a − 69.15 ± 61.98a 最长侧枝长生长量
Growth of the maximum lateral branch length/cm− 9.62 ± 6.66a − 6.59 ± 6.16a − 8.73 ± 5.59a 1 ~ 2 地径生长量
Growth of basal diameter/mm0.28 ± 0.36a 0.27 ± 0.33a 0.22 ± 0.23a 苗高生长量
Growth of seedling height/cm− 0.09 ± 2.5b 0.46 ± 1.44b 1.73 ± 2.45a 冠幅面积生长量
Growth of crown width area/cm21.49 ± 5.21ab 0.23 ± 5.17b 2.78 ± 3.94a 注:同行不同小写字母表示该试验地间同一指标在0.05水平上差异显著。BPF代表杨桦林,PSF代表红松云冷杉林,PTF代表红松紫椴林。下同。Notes: different lowercase letters in the same row indicate significant differences of the same index among varied experimental sample plots at the level of 0.05. BPF represents Betula platyphylla-Populus cathayana forest, PSF represents Pinus koraiensis-spruce-fir forest, PTF represents Pinus koraiensis-Tilia amurensis forest. The same below. 2019年5月,各林型4 ~ 5年生东北红豆杉幼苗苗高生长量、冠幅面积生长量、最长侧枝长生长量以及杨桦林幼苗地径生长量均为负值,说明这些幼苗的苗高、冠幅、最长侧枝长与移栽完成时相比存在一定的损伤。各林型1 ~ 2年生幼苗地径生长量要高于2018年9月,但是苗高生长量、冠幅面积生长量要低于2018年9月。
差异显著性检验可知,2018年9月,各林型4 ~ 5年生幼苗地径生长量、苗高生长量、冠幅面积生长量、最长侧枝长生长量均没有显著性差异(P > 0.05),各林型1 ~ 2年生幼苗地径生长量、冠幅面积生长量没有显著性差异(P > 0.05),红松紫椴林的苗高生长量显著高于(P < 0.05)杨桦林和红松云冷杉林。2019年5月,各林型4 ~ 5年生幼苗地径生长量、冠幅面积生长量、最长侧枝长生长量均没有显著性差异(P > 0.05),各林型1 ~ 2年生东北红豆杉幼苗地径生长量没有显著性差异(P > 0.05),红松紫椴林的苗高生长量显著高于(P < 0.05)杨桦林试验地和红松云冷杉林试验地,红松紫椴林的冠幅面积生长量显著高于红松云冷杉林(P < 0.05),但与杨桦林差异不显著。
2.2 回归幼苗适宜林型评价
根据表2可知,在幼苗移栽的第1个生长季中,各林型4 ~ 5年生幼苗的关联度由高到低依次是红松云冷杉林 > 杨桦林 > 红松紫椴林。说明在2018年5—9月期间4 ~ 5年生东北红豆杉幼苗的各项指标与红松云冷杉林的关联度最高,综合表现最好,红松云冷杉林更适宜开展4 ~ 5年生东北红豆杉幼苗野外回归。各林型1 ~ 2年生幼苗的关联度由高到低依次是红松紫椴林 > 杨桦林 > 红松云冷杉林,说明在观测期间,1 ~ 2年生东北红豆杉幼苗在红松紫椴林的综合表现最好,适应性更强,相比于杨桦林和红松云冷杉林,更适宜开展1 ~ 2年生东北红豆杉野外回归。
表 2 幼苗生长指标生长量和存活率与各林型的关联度及排序Table 2. Correlation degree between growth of seedling growth indexes as well as survival rate and each forest type and their rank苗龄/a
Seedling age/year指标 Index BPF PSF PTF 4 ~ 5 苗高生长量 Growth of seedling height 0.755 2 0.798 3 0.889 8 地径生长量 Growth of basal diameter 0.865 8 0.838 8 0.738 8 冠幅面积生长量 Growth of crown width area 0.830 7 0.784 6 0.818 1 最长侧枝长生长量 Growth of the maximum lateral branch length 0.751 4 1.000 0 0.730 0 存活率 Survival rate 0.823 4 0.791 4 0.817 9 关联度均值 Mean value of correlation degree 0.805 3 0.842 6 0.798 9 排序 Rank 2 1 3 1 ~ 2 苗高生长量 Growth of seedling height 0.445 9 0.535 6 1.000 0 地径生长量 Growth of basal diameter 0.602 5 0.595 8 0.564 5 冠幅面积生长量 Growth of crown width area 0.586 1 0.474 4 0.772 0 存活率 Survival rate 0.615 1 0.610 3 0.541 9 关联度均值 Mean value of correlation degree 0.562 4 0.554 0 0.719 6 排序 Rank 2 3 1 2.3 影响回归幼苗存活的环境因素
2.3.1 不同林型的环境因子
不同林型环境因子(表3)显示,各林型在海拔都在800 ~ 900 m的范围内,郁闭度都在0.5 ~ 0.6之间,坡度在15° ~ 20°之间,坡向均为阴坡或半阴坡。各林型之间的土壤pH值差异不显著(P > 0.05),杨桦林的土壤有机质含量显著高于(P < 0.05)红松云冷杉林和红松紫椴林,全氮含量显著高于(P < 0.05)红松云冷杉林,但与红松紫椴林的土壤全氮含量差异不显著(P > 0.05),杨桦林和红松云冷杉林的全磷含量显著高于红松紫椴林(P < 0.05)。
表 3 不同林型环境因子Table 3. Environmental factors in different forest types林型
Forest type海拔
Altitude/m郁闭度
Canopy density坡度
Slope degree/(°)坡向
Slope aspectpH 有机质含量
Organic matter
content/(g·kg− 1)全氮含量
Total N
content/(g·kg− 1)全磷含量
Total P
content/(g·kg− 1)BPF 838 0.56 19 北偏西53°
North by west 53°4.98 ± 0.34a 198.53 ± 18.3 a 7.55 ± 0.31 a 0.65 ± 0.08 a PSF 846 0.55 16 北偏西3°
North by west 3°4.97 ± 0.26a 148.27 ± 17.78 b 5.41 ± 0.92 b 0.70 ± 0.03 a PTF 863 0.58 18 北偏东12°
North by east 12°4.94 ± 0.28a 143.70 ± 29.74 b 6.24 ± 0.82 ab 0.43 ± 0.11 b 2.3.2 幼苗生长指标和存活状况与环境因子的灰色关联分析
从1 ~ 2年生东北红豆杉幼苗地径生长量、苗高生长量、冠幅面积生长量和存活率与9个环境因子的灰色关联度计算结果(表4)可知,与地径生长量关联度最高的环境因子是坡向和坡度,与苗高生长量关联度最高的环境因子是土壤全磷含量,与冠幅面积生长量关联度最高的环境因子是林型,与存活率关联度最高的环境因子是林型和坡向。关联度均值排前4位主要是生物因子(林型、郁闭度)和地形因子(坡向、坡度),土壤化学性质的排名在后5位。该结果表明,对1 ~ 2年生东北红豆杉幼苗生长指标和存活率影响最大的是林型,其次是坡向、坡度、郁闭度,土壤化学性质的影响较小。土壤化学性质排第1位的是土壤pH值。
表 4 1 ~ 2年生幼苗生长指标和存活率与环境因子的关联度及排序Table 4. Correlation degree between seedling growth indexes as well as survival rate and environmental factors and their rank环境因子
Environmental factor关联度 Correlation degree 关联度均值
Mean value of
correlation degree排序
Rank地径生长量
Growth of basal diameter苗高生长量
Growth of seedling height冠幅面积生长量
Growth of crown
width area存活率
Survival rate林型 Forest type 0.705 8 0.607 6 0.810 9 0.851 6 0.744 0 1 坡向 Slope aspect 0.858 3 0.605 4 0.640 2 0.863 9 0.741 9 2 坡度 Slope degree 0.854 0 0.680 5 0.579 5 0.830 9 0.736 2 3 郁闭度 Canopy density 0.836 6 0.681 7 0.556 0 0.798 1 0.718 1 4 pH 0.780 5 0.689 2 0.561 6 0.765 6 0.699 2 5 海拔 Altitude 0.767 3 0.689 2 0.559 4 0.759 5 0.693 8 6 全氮 Total N 0.829 4 0.624 2 0.675 9 0.560 0 0.672 4 7 有机质 Organic matter 0.638 2 0.642 3 0.685 1 0.666 3 0.658 0 8 全磷 Total P 0.573 8 0.732 8 0.604 2 0.581 6 0.623 1 9 3. 讨论与结论
3.1 回归效果
在不同林型4 ~ 5年生幼苗回归的第1个生长季中,东北红豆杉幼苗有较高的存活率,且苗高、地径、冠幅、最长侧枝长等指标均有一定幅度的增长,但是在越冬期间由于受到狍子(Capreolus capreolus)的啃食,幼苗的枝、叶受到严重的破坏,越冬后杨桦林幼苗存活率下降了60%,存活幼苗最多的红松紫椴林幼苗存活率也下降了28%,各生长指标也出现了大幅度的缩减(表1),越冬后存活的幼苗仅存少量叶片、主枝和侧枝受损严重,没有新芽,基本宣告4 ~ 5年生东北红豆杉幼苗回归失败。这表明东北红豆杉植株在回归过程中,除了考虑到生境中的立地环境、土壤化学性质、光照和树种组成等非生物因子外,还需要考虑动物因子对回归效果产生的不利影响。周绍春等[29]对完达山地区狍子冬季生境选择的研究中发现,狍子冬季偏向于针阔混交林和阔叶林,不喜欢针叶混交林。但是本研究中发现各林型中所有4 ~ 5年生幼苗均受到了不同程度的啃食,
各林型1 ~ 2年生幼苗在回归的第一个生长季中,幼苗存活率较高,且地径、苗高、冠幅均有所增长,说明1 ~ 2年生幼苗在移栽后能适应原生境及杨桦林这种次生生境。但是经历越冬后,各林型1 ~ 2年生幼苗存活率有不同程度的降低,幼苗的苗高、冠幅相对越冬之前都有一定程度的减少,表明1 ~ 2年生幼苗在越冬期间长时间受到低温胁迫其生长受到影响。这与刘彤等[30]、张继武等[31]得出多次经历自然越冬的幼苗对低温的耐受力要远高于初次经历室外越冬的幼苗的结论相一致。
3.2 适宜幼苗回归林型
本研究通过灰色关联分析,对不同苗龄东北红豆杉幼苗适宜回归的林型进行评价,4 ~ 5年生幼苗适应性最好的是红松云冷杉林,1 ~ 2年生幼苗适应性最好的是红松紫椴林,说明东北红豆杉幼苗更适宜在原生境开展回归。4 ~ 5年生幼苗与红松紫椴林的关联度最低,而1 ~ 2年生幼苗与红松云冷杉林的关联度最低,这可能与不同苗龄的幼苗叶生物量以及光合面积的大小不同有关。由于是野外回归试验,杂草与幼苗会存在光照、水分和养分的竞争关系,相对于4 ~ 5年生幼苗,1 ~ 2年生幼苗对竞争杂草更为敏感,这也可能是导致不同苗龄东北红豆杉适宜回归林型不同的原因。
3.3 东北红豆杉回归与环境因子之间的关系
地形因子通过土壤、气候、生物因子在空间上的不同组合而间接地对植物发生作用[32]。本研究中地形因子与东北红豆杉回归幼苗生长和存活关联度均值的排序为坡向 > 坡度 > 海拔,坡向与幼苗的地径生长量和存活率关联度最高,坡度与幼苗地径生长量和存活率关联度较高,海拔与幼苗生长指标和存活率的关联度较低。坡向主要影响太阳辐射而改变热量与水分蒸发,坡度主要影响水分的流失,两者最主要的还是影响土壤含水量。东北红豆杉是典型的耐阴树种,性喜阴湿,幼苗在长时间缺水的环境中叶片光合作用减弱甚至死亡[33]。因此,在进行东北红豆杉种群回归时,选择阴坡或半阴坡以及适宜的坡度有利于幼苗的生长于存活。海拔高度主要影响温度,由于3个林型的海拔变化范围在100 m之内,气温上不会有太大的变化,因此海拔对本研究中幼苗的生长和存活影响不明显。
光是植物进行光合作用的基础,是影响幼苗生存与生长的重要环境因子之一。刘彤等[34]对东北红豆杉幼苗进行遮阴处理发现强光会影响幼苗正常的光合作用,30%的遮阴条件下幼苗光合作用最强。本研究中各林型的郁闭度在0.5 ~ 0.6之间,幼苗也能正常生长,但幼苗回归最适宜的郁闭度范围还需要进一步的试验进行研究。
土壤酸碱度是土壤重要的化学性质。本研究中,土壤pH值与东北红豆杉幼苗生长指标和存活率的关联度较大,说明土壤酸碱度对东北红豆杉回归幼苗的存活和生长有一定的影响。不同林型的土壤营养元素含量也存在差异,杨桦林的土壤有机质含量要显著高于红松云冷杉林和红松紫椴林,全氮含量显著高于红松云冷杉林,这与张雨鉴等[35]得出的土壤各土层C含量表现为阔叶林 > 针叶林,土壤N含量阔叶林 > 针叶林的结论相一致。但是土壤营养元素与东北红豆杉幼苗存活和生长的关联度较小,可能是由于东北红豆杉是慢生树种,1 ~ 2年生幼苗生长缓慢,对营养元素的需求较小有关。
3.4 回归试验的启示
本研究结果表明,动物因子对东北红豆杉回归的影响很大,是导致4 ~ 5年生幼苗回归效果不良的主要原因,也是1 ~ 2年生幼苗在之后的生长发育过程中最重要的威胁因素,这说明在回归过程中不仅要考虑环境因子对幼苗回归的影响,也要考虑生物因子对幼苗回归是否存在影响。因此,预先调查回归试验点是否存在啃食回归幼苗的动物具有非常重要的意义;在回归试验点无法避开野生动物的情况下,针对回归幼苗的生长状况以及回归地动物的生境选择,采取合理有效的保护方法,如干怀新[36]发明的新型造林地幼苗保护装置等,是保护回归幼苗在回归过程中正常生长发育最重要的一环。
-
![]()
图 1 各林型4 ~ 5年生幼苗存活曲线
Figure 1. Survival rate curves of 4−5 years old seedlings for each forest type
![]()
图 2 各林型1 ~ 2年生幼苗存活率曲线
Figure 2. Survival rate curves of 1−2 years old seedlings for each forest type
表 1 不同林型幼苗各生长指标生长量
Table 1 Growth of each seedling growth index for different forest types
时间 Time 苗龄/a
Seedling age/year指标 Index BPF PSF PTF 2018−09 4 ~ 5 地径生长量
Growth of basal diameter/mm0.63 ± 1.32a 0.78 ± 1.10a 1.05 ± 1.15a 苗高生长量
Growth of seedling height/cm2.65 ± 3.23a 2.44 ± 2.84a 1.53 ± 1.80a 冠幅面积生长量
Growth of crown width area/cm266.19 ± 50.87a 55.06 ± 39.95a 63.26 ± 56.97a 最长侧枝长生长量
Growth of the maximum lateral branch length/cm1.28 ± 3.72a 2.72 ± 2.77a 1.11 ± 2.88a 1 ~ 2 地径生长量
Growth of basal diameter/mm0.21 ± 0.20a 0.24 ± 0.31a 0.21 ± 0.27a 苗高生长量
Growth of seedling height/cm0.82 ± 1.43b 1.05 ± 1.09b 1.99 ± 2.02a 冠幅面积生长量
Growth of crown width area/cm22.19 ± 3.97a 1.47 ± 4.36a 2.83 ± 3.75a 2019−05 4 ~ 5 地径生长量
Growth of basal diameter/mm− 0.02 ± 1.11a 0.60 ± 1.99a 0.56 ± 1.03a 苗高生长量
Growth of seedling height/cm− 5.72 ± 5.75b − 1.41 ± 4.35a − 5.40 ± 4.07b 冠幅面积生长量
Growth of crown width area/cm2− 40.45 ± 51.78a − 50.43 ± 41.35a − 69.15 ± 61.98a 最长侧枝长生长量
Growth of the maximum lateral branch length/cm− 9.62 ± 6.66a − 6.59 ± 6.16a − 8.73 ± 5.59a 1 ~ 2 地径生长量
Growth of basal diameter/mm0.28 ± 0.36a 0.27 ± 0.33a 0.22 ± 0.23a 苗高生长量
Growth of seedling height/cm− 0.09 ± 2.5b 0.46 ± 1.44b 1.73 ± 2.45a 冠幅面积生长量
Growth of crown width area/cm21.49 ± 5.21ab 0.23 ± 5.17b 2.78 ± 3.94a 注:同行不同小写字母表示该试验地间同一指标在0.05水平上差异显著。BPF代表杨桦林,PSF代表红松云冷杉林,PTF代表红松紫椴林。下同。Notes: different lowercase letters in the same row indicate significant differences of the same index among varied experimental sample plots at the level of 0.05. BPF represents Betula platyphylla-Populus cathayana forest, PSF represents Pinus koraiensis-spruce-fir forest, PTF represents Pinus koraiensis-Tilia amurensis forest. The same below.  下载: 导出CSV
下载: 导出CSV
表 2 幼苗生长指标生长量和存活率与各林型的关联度及排序
Table 2 Correlation degree between growth of seedling growth indexes as well as survival rate and each forest type and their rank
苗龄/a
Seedling age/year指标 Index BPF PSF PTF 4 ~ 5 苗高生长量 Growth of seedling height 0.755 2 0.798 3 0.889 8 地径生长量 Growth of basal diameter 0.865 8 0.838 8 0.738 8 冠幅面积生长量 Growth of crown width area 0.830 7 0.784 6 0.818 1 最长侧枝长生长量 Growth of the maximum lateral branch length 0.751 4 1.000 0 0.730 0 存活率 Survival rate 0.823 4 0.791 4 0.817 9 关联度均值 Mean value of correlation degree 0.805 3 0.842 6 0.798 9 排序 Rank 2 1 3 1 ~ 2 苗高生长量 Growth of seedling height 0.445 9 0.535 6 1.000 0 地径生长量 Growth of basal diameter 0.602 5 0.595 8 0.564 5 冠幅面积生长量 Growth of crown width area 0.586 1 0.474 4 0.772 0 存活率 Survival rate 0.615 1 0.610 3 0.541 9 关联度均值 Mean value of correlation degree 0.562 4 0.554 0 0.719 6 排序 Rank 2 3 1
下载: 导出CSV
表 3 不同林型环境因子
Table 3 Environmental factors in different forest types
林型
Forest type海拔
Altitude/m郁闭度
Canopy density坡度
Slope degree/(°)坡向
Slope aspectpH 有机质含量
Organic matter
content/(g·kg− 1)全氮含量
Total N
content/(g·kg− 1)全磷含量
Total P
content/(g·kg− 1)BPF 838 0.56 19 北偏西53°
North by west 53°4.98 ± 0.34a 198.53 ± 18.3 a 7.55 ± 0.31 a 0.65 ± 0.08 a PSF 846 0.55 16 北偏西3°
North by west 3°4.97 ± 0.26a 148.27 ± 17.78 b 5.41 ± 0.92 b 0.70 ± 0.03 a PTF 863 0.58 18 北偏东12°
North by east 12°4.94 ± 0.28a 143.70 ± 29.74 b 6.24 ± 0.82 ab 0.43 ± 0.11 b
下载: 导出CSV
表 4 1 ~ 2年生幼苗生长指标和存活率与环境因子的关联度及排序
Table 4 Correlation degree between seedling growth indexes as well as survival rate and environmental factors and their rank
环境因子
Environmental factor关联度 Correlation degree 关联度均值
Mean value of
correlation degree排序
Rank地径生长量
Growth of basal diameter苗高生长量
Growth of seedling height冠幅面积生长量
Growth of crown
width area存活率
Survival rate林型 Forest type 0.705 8 0.607 6 0.810 9 0.851 6 0.744 0 1 坡向 Slope aspect 0.858 3 0.605 4 0.640 2 0.863 9 0.741 9 2 坡度 Slope degree 0.854 0 0.680 5 0.579 5 0.830 9 0.736 2 3 郁闭度 Canopy density 0.836 6 0.681 7 0.556 0 0.798 1 0.718 1 4 pH 0.780 5 0.689 2 0.561 6 0.765 6 0.699 2 5 海拔 Altitude 0.767 3 0.689 2 0.559 4 0.759 5 0.693 8 6 全氮 Total N 0.829 4 0.624 2 0.675 9 0.560 0 0.672 4 7 有机质 Organic matter 0.638 2 0.642 3 0.685 1 0.666 3 0.658 0 8 全磷 Total P 0.573 8 0.732 8 0.604 2 0.581 6 0.623 1 9
下载: 导出CSV
-
[1] Wiens J J. Climate-related local extinctions are already widespread among plant and animal species[J/OL]. PLoS Biology, 2016, 14(12), e2001104 (2016−09−08) [2018−08−14]. http://doi.org/10.1371/journal.pbio.2001104.
[2] Oliver T H, Marshall H H, Morecroft M D, et al. Interacting effects of climate change and habitat fragmentation on drought-sensitive butterflies[J]. Nature Climate Change, 2015, 5(10): 941−945. doi: 10.1038/nclimate2746
[3] Ren H, Zhang Q M, Lu H F. Wild plant species with extremely small populations require conservation and reintroduction in China[J]. Royal Swedish Academy of Sciences, 2012, 41: 913−917.
[4] 臧润国, 董鸣, 李俊清, 等. 典型极小种群野生植物保护与恢复技术研究[J]. 生态学报, 2016, 36(22):7130−7135. Zang R G, Dong M, Li J Q, et al. Conservation and restoration for typical critically endangered wild plants with extremely small population[J]. Acta Ecologica Sinica, 2016, 36(22): 7130−7135.
[5] Ren H, Jian S G, Liu H X, et al. Advances in the reintroduction of rare and endangered wild plant species[J]. Science China Life Sciences, 2014, 57(6): 603−609. doi: 10.1007/s11427-014-4658-6
[6] 周翔, 高江云. 珍稀濒危植物的回归:理论和实践[J]. 生物多样性, 2011, 19(1):97−105. doi: 10.3724/SP.J.1003.2011.09101 Zhou X, Gao J Y. The reintroduction of rare and endangered plants: theory and practice[J]. Biodiversity Science, 2011, 19(1): 97−105. doi: 10.3724/SP.J.1003.2011.09101
[7] Godefroid S, Piazza C, Rossi G, et al. How successful are plant species reintroductions?[J]. Biological Conservation, 2011, 144(2): 672−682. doi: 10.1016/j.biocon.2010.10.003
[8] Albrecht M A, Long Q G. Habitat suitability and herbivores determine reintroduction success of an endangered legume[J]. Plant Diversity, 2019, 41(2): 109−117. doi: 10.1016/j.pld.2018.09.004
[9] Erin J Q, James R K, Kealoha K, et al. Mapping habitat suitability for at-risk plant species and its implications for restoration and reintroduction[J]. Ecological Applications: A Publication of the Ecological Society of America, 2014, 24(2): 385−395. doi: 10.1890/13-0775.1
[10] 汪越, 易慧琳, 邵玲, 等. 紫背天葵(<italic>Begonia fimbristipula</italic> Hance)回归植株存活及叶片生物学特性研究[J]. 生态科学, 2017, 36(2):32−41. Wang Y, Li H L, Shao L, et al. Survival and eco-biological characteristics of <italic>Begonia fimbristipula</italic> Hance in the process of reintroduction[J]. Ecological Science, 2017, 36(2): 32−41.
[11] 李景秀, 崔卫华, 胡枭剑, 等. 濒危植物古林箐秋海棠的扦插繁殖及回归引种初探[J]. 广西植物, 2018, 38(7):851−858. doi: 10.11931/guihaia.gxzw201705019 Li J X, Cui W H, Hu X J, et al. Cutting propagation and regression planting of endangered plant <italic>Begonia gulinqingensis</italic>[J]. Guihaia, 2018, 38(7): 851−858. doi: 10.11931/guihaia.gxzw201705019
[12] 谭美, 杨志玲, 杨旭, 等. 厚朴野生资源的野外回归植株苗期适应性评价[J]. 林业科学研究, 2019, 32(1):125−132. Tan M, Yang Z L, Yang X, et al. Adaptability evaluation of wild <italic>Houpoëa officinalis</italic> seedling in the process of reintroduction[J]. Forest Research, 2019, 32(1): 125−132.
[13] 冯欣欣, 余金昌, 袁志永, 等. 国家Ⅱ级保护植物竹叶兰的迁地保护与野外回归研究[J]. 中国园艺文摘, 2017, 33(3):12−14. doi: 10.3969/j.issn.1672-0873.2017.03.005 Feng X X, Yu J C, Yuan Z Y, et al. The ex-situ conservation and reintroduction of the national second-class protection plant <italic>Arundina graminifolia</italic>[J]. Chinese Horticulture Abstracts, 2017, 33(3): 12−14. doi: 10.3969/j.issn.1672-0873.2017.03.005
[14] 陈芳清, 谢宗强, 熊高明, 等. 三峡濒危植物疏花水柏枝的回归引种和种群重建[J]. 生态学报, 2005, 25(7):1813−1817. Chen F Q, Xie Z Q, Xiong G M, et al. Reintroduction and population reconstruction of an endangered plant <italic>Myricaria laxiflora</italic> in the Three Gorges Reservoir Area, China[J]. Acta Ecologica Sinica, 2005, 25(7): 1813−1817.
[15] 简尊吉, 马凡强, 郭泉水, 等. 回归崖柏苗木存活和生长对海拔梯度的响应[J]. 林业科学, 2017, 53(11):1−11. doi: 10.11707/j.1001-7488.20171101 Jian Z J, Ma F Q, Guo Q S, et al. Responses of survival and growth of <italic>Thuja sutchuenensis</italic> reintroduction seedlings to altitude gradient[J]. Scientia Silave Sinicae, 2017, 53(11): 1−11. doi: 10.11707/j.1001-7488.20171101
[16] 周志强, 刘彤, 胡林林, 等. 穆棱东北红豆杉年轮—气候关系及其濒危机制[J]. 生态学报, 2010, 30(9):2304−2310. Zhou Z Q, Liu T, Hu L L, et al. Tree ring-climate response and endangered mechanism deliberation of Japanese yew (<italic>Taxus cuspidata</italic>) in Muling Nature Reserve[J]. Acta Ecologica Sinica, 2010, 30(9): 2304−2310.
[17] 刘彤. 天然东北红豆杉种群生态学研究[D]. 哈尔滨: 东北林业大学, 2007. Liu T. Population ecology of natural Japanese yew[D]. Harbin: Northeast Forestry University, 2007.
[18] 李云灵.东北红豆杉种间关系研究[D].哈尔滨: 东北林业大学, 2008. Li Y L. Interspecific relationship of the Taxus cuspidata[D]. Harbin: Northeast Forestry University, 2008.
[19] 周志强, 刘彤, 袁继连. 黑龙江穆棱天然东北红豆杉种群资源特征研究[J]. 植物生态学报, 2004, 28(4):476−482. doi: 10.3321/j.issn:1005-264X.2004.04.005 Zhou Z Q, Liu T, Yuan J L. Population characteristics of yew(<italic>Taxus cuspidata</italic>) in the Muling Yew Nature Reserve, Heilongjiang Province[J]. Acta Phytoecologica Sinica, 2004, 28(4): 476−482. doi: 10.3321/j.issn:1005-264X.2004.04.005
[20] 陈杰, 龙婷, 杨蓝, 等. 东北红豆杉生境适宜性评价[J]. 北京林业大学学报, 2019, 41(4):51−59. Chen J, Long T, Yang L, et al. Habitat suitability assessment of <italic>Taxus cuspidate</italic>[J]. Journal of Beijing Forestry University, 2019, 41(4): 51−59.
[21] 杨占, 张强, 卢元, 等. 辽宁省东北红豆杉天然种群特征研究[J]. 防护林科技, 2017(3):16−17. Yang Z, Zhang Q, Lu Y, et al. Distribution and characteristics of wild natural <italic>Taxus cuspidata</italic> in Liaoning Province[J]. Protection Forest Science and Technology, 2017(3): 16−17.
[22] 刁云飞. 东北红豆杉−红松林群落结构与空间关联性研究[D].哈尔滨: 东北林业大学, 2015. Diao Y F. Community structure and spatial correlation of a Taxus cuspidata-Pinus koraiehsis forest[D]. Harbin: Northeast Forestry University, 2015.
[23] 王兆东, 孙习华, 张荣哲. 东北红豆杉扦插繁殖技术研究[J]. 现代园林, 2007(12):82−83. Wang Z D, Sun X H, Zhang R Z. Research on cottage propagation techniques of <italic>Taxus cuspidata</italic>[J]. Modern Landscape Architecture, 2007(12): 82−83.
[24] 马盈, 丰庆义, 杨凯. 东北红豆杉繁殖技术研究进展评述[J]. 林业勘查设计, 2013(1):62−64. doi: 10.3969/j.issn.1673-4505.2013.01.030 Ma Y, Feng Q Y, Yang K. Review on the research on propagation technique of <italic>Taxus cuspidata</italic>[J]. Forest Investigation Design, 2013(1): 62−64. doi: 10.3969/j.issn.1673-4505.2013.01.030
[25] 吴杰, 汤欢, 黄林芳, 等. 红豆杉属植物全球生态适宜性分析研究[J]. 药学学报, 2017, 52(7):1186−1195. Wu J, Tang H, Huang L F, et al. Global ecological suitability analysis of <italic>Taxus</italic>[J]. Acta Pharmaceutica Sinica, 2017, 52(7): 1186−1195.
[26] 孔维尧, 孙权, 刘鑫鑫, 等. 基于红外相机监测的汪清自然保护区东北豹种群动态[J]. 林业科学, 2019, 55(5):188−196. doi: 10.11707/j.1001-7488.20190521 Sun W Y, Sun Q, Liu X X, et al. Population dynamic of dar eastern leopard (<italic>Panthera pardus orientalis</italic>) in Wangqing Nature Reserve based on infrared camera monitoring[J]. Scientia Silave Sinicae, 2019, 55(5): 188−196. doi: 10.11707/j.1001-7488.20190521
[27] 国家林业局. 中华人民共和国林业行业标准: 森林土壤分析方法[S]. 北京: 中国标准出版社, 1999. State Forestry Administration. Forestry industry standard of the People ’s Republic of China: forest soil analysis method[S]. Beijing: China Standard Press, 1999.
[28] 余敏, 周志勇, 康峰峰, 等. 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释[J]. 植物生态学报, 2013, 37(5):373−383. doi: 10.3724/SP.J.1258.2013.00373 Yu M, Zhou Z Y, Kang F F, et al. Gradient analysis and environmental interpretation of understory herb-layer communities in Xiaoshegou of Lingkong Mountain, Shanxi, China[J]. Chinese Journal of Plant Ecology, 2013, 37(5): 373−383. doi: 10.3724/SP.J.1258.2013.00373
[29] 周绍春, 张明海, 尹远新, 等. 黑龙江完达山地区狍子冬季生境选择[J]. 北京林业大学学报, 2010, 32(3):122−127. Zhou S C, Zhang M H, Yin Y X, et al. Habitat selection of roe deer (<italic>Capreolus capreolus</italic>) in winter in the eastern Wandashan Mountains, Heilongjiang Province[J]. Journal of Beijing Forestry University, 2010, 32(3): 122−127.
[30] 刘彤, 祝佳媛, 李鹏, 等. 秋冬季自然降温过程中东北红豆杉幼苗的生理生化特性[J]. 北京林业大学学报, 2013, 35(2):51−56. Liu T, Zhu J Y, Li P, et al. Physiological and biochemical characteristics of Japanese yew seedlings as natural temperature falling in autumn and winter[J]. Journal of Beijing Forestry University, 2013, 35(2): 51−56.
[31] 张继武. 东北红豆杉幼苗越冬过程中理化特性研究[J]. 黑龙江科技信息, 2013(12):261. Zhang J W. Physical and chemical characteristics of <italic>Taxus cuspidata</italic> seedlings during overwintering[J]. Heilongjiang Scientific and Technological Information, 2013(12): 261.
[32] 李俊清. 森林生态学[M]. 北京: 高等教育出版社, 2010. Li J Q. Forest ecology[M]. Beijing: Higher Education Press, 2010.
[33] 徐博超, 周志强, 李威, 等. 东北红豆杉幼苗对不同水分条件的光合和生理响应[J]. 北京林业大学学, 2012, 34(4):73−78. Xu B C, Zhou Z Q, Li W, et al. Physiological and photosynthetic response to different water conditions of <italic>Taxus cuspidate</italic> seedings[J]. Journal of Beijing Forestry University, 2012, 34(4): 73−78.
[34] 刘彤, 崔海娇, 吴淑杰, 等. 东北红豆杉幼苗光合和荧光特性对不同光照条件的响应[J]. 北京林业大学学报, 2013, 35(3):65−70. Liu T, Cui H J, Wu S J, et al. Response of photosynthetic and fluorescence characteristics of Japanese yew seedlings to different light conditions[J]. Journal of Beijing Forestry University, 2013, 35(3): 65−70.
[35] 张雨鉴, 王克勤, 宋娅丽, 等. 滇中亚高山5种林型土壤碳氮磷生态化学计量特征[J]. 生态环境学报, 2019, 28(1):73−82. Zhang Y J, Wang K Q, Song Y L, et al. Ecological stoichiometry of soil carbon, nitrogen and phosphorus in five forest types in subalpine of middle Yunnan Province[J]. Ecology and Environmental Sciences, 2019, 28(1): 73−82.
[36] 干怀新. 一种新型造林地幼苗保护装置: CN209435912U[P]. 2019−09−27. Gan H X. The utility model relates to a new seedling protection device in constructed woodland: CN209435912U[P]. 2019−09−27.
-
期刊类型引用(4)
1. 杨维红,刘忠,杨博文,刘晓岚. 一种重要的造纸助剂——湿强剂. 天津造纸. 2022(02): 39-45 .  百度学术
百度学术
2. 宁晓,符庆金,王燕云,姚春丽,梁帅博,袁涛,顿旭继. 聚丙烯酰胺环氧氯丙烷-膨润土的二元体系增加湿强度效果及其机理. 东北林业大学学报. 2020(06): 110-114+119 . 百度学术
3. 梁帅博,姚春丽,符庆金,刘倩,袁涛. 纸张二元增强体系的研究进展. 中国造纸学报. 2020(02): 89-95 . 百度学术
4. 宁晓,姚春丽,管丽娜. 聚酰胺多胺环氧氯丙烷/荧光海藻酸钠体系对二次纤维增湿强的机理. 造纸科学与技术. 2018(03): 46-51+61 . 百度学术
其他类型引用(4)
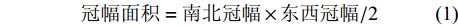
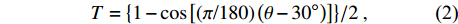
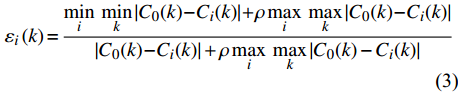
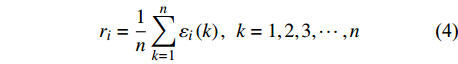
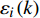
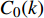
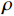
计量
- 文章访问数: 1494
- HTML全文浏览量: 486
- PDF下载量: 79
- 被引次数: 8
