
Spatial heterogeneity of photosynthetic characteristics in Chinese chestnut canopy
-
摘要:目的 冠层是林木与外部环境相互作用的重要部分,研究板栗树冠不同部位光合作用的空间变化规律,为揭示植物冠层生产力提供理论依据。方法 以8年生板栗树为试验材料,采用Li-6400便携式光合作用测量系统,于7月对板栗树冠不同冠层和不同方位进行光合日变化测定,并取光合参数的日均值进行方差分析及通径分析,于9月对不同冠层和不同方位的刺苞进行采集,测定坚果经济性状,并进行方差分析及相关性分析。结果 (1)东上、东中、南中及南下冠层的净光合速率日变化呈单峰曲线分布,其余部位均表现为双峰曲线,呈双峰曲线分布的区域均于13:00出现“午休”现象;每一冠层区域的水分利用效率日动态均呈双峰曲线。(2)在冠层垂直方向上,水汽压亏缺及胞间CO2浓度均呈“下部 > 中部 > 上部”的规律,其最大值分别为2.13 kPa、274.93 μmol/mol,最小值分别为1.95 kPa、258.75 μmol/mol;光合速率、气孔导度、蒸腾速率、光合有效辐射、水分利用效率及羧化效率均呈“上部 > 中部 > 下部”的规律,同时,相对叶绿素含量(SPAD)和比叶面积(SLA)在垂直冠层上均有显著差异,表现为“下层 > 中层 > 上层”的规律。(3)在树冠不同方位,东面光合能力最强,各冠层光合生理生态指标有一定差异性,但无垂直方向差异显著;SLA在不同方位无显著差异,仅SPAD在东西向与北向差异显著。(4)通径分析得出,影响板栗净光合速率的主要因子为蒸腾速率、水分利用效率、羧化效率、水汽压亏缺、气孔导度、气温及胞间CO2浓度,其中,只有气温对净光合速率起抑制作用,蒸腾速率是对板栗光合作用影响最大的生理因子,水汽压亏缺是对板栗光合作用影响最大的环境因子。(5)垂直冠层上,净光合速率与单粒质量、出实率和单位面积产量均呈显著或极显著正相关,相关系数分别为0.872、0.965、0.958;水平冠层上,净光合速率与单粒质量、单位面积产量呈显著正相关,相关系数分别为0.777、0.487;对水平和垂直冠层进行综合分析发现,净光合速率与单粒质量、出实率、单位面积产量均呈显著或极显著正相关关系,相关系数分别为0.600、0.669与0.532。结论 板栗光合具有明显的空间异质性,各光合指标与果实产量的差异性是垂直冠层和水平冠层综合作用的结果。因此,在估算冠层生产力时,应充分考虑到冠层光合作用的垂直变化和水平变化,在实际生产中,应合理修枝,提高冠层中下部的光能利用率,从而促进果实产量的提高。Abstract:Objective Canopy is an important part of the interaction between trees and external environment. The study on spatial variation law of photosynthesis in different parts of Chinese chestnut canopy provides a theoretical basis for revealing Chinese chestnut canopy productivity.Method The 8-year-old Chinese chestnut trees were used as tested materials, the Li-6400 portable photosynthesis measurement system was used to determine the diurnal changes of photosynthesis in different canopies and positions of the Chinese chestnut canopy in July, fruit collection was carried out to determine the economic traits of nuts in September. Variance analysis, path analysis and regression analysis were done on the base of indexes.Result (1) The daily dynamics of photosynthetic rate (Pn) in the upper east, middle east, south middle, and lower south showed a unimodal curve distribution; the rest of the region showed a bimodal curve, and the areas showing a bimodal curve distribution all presented a “midday depression” phenomenon at 13:00. The daily dynamics of water use efficiency (WUE) in each canopy area presented a bimodal curve. (2) In the vertical direction of the canopy, the deficient vapor pressure (VpdL) and intercellular CO2 concentration (Ci) were shown as “lower > middle > upper”. The maximum values were 2.13 kPa, 274.93 μmol/mol, and the minimum values were 1.95 kPa, 258.75 μmol/mol; photosynthetic rate, stomatal conductance (Gs), transpiration rate (Tr), photosynthetically active radiation (PAR), WUE and carboxylation efficiency (Vc) presented a “upper > middle > lower” phenomenon. The SPAD and specific leaf area (SLA) had significant differences in the vertical canopy (P < 0.05), and both showed “lower > middle > upper”. (3) In different directions in the canopy, the strongest photosynthetic capacity was in the east, but the photosynthetic indexes were not significantly different compared with the vertical canopy. The SPAD only had significantly difference in the east, west and north directions. The SLA was not significantly different in varied directions. (4) Path analysis showed that the main factors affecting Pn were Tr, WUE, Vc, VpdL, Gs, air temperature (Ta) and Ci, only Ta had an inhibitory effect on the Pn; Tr and VpdL were the physiological and environmental factors that had the greatest influence on Chinese chestnut photosynthesis.(5)In the vertical canopy, the Pn was significantly or extremely significantly and positively correlated with the single fruit mass, bur nut rate and yield per unit area, and the correlation coefficients were 0.872, 0.965 and 0.958, respectively. In the horizontal canopy, the Pn was significantly positively correlated with the single fruit mass and yield per unit area, and the correlation coefficients were 0.777 and 0.487, respectively. A comprehensive analysis of the horizontal and vertical canopy showed that the Pn was significantly or extremely significantly and positively correlated with the single fruit mass, bur nut rate and yield per unit area, with correlation coefficients of 0.600, 0.669 and 0.532, respectively.Conclusion The photosynthesis of Chinese chestnut has obvious spatial heterogeneity, and the difference between photosynthetic index and fruit yield is the result of synthesis of vertical and horizontal canopy. In Chinese chestnut production, pruning should be reasonable to improve the efficiency for solar energy utilization in the lower and middle part of canopy, so as to increase the fruit yield.
-
Keywords:
- Castanea mollissima /
- photosynthesis /
- canopy /
- spatial heterogeneity
-
在当前全球气候不断变暖的背景下,中国气候发展趋势变化明显,其升温速度远远超过了同期的全球平均水平[1];作为由中纬度向高纬度过渡的地区,东北地区在过去50年中平均温度每10年增加0.3 ℃[2]。长白山位于我国东北温带季风气候区,其森林面积大,生境条件多样,且受到人为活动干扰少,是研究气候变化对森林生态系统影响的理想区域[3],近年来该地区生态环境受气候变化的影响被广泛关注[2]。
树木的生长和发育与气候因子紧密相关,它可以对环境变化和极端气候产生长期的响应并将其记录在年轮宽度信息之中[4-5]。树木年轮由于具有定年准确、分辨率高、连续性强、分布广泛、易于采样等特点[6],已成为研究气候变化对树木生长的影响和重建过去气候变化的关键资源[7-8]。近年来,已有大量研究运用树木年轮学从不同角度探讨长白山森林生长对全球气候变化的响应。例如:高露双等[9]通过研究阔叶红松(Pinus koraiensis) 林建群树种的生长−气候关系指出,不同树种的径向生长与气候因子之间的关系有所不同;王守乐等[10]研究了同一生境下不同树种对季节性气候的响应及其动态关系;于健等[11]研究了长白山地区群落交错带不同树种的径向生长对气候变暖的响应,但大部分研究仍集中于阔叶红松林中共存的两个树种与气候因子之间的关系,还缺乏对于云冷杉针阔混交林中多个共存树种径向生长对气候变化响应的研究。
汪清地区是长白山云冷杉针阔混交林的重要分布区,已有众多学者对该地区云冷杉针阔混交林进行研究,包括林分结构[12]、进界模型[13]、林下更新[14]等多个方面。为了解该地区云冷杉针阔混交林中不同树种对气候因子的响应特征及升温后气候因子对不同树种生长的影响。本研究以吉林汪清金沟岭林场云冷杉针阔混交林为研究对象,对臭冷杉(Abies nephrolepis)、鱼鳞云杉(Picea jezoensis)和红松3种常见树种进行采样,建立树木年轮宽度标准年表,分析其对气候因子的响应特征,探讨不同树种径向生长与气候因子的关系及升温对其生长的影响,为该地区树木的保护提供依据。
1. 研究区概况与研究方法
1.1 研究区概况
研究区位于吉林省延边汪清林业局金沟岭林场(43°17′ ~ 43°25′N、130°05′ ~ 130°20′E),属长白山系老爷岭山脉雪岭支脉,经营面积16 286 hm2。全场四面环山,以低山丘陵地貌为主,海拔500 ~ 1 200 m,坡度多在 5° ~ 25°。林区属温带大陆性季风气候,年均温4 ℃左右,年降水量600 ~ 700 mm。林区属低山灰化土灰棕壤区,母岩为玄武岩。研究区内的主要树种有臭冷杉、鱼鳞云杉、红松、红皮云杉(Picea koraiensis)、白桦(Betula platyphylla)、落叶松(Larix gmelinii)、枫桦(Betula costata)、紫椴(Tilia amurensis)、花楷槭(Acer ukurunduense)、色木槭(Acer mono)、青楷槭(Acer tegmentosum)、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、黄檗(Phellodendron amurense)等;草本植物以苔草(Carex tristachya)和禾本科(Gramineae)为主。
1.2 样品的采集与处理
2019年8月在典型云冷杉天然林,海拔710 m,西北坡向,按照径级选取生长良好的臭冷杉、鱼鳞云杉和红松作为取样目标,利用口径为5.15 mm的树木生长锥在胸高(1.3 m)位置钻取年轮样芯(尽可能通过髓芯)。为减少对树木的损害,每株树仅钻取一根样芯。臭冷杉、鱼鳞云杉和红松分别钻取了65、25和55根样芯。将采集的样芯进行编号并记录每株树的生境信息。将样品带回实验室后,对年轮样芯进行风干和打磨等处理。打磨时用200目、400目、600目的砂纸依次打磨,直到年轮界限在显微镜下清晰可见。
1.3 建立年表
1.3.1 年表的制作方法
利用LINTABTM6.0年轮宽度测量仪(测量精度为0.001 mm)及配套的TSAP软件测量年轮宽度。利用 COFECHA 程序[15]对定年和测量结果进行检验,剔除与主序列相关性差以及难以交叉定年的样芯,将保留的样芯用于建立年表。为消除其他非气候因素导致的生长趋势,利用R语言“dplr”包中的ModNegExp方法对树轮宽度序列进行去趋势和标准化处理[16],建立3个树种标准年表。
![]() 图 3 3种常见树种的标准年表和样本量Figure 3. Standard chronology and sample size of three common tree species
图 3 3种常见树种的标准年表和样本量Figure 3. Standard chronology and sample size of three common tree species1.3.2 标准年表统计参数
年轮宽度指数、信噪比、样本一阶自相关系数、样本间相关系数、平均敏感度、树木年轮的标准差、样本总体代表性等见文献。
(1)年轮宽度指数。在建立树木年轮宽度年表时,将树木年轮宽度序列用统计学方法进行曲线拟合,得到树木生长的期望值(Yi),树木年轮实际宽度值(Wi)与期望值(Yi)的比值为年轮宽度指数(Ii)。
Ii=WiYi (1) (2)信噪比。信噪比(signal-to-noise ratio,RSN)用于表示样本中所包含环境信息多少的统计量,其值为树木年表中气候信息与非气候噪声之比,若数值越大,则年表中包含的气候信息越多。
RSN=N¯rbt1−¯rbt (2) 式中:N代表样本数,
¯rbt 代表不同样芯间的平均相关系数。(3)平均敏感度。平均敏感度(mean sensitivity,SM)用于衡量年轮宽度的逐年变化,是无量纲值,其值越高,树木生长对气候变化越敏感,受到气候因子的限制作用越强。
SM=1n−1n−1∑i=1|2(xi+1−xi)xi+1+xi| (3) 式中:xi和xi + 1分别是第i个和第i + 1个年轮的宽度值;n为该树轮样本的年轮总数。
(4)样本总体代表性。样本总体代表性(expressed population signal,SEP)表示采集的树轮样本能够代表采样点森林总体生长趋势的百分比值,是决定采样是否成功的一项指标,其值越大,建立的树轮年表越具有理论年表的特征,采样工作越成功。
SEP=¯rbt¯rbt+(1−¯rbt)/N (4) 样本总体代表性的阈值通常设定为0.85[17]。
1.4 气象数据
从中国气象科学数据共享网(http://cdc.cma.gov.cn/home.do)获取距离采样点最近的汪清气象站(43°18′N、129°47′E,海拔244.8 m)的气象资料,时间为60年(1958—2018 年)。气象要素包含逐月的最低气温、最高气温、平均气温和总降水量。根据气象站内的降水量和温度数据使用R软件的“SPEI”包来计算标准化降水蒸散指数(standardized precipitation evapotranspiration index,SPEI)。利用 Mann-Kendall(M-K)方法[18]对年平均气温进行突变分析。
1.5 统计分析
利用Pearson相关分析分别对臭冷杉、鱼鳞云杉及红松的径向生长对逐月气候因子的响应进行分析。考虑到前一年气候的条件可能对树木当年径向生长存在影响[19],故选用上一年6月至当年9月的气候数据用于分析。为研究升温突变(1985年)后树木生长与气候关系的变化,对1985年升温前后主要气候因子与臭冷杉、鱼鳞云杉、红松径向生长的关系进行比较。利用滑动相关分析探讨径向生长与气候因子的动态关系,滑动窗口为32年。利用R软件计算各年表的参数,所有相关分析利用DendroClim2002程序[20]进行,使用Origin 2019软件绘制图表。
2. 结果与分析
2.1 气候因子变化特征分析
该区域具有雨热同期的规律,最低平均气温出现在1月(约−18 ℃),最高平均气温在7月(约6 ℃)。降水集中在5—9月,最大降水量在8月,10 月份开始降水逐渐减少,12月份降水量最低(图1)。
![]() 图 1 研究区1958—2018年气象数据T.平均气温;Tmin.最低气温;Tmax.最高气温;SPEI.标准化降水蒸散指数。T, mean temperature; Tmin, minimum temperature; Tmax, maximum temperature; SPEI, standardized precipitation evapotranspiration index.Figure 1. Meteorological data of research area from 1958 to 2018
图 1 研究区1958—2018年气象数据T.平均气温;Tmin.最低气温;Tmax.最高气温;SPEI.标准化降水蒸散指数。T, mean temperature; Tmin, minimum temperature; Tmax, maximum temperature; SPEI, standardized precipitation evapotranspiration index.Figure 1. Meteorological data of research area from 1958 to 2018Mann-Kendall的检验结果为1978年后正常的统计值时间序列UFk > 0,表明近40年平均气温呈增加趋势,且1985年为年平均气温升高的突变点(图2)。1958—1985年年均气温为4 ℃,1986—2018年年均气温为4.65 ℃,比突变前增加了0.65 ℃。1958—1985年年均降水量为44.58 mm,1986—2018年年均降水量为47.51 mm,增温突变后的年均降水量大于突变前。1958—1985年期间,平均气温以0.01 ℃/a的速率上升,但变化趋势不显著,平均最低气温以0.04 ℃/a的速率显著上升,平均最高气温变化趋势不显著,降水量以0.44 mm/a的速率显著下降,标准化蒸散指数以0.01的速率显著下降。1986—2018年期间平均最低气温以0.03 ℃/a的速率显著下降,平均最高气温以0.04 ℃/a的速率显著上升,平均气温、降水量和标准化蒸散指数变化均不显著。总体而言,汪清地区在1958—2018年期间高温变高,低温变低,平均气温升高,降水量及干旱情况不变。
![]() 图 2 1958—2018年年均气温 Mann-Kendall检验结果UFk.正常的统计值时间序列 ;UBk.逆序的统计值时间序列。UFk, normal statistical time sequence; UBk, inverted statistical time sequence.Figure 2. Mann-Kendall test results of average annual temperature from 1958 to 2018
图 2 1958—2018年年均气温 Mann-Kendall检验结果UFk.正常的统计值时间序列 ;UBk.逆序的统计值时间序列。UFk, normal statistical time sequence; UBk, inverted statistical time sequence.Figure 2. Mann-Kendall test results of average annual temperature from 1958 to 20182.2 标准年表的统计特征
臭冷杉、鱼鳞云杉和红松年表整体变化趋势差异显著,特别是在1985年前后(图3)。1985年前臭冷杉年轮指数有明显下降趋势(−0.03 a−1,R2 = 0.77),鱼鳞云杉和红松年轮指数下降趋势较弱;1985年后臭冷杉年轮指数出现显著上升趋势(0.05 a−1,R2 = 0.89),鱼鳞云杉次之(0.04 a−1,R2 = 0.81),红松年轮指数上升趋势较弱。
如表1所示,3个树种年表序列长度的平均值为110年,其中红松年表的时间跨度最长,达120年(1898—2018年),臭冷杉、鱼鳞云杉年表时间跨度分别为118年(1900—2018年)、92年(1926—2018年)。平均敏感度(SM)为0.16 ~ 0.27,信噪比(RSN)为6.14 ~ 19.98,臭冷杉的值均为最大。3个树种的平均敏感度、信噪比等指标均较高,说明所建立的年表包含较多的气候信息,对气候变化敏感,适用于气候因子分析。一阶自相关系数为0.81 ~ 0.86,说明汪清地区臭冷杉、鱼鳞云杉、红松年轮受前一年气候因子影响较大。样本总体代表性(SEP)高达0.86 ~ 0.92,均超过了可接受的临界阈值,表明样本包含的信号基本代表当地的总体特征。3个树种相比,虽然臭冷杉标准年表的一阶自相关系数低于鱼鳞云杉,但其平均敏感度、标准差、信噪比、样本总体代表性均高于另外两个树种,说明臭冷杉更适用于气候研究。总体上,各项年表特征均表明,臭冷杉、鱼鳞云杉和红松年表均适用于树木年轮气候学分析。
表 1 标准年表的统计特征及公共区间分析Table 1. Statistical characteristics and common interval analysis of the standard chronology统计特征 Statistic characteristics 臭冷杉 Abies nephrolepis 鱼鳞云杉 Picea jezoensis 红松 Pinus koraiensis 时间跨度 Time span 1900—2018 1926—2018 1898—2018 平均值 Mean value 0.93 0.96 0.95 平均敏感度 Mean sensitivity (SM) 0.27 0.16 0.22 标准差 Standard deviation (SD) 0.40 0.38 0.30 一阶自相关系数 First-order autocorrelation coefficient (AC) 0.81 0.86 0.81 公共区间 Common interval 1969—2011 1969—2011 1959—2013 信噪比 Signal-to-noise ratio (RSN) 19.98 6.14 11.21 样本间相关系数 Correlation coefficient between samples (Rbar) 0.40 0.31 0.32 样本总体代表性 Sample population representativeness (SEP) 0.95 0.86 0.92 2.3 不同树种径向生长对气候因子的响应特征
由图4、5可知,臭冷杉、鱼鳞云杉和红松径向生长均与上年生长季末期(9月)平均气温及上年、当年7月最低气温呈显著正相关(P < 0.05);鱼鳞云杉和红松与降水、标准化降水蒸散指数(SPEI)无显著性关系(P > 0.05);臭冷杉和红松与生长季前期(4—5月)最高气温呈显著正相关(P < 0.05)。此外臭冷杉径向生长与当年7月和上年7月平均气温呈显著正相关(P < 0.05),与当年5月和上年11月降水、SPEI呈显著正相关(P < 0.05);鱼鳞云杉与上年6月平均气温呈显著正相关(P < 0.05),与当年5月最高气温呈显著正相关(P < 0.05)。这表明臭冷杉年表较鱼鳞云杉和红松年表对气候因子响应更加敏感。臭冷杉径向生长受到气温和降水的制约,而鱼鳞云杉和红松径向生长的主要限制因子为温度。
![]() 图 4 标准年表与月最低气温、平均气温和最高气温的相关系数p.上年;c.当年。下同。p, previous year; c, current year. The same below.Figure 4. Correlation coefficients of the standard chronology with monthly minimum temperature, mean temperature and maximum temperature
图 4 标准年表与月最低气温、平均气温和最高气温的相关系数p.上年;c.当年。下同。p, previous year; c, current year. The same below.Figure 4. Correlation coefficients of the standard chronology with monthly minimum temperature, mean temperature and maximum temperature![]() 图 5 标准年表与月降水量、SPEI的相关系数Figure 5. Correlation coefficients of standard chronology with monthly precipitation and SPEI
图 5 标准年表与月降水量、SPEI的相关系数Figure 5. Correlation coefficients of standard chronology with monthly precipitation and SPEI2.4 不同树种径向生长对升温的响应差异
由于汪清地区的气候因子(气温和降水量)在1985年前后具有明显差别(图1、2),因此分两个时间段(1985—1985年和1986—2018年)分别评估温度和降水量对3个树种生长的影响。由图6可知,1958—1985年,臭冷杉和红松与上年9月、当年6月降水量呈显著正相关(P < 0.05),鱼鳞云杉与上年8、9月降水量呈显著正相关(P < 0.05),各树种与平均气温均未达到显著性水平(P > 0.05)。1986—2018年,臭冷杉径向生长与当年4月和7月平均气温呈显著正相关(P < 0.05),鱼鳞云杉径向生长与平均气温均未达到显著水平(P > 0.05),红松径向生长与当年4月平均气温呈显著正相关(P < 0.05),各树种与降水量均未达到显著性水平(P > 0.05)。由此可见,升温突变前后臭冷杉、鱼鳞云杉和红松径向生长与气候因子相关性均发生改变,主要表现为升温突变后径向生长与气温相关性增强但是与降水量相关性减弱。
![]() 图 6 升温突变前后3种常见树种标准年表与气候因子的相关分析*表示的显著相关(P < 0.05)。* means significant correlation (P < 0.05).Figure 6. Correlation analysis between the standard chronology of three common tree species and climate factors before and after the abrupt warming change
图 6 升温突变前后3种常见树种标准年表与气候因子的相关分析*表示的显著相关(P < 0.05)。* means significant correlation (P < 0.05).Figure 6. Correlation analysis between the standard chronology of three common tree species and climate factors before and after the abrupt warming change2.5 不同树种径向生长−气候关系的时间稳定性
由图7可知,上年6月和当年4、6月平均气温对臭冷杉径向生长的影响具有一定的长期稳定性。臭冷杉标准年表与上年9月、当年9月平均气温由显著负相关转为显著正相关,与当年6、7月平均气温由不显著相关转为显著正相关,说明夏季气温对臭冷杉生长的影响逐渐增强,秋季气温对臭冷杉径向生长由抑制作用转为促进作用。气温对红松的影响与臭冷杉和鱼鳞云杉相比,抑制作用较少。当年7月降水量与臭冷杉由显著负相关转为不显著相关,而与鱼鳞云杉和红松由不显著相关转为显著正相关。当年9月降水量对红松由促进作用变为抑制作用。上年6、7、12月和当年6、7、9月降水量对臭冷杉径向生长具有较明显的抑制作用,尤其是上年6月,显著相关关系持续时间长达11年(2005—2016)。
![]() 图 7 3种常见树种与气候因子的滑动相关分析Figure 7. Moving correlation analysis of three common tree species and climate factors
图 7 3种常见树种与气候因子的滑动相关分析Figure 7. Moving correlation analysis of three common tree species and climate factors3. 讨 论
3.1 臭冷杉、鱼鳞云杉和红松径向生长−气候关系差异
研究结果表明:臭冷杉、鱼鳞云杉和红松径向生长与均与上年9月平均气温呈显著正相关,这主要因为树木的生长集中在6—9月,上年9月为上个生长季的末期,而当上个生长季结束时较高的温度有利于延长生长季,保证树木正常的代谢活动,使树木能够储存更多的营养物质以供下一个生长季开始时树木的生长,从而表现出促进作用[21]。3个树种径向生长均与当年7月最低气温呈显著正相关,这可能是因为研究区在7、8月降水集中,水分充足,有利于增强植物的蒸腾作用,促进叶片气孔开放和CO2吸收,在水分充足的条件下,生长季的温度升高有助于提高酶的活性,增加细胞生产速率,进而促进树木径向生长[22]。以上结果表明同一区域不同树种径向生长对气候的响应具有一定的相似性,与杨婧雯等[23]的研究结果一致,这可以解释为树木对区域气候信号的共同响应[24]。
本研究中,臭冷杉径向生长与生长季前期降水有较强相关性,生长季前期(当年5月)降水量的增加,不仅可以有效补充土壤水分,缓解生长季温度过高造成的水分不足,还可以促进细胞运输营养物质和合成蛋白质,从而促进宽轮的形成[25]。树木需要通过根系从土壤吸收足够的水分来维持自身生理活动,作为喜湿、耐荫、浅根性树种,臭冷杉的生长很容易受到水分胁迫[24],因此臭冷杉生长受温度和降水的共同影响。鱼鳞云杉径向生长与5月最高气温均呈显著正相关,可能是因为5月温度的升高不仅可以促进地面积雪的融化,温暖土壤和茎分生组织,使土壤早日解冻,还可以提高光合作用,增加细胞生产速率,进而促进早期树木的生长[26-27]。红松径向生长与当年4月最高气温呈显著正相关,春季树木结束休眠转入生长季,较高的生长季前期温度可以促进生长季提前开始,从而延长生长季,而适宜的温度可以促进树木形成层活动,因此生长季前期足够的热量有利于树木形成宽轮[24]。该地区不同树种的生长与气候因子之间的关系存在一定差异可能是受树种遗传特性的影响[28]。此外,韩艳刚等[29]对长白山关于径向生长−气候关系已发表的文献总结的结果也显示长白山地区不同树种对气候因子的响应有所不同,在川西高原[30]、大兴安岭[23]等地的研究同样表明不同树种对气候因子的响应有差异。
3.2 气候变暖对不同树种径向生长的影响
本研究结果显示:升温突变(1985年)后,臭冷杉、鱼鳞云杉和红松径向生长与气温的正相关性增加,即升温突变后3个树种径向生长对气温的敏感性增加,且其径向生长具有与气温升高相一致的趋势;这与Andreu等[24]研究发现的树木生长随着气温升高对气候的敏感性增加的结果一致,而与王守乐等[10]研究显示的树木径向对气候的敏感性降低的结果不同。目前,已有很多研究提出气候变暖对树木径向生长影响。树木的径向生长随着气候变暖可能出现加快[31]、降低[32]、及变化不明显[33]等多种情况。这也验证了气候变暖后树木径向生长对气候因子的响应存在差异。
此外,结果显示,气温升高有利于3个树种的径向生长,这与于健等[11]在位于1 125 m海拔的研究地得出的气候变暖抑制鱼鳞云杉生长的研究结果不同。通常情况下,海拔可以改变树木的生长速度,进而影响树木生长与气候的关系。Zhu等[34]研究表明鱼鳞云杉对气候变暖的响应存在海拔差异,快速升温加速了高海拔鱼鳞云杉的径向生长,而减少了低海拔鱼鳞云杉的径向生长。但是本研究在海拔低于于健等[11]研究的情况下气候变暖仍然促进树木径向生长,这可能是由于气温的阈值效应,树木生长之所以会出现对气候因子响应的敏感性变化主要是因为该效应,气温的升高满足了树木生长的下限阈值,而逐渐突破了树木生长所需要的上限阈值[35]。当气温高于树木生长的上限阈值后,会使其蒸发、蒸腾作用加剧,从而导致土壤水分有效性降低,此时水分可能变成限制因素,出现干旱胁迫现象[31]。目前升温可能未超过臭冷杉、鱼鳞云杉和红松径向生长的临界温度,且由于汪清地区降水量充足(图1),故升温并未对该地区云冷杉林造成干旱胁迫,进而表现出对其径向生长的促进作用。李波[13]在2017年对汪清地区云冷杉林的研究中同样发现该地区年均温促进树木的径向生长。即使目前汪清地区树木生长并未受到气温升高的抑制作用,但是随着未来气候的持续变暖,该地区树木生长对气候的响应仍需持续关注。
4. 结 论
本研究表明,相比较而言,臭冷杉标准年表统计参数值高于鱼鳞云杉和红松,其对气候变化的响应更敏感,更适用于树轮气候学的研究。长白山汪清地区臭冷杉、鱼鳞云杉和红松径向生长对于特定气候因子的响应有一致性,上年生长季末期(9月)平均气温及上年、当年7月最低气温促进3个树种径向生长。同时,该地区树木径向生长−气候因子之间的关系也有着显著的差异,臭冷杉生长受气温和降水的制约,鱼鳞云杉和红松生长的主要限制因子为温度。这可能主要是因为树种的生理特征以及种间遗传因素存在差异。1985年升温前树木径向生长有显著下降趋势,升温后则出现上升趋势。升温突变后,臭冷杉、鱼鳞云杉和红松与气温和降水的相关性变化存在明显差异,气温对树木径向生长的促进作用增加而降水的促进作用减弱。总体而言,目前气温的升高促进汪清地区云冷杉林常见树种的径向生长。
-
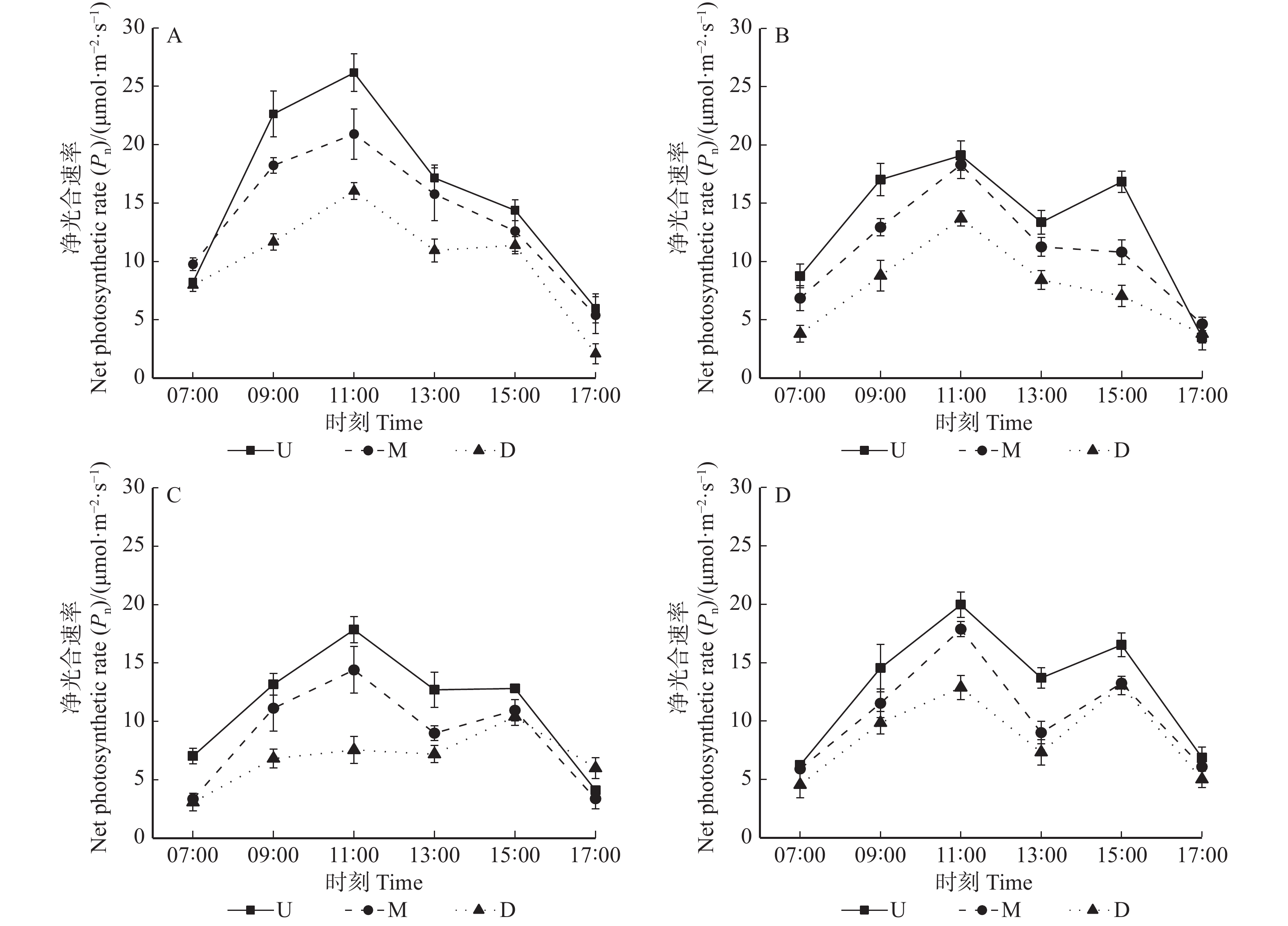
![]()
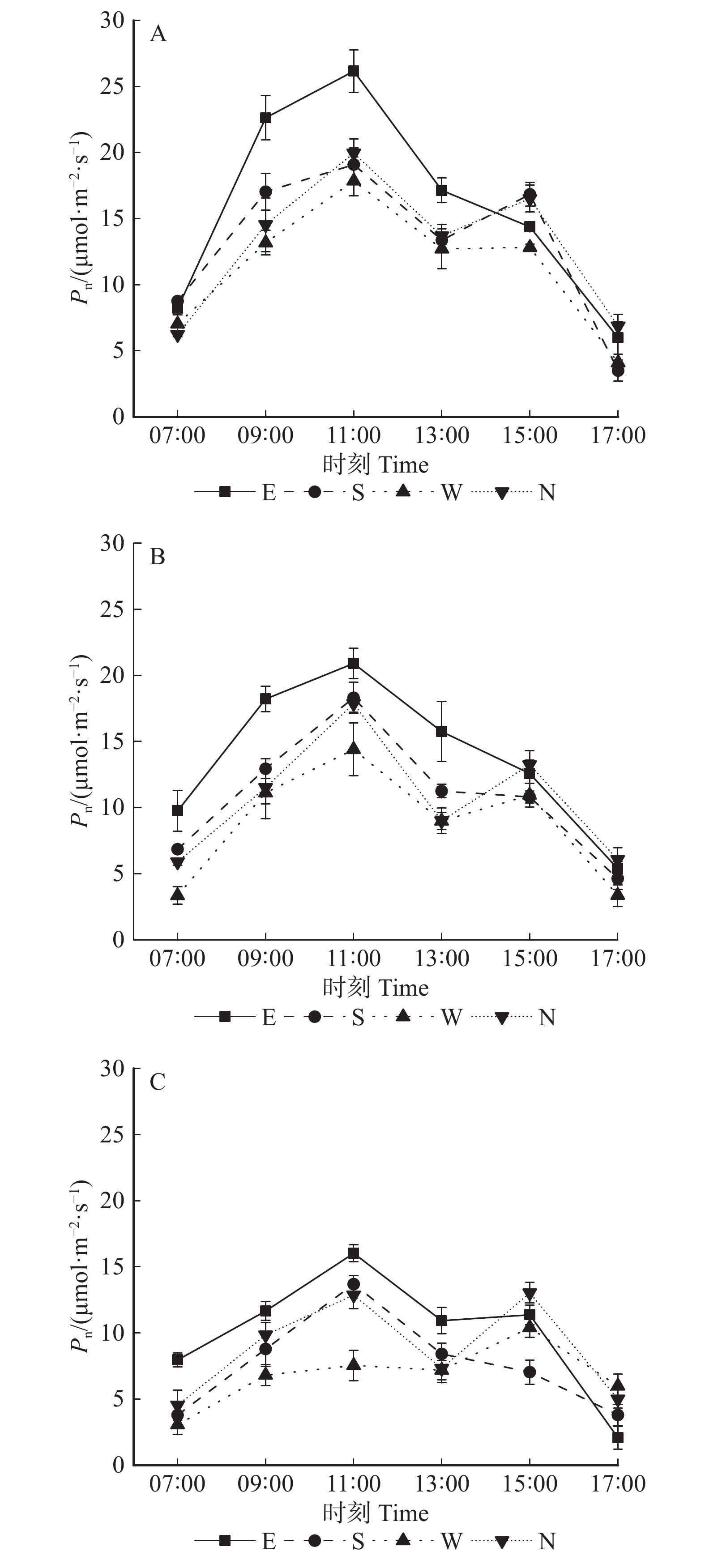
图 1 垂直冠层内净光合速率在不同方位条件下的日变化
A、B、C、D分别表示东、南、西、北方向下上、中、下冠层的净光合速率日变化;图中字母U、M、D分别指树冠上部、中部、下部。下同。A, B, C and D show the daily changes of Pn in the upper, middle, and lower canopies under the conditions of east, south, west, and north, respectively;U, M, D refer to the upper, middle and lower canopy, respectively. The same below.
Figure 1. Diurnal changes of Pn in vertical canopy under different directions
![]()
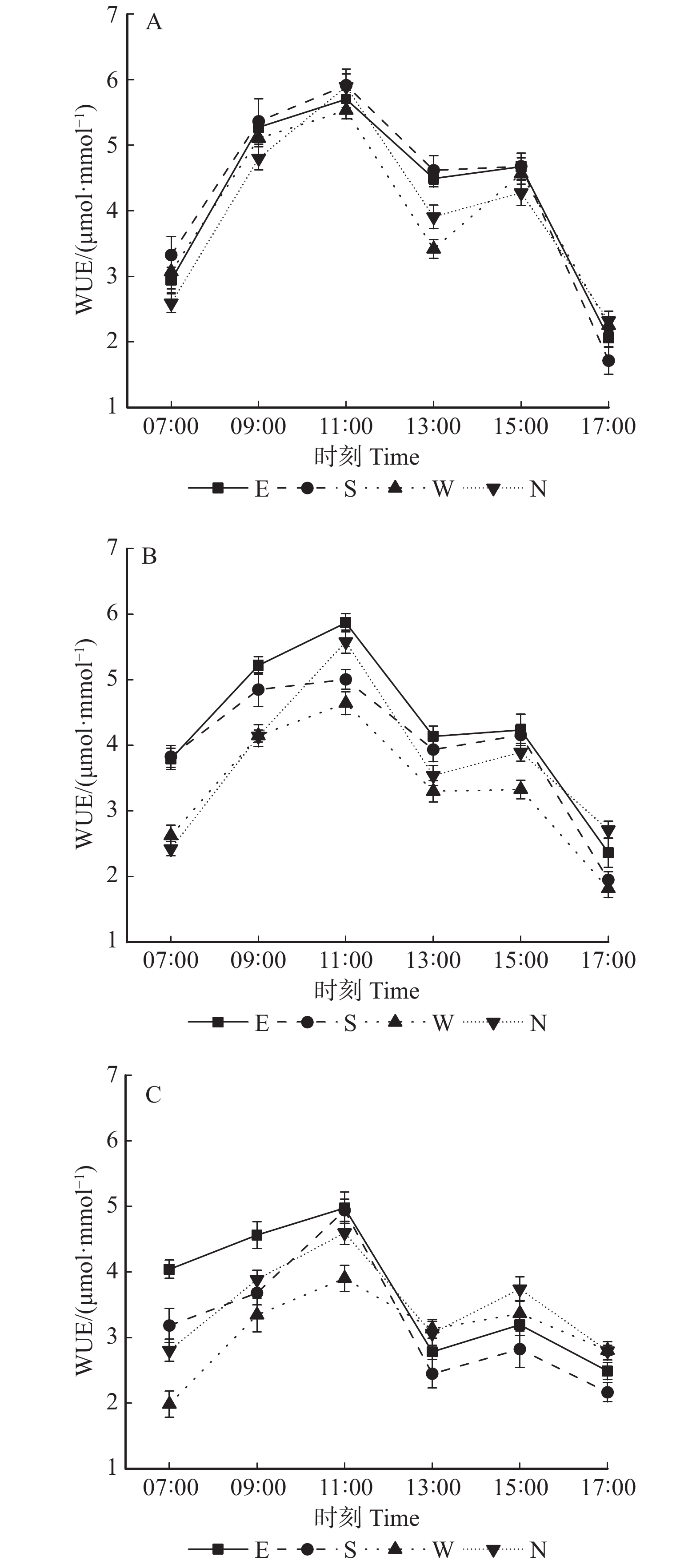
图 2 同一冠层不同方位的净光合速率日变化
A、B、C分别表示上、中、下冠层在东西南北方向的净光合速率日变化;图中字母E、S、W、N分别指树冠东向、树冠南向、树冠西向、树冠北向。下同。A, B and C show the daily changes in the net photosynthetic rate of the east, west, south, and north directions in the upper, middle, and lower canopies, respectively; E, S, W, N refer to the east, south, west and north direction of the canopy, respectively. The same below.
Figure 2. Diurnal changes of Pn in different directions at the same canopy
![]()
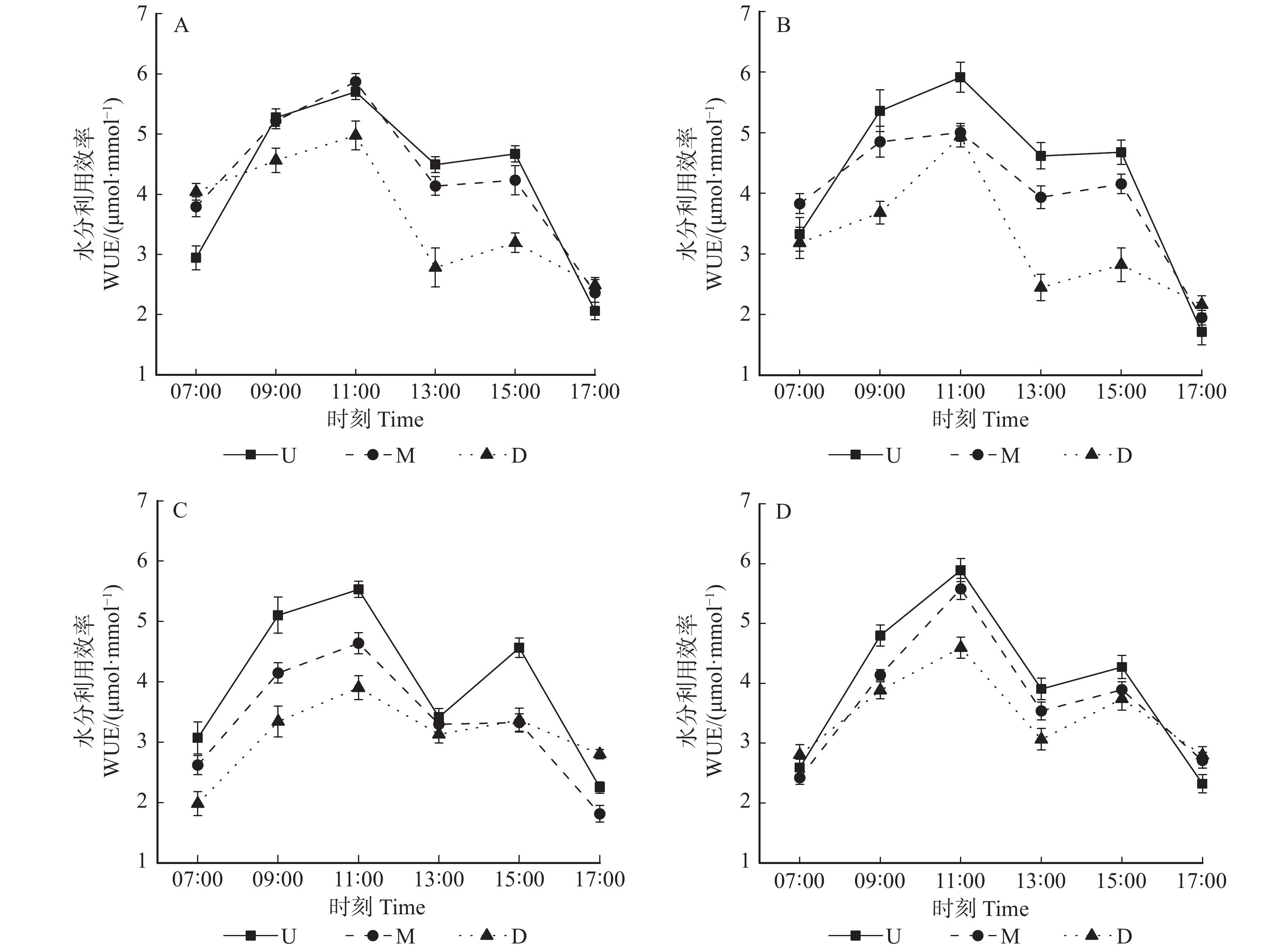
图 3 垂直冠层内水分利用效率在不同方位的日变化
A、B、C、D分别表示东、南、西、北方向下上、中、下冠层的水分利用速率日变化。A, B, C and D show the daily changes of the WUE in the upper, middle, and lower canopies of the east, south, west, and north directions, respectively.
Figure 3. Diurnal variations of WUE in vertical canopy under different directions
![]()
图 4 同一冠层不同方位的水分利用效率日变化
A、B、C分别表示上、中、下冠层条件下东、西、南、北方向的水分利用效率日变化。A, B and C show the daily changes of the WUE in the east, west, south and north directions of the upper, middle, and lower canopies, respectively.
Figure 4. Diurnal changes of WUE in different directionsat the same canopy
![]()
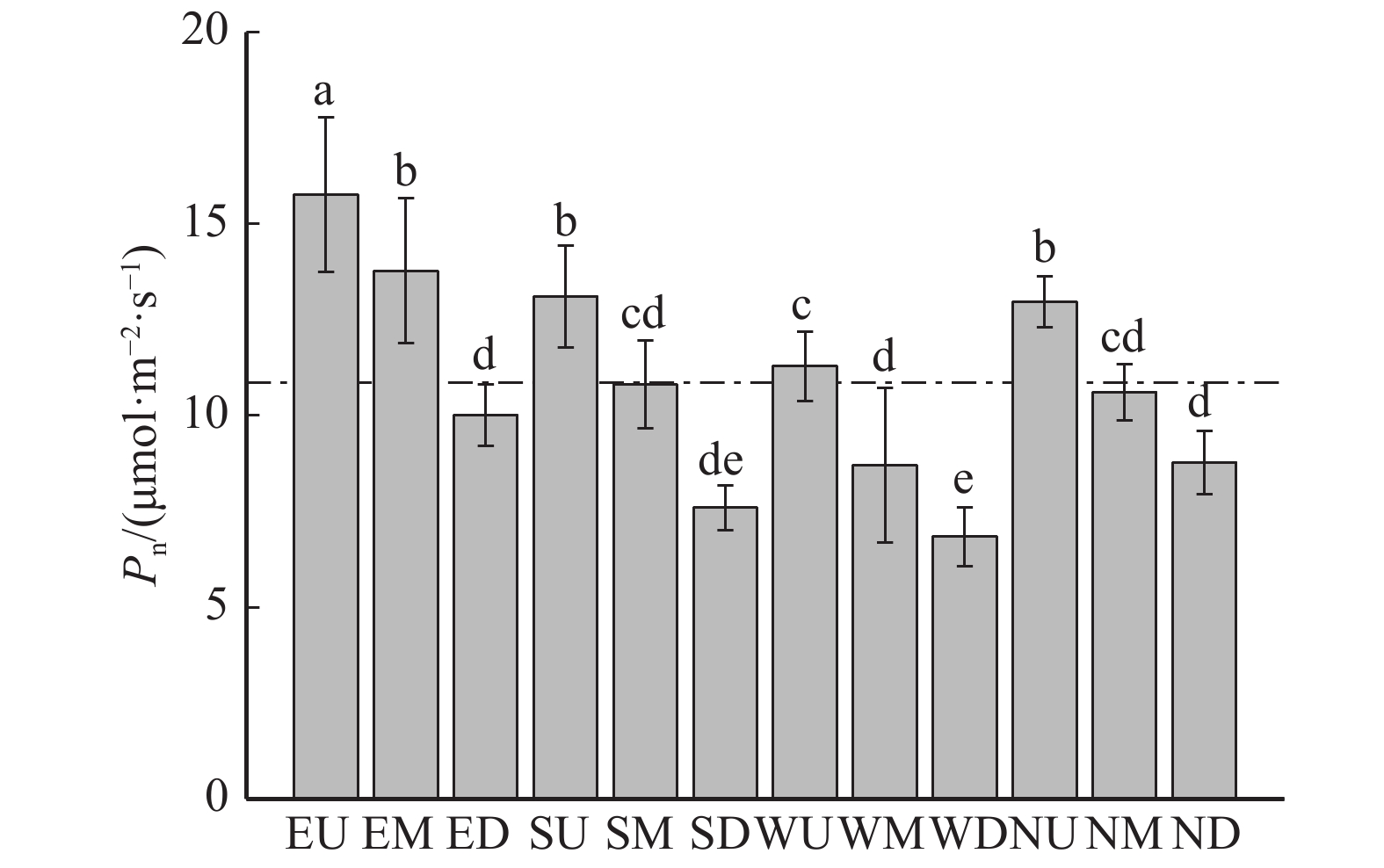
图 5 冠层不同区域净光合速率的分析对比
虚线表示冠层平均光合速率。E、S、W、N 分别代表东、南、西、北,U、M、D 分布代表冠层上部、冠层中部、冠层下部,下同。The dotted line in the figure indicates the average canopy photosynthetic rate. E, S, W and N represent east, south, west and north, while distribution of U, M and D represent upper, middle and lower canopy, the same below.
Figure 5. Analysis of net photosynthetic rate in different regions of the canopy
![]()
图 6 不同层次、不同方位的相对叶绿素含量及比叶面积分析对比
Figure 6. Analysis and comparison of SPAD and SLA at different canopies and directions
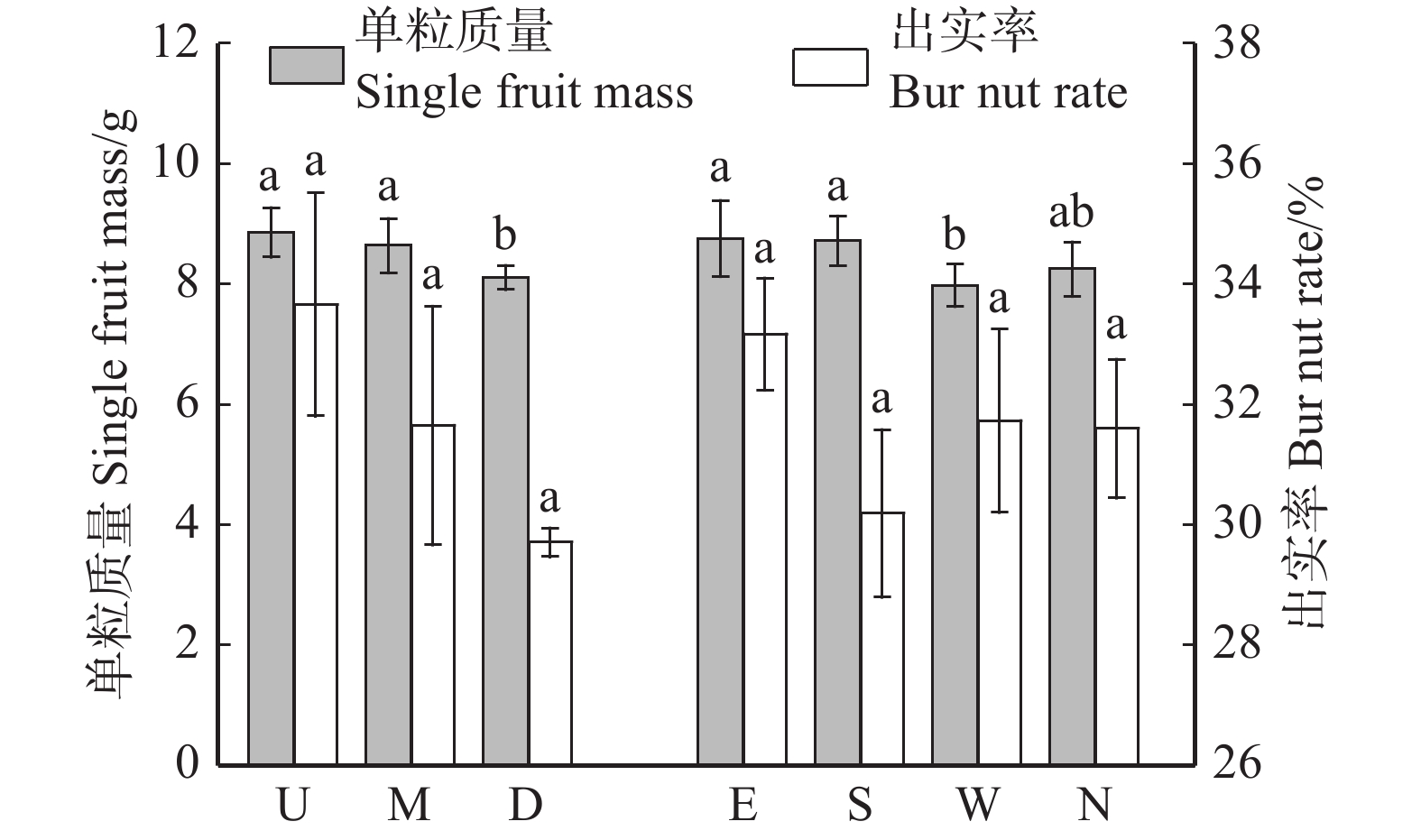
![]()
图 7 板栗果实单粒质量及出实率在垂直冠层和不同方位的差异性
Figure 7. Differences in single fruit mass and bur nut rate between vertical canopies and different directions
![]()
图 8 板栗果实单位面积产量在垂直冠层和水平冠层的差异性
Figure 8. Differences of yield per unit area of Chinese chestnut fruit in vertical and horizontal canopies
表 1 板栗光合测定的树体特征
Table 1 Characteristics of sample trees for measuring photosynthesis of Chinese chestnut
m 样木编号
Sample
tree No.主干高
Trunk
height树高
Tree
height冠高
Canopy
height冠幅
Canopy
width分层宽度
Length of
each layer1 0.40 2.61 2.21 2.56 0.85 2 0.44 2.79 2.35 2.56 0.85 3 0.42 2.68 2.26 2.60 0.87 4 0.45 2.75 2.30 2.54 0.85 5 0.43 2.60 2.17 2.44 0.81  下载: 导出CSV
下载: 导出CSV
表 2 板栗不同方位光合生理指标的比较
Table 2 Comparison of photosynthetic physiological indexes in different directions of Chinese chestnut
光合生理生态指标
Physiological and ecological index of photosynthesis方向 Direction 东 East 南 South 西 West 北 North 净光合速率 Net photosynthetic rate (Pn)/(μmol·m−2·s−1) 13.18 ± 2.95a 10.50 ± 2.54b 8.94 ± 2.30c 10.78 ± 1.90b 气孔导度 Stomatal conductance (Gs)/(mol·m−2·s−1) 0.18 ± 0.05a 0.15 ± 0.02b 0.14 ± 0.02b 0.15 ± 0.04b 胞间CO2浓度 Intercellular CO2 concentration (Ci)/(μmol·mol−1) 286.67 ± 22.00a 272.17 ± 18.24b 263.60 ± 10.53b 251.00 ± 18.41c 蒸腾速率 Transpiration rate (Tr)/(mmol·m−2·s−1) 3.12 ± 0.54a 2.64 ± 0.35bc 2.47 ± 0.39c 2.80 ± 0.63b 水汽压亏缺 Vapor pressure deficit (VpdL)/ kPa 1.89 ± 0.18a 2.05 ± 0.12b 2.08 ± 0.14b 2.10 ± 0.19b 气温 Air temperature (Ta)/ ℃ 31.67 ± 0.50a 31.68 ± 0.44a 31.98 ± 0.48ab 32.12 ± 0.98b 叶温 Leaf temperature (Tl)/ ℃ 31.52 ± 0.52a 31.66 ± 0.56ab 31.87 ± 0.49ab 32.01 ± 0.99b 大气CO2浓度 Atmospheric CO2 concentration (Ca)/(μmol·mol−1) 441.44 ± 20.25a 415.50 ± 8.62b 409.12 ± 8.21c 414.70 ± 3.94b 空气相对湿度 Relative air humidity (RH)/% 52.15 ± 2.03a 50.66 ± 1.37b 50.37 ± 1.74b 50.82 ± 3.07b 光合有效辐射 Photosynthetically active radiation (PAR)/(μmol·m−2·s−1) 731.63 ± 58.24a 747.22 ± 64.59a 749.90 ± 31.65b 703.40 ± 56.30b 气孔限制值 Stomatal limitation (Ls) 0.350 ± 0.05a 0.345 ± 0.04a 0.356 ± 0.02a 0.395 ± 0.04b 水分利用效率 Water use efficiency (WUE) 4.20 ± 0.52a 3.97 ± 0.79ab 3.65 ± 0.90b 3.99 ± 0.86ab 羧化效率 Carboxylation efficiency (Vc) 0.046 ± 0.01a 0.040 ± 0.01b 0.033 ± 0.01c 0.043 ± 0.01a 注:不同小写字母代表各光合生理生态因子在不同处理间差异显著(P < 0.05)。下同。Notes: different lowercase letters represent significant differences among photosynthetic physiological and ecological factors in different treatments (P < 0.05). The same below.
下载: 导出CSV
表 3 板栗不同冠层光合生理指标的比较
Table 3 Comparison of photosynthetic physiological indexes in different canopies of Chinese chestnut
光合生理生态指标
Physiological and ecological index of photosynthesis冠层 Canopy 上 Upper 中 Middle 下 Lower Pn/(μmol·m−2·s−1) 13.28 ± 2.11a 10.97 ± 2.41b 8.30 ± 1.43c Gs/(mol·m−2·s−1) 0.18 ± 0.04a 0.15 ± 0.03b 0.13 ± 0.02c Ci/(μmol·mol−1) 258.75 ± 23.68a 271.40 ± 20.19b 274.93 ± 18.53b Tr/(mmol·m−2·s−1) 3.11 ± 0.45a 2.76 ± 0.49b 2.41 ± 0.46c VpdL/kPa 1.95 ± 0.19a 2.02 ± 0.15b 2.13 ± 0.15c Ta/℃ 31.90 ± 0.62a 31.82 ± 0.59a 31.86 ± 0.76a Tl/℃ 31.77 ± 0.64a 31.69 ± 0.62a 31.83 ± 0.81a Ca/(μmol·mol−1) 423.15 ± 20.96a 419.65 ± 17.52a 417.78 ± 11.93a RH/% 51.29 ± 2.19a 51.14 ± 1.85a 50.57 ± 2.58a PAR/(μmol·m−2·s−1) 757.88 ± 34.19a 723.43 ± 61.08b 678.23 ± 62.78c Ls 0.389 ± 0.05a 0.353 ± 0.04b 0.364 ± 0.04b WUE 4.29 ± 0.53a 4.02 ± 0.83a 3.54 ± 0.82b Vc 0.052 ± 0.01a 0.040 ± 0.01b 0.030 ± 0.01c
下载: 导出CSV
表 4 生理生态因子对板栗净光合速率的通径分析结果
Table 4 Path analysis results of Chinese chestnut net photosynthetic rate with physiological and ecological factors
因子
Factor简单相关系数
Simple correlation
coefficient直接通径系数
Direct path
coefficient间接通径系数 Indirect path coefficient 决策系数
Decision
coefficientVc Ci Gs WUE Tr VpdL Ta 总计 Total Vc 0.904 5 0.178 −0.020 4 0.018 8 0.310 2 0.358 6 0.056 6 0.002 7 0.726 5 0.290 3 Ci 0.0650 0.074 −0.049 1 0.005 5 −0.059 2 0.077 8 0.029 4 −0.013 4 −0.009 0 0.004 1 Gs 0.7626 0.029 0.115 5 0.014 0 0.088 2 0.441 0 0.069 1 0.005 7 0.733 6 0.043 4 WUE 0.5828 0.548 0.100 7 −0.008 0 0.004 7 −0.072 6 0.051 0 −0.041 0 0.034 8 0.338 4 Tr 0.7200 0.572 0.111 6 0.010 1 0.022 4 −0.069 6 0.041 0 0.032 5 0.148 0 0.496 5 VpdL −0.5450 −0.135 −0.074 6 −0.016 1 −0.014 8 −0.207 1 −0.173 9 0.076 6 −0.410 0 0.128 9 Ta −0.0270 0.108 0.004 5 −0.009 2 0.001 5 −0.208 2 0.172 2 −0.095 7 −0.135 0 −0.017 5
下载: 导出CSV
表 5 果实单粒质量、出实率及单位面积产量与光合指标的相关性
Table 5 Correlations between single fruit mass, bur nut rate and yield per unit area of fruit and photosynthetic index
冠层类型 Canopy type 指标 Index Pn Gs Ci Tr WUE 垂直冠层
Vertical canopy单粒质量 Single fruit mass 0.872* −0.799 0.946 0.986* 0.061 出实率 Bur nut rate 0.965** −0.921* 0.996 0.917 0.301 单位面积产量 Yield per unit area 0.958** −0.988 0.892 0.569 0.759** 水平冠层
Horizontal canopy单粒质量 Single fruit mass 0.777* 0.720 0.728 0.668 0.842** 出实率 Bur nut rate 0.610 0.720 0.416 0.687 0.393 单位面积产量 Yield per unit area 0.487* 0.527 0.849 0.337 0.499 水平 × 垂直
Horizontal canopy × vertical canopy单粒质量 Single fruit mass 0.600* 0.363 0.206 0.490 0.699* 出实率 Bur nut rate 0.669* 0.536 0.112 0.654* 0.606* 单位面积产量 Yield per unit area 0.532** 0.460 0.572* 0.441 0.498*
下载: 导出CSV
-
[1] 夏国威, 孙晓梅, 陈东升, 等. 日本落叶松冠层光合特性的空间变化[J]. 林业科学, 2019, 55(6):13−21. doi: 10.11707/j.1001-7488.20190602. Xia G W, Sun X M, Chen D S, et al. Spatial variation of photosynthetic characteristics in canopy of Larix kaempferi[J]. Scientia Silvae Sinicae, 2019, 55(6): 13−21. doi: 10.11707/j.1001-7488.20190602.
[2] 夏国威, 陈东升, 孙晓梅, 等. 日本落叶松冠层光合生理参数的空间异质性研究[J]. 林业科学研究, 2018, 31(6):130−137. Xia G W, Chen D S, Sun X M, et al. Spatial heterogeneity of photosynthetic and physiological parameters in Larix kaempferi crown[J]. Forest Research, 2018, 31(6): 130−137.
[3] Hirose T, Werger M J A. Maximizing daily canopy photosynthesis with respect to the leaf nitrogen allocation pattern in the canopy[J]. Oecologia, 1987, 72(4): 520−526. doi: 10.1007/BF00378977.
[4] Sakowska K, Alberti G, Genesio L, et al. Leaf and canopy photosynthesis of a chlorophyll deficient soybean mutant[J]. Plant Cell and Environment, 2018, 41(6): 1427−1437. doi: 10.1111/pce.13180.
[5] O’Rourke P A, Terjung W H. Relative influence of city structure on canopy photosynthesis[J]. International Journal of Biometeorology, 1981, 25(1): 1−19. doi: 10.1007/BF02184432.
[6] Kurachi N, Hagihara A, Hozumi K. Canopy photosynthetic production in a Japanese larch forest (Ⅱ): estimation of the canopy photosynthetic production[J]. Ecological Research, 1993, 8(3): 349−361. doi: 10.1007/BF02347194.
[7] Suwa R, Hagihara A. Seasonal changes in canopy photosynthesis and foliage respiration in a Rhizophora stylosa stand at the northern limit of its natural distribution[J]. Wetlands Ecology and Management, 2007, 16(4): 313−321.
[8] 胡耀升, 么旭阳, 刘艳红. 辽东栎冠层光合生理特性的空间异质性[J]. 生物学杂志, 2014, 31(1):33−36. doi: 10.3969/j.issn.2095-1736.2014.01.033. Hu Y S, Yao X Y, Liu Y H. Spatial heterogeneity of photosynthetic characteristics of Quercus wutaishanica canopy[J]. Journal of Biology, 2014, 31(1): 33−36. doi: 10.3969/j.issn.2095-1736.2014.01.033.
[9] 刘强, 董利虎, 李凤日, 等. 长白落叶松冠层光合作用的空间异质性[J]. 应用生态学报, 2016, 27(9):2789−2796. Liu Q, Dong L H, Li F R, et al. Spatial heterogeneity of canopy photosynthesis for Larix olgensis[J]. Chinese Journal of Applied Ecology, 2016, 27(9): 2789−2796.
[10] 张赟齐, 高世轮, 卫星杓, 等. 光合模型对无患子叶片光合响应参数计算结果的影响[J]. 北京林业大学学报, 2019, 41(4):32−40. Zhang Y Q, Gao S L, Wei X B, et al. Effects of photosynthetic models on the calculation results of photosynthetic response parameters in Sapindus mukorossi leaves[J]. Journal of Beijing Forestry University, 2019, 41(4): 32−40.
[11] 李六林, 季兰. 欧榛光合特性及影响因子的研究[C]//中国园艺学会第七届青年学术讨论会论文集. 北京: 中国园艺学会, 2006: 52−57. Li L L, Ji L. Studies on photosynthetic characteristics and influencing factors of Corylus avella[C]//Proceedings of the 7th Youth Symposium of Chinese Horticultural Society. Beijing: Chinese Society for Horticultural Society, 2006: 52−57.
[12] 牛莹莹, 廖康, 赵世荣, 等. 不同栽植密度库尔勒香梨冠层光合有效辐射变化规律研究[J]. 新疆农业科学, 2016, 53(3):420−428. Niu Y Y, Liao K, Zhao S R, et al. Study on PAR change rule of Korla fragrant pear canopy in different planting densities[J]. Xinjiang Agricultural Sciences, 2016, 53(3): 420−428.
[13] 张继祥, 魏钦平, 张静, 等. 苹果密植园与间伐园树冠层内叶片光合潜力比较[J]. 应用生态学报, 2009, 20(12):2898−2904. Zhang J X, Wei Q P, Zhang J, et al. Leaf photosynthetic potential in canopy layers of un-thinned and thinned apple orchards[J]. Chinese Journal of Applied Ecology, 2009, 20(12): 2898−2904.
[14] 周连第, 兰彦平, 韩振海. 板栗品种资源分子水平遗传多样性研究[J]. 华北农学报, 2006, 21(3):81−85. doi: 10.3321/j.issn:1000-7091.2006.03.019. Zhou L D, Lan Y P, Han Z H. Study on heritance diversity of Chinese chestnut (Castanea mollissima) variety resources at molecula level[J]. Acta Agriculturae Boreali-Sinica, 2006, 21(3): 81−85. doi: 10.3321/j.issn:1000-7091.2006.03.019.
[15] 张丽, 郭素娟, 孙慧娟, 等. 硼砂和蔗糖对板栗果实非结构性碳水化合物含量的影响[J]. 果树学报, 2018, 35(3):319−325. Zhang L, Guo S J, Sun H J, et al. Effects of borax and sucrose on the non-structural carbohydrate content in Chinese chestnut fruit[J]. Journal of Fruit Science, 2018, 35(3): 319−325.
[16] 张俊杰, 陈宗游, 韩愈, 等. 不同砧木嫁接对‘桂栗一号’大果锥栗苗木生长及光合特性的影响[J]. 中国农业科技导报, 2018, 20(3):10−19. Zhang J J, Chen Z Y, Han Y, et al. Effects of different rootstocks on growth and photosynthetic characteristics of large-fruit Castanen henryi ‘Guili 1’[J]. Journal of Agricultural Science and Technology, 2018, 20(3): 10−19.
[17] 田寿乐, 孙晓莉, 沈广宁, 等. 尿素与磷酸二氢钾配施对板栗光合特性及生长结实的影响[J]. 应用生态学报, 2015, 26(3):747−754. Tian S L, Sun X L, Shen G N, et al. Effects of compound fertilizer of (NH2)2CO and KH2PO4 on the chestnut photosynthesis characteristics, growth and fruiting[J]. Chinese Journal of Applied Ecology, 2015, 26(3): 747−754.
[18] 陈锦璞. 锰对‘石门早硕’板栗光合特性及矿质营养的影响[D]. 泰安: 山东农业大学, 2013. Chen J P. Effect of MnSO4 on photosynthetic characteristics and mineral nutrition in ‘Shi-men-zao-shuo’ chestnut tree[D]. Taian: Shandong Agriculture University, 2013.
[19] 彭方仁, 黄宝龙. 密植板栗树光合特性的研究[J]. 浙江农林大学学报, 1997, 14(2):151−154. Peng F R, Huang B L. Photosynthetic characteristics of chestnuts in high density planting orchards[J]. Journal of Zhejiang Forestry College, 1997, 14(2): 151−154.
[20] 张亦弛, 郭素娟, 孙传昊. 生长延缓剂对板栗叶片解剖结构及非结构性碳水化合物的影响[J]. 北京林业大学学报, 2020, 42(1):46−53. doi: 10.12171/j.1000-1522.20180437. Zhang Y C, Guo S J, Sun C H. Effects of growth retardants on anatomy and non-structural carbohydrates of chestnut leaves[J]. Journal of Beijing Forestry University, 2020, 42(1): 46−53. doi: 10.12171/j.1000-1522.20180437.
[21] 郑元, 赵忠, 周慧, 等. 刺槐树冠光合作用的空间异质性[J]. 生态学报, 2010, 30(23):6399−6408. Zheng Y, Zhao Z, Zhou H, et al. Spatial heterogeneity of canopy photosynthesis in black locust plantations[J]. Acta Ecologica Sinica, 2010, 30(23): 6399−6408.
[22] 姜丽芬, 石福臣, 王化田, 等. 叶绿素计SPAD-502在林业上应用[J]. 生态学杂志, 2005, 24(12):1543−1548. doi: 10.3321/j.issn:1000-4890.2005.12.034. Jiang L F, Shi F C, Wang H T, et al. Application tryout of chlorophyll meter SPAD-502 in forestry[J]. Chinese Journal of Ecology, 2005, 24(12): 1543−1548. doi: 10.3321/j.issn:1000-4890.2005.12.034.
[23] 张亦弛, 郭素娟, 孙传昊. 生长延缓剂对板栗枝条的促壮效应和叶片发育及生理的影响[J]. 西北农林科技大学学报(自然科学版), 2019, 47(9):79−89. Zhang Y C, Guo S J, Sun C H. Effects of growth retardants on branches growth, leaves development andphysiology of chestnut[J]. Journal of Northwest A&F University (Natural Science Edition), 2019, 47(9): 79−89.
[24] 宋小园, 朱仲元, 刘艳伟, 等. 通径分析在SPSS逐步线性回归中的实现[J]. 干旱区研究, 2016, 33(1):108−113. Song X Y, Zhu Z Y, Liu Y W, et al. Application of path analysis in stepwise linear regression SPSS[J]. Arid Zone Research, 2016, 33(1): 108−113.
[25] 李六林, 季兰. 杂种榛子不同方位叶片光合作用的日变化[J]. 林业科学, 2006, 42(12):50−56. Li L L, Ji L. Diurnal variation in photosynthesis of differently directional leaves in hybrid hazels (Corylus heterophylla × Corylus avellana)[J]. Scientia Silvae Sinicae, 2006, 42(12): 50−56.
[26] 杨晓玲, 杨晴, 郭守华, 等. 燕龙板栗光合作用及其相关因素的日变化[J]. 经济林研究, 2008, 26(1):67−70. doi: 10.3969/j.issn.1003-8981.2008.01.014. Yang X L, Yang Q, Guo S H, et al. Diurnal variations of photosynthesis in Yanlong chestnut and some related factors[J]. Nonwood Forest Research, 2008, 26(1): 67−70. doi: 10.3969/j.issn.1003-8981.2008.01.014.
[27] 刘庆忠, 董合敏, 刘鹏, 等. 板栗的光合特性研究[J]. 果树学报, 2005, 22(4):335−338. Liu Q Z, Dong H M, Liu P, et al. Studies on the photosynthetic characteristics of chestnut trees[J]. Journal of Fruit Science, 2005, 22(4): 335−338.
[28] 曹阳, 文国宇, 李茂军, 等. 种植密度对烤烟光合特性日变化及其主要化学成分的影响[J]. 南京农业大学学报, 2019, 42(4):641−647. doi: 10.7685/jnau.201811015. Cao Y, Wen G Y, Li M J, et al. Effect of plant density on diurnal changes of photosynthetic characteristics and its main chemical components of flue-cured tobacco (Nicotiana tabacum L.)[J]. Journal of Nanjing Agricultural University, 2019, 42(4): 641−647. doi: 10.7685/jnau.201811015.
[29] 唐雨薇, 魏新宇, 李晨, 等. 三峡库区柑橘树冠光合特征的空间异质性[J]. 湖北农业科学, 2016, 55(19):5025−5029. Tang Y W, Wei X Y, Li C, et al. Spatial heterogeneity of canopy photosynthesis of citrus plantation in Three Gorges Reservior Region[J]. Hubei Agricultural Sciences, 2016, 55(19): 5025−5029.
[30] 黄良帅, 韩海荣, 牛树奎, 等. 华北落叶松冠层光合生理特性的空间异质性[J]. 南京林业大学学报(自然科学版), 2017, 41(2):193−197. Huang L S, Han H R, Niu S K, et al. Spatial heterogeneity of canopy photosynthesis in Larix principis-rupprechtii Mayr. plantations[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2017, 41(2): 193−197.
[31] 臧润国, 刘华, 张新平, 等. 天山中部天然林分中不同龄级天山云杉光合特性[J]. 林业科学, 2009, 45(5):60−68. doi: 10.3321/j.issn:1001-7488.2009.05.009. Zang R G, Liu H, Zhang X P, et al. Photosynthetic characteristics of Picea schrenkiana var. tianschanica of different age classes in a natural stand in the central part of Tianshan Mountains[J]. Scientia Silvae Sinicae, 2009, 45(5): 60−68. doi: 10.3321/j.issn:1001-7488.2009.05.009.
[32] Gao Y B, Zheng W W, Zhang C, et al. High temperature and high light intensity induced photoinhibition of bayberry (Myrica rubra Sieb
. et Zucc.) by disruption of D1 turnover in photosystem Ⅱ[J]. Scientia Horticulturae, 2019, 248: 132−137. doi: 10.1016/j.scienta.2019.01.007. [33] 高清华, 叶正文, 章镇, 等. 设施栽培中早熟甜油桃的光合特性研究[J]. 农业现代化研究, 2006, 27(4):307−310. doi: 10.3969/j.issn.1000-0275.2006.04.018. Gao Q H, Ye Z W, Zhang Z, et al. Photosynthetic characteristic in precocious and sweet nectarine trees in greenhouse[J]. Research of Agricultural Modernization, 2006, 27(4): 307−310. doi: 10.3969/j.issn.1000-0275.2006.04.018.
[34] 穆宏平, 叶万辉, 陈贻竹, 等. 不同磷营养水平对朱砂根和山血丹光合作用及生长的影响[J]. 武汉植物学研究, 2008, 26(5):514−519. Mu H P, Ye W H, Chen Y Z, et al. Photosynthesis and growth of and under different phosphorus nutrition levels[J]. Journal of Wuhan Botanical Research, 2008, 26(5): 514−519.
[35] 唐凤德, 武耀祥, 韩士杰, 等. 长白山阔叶红松林叶片气孔导度与环境因子的关系[J]. 生态学报, 2008, 28(11):5649−5655. doi: 10.3321/j.issn:1000-0933.2008.11.050. Tang F D, Wu Y X, Han S J, et al. Relationship of stamatal conductance of leaf with environmental factors in broadlcaved Korean pine forest at Changbai Mountain[J]. Acta Ecologica Sinica, 2008, 28(11): 5649−5655. doi: 10.3321/j.issn:1000-0933.2008.11.050.
[36] 胡昌浩, 董树亭, 岳寿松, 等. 高产夏玉米群体光合速率与产量关系的研究[J]. 作物学报, 1993, 19(1):63−69. doi: 10.3321/j.issn:0496-3490.1993.01.009 Hu C H, Dong S T, Yue S S, et al. Studies on the relationship between canopy apparent photosynthesis rate and grain yield in high yielding summer corn (Zea may L.)[J]. Acta Agronomica Sinica, 1993, 19(1): 63−69. doi: 10.3321/j.issn:0496-3490.1993.01.009
[37] 郭素娟, 熊欢, 李广会, 等. 树体结构对板栗冠层光辐射与光合特征及产量的影响[J]. 东北林业大学学报, 2014, 42(1):14−18. doi: 10.3969/j.issn.1000-5382.2014.01.004. Guo S J, Xiong H, Li G H, et al. Effect of chestnut (Castanea mollissima Bl.) tree structure on canopy light radiation, photosynthesis and yield[J]. Journal of Northeast Forestry University, 2014, 42(1): 14−18. doi: 10.3969/j.issn.1000-5382.2014.01.004.
[38] 曹永慧, 周本智, 王小明, 等. 冠层高度对毛竹叶片光合生理特性的影响[J]. 西北植物学报, 2016, 36(11):2256−2266. doi: 10.7606/j.issn.1000-4025.2016.11.2256. Cao Y H, Zhou B Z, Wang X M, et al. Effects of canopy height on photosynthetic physiology characteristics of Phyllostachys pubescens leaves[J]. Acta Botanica Boreali-Occidentalia Sinica, 2016, 36(11): 2256−2266. doi: 10.7606/j.issn.1000-4025.2016.11.2256.
[39] 霍宏, 王传宽. 冠层部位和叶龄对红松光合蒸腾特性的影响[J]. 应用生态学报, 2007, 18(6):1181−1186. doi: 10.3321/j.issn:1001-9332.2007.06.002. Huo H, Wang C K. Effects of canopy position and leaf age on photosynthesis and transpiration of Pinus koraiensis[J]. Chinese Journal of Applied Ecology, 2007, 18(6): 1181−1186. doi: 10.3321/j.issn:1001-9332.2007.06.002.
[40] 商天其, 孙志鸿. 香樟幼龄林不同叶龄叶片的光合特征和单萜释放规律[J]. 应用与环境生物学报, 2019, 25(1):89−99. Shang T Q, Sun Z H. Photosynthetic characteristics and monoterpenes emission are related to foliage ontogeny in Cinnamomum camphora[J]. Chin J Appl Environ Biol, 2019, 25(1): 89−99.
[41] 张小全, 徐德应, 赵茂盛. 林冠结构、辐射传输与冠层光合作用研究综述[J]. 林业科学研究, 1999, 12(4):411−421. doi: 10.3321/j.issn:1001-1498.1999.04.014. Zhang X Q, Xu D Y, Zhao M S. Review of canopy structure, radiation transmission and canopy photosynthesis[J]. Forest Research, 1999, 12(4): 411−421. doi: 10.3321/j.issn:1001-1498.1999.04.014.
[42] Wei X, Otieno D, Ko J, et al. Conditional variations in temperature response of photosynthesis, mesophyll and stomatal control of water use in rice and winter wheat[J]. Field Crops Research, 2016, 199: 77−88. doi: 10.1016/j.fcr.2016.09.016.
[43] 李勇, 韩海荣, 康峰峰, 等. 油松人工林冠层光合生理特性的空间异质性[J]. 东北林业大学学报, 2013, 41(4):32−35. doi: 10.3969/j.issn.1000-5382.2013.04.008. Li Y, Han H R, Kang F F, et al. Spatial heterogeneity of photosynthetic characterisitics of Pinus tabulaeformis canopy[J]. Journal of Northeast Forestry University, 2013, 41(4): 32−35. doi: 10.3969/j.issn.1000-5382.2013.04.008.
-
期刊类型引用(3)
1. 刘索名,王君杰,燕云飞,姜立春. 小兴安岭兴安落叶松人工林冠幅模型构建. 北京林业大学学报. 2023(05): 79-87 .  本站查看
本站查看
2. 李应涛,刘时良,孙海龙,王卫霞,向玮. 云冷杉针阔混交林单木枝下高和冠幅模型构建. 森林与环境学报. 2022(03): 289-296 . 百度学术
3. 佟艺玟,陈东升,冯健,高慧淋. 基于线性分位数混合效应的辽东山区红松冠幅模型. 应用生态学报. 2022(09): 2321-2330 . 百度学术
其他类型引用(1)














计量
- 文章访问数: 1276
- HTML全文浏览量: 408
- PDF下载量: 85
- 被引次数: 4
