
Spatiotemporal distribution and seed germination characteristics of Betula costata seed rain in the spruce-fir mixed forest
-
摘要:目的 研究东北云冷杉针阔混交林枫桦种子雨时空分布规律和种子萌发状况,为云冷杉针阔混交林物种多样性保护提供理论依据。方法 基于在吉林省汪清林业局设立的9块1 hm2固定样地,按对角线五点取样法在每块样地布设5个(9块样地共布设45个)种子收集器收集枫桦种子,采用方差/均值(Cx
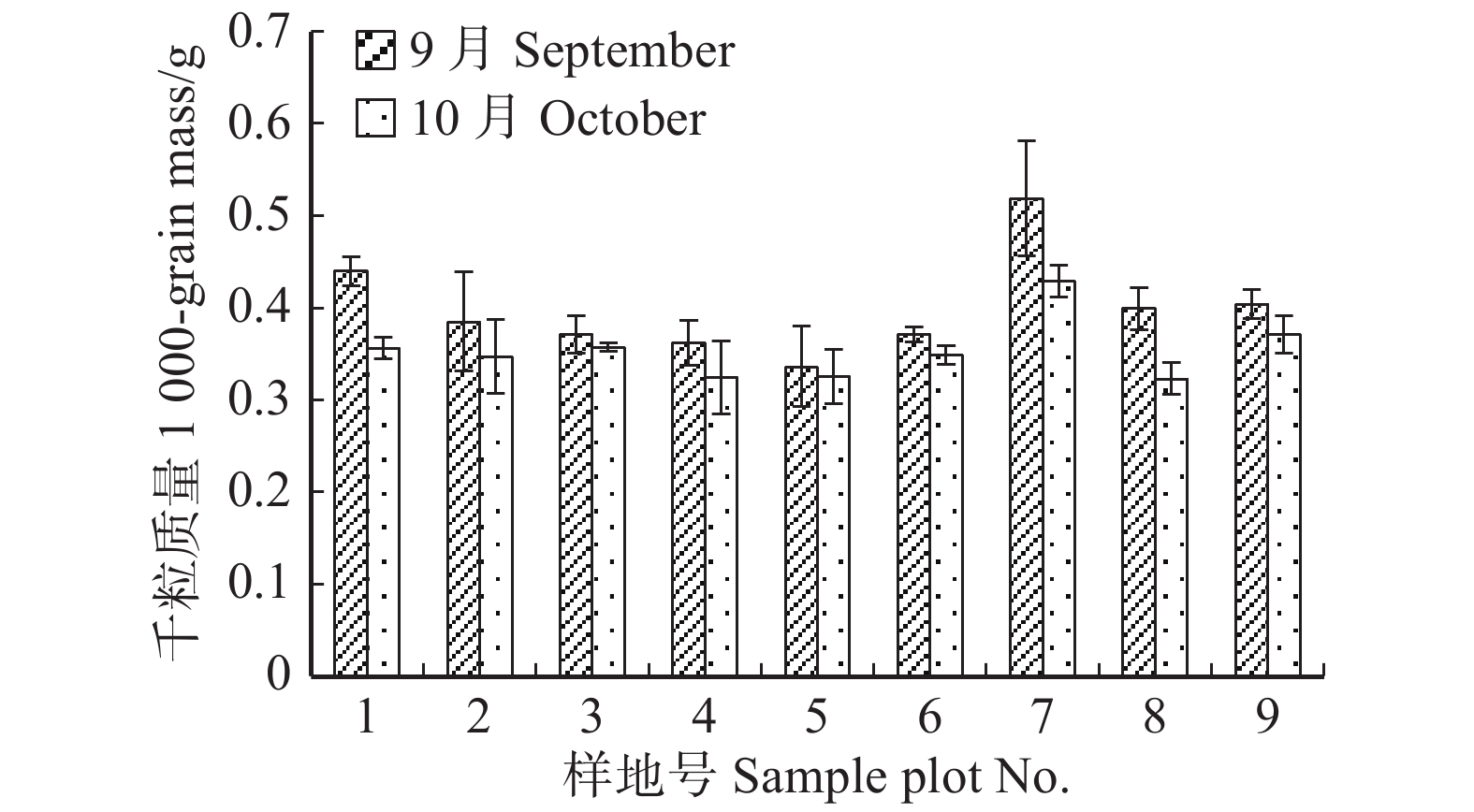
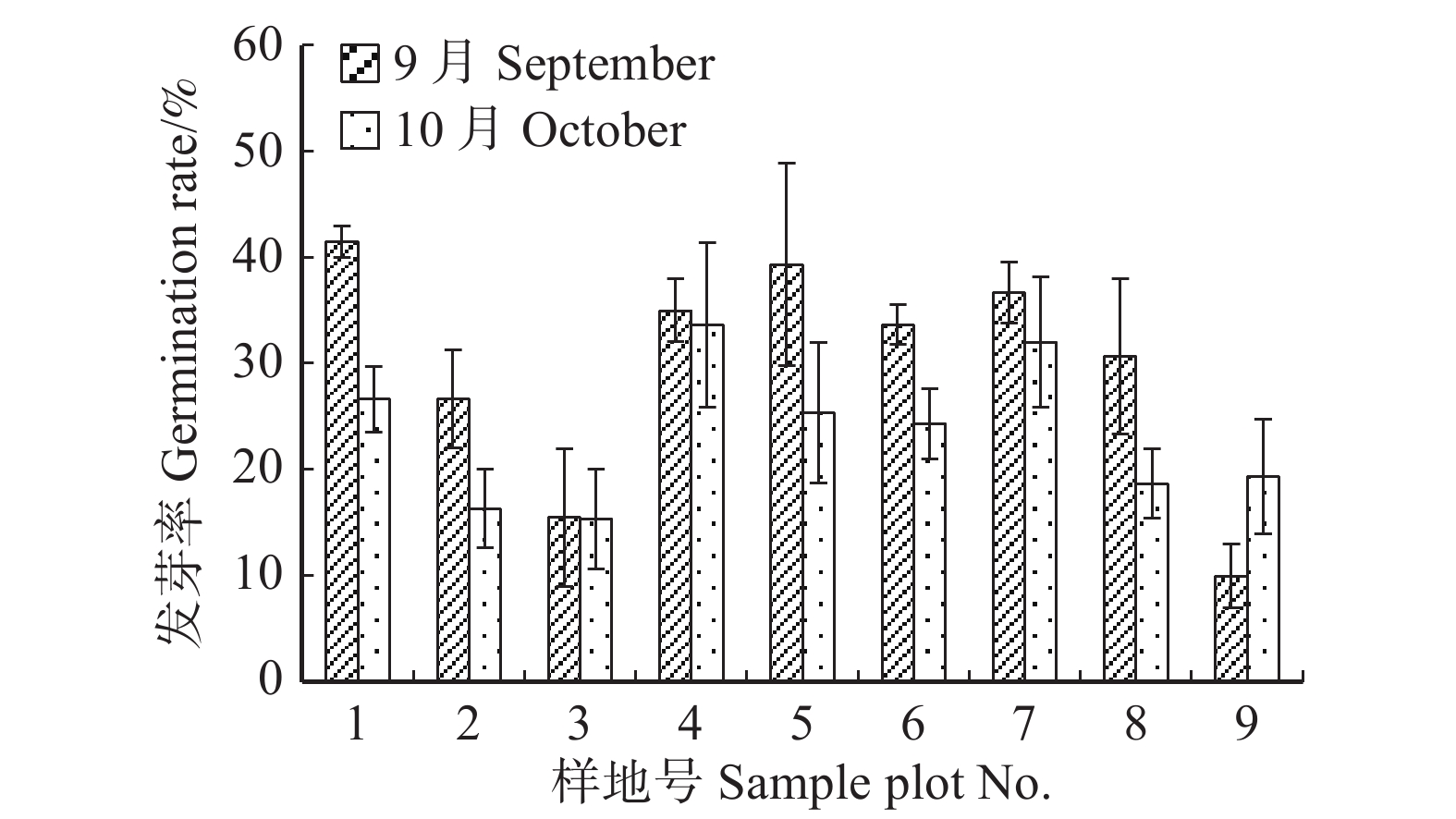
)、负二项指数(K)、聚集度指标(David和Moore丛生指标,I)、Cassie. R. M.指标(CA)等4种空间分布类型评价方法判别种子雨空间分布状态,此外,从9、10月份收集的种子中,每块样地随机抽取300粒枫桦种子进行种子萌发试验,分别测定种子千粒质量、发芽势和发芽率。 结果 从7—11月份,共收集到69 100粒种子;枫桦落种高峰期为10月份;种子分布方差/均值(Cx)达1116.30、负二项指数(K)达4.75、聚集度指标(David和Moore丛生指标,I)达1115.30、Cassie. R. M.指标(CA)达0.41,4个指标均显示种子空间分布呈聚集分布;9月份与10月份收集的平均种子千粒质量分别为(0.398 ± 0.064)g和(0.353 ± 0.040)g,9月份种子千粒质量显著高于10月份(P < 0.01);9月份收集的种子平均发芽率为(29.84 ± 11.68)%,显著高于10月份(23.52 ± 8.23)%(P < 0.05)。 结论 在云冷杉针阔混交林中,枫桦种子小结实量大,且具有明显的时间分布特征,由于在一定程度上种子扩散受到限制,使得枫桦种子呈聚集分布状态。该区域枫桦种源丰富但种子萌发能力差,这些特性将会影响枫桦分布格局,从而影响该地区云冷杉针阔混交林群落结构和物种多样性。Abstract:Objective The population of Betula costata in spruce-fir mixed forest in northeastern China was taken as the research object. The spatiotemporal distribution of Betula costata seed rain and seed germination characteristics were studied to provide theoretical basis for species diversity protection of the forest.Method Based on the 9 sample plots, 1 ha (100 m × 100 m) each set in Wangqing Forestry Bureau, Jilin Province of northeastern China, sampling by diagonal five points, we set 5 seed traps in each sample plot, totally 45 in 9 sample plots to collect Betula costata seeds. We used 4 evaluation methods on spatial distribution types to distinguish the spatial distribution state of seed rain, including variance-mean ratio (Cx), negative binomial parameter (K), aggregation index (I), Cassie. R.M. index (CA). In addition, 300 Betula costata seeds collected in September and October were randomly selected from each sample plot for seed germination experiment, and the 1 000-grain mass, germination potential and germination percentage were measured, respectively.Result From July to November, 69 100 seeds were collected in total. The peak period of seed falling for Betula costata was October. We found that the seeds of Betula costata presented aggregation distribution at the spatial level based on 4 indexes, i.e. Cx reaching 1 116.30, K reaching 4.75, I reaching 1 115.30, CA reaching 0.41, respectively. The average 1000-grain mass value of Betula costata seeds in September was 0.398 g, which was significantly higher than that in October (0.353 g) (P<0.01). Similarly, the average germination rate in September was (29.84 ± 11.68)%, which was significantly higher than that in October (23.52 ± 8.23)% (P < 0.05).Conclusion In spruce-fir coniferous-broadleaved mixed forest, the seeds are small and fruiting for Betula costata and have obvious time distribution characteristics. Because the diffusion of Betula costata seeds is limited, as a result the seeds show an aggregated distribution pattern. In this region, the characteristics of rich provenance and poor germination ability of Betula costata will affect the distribution pattern of Betula costata, thus affect the community structure and species diversity of spruce-fir coniferous-broadleaved mixed forest in this area. -
蒿柳(Salix viminalis)为柳属蒿柳组灌木,主要分布在北半球高纬度地区,是重要的能源、蜜源树种及沼泽地带的先锋树种[1]。目前,关于蒿柳的研究主要集中在群体遗传多样性、污染治理和能源林建设等方面[2-5],且研究材料多以自然群体或无性繁殖的扦插材料为主。为解决能源短缺问题,瑞典等国家很早就开展了以蒿柳为主要树种的能源柳育种工作,虽然获得了一些优良的能源柳品种,但由于大部分蒿柳品系都是瑞典起源的,杂交后代已出现衰退现象[1-6]。我国地域广阔,具有丰富野生蒿柳资源,但由于蒿柳为雌雄异株植物,雌、雄花开放时间不同步,且不同地理居群的蒿柳间的杂交亲和性有差异[2]。目前,我国蒿柳资源多处于野生状态,利用天然优株进行杂交育种的工作亟待开展。花粉是植物的雄配子体,在有性生殖中具有重要作用[7],研究花粉萌发特性是了解植物有性生殖,进行杂交育种的前提。因此,进行蒿柳花粉离体萌发特性研究,对深入开展能源柳育种工作具有重要意义。
培养条件(温度、光照、pH值等)和培养基组分(蔗糖、H3BO3、激素、Ca2+ 等)对花粉离体萌发均有影响,但不同植物花粉所需的最佳培养条件及浓度范围存在一定差异。如黄连木(Pistacia chinensis)花粉生长的最适温度为25 ℃,最适蔗糖浓度为15%[8];枣(Ziziphus jujuba)花粉在15.0 mg/L H3BO4+5.0 mg/L IAA+10.0 mg/L GA3的培养基上萌发率达到了90.23%[9];小黑杨(Populus simonii×P. nigra)花粉萌发最适培养基为150 g/L蔗糖+20 mg/LH 3BO3+40 mg/LCaCl2[10];梨(Pyrus pyrifolia)花粉在30 mmol/L MES缓冲液中添加0. 01%硼酸,0.03% CaCl 2·2H2O,15% PEG-4000,10%蔗糖,pH值在6.5左右的条件下的萌发率为59.2%,花粉管长度为966.3 μm[11];核桃(Juglans regia)花粉萌发和花粉管生长的最适培养基为20%蔗糖+0.02%~0.03%的硼酸+0.05%Ca(NO3)2,最佳培养条件为25 ℃,24 h后花粉萌发率达74.46%[12];月季(Rosa chinensis)花粉萌发的适宜条件为20%蔗糖+50~100 mg/L硼酸+100~200 mg/L Ca(NO3)2[13]。适宜的萌发条件可以提高花粉的萌发率和花粉管的伸长速度,显著增加授粉效率和座果率,加快育种进程[14],Stanley等[15]认为离体萌发与在柱头萌发时的花粉活力是否接近,是判断花粉离体萌发条件及培养基成分最为可靠的方法。本实验研究蒿柳花粉离体萌发的培养条件及培养基组分对花粉萌发率和花粉管生长的影响,并与花粉在柱头萌发情况相对比,获得蒿柳花粉萌发的最佳培养条件和培养基组分,可为蒿柳的科学授粉和杂交育种奠定基础。
1. 材料与方法
1.1 材料
中国林业科学研究院玉泉山苗圃栽种的蒿柳,在雄花序处于半开或全开状态时采集花粉带回实验室立即培养。
1.2 方法
1.2.1 温度和光照对蒿柳花粉萌发的影响
采用单因子实验设计,采集成熟蒿柳花粉,用仅含有2%琼脂粉的固体培养基培养花粉,温度处理设5、10、15、20、25、30和35 ℃共7个梯度,光照强度均为1 500 lx;光照强度处理设0、300、600、900、1 200、1 500和2 000 lx共7个梯度,培养温度均为20 ℃。
1.2.2 培养基成分对蒿柳花粉萌发的影响
采用正交实验设计,2%琼脂粉的固体培养基中添加不同质量浓度的蔗糖、H3BO3并调整培养基的pH值,光照强度1 000 lx,培养温度25 ℃,设计3因素3水平L9(34)正交试验,共9个组合。
1.2.3 花粉萌发率统计及花粉管长度测量
所有处理的花粉培养时间均为4 h,然后在ZEISS Axio lmager A1光学显微镜下统计花粉萌发率,测量花粉管长度。萌发率=(萌发的花粉数/总的花粉数)×100%,花粉管长度用软件自带标尺测量。每处理重复3次,每次观察1个载玻片,每个载玻片观察3个视野,每个视野花粉粒不少于30粒。
1.2.4 花粉在柱头上萌发
选取处于全开状态的雌花序进行人工授粉,于授粉后1、2、3和4 h取整个雌花序在FAA固定液中固定24 h,50 mmol/L的偏磷酸钾溶液冲洗3次,然后用4 mol/L的NaOH透明和软化24 h,蒸馏水多次换水浸泡除净NaOH,转移到0.1%水溶性苯胺蓝染液中染色4 h。用镊子从花序上取小花(子房),放在载玻片上,用滤纸吸出染液,滴一滴甘油,盖上盖玻片,轻轻敲压盖玻片,使花柱展开,最后在荧光显微镜观察,拍照。
1.3 数据统计及处理
采用SPSS19.0对数据进行处理并进行LSD显著性分析。
2. 结果与分析
2.1 温度对蒿柳花粉萌发的影响
表 1显示,在5~35 ℃范围内,随着培养温度的升高,花粉萌发率先上升后下降。培养温度为5和35 ℃时,花粉萌发率均低于10%,与其他培养温度下的差异显著(P<0.05);培养温度为20和25 ℃时的蒿柳花粉萌发率分别为72%和64.9%,与其他培养温度下的萌发率相比差异显著(P<0.05)。蒿柳花粉管长度的变化与萌发率变化一致,随着培养温度的升高,花粉管长度先上升后下降。培养温度为20和25 ℃时,花粉管长度分别为306.32和301.18 μm,伸长速度均大于75 μm/h,与其他温度下的长度和伸长速度差异显著(P<0.05)。结果表明,温度影响蒿柳花粉萌发及花粉管伸长,15~25 ℃时,蒿柳花粉萌发率较高,而20~25 ℃为蒿柳花粉萌发的最适温度区间,培养温度过高或过低,均不利于蒿柳花粉萌发。
表 1 温度对蒿柳花粉萌发的影响Table 1. Effects of different temperatures on the pollen germination of S. viminalis温度
Temperature/℃萌发率
Germination rate/%花粉管长度
Pollen tube length/μm5 4.8±2.25d 19.76±1.94d 10 20.2±8.11c 44.92±4.80c 15 42.1±5.85b 160.64±13.28b 20 72.0±4.90a 306.32±12.82a 25 64.9±1.64a 301.18±25.62a 30 22.9±2.05c 53.68±7.36c 35 8.3±1.65d 29.46±7.16d 2.2 光照强度对蒿柳花粉萌发的影响
表 2显示,在黑暗和300 lx光照强度时,蒿柳花粉萌发率分别为15.16%和11.27%,显著低于其他光照强度下的萌发率;光照强度为1 500 lx时,蒿柳花粉萌发率最高为64.14%,与2 000 lx的萌发率差异不显著,与其他光照强度下的萌发率差异显著(P<0.05)。光照强度低于1 200 lx时,蒿柳花粉管平均长度为182.07~227.31 μm,各处理间无显著差异;光照强度为1 500和2 000 lx时的花粉管长度分别为325.95和320.77 μm,伸长速度均大于75 μm/h,显著大于其他处理的长度和伸长速度(P<0.05)。结果表明,光照强度影响蒿柳花粉萌发率和花粉管伸长速度,低于1 200 lx不利于花粉萌发,1 500和2 000 lx是蒿柳花粉萌发的最佳光照强度。
表 2 光照强度对蒿柳花粉萌发的影响Table 2. Effects of light intensity on the pollen germination of S. viminalis光照强度
Light intensity/lx萌发率
Germination rate/%花粉管长度
Pollen tube length/μm0 15.16±2.01e 198.70±15.66b 300 11.27±3.91e 182.41±12.83b 600 36.63±4.35d 182.07±16.90b 900 43.07±2.50c 199.45±16.39b 1 200 52.00±6.14b 227.31±17.71b 1 500 64.14±0.84a 325.95±15.30a 2 000 59.85±2.26a 320.77±15.58a 2.3 培养基成分对蒿柳花粉萌发的影响
将蔗糖、硼酸及pH值3个因素进行L 9(34)正交实验(表 3)。对表 3中数据进行方差分析。结果表明,3个因子均对蒿柳花粉萌发具有显著的影响:蔗糖F=3.268,P<0.05,H3BO3的F=37.368,P<0.05,pH值的F=91.427,P<0.05。极差分析显示,3因素对蒿柳花粉萌发的影响为:pH>H3BO3>蔗糖。正交实验平方和比较分析的结果表明:蔗糖各种质量浓度下的花粉萌发率以100 g/L最高,H3BO3以200 mg/L最高,pH值以5.0最高,由此推论3个因子最佳组合为蔗糖100 g/L、H3BO3 200 mg/L和pH 5.0。
表 3 正交实验组合对蒿柳花粉的萌发率及花粉管长度的影响Table 3. Pollen germination rate and tube length of S. viminalis in orthogonal design experiment处理编号
Treatment No.蔗糖
Sucrose/(g·L-1)H3BO3/(mg·L-1) pH 萌发率
Germination rate/%花粉管长度
Pollen tube length/μm1 50 100 5.0 40.70±5.86b 257.25±15.27b 2 50 200 7.0 17.64±3.77d 57.75±12.36e 3 50 500 9.0 3.63±0.72e 41.00±9.43f 4 100 100 7.0 22.18±0.69cd 102.44±8.64d 5 100 200 9.0 29.29±4.02c 142.32±17.10c 6 100 500 5.0 26.66±3.14c 146.20±9.93c 7 150 100 9.0 3.33±2.18e 28.66±4.90f 8 150 200 5.0 48.80±4.20a 288.50±13.50a 9 150 500 7.0 7.39±0.52de 59.85±7.18e 2.4 蒿柳花粉离体萌发与柱头萌发比较
利用单因素和正交实验筛选的最佳培养组合,培养蒿柳花粉,分别在培养1、2、3和4 h时取样,测量花粉管长度。由图 1、2结果所示,蒿柳花粉在适宜条件下可迅速萌发,柱头授粉1 h时,已有大量花粉黏附在柱头上(图 2A),表明花粉已经开始萌发,花粉管平均长度为48.07 μm;而离体培养1 h时,花粉吸水膨胀,部分花粉已伸出花粉管,花粉管平均长度45.83 μm(图 2E),与柱头萌发相比无显著差异。柱头授粉2、3和4 h时,花粉管不断生长,长度分别为85.42、224.46和320.19 μm,而同时期离体培养的花粉管长度分别达到了90.34、202.65和314.88 μm,相同时间的离体与柱头萌发差异不显著。结果表明,本实验筛选的蒿柳花粉萌发培养条件接近于在柱头萌发的活体状态,适合蒿柳花粉的萌发。
![]() 图 1 蒿柳花粉离体萌发与柱头萌发的花粉管长度比较Figure 1. Pollen germination length of S. viminalis both in vitro and in vivo
图 1 蒿柳花粉离体萌发与柱头萌发的花粉管长度比较Figure 1. Pollen germination length of S. viminalis both in vitro and in vivo![]() 图 2 蒿柳花粉离体萌发与柱头萌发过程A~D分别为花粉在柱头萌发1、2、3、4 h的状态和过程,40×;E~H分别为花粉离体萌发1、2、3、4 h的状态和过程,E、H为100×,F、G为200×。Figure 2. Pollen germination process and stigma germination of S. viminalis both in vitro and in vivoA-D, Pollen germination state and process in vivo at 1, 2, 3, 4 hours respectively, 40×; E-H, Pollen germination state and process in vitro at 1, 2, 3, 4 hours respectively, E and H, 100×, F and G 200×.
图 2 蒿柳花粉离体萌发与柱头萌发过程A~D分别为花粉在柱头萌发1、2、3、4 h的状态和过程,40×;E~H分别为花粉离体萌发1、2、3、4 h的状态和过程,E、H为100×,F、G为200×。Figure 2. Pollen germination process and stigma germination of S. viminalis both in vitro and in vivoA-D, Pollen germination state and process in vivo at 1, 2, 3, 4 hours respectively, 40×; E-H, Pollen germination state and process in vitro at 1, 2, 3, 4 hours respectively, E and H, 100×, F and G 200×.3. 讨论与结论
温度是影响花粉萌发率的重要因素,许多研究指出[16-18],25 ℃是大多植物花粉萌发的最适温度,但分布于不同地区的植物,其花粉最适萌发温度差异明显。油橄榄(Olea europaea)花粉萌发的适宜温度范围为25~30 ℃,低于10 ℃不萌发,而高于35 ℃萌发率下降[19];董易等[20]报道,荷兰水仙(Narcissus pseudonarcissus)花粉的最适萌发温度为20 ℃;而与蒿柳同属杨柳科(Salicaceae)的小黑杨花粉萌发的最适温度为21 ℃[10];由于不同地域的菊花(Chrysanthemum morifolium)长期适应各自区域的气候特点,导致花粉对温度的响应不同。低温或高温下花粉的萌发率均下降,低温下花粉代谢过程中酶活力下降,膜流动性下降,代谢速率降低,抑制了花粉的萌发[21];而高温下,花粉失水速度加快,含水量降低,花粉代谢过程中一些对温度敏感的酶活性受高温钝化失活,膜完整性被破坏,代谢速率下降,抑制花粉萌发[22]。蒿柳多分布在北半球高纬度地区,花期在4—5月,此时该区域的最高温度一般在15~25 ℃,本实验也表明,温度区间为15~25 ℃时蒿柳花粉萌发率较高,这是蒿柳长期适应分布区的气候特点的结果。
关于光照与花粉萌发的关系至今尚无一致结论。杨中汉等[23]认为,照光不仅能促进兰州百合(Lilium davidii)花粉的萌发也促进花粉管的伸长。郭思佳等[24]也发现,各居群有斑百合(L. concolor var. ulchellum)在太阳光下散粉萌发率最高,在日光灯下散粉萌发率降低近一半,在室内散射光散粉花粉的萌发率最低。与此相反,徐进等[25]报道,在黑暗条件下,马尾松(Pinus massoniana)花粉发芽率较其他光照条件下高,但它们的差异未达到显著水平。许珂等[26]也发现,在弱光(0.562 5 μmol/(s·m2))与黑暗下培养,金银忍冬(Lonicera maackii)花粉的萌发率无明显变化,但黑暗条件更利于花粉管的生长。光照能为植物提供更多的能量,提高植株温度,使一些酶的活性(比如G蛋白)升高,调节细胞外钙调素启动花粉萌发和花粉管伸长,促进花粉管伸长[27]。本研究发现,蒿柳花粉萌发率及花粉管长度均受到光照强度的影响,且在一定范围内,光照强度越大,花粉萌发率越高,花粉管伸长速度越快,低于1 200 lx不利于花粉萌发,1 500~2 000 lx是蒿柳花粉萌发的最佳光照强度,本实验未就高于2 000 lx的光照强度对蒿柳花粉萌发情况进行研究。
花粉离体培养一般在培养基上进行,培养基组分如蔗糖、硼酸及pH值等均影响花粉萌发。蔗糖作为植物生长的重要的能源物质和渗透调节物质,在花粉萌发和生长过程中具有重要作用,15%的蔗糖利于黄连木花粉的萌发和生长[8];10%的蔗糖或葡萄糖适宜梨花粉萌发,蔗糖浓度过低或过高对花粉的萌发和花粉管生长不利[11]。蔗糖浓度过低时,花粉细胞破裂,细胞内容物散出,浓度过高会造成花粉细胞的质壁分离,抑制花粉萌发生长。本实验中蔗糖浓度为10%时,蒿柳花粉萌发率显著高于其他浓度,与上述研究结果一致。
培养基中添加一定浓度硼酸有利于花粉萌发和花粉管的生长。尚宏芹[28]研究发现,20 mg/L的硼酸对两种芍药花粉萌发具有显著促进作用。李旭新等[8]报道,硼酸处理浓度为100 mg/L 时,黄连木花粉萌发率和花粉管生长均达最大值,硼酸浓度继续升高,花粉萌发率和花粉管生长反而显著下降。韩志强等[29]对不同营养元素及其配比对枣花粉萌发与花粉管生长的影响进行研究后得出,20 mg/L的硼酸是促进花粉管生长最适宜质量浓度,本实验在硼酸对蒿柳花粉萌发的研究中也得出了相似的结论。
培养基pH值影响花粉萌发,适合的花粉离体培养基多为弱酸性,果梅花粉离体萌发培养基的适宜pH值为6.5[30];芍药(Peaonia lactiflora)花粉生长的最适pH值为6~7[28];培养基pH值为6.5时,促进梨花粉萌发及花粉管生长[11]。本实验结果表明,在pH值为5.0时,蒿柳花粉的萌发率和花粉管长度最高,虽略低于上述研究结果,但在实验筛选的培养条件下,蒿柳花粉离体萌发与柱头萌发无显著差异。提高培养基的pH值是否会进一步促进蒿柳花粉萌发率和花粉管伸长,由于本实验设置的pH值间隔偏大,无法更加精确地判断影响蒿柳花粉萌发的最适pH值,这还需要进一步细化研究。
综上所述,蒿柳花粉离体萌发受到光照、温度等环境因素及培养基的影响。利用筛选出的蒿柳花粉最佳离体培养组合,即利用pH值为5.0的含有蔗糖100 g/L和H3BO3 200 mg/L的培养基,在培养温度为20 ℃,光照强度为1 500 lx的培养条件下培养花粉,并与花粉在柱头上活体萌发相比较,二者花粉管的伸长无显著差异。
-
![]()
图 2 云冷杉针阔混交林各样地种子千粒质量
Figure 2. 1 000-grain mass of spruce-firconiferous-broadleaved forest in each sample plot
![]()
图 3 云冷杉针阔混交林各样地种子发芽率
Figure 3. Seed germination rate of spruce-fir coniferous-broadleaved forest in each sample plot
表 1 云冷杉针阔混交林样地基本概况
Table 1 Basic condition of mixed spruce-fir coniferous-broadleaved forest
样地
Sample plot地理坐标
Geographical coordinate海拔
Altitude/m坡向
Slope
aspect坡度
Slope
degree/(°)平均胸径
Mean
DBH/cm平均树高
Mean tree
height/m林分密度/
(株·hm−2)
Stand density/
(tree·ha−1)枫桦密度
(DBH > 15 cm)/(株·hm−2)
Density of Betula costata
(DBH > 15 cm)/(tree·ha−1)YLK-1 130°15′09″E 43°22′46″N 760 东北 Northeast 15 19.0 15.1 1 232 72 YLK-2 130°15′16″E 43°22′50″N 773 东北 Northeast 16 18.2 15.1 1 165 56 YLK-3 130°15′14″E 43°22′46″N 780 东北 Northeast 5 17.4 15.8 1 328 83 YLK-4 130°15′13″E 43°22′43″N 771 东北 Northeast 5 18.5 15.5 1 539 123 YLK-5 130°15′00″E 43°23′05″N 732 东北 Northeast 5 18.6 14.9 1 309 27 YLK-6 130°15′05″E 43°23′04″N 749 东北 Northeast 5 19.9 16.5 1 436 60 YLK-7 130°15′09″E 43°23′04″N 759 东北 Northeast 5 18.8 15.7 1 195 73 YLK-8 130°15′15″E 43°23′04″N 769 东北 Northeast 5 18.4 16.3 1 301 56 YLK-9 130°15′20″E 43°23′05″N 773 东北 Northeast 3 17.6 16.5 1 437 70  下载: 导出CSV
下载: 导出CSV
表 2 各空间变异参数计算公式与分级
Table 2 Formulas and classification of spatial variation parameters
参数 Parameter 公式 Formula 分级 Classification 方差/均值 Variance/mean (Cx) Cx=S2ˉx Cx < 1 (均匀分布 Regular distribution);Cx > 1 (聚集分布 Aggregated distribution);Cx = 1 (随机分布 Random distribution)负二项指数 Negative binomial parameter (K) K=ˉx2(S2−ˉx) K < 0 (非聚集分布 Non-aggregated distribution);K > 0 (聚集分布 Aggregated distribution)聚集度指标 Aggregation index (I) I=S2ˉx−1 I < 0 (均匀分布 Regular distribution);I > 0 (聚集分布 Aggregated distribution);I = 0 (随机分布 Random distribution)Cassie. R. M.指标 Cassie. R. M. index (CA) CA=(S2−ˉx)ˉx2 CA < 0 (均匀分布 Regular distribution);CA > 0 (聚集分布 Aggregated distribution);CA = 0 (随机分布 Random distribution)注:S2为样地所有收集器收集到种子总数的方差, ˉx 为样地收集器收集到种子数量的均值。Notes: S2 is the variance of total number of seeds collected by all collectors in the sample plot,ˉx is the average number of seeds collected by all collectors in the sample plot.
下载: 导出CSV
表 3 各样地枫桦种子雨密度 粒/m2
Table 3 Density of Betula costata seed rain of each sample plot
seed/m2 样地号 Sample plot No. 7月 July 8月 August 9月 September 10月 October 11月 November 1 0.00 ± 0.00b 0.00 ± 0.00b 87.5 ± 32.07ab 222.92 ± 9.36a 0.00 ± 0.00b 2 0.00 ± 0.00b 4.69 ± 4.19b 158.75 ± 14.37a 108.98 ± 71.75ab 0.00 ± 0.00b 3 0.00 ± 0.00b 2.81 ± 1.68b 172.27 ± 7.21ab 661.88 ± 212.92a 0.00 ± 0.00b 4 0.00 ± 0.00b 15.31 ± 13.35b 118.36 ± 34.18ab 1 645.63 ± 481.73a 0.00 ± 0.00b 5 0.00 ± 0.00b 1.25 ± 1.12b 492.19 ± 139.33ab 1 405.94 ± 487.45a 0.00 ± 0.00b 6 0.00 ± 0.00b 2.19 ± 1.96b 212.5 ± 172.62ab 1 882.81 ± 455.40a 0.00 ± 0.00b 7 0.00 ± 0.00b 1.56 ± 1.40b 309.06 ± 110.07ab 4 303.75 ± 980.00a 0.00 ± 0.00b 8 0.00 ± 0.00b 53.13 ± 21.47ab 426.25 ± 96.68ab 4 313.44 ± 1 302.08ab 0.00 ± 0.00b 9 0.00 ± 0.00b 1.88 ± 1.12b 678.13 ± 199.32ab 4856.25 ± 558.52a 0.00 ± 0.00b 平均值 Mean 0.00 9.20 295.00 2155.73 0.00 注:数据为平均值±标准误。不同字母表示同一样地不同收集时间之间种子密度差异显著(P < 0.05)。Notes: data is mean ± SE. Different letters mean significant differences in same sample plot at varied collecting time (P < 0.05).
下载: 导出CSV
表 4 枫桦种子雨空间分布特征
Table 4 Spatial distribution characteristics of seed rain of Betula costata
样地号
Sample plot No.Cx 分布格局
Distribution patternK 分布格局
Distribution patternI 分布格局
Distribution patternCA 分布格局
Distribution pattern1 20.24 聚集 Aggregated 17.19 聚集 Aggregated 19.24 聚集 Aggregated 0.06 聚集 Aggregated 2 86.13 聚集 Aggregated 3.01 聚集 Aggregated 85.13 聚集 Aggregated 0.33 聚集 Aggregated 3 462.49 聚集 Aggregated 1.78 聚集 Aggregated 461.49 聚集 Aggregated 0.56 聚集 Aggregated 4 3 166.31 聚集 Aggregated 1.04 聚集 Aggregated 3 165.31 聚集 Aggregated 0.96 聚集 Aggregated 5 971.74 聚集 Aggregated 1.96 聚集 Aggregated 970.74 聚集 Aggregated 0.51 聚集 Aggregated 6 763.18 聚集 Aggregated 2.82 聚集 Aggregated 762.18 聚集 Aggregated 0.35 聚集 Aggregated 7 1 532.86 聚集 Aggregated 3.01 聚集 Aggregated 1 531.86 聚集 Aggregated 0.33 聚集 Aggregated 8 2 489.16 聚集 Aggregated 1.93 聚集 Aggregated 2 488.16 聚集 Aggregated 0.52 聚集 Aggregated 9 554.61 聚集 Aggregated 10.00 聚集 Aggregated 553.61 聚集 Aggregated 0.10 聚集 Aggregated 均值 Mean 1 116.30 4.75 1 115.30 0.41
下载: 导出CSV
-
[1] 杜彦君, 马克平. 森林种子雨研究进展与展望[J]. 生物多样性, 2012, 20(1):94−107. doi: 10.3724/SP.J.1003.2012.14161. Du Y J, Ma K P. Advancements and prospects in forest seed rain studies[J]. Biodiversity Science, 2012, 20(1): 94−107. doi: 10.3724/SP.J.1003.2012.14161.
[2] 于顺利, 郎南军, 彭明俊, 等. 种子雨研究进展[J]. 生态学杂志, 2007, 26(10):1646−1652. Yu S L, Lang N J, Peng M J, et al. Research advances in seed rain[J]. Chinese Journal of Ecology, 2007, 26(10): 1646−1652.
[3] Martinez R M, Castro A S. Seed rain and advanced regeneration in a tropical rain forest[M]//Frugivory and seed dispersal: ecological and evolutionary aspects. Dordrecht: Springer, 1993: 299−318.
[4] Wang B C, Smith T B. Closing the seed dispersal loop[J]. Trends in Ecology & Evolution, 2002, 17: 379−386.
[5] Perini M, Dias H M, Kunz S H. The role of environmental heterogeneity in the seed rain pattern[J]. Florestae Ambiente, 2019, 26(9): 1−10.
[6] Seidler T G, Plotkin J B. Seed dispersal and spatial pattern in tropical trees[J]. PLoS Biology, 2006, 4(11): 2132−2137.
[7] Traveset A, Heleno R, Nogales M. The ecology of seed dispersal[M]// Seeds: the ecology of regeneration in plant communities. New York: CAB International Publishing, 2014: 85−110.
[8] Snell R S, Beckman N G, Fricke E, et al. Consequences of intraspecific variation in seed dispersal for plant demography, communities, evolution and global change[J]. AoB Plants, 2019, 11(4): 1−19.
[9] Jara-Guerrero A, Escribano-Avila G, Espinosa C I, et al. White tailed deer as the last megafauna dispersing seeds in Neotropical dry forests: the role of fruit and seed traits[J]. Biotropica, 2018, 50(1): 169−177. doi: 10.1111/btp.12507.
[10] Urrea-Galeano L A, Andresen E, Coates R, et al. Dung beetle activity affects rain forest seed bank dynamics and seedling establishment[J]. Biotropica, 2019, 51(2): 186−195. doi: 10.1111/btp.12631.
[11] Janzen D H. Seed predation by animals[J]. Annual Review of Ecology and Systematics, 1971, 2: 465−492. doi: 10.1146/annurev.es.02.110171.002341.
[12] Solvertown J W. The evolutionary ecology of mast seeding in trees[J]. Biological Journal of the Linnean Society, 1980(14): 235−250.
[13] Silvertown J W. The evolutionary ecology of mast seeding in trees[J]. Biological Journal of the Linnean Society, 1980, 14(2): 235−250.
[14] 程积民, 赵凌平, 程杰. 子午岭60年辽东栎林种子质量与森林更新[J]. 北京林业大学学报, 2009, 31(2):10−16. doi: 10.3321/j.issn:1000-1522.2009.02.002. Cheng J M, Zhao L P, Cheng J. Seed quality and forest regeneration of a 60-year Quercus liaotungensis forest in the Ziwuling region, northwestern China[J]. Journal of Beijing Forestry University, 2009, 31(2): 10−16. doi: 10.3321/j.issn:1000-1522.2009.02.002.
[15] Houle G. Seed dispersal and seedling recruitment of Betula alleghaniensis: spatial inconsistency in time[J]. Ecology, 1998, 79(3): 807−818. doi: 10.1890/0012-9658(1998)079[0807:SDASRO]2.0.CO;2.
[16] 陈智平, 张涛, 赵万奎, 等. 黄土高原子午岭林区油松林种子雨强度及时空动态特征[J]. 中国沙漠, 2020(3):1−9. Chen Z P, Zhang T, Zhao W K, et al. Seed rain intensity and spatio-temporal dynamic characteristics of Pinus tabulifornis in Ziwu Mountain forest area of Loess Plateau of China[J]. Journal of Desert Research, 2020(3): 1−9.
[17] 钱多多, 匡旭, 王绪高, 等. 2006—2017年长白山阔叶红松林木本植物种子雨的时空动态[J]. 应用生态学报, 2019, 30(5):1487−1493. Qian D D, Kuang X, Wang X G, et al. Spatio-temporal dynamics of woody plants seed rains in broad-leaved Korean pine mixed forest in Changbai Mountains form 2006 to 2017, China[J]. The Journal of Applied Ecology, 2019, 30(5): 1487−1493.
[18] 杨彬, 王玉, 郝清玉. 木麻黄海防林种子雨的时空动态[J]. 热带亚热带植物学报, 2019, 27(4):367−375. doi: 10.11926/jtsb.4033. Yang B, Wang Y, Hao Q Y. Spatial and temporal dynamics of seed rain of Casuarina equisetifolia coastal protection forest[J]. Journal of Tropical and Subtropical Botany, 2019, 27(4): 367−375. doi: 10.11926/jtsb.4033.
[19] 肖静, 黄力, 杨超, 等. 缙云山常绿阔叶林种子雨组成及其时空动态[J]. 林业科学, 2019, 55(7):163−169. doi: 10.11707/j.1001-7488.20190718. Xiao J, Huang L, Yang C, et al. Composition and spatial-temporal variation of the seed rain in an evergreen broad-leaved forest on Jinyun Mountain[J]. Scientia Silvae Sinicae, 2019, 55(7): 163−169. doi: 10.11707/j.1001-7488.20190718.
[20] 张春雨, 赵秀海, 夏富才. 长白山次生林树种空间分布及环境解释[J]. 林业科学, 2008, 44(8):1−8. doi: 10.3321/j.issn:1001-7488.2008.08.001. Zhang C Y, Zhao X H, Xia F C. Spatial distribution of tree species and environmental interpretations of secondary forest in Changbai Mountains[J]. Scientia Silvae Sinicae, 2008, 44(8): 1−8. doi: 10.3321/j.issn:1001-7488.2008.08.001.
[21] 杨亮庆, 刘一星, 蔡英春. PEG400预处理枫桦圆盘干燥特性研究[J]. 西北林学院学报, 2013, 28(4):153−157. doi: 10.3969/j.issn.1001-7461.2013.04.31. Yang L Q, Liu Y X, Cai Y C. Drying characteristics of Betula costata disk pretreated with PEG400[J]. Journal of Northwest Forestry University, 2013, 28(4): 153−157. doi: 10.3969/j.issn.1001-7461.2013.04.31.
[22] 安慧君. 阔叶红松林空间结构研究[D]. 北京: 北京林业大学, 2003. An H J. Study on the spatial structure of the broad-leaved Korean pine forest[D]. Beijing: Beijing Forestry University, 2003.
[23] 刘宇, 徐焕文, 姜静, 等. 基于种子活力及苗期生长性状的白桦四倍体半同胞家系初选[J]. 北京林业大学学报, 2014, 36(2):74−80. Liu Y, Xu H W, Jiang J, et al. Family selection of birch tetraploid half-sibling based on seed vigor and seedling growth traits[J]. Journal of Beijing Forestry University, 2014, 36(2): 74−80.
[24] David F N, Moore P G. Notes on contagious distributions in plant populations[J]. Annals of Botany, 1954, 18(1): 47−53. doi: 10.1093/oxfordjournals.aob.a083381.
[25] Comita L S, Hubbell S P. Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank[J]. Ecology, 2009, 90(2): 328−334. doi: 10.1890/08-0451.1.
[26] 王蕾. 长白山针阔混交林种子雨动态及扩散限制研究[D]. 北京: 北京林业大学, 2010. Wang L. Seed rain dynamics and dispersal limitations in mixed broadleaf-conifer forest in Changbai Mountain[D]. Beijing: Beijing Forestry University, 2010.
[27] Yang Y C, Huang L, Qian S H, et al. Completing the life history of Castanopsis fargesii: changes in the seed dispersal, seedling and sapling recruitment patterns[J]. European Journal of Forest Research, 2015, 134(6): 1143−1154. doi: 10.1007/s10342-015-0916-9.
[28] 张健, 郝占庆, 李步杭, 等. 长白山阔叶红松(Pinus koraiensis)林种子雨组成及其季节动态[J]. 生态学报, 2008, 28(6):2445−2454. doi: 10.3321/j.issn:1000-0933.2008.06.002. Zhang J, Hao Z Q, Li B H, et al. Composition and seasonal dynamics of seed rain in broad-leaved Korean pine (Pinus koraiensis) mixed forest, Changbai Mountain[J]. Acta Ecologica Sinica, 2008, 28(6): 2445−2454. doi: 10.3321/j.issn:1000-0933.2008.06.002.
[29] 杜彦君, 马克平. 浙江古田山自然保护区常绿阔叶林种子雨的时空变异[J]. 植物生态学报, 2012, 36(8):717−728. Du Y J, Ma K P. Temporal and spatial variation of seedfall in a broad-leaved evergreen forest in Gutianshan Nature Reserve of Zhejiang Province, China[J]. Chinese Journal of Plant Ecology, 2012, 36(8): 717−728.
[30] 陈香茗, 赵秀海, 夏富才, 等. 长白山紫椴种子雨的时空分布格局[J]. 东北林业大学学报, 2011, 39(1):7−10. doi: 10.3969/j.issn.1000-5382.2011.01.003. Chen X M, Zhao X H, Xia F C, et al. Spatiotemporal dynamics of seed rain of Tilia amurensis in forest stands in Changbai Mountain[J]. Journal of Northeast Forestry University, 2011, 39(1): 7−10. doi: 10.3969/j.issn.1000-5382.2011.01.003.
[31] Saastamoinen M, Bocedi G, Cote J, et al. Genetics of dispersal[J]. Biological Reviews, 2018, 93(1): 574−599. doi: 10.1111/brv.12356.
[32] Masaki T, Nakashizuka T, Niiyama K, et al. Impact of the spatial uncertainty of seed dispersal on tree colonization dynamics in a temperate forest[J]. Oikos, 2019, 128(12): 1816−1828. doi: 10.1111/oik.06236.
[33] Lin L, Fu J. Relationship between the accumulation of storage proteins and formation of vigor of peanut seed[J]. Journal of Tropical and Subtropical Botany, 1996, 4(1): 57−60.
[34] 姜文, 姚大年, 张文明, 等. 小麦种子萌发过程贮藏蛋白变化及其与活力关系的研究[J]. 种子, 2006, 25(7):16−19. doi: 10.3969/j.issn.1001-4705.2006.07.005. Jiang W, Yao D N, Zhang W M, et al. Study on relationship between seed vigor and storage protein in germinating wheat seeds[J]. Seed, 2006, 25(7): 16−19. doi: 10.3969/j.issn.1001-4705.2006.07.005.
[35] Wendt A L. Seed dispersal, seed size and seed predation in tropical regrowth forests[D]. Connecticut: University of Connecticut-Storrs, 2014.
[36] 任坚毅, 林玥, 岳明. 太白山红桦种子的萌发特性[J]. 植物生态学报, 2008, 32(4):883−890. doi: 10.3773/j.issn.1005-264x.2008.04.017. Ren J Y, Lin Y, Yue M. Seed germination characteristics of Betula albosinensis at Mountain Taibai, China[J]. Chinese Journal of Plant Ecology, 2008, 32(4): 883−890. doi: 10.3773/j.issn.1005-264x.2008.04.017.
-
期刊类型引用(5)
1. 吕焕哲,彭渤,孙旭,陈希亚,胡雨萱. 国土空间规划视域下国家公园适宜范围识别研究——以拟建洞庭湖国家公园为例. 湖南师范大学自然科学学报. 2024(03): 37-44 .  百度学术
百度学术
2. 邱胜荣,唐小平,刘洋,缐海彦,黄璐,高琦,樊远征. 国家公园范围确定方法及在秦岭国家公园候选区的实践. 国家公园(中英文). 2024(10): 620-631 . 百度学术
3. 曾丽婷 ,戴冰武 ,曾毅 ,吕焕哲 ,谭畅 ,王一淳 . 基于生态安全格局的国家公园边界划定研究——以拟建洞庭湖国家公园为例. 国土资源导刊. 2023(01): 6-13 . 百度学术
4. 孙乔昀,李娜,张玉钧. 面向国土景观风貌管控的中国国家公园空间布局研究. 中国园林. 2022(04): 26-31 . 百度学术
5. 黄晗雯,刘洋,陈君帜,李晓民. 拟建松嫩鹤乡国家公园范围划定方法探讨. 风景园林. 2022(07): 26-29 . 百度学术
其他类型引用(3)


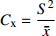
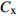
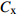
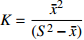
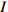
计量
- 文章访问数: 2152
- HTML全文浏览量: 607
- PDF下载量: 76
- 被引次数: 8
