
Ploidy, genotype and gender effects of functional leaf and stomatal traits on short branches in full-sib hybrids between section Tacamahaca and sect. Aigeiros of Populus
-
摘要:目的 探讨倍性水平、性别、基因型及环境效应对青黑杨全同胞杂种短枝功能叶片及气孔性状变异的影响,解析杨树异源三倍体的性状遗传与变异规律。方法 以‘哲引3号杨’× ‘北京杨’全同胞杂种二倍体和三倍体的10年生种质保存林为材料,对其短枝功能叶片和气孔相关性状变异进行分析。结果 短枝功能叶片和气孔各性状的无性系重复力均较高,介于0.789 ~ 0.960之间,受较强的遗传控制,环境影响较小。叶片和气孔各性状在无性系间均存在极显著差异,其中叶片宽度和气孔宽度变异系数较低,是所测性状中较稳定的遗传特征。三倍体植株叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度均极显著大于二倍体,气孔密度极显著小于二倍体,而三倍体植株的叶柄长度和锯齿数与二倍体无显著差异。叶片及气孔性状在不同性别植株之间均无显著差异。相关性分析结果显示,叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度与倍性水平呈极显著正相关;气孔密度与倍性水平呈极显著负相关;而叶柄长度和锯齿数与倍性水平无显著相关性。方差贡献率显示,无性系性别对叶片及气孔性状的影响最小;气孔性状受倍性效应影响最大,其次是基因型效应;叶片性状主要受基因型效应的影响,其次是倍性效应和环境效应。结论 青黑杨全同胞杂种三倍体植株短枝功能叶片及气孔性状变异主要来源于倍性效应和基因型效应,环境和性别影响较小,无性系间存在丰富的性状变异,在杨树三倍体育种时基于遗传测定进行“大群体,强选择”育种十分必要。Abstract:Objective This paper aims to analyze the inheritance and variation rules of allotriploid poplar, explore the effects of ploidy level, gender, genotype and environmental effects on the variation of short branch functional leaf and stomatal characters in full-sib diploid and triploid hybrids between section Tacamahaca and sect. Aigeiros of Populus.Method In this study, variations of short branch functional leaf and stomatal traits between full-sib diploid and triploid hybrids of 10-year-old clone experimental forest of P. pseudo-simonii × P. nigra ‘Zheyin3#’ and P. × beijingensis were analyzed.Result Clonal repeatability of short branch functional leaf and stomata was high, ranging from 0.789 to 0.960, which was controlled by strong heredity and had little environmental impact. There were very significant differences among clones in leaf and stomatal traits. The variation coefficient of leaf width and stomatal width was lower, which was a relatively stable genetic feature among the tested traits. The leaf length, leaf width, leaf area, stomatal length and stomatal width of triploid were extremely significantly larger than those of diploid, and the stomatal density was extremely significantly smaller than diploids, while the petiole length and serration number of triploids were not significantly different from diploid. There was no significant difference in leaf and stomatal traits between different gender plants. Correlation analysis results showed that leaf length, leaf width, leaf area, stomatal length and width were very significantly positively correlated with ploidy level, stomatal density was very significantly negatively correlated with ploidy level, while petiole length and serration number were not significantly correlated with ploidy level. The variance contribution rate showed that the gender effect of clones had the least influence on leaf and stomatal characters; stomatal traits were mostly affected by ploidy effect, followed by genotype effect; leaf traits were mainly affected by genotype effect, followed by ploidy and environmental effects.Conclusion The variation of short branch functional leaf and stomatal traits of full-sib hybrid between section Tacamahaca and sect. Aigeiros of Populus was mainly come from ploidy effect and genotype effect, but the influence of gender and environment was small. There were abundant trait variations among clones. It is necessary to carry out “large population, strong selection” breeding based on genetic determination in triploid breeding of Populus.
-
Keywords:
- poplar /
- allotriploid /
- ploidy effect /
- genotype effect /
- gender effect /
- functional leaf of short branch /
- stomata
-
多倍体育种是杨树(Populus spp.)遗传改良的重要途径。‘三毛杨’(P. tomentosa ‘Sanmaoyang’)、‘中林46’(P. × euramericana ‘Zhonglin-46’)、‘银中杨’(P. alba × P. berolinensis ‘Yinzhong’)、‘北林雄株1号’(P. × ‘Beilinxiongzhu 1’)、‘中怀1号’(P. × ‘Zhonghuai 1’)等三倍体品种均在我国林业生产中广泛利用[1-5],对推动我国林业产业升级、改善生态环境做出了重要贡献。为了指导林木多倍体品种选育,朱之悌[6]曾提出了“大群体,强选择”的育种策略,认为多倍体育种同样要基于大群体,依据主要目标性状对多倍体种质进行选择,经过遗传测定,才能形成表现优良的新品种。因此,在杨树多倍体品种选育过程中,对性状变异规律进行遗传分析十分必要。
就杨树多倍体育种而言,基于2n配子的有性多倍化途径综合了杂合效应和倍性效应,提供了更加丰富的遗传变异[7]。植物生长通过叶片的光合作用产生物质和能量[8],叶片大小和形状影响植物的气体交换和蒸腾作用[9],有关植物叶片和气孔性状的研究可间接反映植物的植物生长发育及生理代谢特征[10]。Liao等[11]分析了‘哲引3号杨’ × ‘北京杨’全同胞杂种三倍体和二倍体的苗期表型,发现三倍体群体整体具有生长和光合优势,其中也存在一些光合效率高但生长较差的个体和光合效率低但生长较好的个体,可见,异源三倍体杨树的性状变异规律非常复杂。已有研究表明,雌雄异株植物在叶面积、生长势、生物量等方面存在差异[12],性状表现也会受到性别的影响。然而,关于叶片及气孔性状的研究多集中在苗期植株的分析,尚缺少对于节间生长缓慢且已进入开花成熟期的杨树枝条上的短枝功能叶片及气孔性状的变异规律的研究,且对于异源三倍体杨树,性别效应对性状表现产生的影响尚不明确。因此,为进一步丰富杨树三倍体叶片及气孔性状的遗传变异特征,本研究以‘哲引3号杨’ × ‘北京杨’全同胞杂种二倍体和三倍体的10年生种质保存林为材料,对其大树短枝功能叶片及气孔性状变异规律进行系统研究,解析倍性、基因型和性别效应对叶片及气孔性状变异的影响大小,为杨树三倍体育种提供理论依据。
1. 材料与方法
1.1 试验材料
试验材料采集自河北威县的‘哲引3号杨’ × ‘北京杨’全同胞杂种二倍体和三倍体种质保存林,4株小区,3次重复,树龄10年,其中包括二倍体无性系17个(雌株7个,编号为D9、D24、D25、D34、D44、D45、D52;雄株10个,编号为D2、D3、D4、D11、D14、D20、D21、D38、D41、D43),三倍体无性系19个(雌株9个,编号为T6、T12、T15、T16、T19、T23、T26、T32、T40;雄株10个,编号为T13、T18、T22、T27、T30、T31、T33、T36、T37、T42)。每个无性系从每小区的4株中随机选择1株作为采样树,共3次重复,采集位于树冠中部北侧的无病虫害短枝功能叶片进行叶片长度、叶片宽度、叶面积、叶柄长度、叶缘锯齿数、气孔长度、气孔宽度、气孔密度等性状测量。
1.2 试验方法
1.2.1 短枝叶片性状测量
每株采样树随机选取30片形态完整的短枝叶片,每个无性系重复3株采样树,用CI-203便携式激光叶面积仪(美国CID)测量叶片长度、叶片宽度、叶面积和叶柄长度,并对叶缘锯齿数进行统计。
1.2.2 气孔性状
每株采样树随机选取5片形态完整的短枝叶片,每个无性系重复3株采样树,采用指甲油涂抹撕取法进行气孔性状观察[13]。每个叶片随机选择50个处于关闭状态的气孔,于Olympus BX51显微镜下测量气孔长度和气孔宽度;随机选择10个视野统计气孔密度。
1.2.3 数据统计分析
利用Excel整理数据,由于各性状数据均不满足正态分布或方差齐次,应用SPSS 17.0软件的非参数Kruskal−Wallis检验进行差异显著性分析。应用R语言对各性状进行Spearman相关分析和方差贡献率的计算,将方差分量的剩余项视为环境效应。
无性系重复力(R)计算公式:
R=σc2/(σc2+σe2/k) 式中:σc2为无性系方差,σe2为随机误差,k为每个无性系采集样本数的调和平均值[14]。
2. 结果与分析
2.1 青黑杨全同胞杂种短枝叶片及气孔性状基因型间的变异分析
青黑杨全同胞杂种植株短枝叶片及气孔各性状在无性系之间均存在极显著差异(表1)。无性系之间叶片性状变异较大,变异系数介于14.23% ~ 27.20%之间,其中锯齿数变异系数最高,叶片宽度变异系数最小;气孔性状变异系数在8.73% ~ 32.73%之间,其中气孔密度变异系数最高,气孔宽度变异系数最低,表明叶片宽度和气孔宽度是所测性状中较稳定的遗传特征。
表 1 ‘哲引3号杨’ × ‘北京杨’杂种二倍体和三倍体叶片及气孔性状无性系重复力及变异系数Table 1. Repeatability and variation coefficients of leaves and stomatal traits of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis性状 Trait 二倍体 Diploid 三倍体 Triploid P值 P value 无性系重复力 Repeatability of clone 变异系数 CV/% 变化范围
Variation range均值 ± 标准误 Mean ± SE 变化范围
Variation range均值 ± 标准误 Mean ± SE 倍性 Ploidy 性别Gender 基因型Genotype 叶片长度
Leaf length/cm6.26 ~ 9.35 7.52 ± 0.14 6.59 ~ 10.51 8.65 ± 0.18 < 0.01 0.86 < 0.01 0.903 15.23 叶片宽度
Leaf width/cm4.83 ~ 7.48 6.05 ± 0.13 5.45 ~ 7.92 6.85 ± 0.11 < 0.01 0.33 < 0.01 0.836 14.23 叶面积
Leaf area/cm217.76 ~ 39.92 27.32 ± 0.97 21.61 ~ 41.97 33.41 ± 1.09 < 0.01 0.72 < 0.01 0.789 25.32 叶柄长度
Petiole length/cm3.32 ~ 5.33 4.35 ± 0.10 3.28 ~ 5.95 4.48 ± 0.12 0.41 0.29 < 0.01 0.829 16.95 锯齿数
Serration number19 ~ 74 42.1 ± 2.0 30 ~ 55 38.4 ± 1.1 0.10 0.34 < 0.01 0.924 27.20 气孔长度
Stomatal length/μm20.67 ~ 28.29 23.58 ± 0.30 24.08 ~ 30.66 27.59 ± 0.30 < 0.01 0.79 < 0.01 0.954 11.13 气孔宽度
Stomatal width/μm12.82 ~ 16.17 14.53 ± 0.14 14.15 ~ 17.66 16.27 ± 0.16 < 0.01 0.28 < 0.01 0.952 8.73 气孔密度/(个·mm−2)
Stomatal density/
(number·mm−2)87 ~ 279 188.6 ± 8.1 95 ~ 191 128.5 ± 3.8 < 0.01 0.06 < 0.01 0.947 32.73 重复力作为一个重要的遗传参数,反映了无性系优良性状受特殊环境影响的程度[15],其大小还表明了性状稳定性的高低[16]。本研究中,叶片性状无性系重复力介于0.789 ~ 0.924之间,气孔性状无性系重复力介于0.947 ~ 0.960之间,气孔性状的遗传效应略大于叶片性状。可见,青黑杨全同胞杂种植株叶片和气孔性状受到强度的遗传控制,环境影响较小,有利于无性系的评价选择,更利于较为准确地解析各效应对性状变异的影响。
2.2 青黑杨全同胞杂种植株短枝叶片性状变异规律分析
不同倍性群体间比较,三倍体植株平均叶片长度、叶片宽度、叶面积均显著大于二倍体植株,呈现出一定的巨大性特征(表1,图1a、b);而叶柄长度和锯齿数性状则无显著差异(表1)。在不同性别间,尽管雄株的叶片宽度、叶面积和叶柄长度比雌株分别大3.14%、2.02%和3.70%,叶片长度和锯齿数分别小1.10%和1.97%,但是这些叶片性状在雌、雄株之间并未呈现显著差异。
![]() 图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis
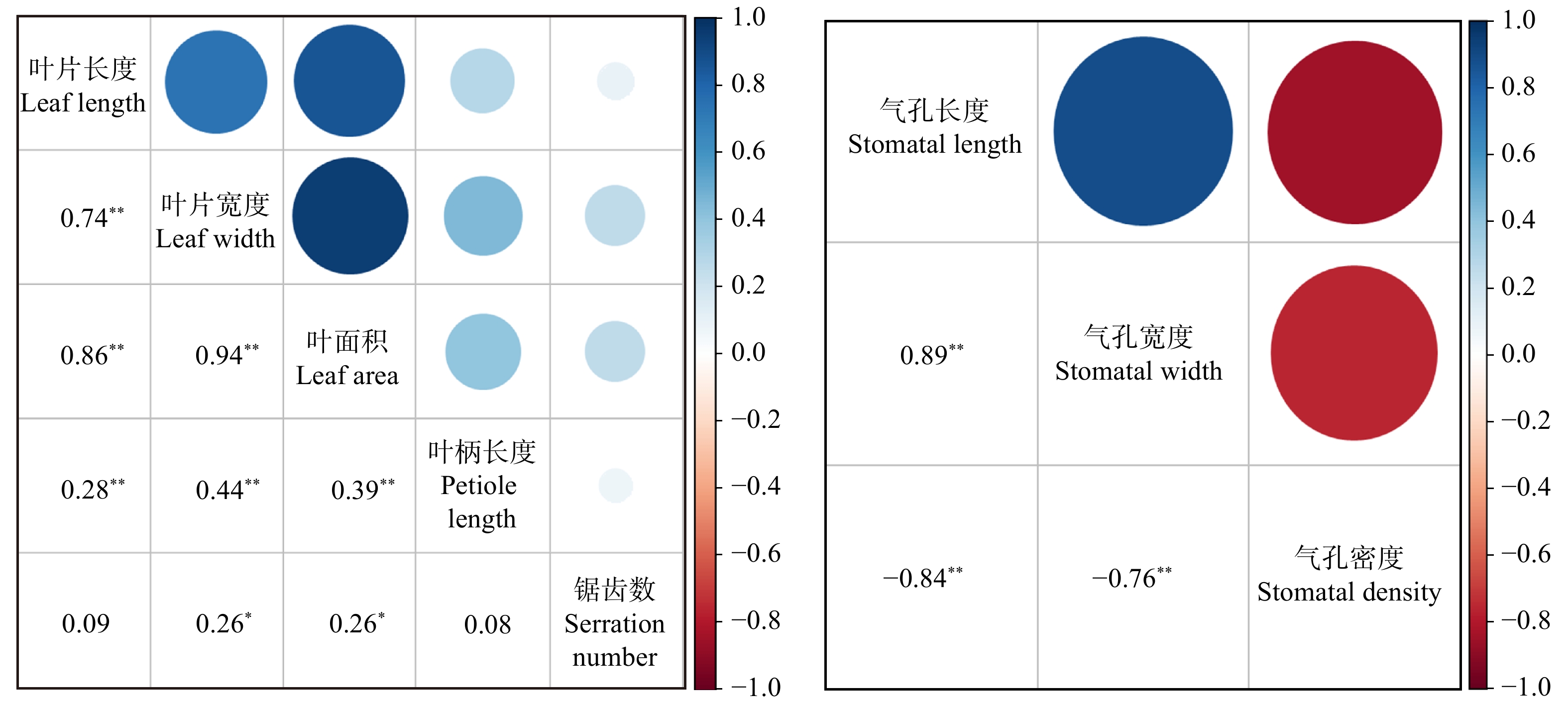
图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis从叶片各性状相关分析可知(图2),叶面积与叶片长度、叶片宽度呈极显著的正相关,分别达0.86和0.94;叶柄长度与叶片长度、叶片宽度、叶面积之间呈显著弱正相关,与锯齿数相关关系不显著;锯齿数与叶片宽度、叶面积呈显著弱正相关,与叶片长度的相关关系不显著。进一步分析发现(表2),叶片长度、叶片宽度和叶面积与倍性水平呈极显著的正相关(r分别为0.44、0.43、0.38,P < 0.01),叶柄长度和锯齿数与倍性水平之间无显著相关性(分别为r = 0.09,P = 0.41和r = −0.17,P = 0.10),而性别与叶片性状之间均没有显著的相关性(P > 0.05)。
![]() 图 2 叶片和气孔性状的Spearman相关分析*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.Figure 2. Spearman correlation analysis of leaves and stomatal traits表 2 叶片及气孔性状与倍性、性别的相关性分析Table 2. Correlation analysis of leaves and stomatal traits with ploidy and gender
图 2 叶片和气孔性状的Spearman相关分析*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.Figure 2. Spearman correlation analysis of leaves and stomatal traits表 2 叶片及气孔性状与倍性、性别的相关性分析Table 2. Correlation analysis of leaves and stomatal traits with ploidy and gender项目
Item叶片长度
Leaf length叶片宽度
Leaf width叶面积
Leaf area叶柄长度
Petiole length锯齿数
Serration number气孔长度
Stomatal length气孔宽度
Stomatal width气孔密度
Stomatal density倍性 Ploidy r值 r value 0.44 0.43 0.38 0.09 −0.17 0.72 0.65 −0.60 P值 P value < 0.01 < 0.01 < 0.01 0.41 0.10 < 0.01 < 0.01 < 0.01 性别 Gender r值 r value −0.02 0.10 0.04 0.11 −0.10 0.03 −0.11 0.20 P值 P value 0.86 0.33 0.72 0.29 0.34 0.80 0.28 0.06 2.3 青黑杨全同胞杂种植株短枝叶气孔性状变异规律分析
三倍体植株的平均气孔长度和气孔宽度均极显著大于二倍体,而平均气孔密度极显著小于二倍体(表1,图1c、d)。在不同性别间比较发现,雄株群体的平均气孔长度和气孔密度均大于雌株,气孔宽度小于雌株,但是均为呈现显著差异。
从气孔各性状的相关分析可知(图2),气孔长度与气孔宽度呈极显著的正相关,达0.89,气孔密度与气孔长度和气孔宽度呈极显著的负相关,分别为−0.84和−0.76。进一步分析发现(表2),气孔长度和气孔宽度与倍性水平间均呈极显著正相关(r分别为0.72和0.65,P < 0.01),气孔密度与倍性水平呈极显著负相关(r = −0.60,P < 0.01),而性别与气孔性状之间均没有显著的相关性(P > 0.05),表明气孔性状与倍性水平密切相关,可作为鉴定杨树多倍体的有效指标。
2.4 影响青黑杨全同胞杂种叶片及气孔性状各效应的方差贡献率解析
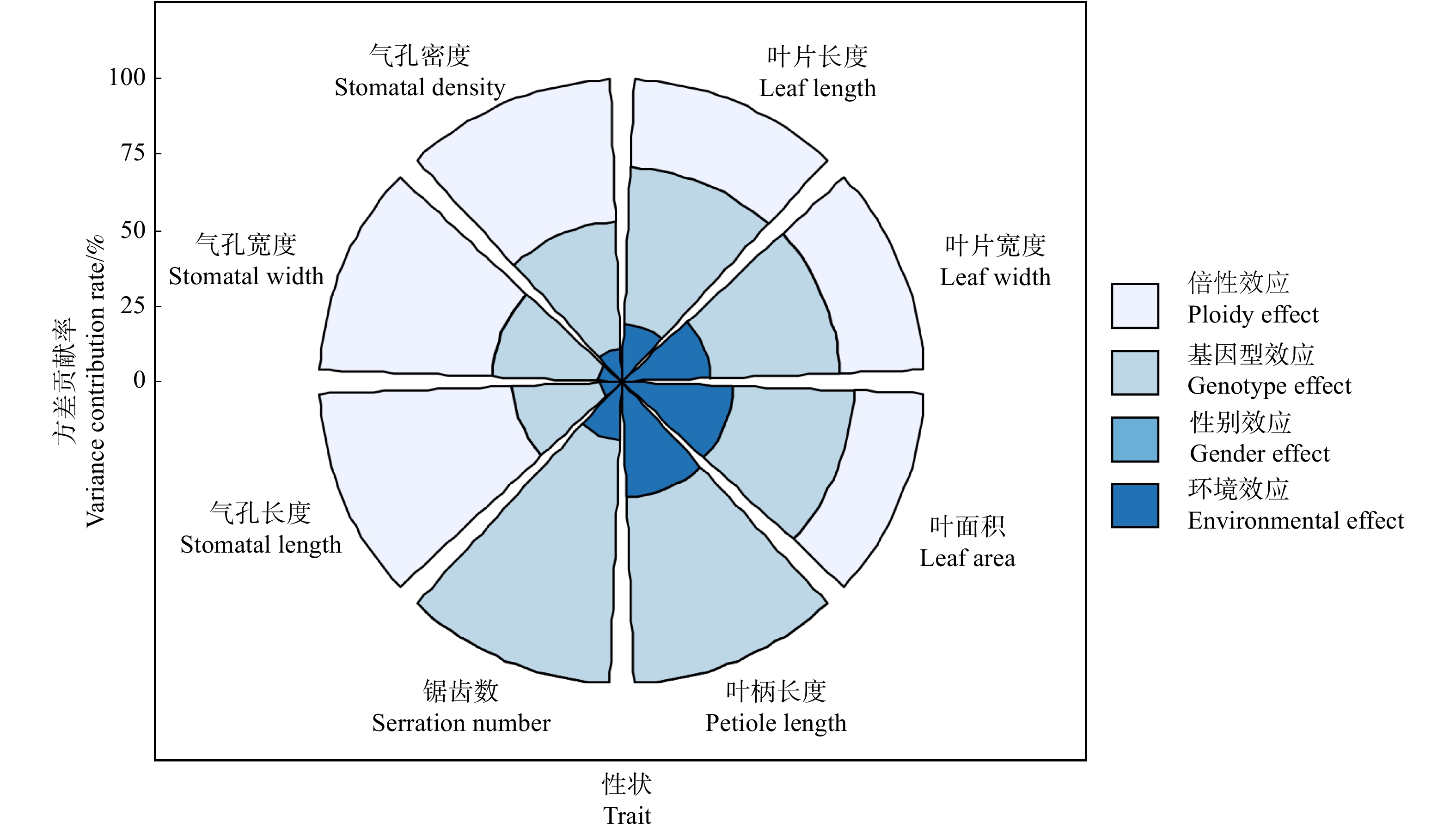
为深入解析倍性、性别和基因型效应对各性状表现的影响程度,计算各效应的方差分量及方差贡献率后发现(图3),气孔性状的变异主要来源于倍性效应,其次是基因型效应和环境效应。其中,气孔长度的倍性效应方差分量在表型总变异中所占比例高达63.79%,其次是气孔宽度的57.05%,气孔密度的47.10%,表明这些性状受倍性水平的影响最大;叶片性状变异的主要来源于基因型效应,其中,锯齿数变异的基因型效应方差贡献率最高,达80.72%,其次是叶柄长度变异的基因型效应为61.66%,表明叶片性状主要受基因型的影响。性别效应对叶片及气孔性状的影响均很小,其中,性别效应对气孔密度的方差贡献率仅为0.06%,对锯齿数和叶柄长度变异的方差贡献率甚至均为0。
3. 讨 论
多倍化是植物进化和适应性改变的重要力量[17-18]。由于基因组剂量的增加所带来的倍性效应往往导致多倍体植物产生广泛的性状变异[19-20]。同源四倍体苏丹凤仙花(Impatiens walleriana)在叶面积、叶片厚度、子房宽度、花粉粒长度和气孔大小等性状上增加,花瓣总花青素含量较高,而株高、气孔保卫细胞密度和花数减少[21]。滇杨(P. yunnanensis)同源四倍体和嵌合体的苗期长枝叶分析发现,与叶片长度、宽度和气孔性状大于二倍体,而气孔密度则显著小于二倍体[22]。本研究发现青黑杨杂种的叶柄长度和锯齿数变异受倍性效应影响较小,而三倍体植株平均叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度均极显著大于二倍体,气孔密度则极显著小于二倍体,且气孔性状变异的倍性效应方差贡献率最高,表明倍性效应对短枝叶的性状表现同样产生重要影响。而且,叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度与倍性水平之间均存在极显著正相关性,气孔密度与倍性水平呈极显著负相关,叶柄长度和锯齿数与倍性水平无显著相关性,因此,在倍性检测时可通过叶片大小和气孔性状相结合作为初步鉴定。此外,值得注意的是,本研究测得的气孔长度和气孔宽度大小与王君等[23]测得的数据相比均偏小,可能是由于杨树长枝和短枝叶片的气孔长宽大小存在差异。
基因型差异也是影响植物性状的重要变异来源。白凤莹等[24]发现,受基因型的影响,不同天然三倍体毛白杨无性系间气孔性状的差异达到了极显著水平,其中存在部分个体表现出类似二倍体的特征。本研究发现,青黑杨全同胞杂种无性系间短枝叶片和气孔各性状均存在极显著差异,其中部分二倍体无性系的叶片和气孔性状表现甚至优于一些三倍体无性系,可见三倍体也并非株株皆优,无性系基因型对性状表现的影响也较大。
对于雌雄异株植物而言,植株性别会对表型产生一定的影响。相关研究表明,多年生植物雄株在叶片数量及叶面积、生长势、树冠体积、生物量等方面都超过雌株[12, 25]。陈珂等[26]发现成年银杏(Ginkgo biloba)雌株叶片长度、叶片宽度、叶面积显著小于雄株,而叶柄长度和气孔密度显著大于雄株。两年生青杨(P. cathayana)雌株叶片总数显著高于雄株,雌株叶片性状较雄株有优势[27];毛白杨雄株叶片长度和叶面积显著大于雌株,而叶片宽度则无显著差异[28]。本研究中,青黑杨全同胞杂种叶片和气孔性状在雌、雄株之间虽存在一定差异,但均未达显著性水平,性状变异与性别之间也不存在显著相关性,且性别效应方差贡献率几乎为0,说明性别对叶片及气孔性状的影响较小。
异源多倍化在植物遗传改良和种质创新中发挥着重要作用,特别是利用2n配子杂交的异源多倍化途径,结合了倍性优势和杂种优势,在杨树多倍体育种已取得了极大的成就[29]。Liao等[11]发现,杨树异源三倍体群体苗期整体具有生长和光合优势,但也存在一些光合效率高但生长较差的个体和光合效率低但生长较好的个体。本研究利用一个包含杂种二倍体和三倍体的青黑杨全同胞群体,证明了杨树经历异源多倍化后,倍性效应和基因型效应均对性状变异产生重要影响,而性别对性状变异的贡献相对较小,并初步明确了倍性、基因型和性别效应对功能叶片和气孔性状的贡献大小,为我们进一步开展多倍体性状变异的遗传解析提供了借鉴。显然,在进行杨树三倍体育种时,对大量变异材料进行“大群体,强选择”是进一步品种选育的必要环节,异源三倍体遗传变异的复杂性也为杨树三倍体选育提供了丰富的选择基础。
-
![]()
图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔
a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.
Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis
![]()
图 2 叶片和气孔性状的Spearman相关分析
*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.
Figure 2. Spearman correlation analysis of leaves and stomatal traits
表 1 ‘哲引3号杨’ × ‘北京杨’杂种二倍体和三倍体叶片及气孔性状无性系重复力及变异系数
Table 1 Repeatability and variation coefficients of leaves and stomatal traits of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis
性状 Trait 二倍体 Diploid 三倍体 Triploid P值 P value 无性系重复力 Repeatability of clone 变异系数 CV/% 变化范围
Variation range均值 ± 标准误 Mean ± SE 变化范围
Variation range均值 ± 标准误 Mean ± SE 倍性 Ploidy 性别Gender 基因型Genotype 叶片长度
Leaf length/cm6.26 ~ 9.35 7.52 ± 0.14 6.59 ~ 10.51 8.65 ± 0.18 < 0.01 0.86 < 0.01 0.903 15.23 叶片宽度
Leaf width/cm4.83 ~ 7.48 6.05 ± 0.13 5.45 ~ 7.92 6.85 ± 0.11 < 0.01 0.33 < 0.01 0.836 14.23 叶面积
Leaf area/cm217.76 ~ 39.92 27.32 ± 0.97 21.61 ~ 41.97 33.41 ± 1.09 < 0.01 0.72 < 0.01 0.789 25.32 叶柄长度
Petiole length/cm3.32 ~ 5.33 4.35 ± 0.10 3.28 ~ 5.95 4.48 ± 0.12 0.41 0.29 < 0.01 0.829 16.95 锯齿数
Serration number19 ~ 74 42.1 ± 2.0 30 ~ 55 38.4 ± 1.1 0.10 0.34 < 0.01 0.924 27.20 气孔长度
Stomatal length/μm20.67 ~ 28.29 23.58 ± 0.30 24.08 ~ 30.66 27.59 ± 0.30 < 0.01 0.79 < 0.01 0.954 11.13 气孔宽度
Stomatal width/μm12.82 ~ 16.17 14.53 ± 0.14 14.15 ~ 17.66 16.27 ± 0.16 < 0.01 0.28 < 0.01 0.952 8.73 气孔密度/(个·mm−2)
Stomatal density/
(number·mm−2)87 ~ 279 188.6 ± 8.1 95 ~ 191 128.5 ± 3.8 < 0.01 0.06 < 0.01 0.947 32.73  下载: 导出CSV
下载: 导出CSV
表 2 叶片及气孔性状与倍性、性别的相关性分析
Table 2 Correlation analysis of leaves and stomatal traits with ploidy and gender
项目
Item叶片长度
Leaf length叶片宽度
Leaf width叶面积
Leaf area叶柄长度
Petiole length锯齿数
Serration number气孔长度
Stomatal length气孔宽度
Stomatal width气孔密度
Stomatal density倍性 Ploidy r值 r value 0.44 0.43 0.38 0.09 −0.17 0.72 0.65 −0.60 P值 P value < 0.01 < 0.01 < 0.01 0.41 0.10 < 0.01 < 0.01 < 0.01 性别 Gender r值 r value −0.02 0.10 0.04 0.11 −0.10 0.03 −0.11 0.20 P值 P value 0.86 0.33 0.72 0.29 0.34 0.80 0.28 0.06
下载: 导出CSV
-
[1] 朱之悌, 林惠斌, 康向阳. 毛白杨异源三倍体B301等无性系选育的研究[J]. 林业科学, 1995, 31(6):499. Zhu Z T, Lin H B, Kang X Y. Studied on allotriploid breeding of Populus tomentosa B301 clones[J]. Scientia Silvae Sinicae, 1995, 31(6): 499.
[2] Zhang S, Qi L, Chen C, et al. A report of triploid Populus of the section Aigeiros[J]. Silvae Genetica, 2004, 53(2): 69−75.
[3] 张守攻, 陈成彬, 韩素英, 等. 中国部分杨属植物的染色体数目[J]. 植物分类学报, 2005, 43(6):539−544. doi: 10.1360/aps040008 Zhang S G, Chen C B, Han S Y, et al. Chromosome numbers of some Populus taxa from China[J]. Acta Phytotaxonomica Sinica, 2005, 43(6): 539−544. doi: 10.1360/aps040008
[4] 贾会霞, 姬慧娟, 胡建军, 等. 杨树新品种的SSR指纹图谱构建和倍性检测[J]. 林业科学, 2015, 51(2):69−79. Jia H X, Ji H J, Hu J J, et al. Fingerprints of SSR markers and ploidy detection for new Populus varieties[J]. Scientia Silvae Sinicae, 2015, 51(2): 69−79.
[5] Kang X Y. Polyploid induction techniques and breeding strategies in poplar[M]// Mason A S. Polyploidy and hybridization for crop improvement. Boca Raton: CRC Press, 2016: 76−96.
[6] 朱之悌. 毛白杨遗传改良[M]. 北京: 中国林业出版社, 2006. Zhu Z T. Genetic improvement of Populus tomentosa[M]. Beijing: China Forestry Publishing House, 2006.
[7] 康向阳. 关于杨树多倍体育种的几点认识[J]. 北京林业大学学报, 2010, 32(5):149−153. Kang X Y. Some understandings on polyploid breeding of poplars[J]. Journal of Beijing Forestry University, 2010, 32(5): 149−153.
[8] Zhu X G, Long S P, Ort D R. Improving photosynthetic efficiency for greater yield[J]. Annual Review of Plant Biology, 2010, 61: 235−261. doi: 10.1146/annurev-arplant-042809-112206
[9] Tsukaya H. Leaf shape: genetic controls and environmental factors[J]. International Journal of Developmental Biology, 2005, 49(5−6): 547−555. doi: 10.1387/ijdb.041921ht
[10] 苏晓华, 李金花, 陈伯望, 等. 杨树叶片数量性状相关联标记及其图谱定位研究[J]. 林业科学, 2000, 36(1):33−40. Su X H, Li J H, Chen B W, et al. Detection and identification of molecular markers associated with quantitative traits of leaf in poplar[J]. Scientia Silvae Sinicae, 2000, 36(1): 33−40.
[11] Liao T, Cheng S P, Zhu X H, et al. Effects of triploid status on growth, photosynthesis, and leaf area in Populus[J]. Trees, 2016, 30(4): 1137−1147. doi: 10.1007/s00468-016-1352-2
[12] Barrett S C H, Hough J. Sexual dimorphism in flowering plants[J]. Journal of Experimental Botany, 2013, 64(1): 67−82. doi: 10.1093/jxb/ers308
[13] Christopher S C. Laboratory techniques for determining ploidy in plants[J]. HortTechnology, 1999, 9(4): 594−596. doi: 10.21273/HORTTECH.9.4.594
[14] Zhang P D, Wu F, Kang X Y. Genetic control of fiber properties and growth in triploid hybrid clones of Populus tomentosa[J]. Scandinavian Journal of Forest Research, 2013, 28(7): 621−630. doi: 10.1080/02827581.2013.829868
[15] 彭兴民, 吴疆翀, 郑益兴, 等. 印楝无性系当代种子农药品质性状的遗传变异及农药型印楝优株评价[J]. 林业科学研究, 2017, 30(6):921−928. Peng X M, Wu J C, Zheng Y X, et al. Genetic variation of azadirachtin quality in seeds of neem clones and evaluation of superior neem trees for pesticides[J]. Forest Research, 2017, 30(6): 921−928.
[16] 王琦, 朱之悌. 林木无性系育种若干遗传参数估算的研究进展[J]. 林业科学, 1995, 31(2):169−176. Wang Q, Zhu Z T. Advances in research of genetic parameter estimations for clonal breeding of forest trees[J]. Scientia Silvae Sinicae, 1995, 31(2): 169−176.
[17] Ruprecht C, Lohaus R, Vanneste K, et al. Revisiting ancestral polyploidy in plants[J/OL]. Science Advances, 2017, 3(7): e1603195 [2020−03−25]. https://advances.sciencemag.org/content/3/7/e1603195.full.
[18] Van Drunen W E, Husband B C. Immediate vs. evolutionary consequences of polyploidy on clonal reproduction in an autopolyploid plant[J]. Annals of Botany, 2018, 122(1): 195−205. doi: 10.1093/aob/mcy071
[19] Münzbergová Z. Colchicine application significantly affects plants performance in the second generation of synthetic polyploids and its effects vary between populations[J]. Annals of Botany, 2017, 120(2): 329−339. doi: 10.1093/aob/mcx070
[20] Ding M Q, Chen Z J. Epigenetic perspectives on the evolution and domestication of polyploid plant and crops[J]. Current Opinion in Plant Biology, 2018, 42(1): 37−48.
[21] Ghanbari M A, Jowkar A, Salehi H, et al. Effects of polyploidization on petal characteristics and optical properties of Impatiens walleriana (Hook.)[J]. Plant Cell, Tissue and Organ Culture, 2019, 138(2): 299−310. doi: 10.1007/s11240-019-01625-3
[22] 唐军荣, 李斌, 朱丽娜, 等. 滇杨多倍体苗期叶片形态及光合生理比较分析[J]. 林业科学研究, 2016, 29(1):103−109. Tang J R, Li B, Zhu L N, et al. Analysis on leaf morphology and photosynthesis physiology of polyploidized Populus yunnanensis seedlings[J]. Forest Research, 2016, 29(1): 103−109.
[23] 王君, 康向阳. 青杨派树种多倍体诱导技术研究[M]. 北京: 中国环境出版社, 2017. Wang J, Kang X Y. Study on polyploid induction technology of Tacamahaca[M]. Beijing: China Environmental Science Press, 2017.
[24] 白凤莹, 曾青青, 康宁, 等. 毛白杨基因库优树倍性检测及性状对比分析[J]. 北京林业大学学报, 2015, 37(4):113−119. Bai F Y, Zeng Q Q, Kang N, et al. Ploidy level and contrast analysis of the traits for superior trees of Populus tomentosa Carr. in gene pool[J]. Journal of Beijing Forestry University, 2015, 37(4): 113−119.
[25] Li C, Xu G, Zang R, et al. Sex-related differences in leaf morphological and physiological responses in Hippophae rhamnoides along an altitudinal gradient[J]. Tree Physiology, 2007, 27(3): 399−406. doi: 10.1093/treephys/27.3.399
[26] 陈珂, 刘霁瑶, 蒋园园, 等. 成年银杏雌雄株非花果期形态学差异性比较分析[J]. 西南大学学报(自然科学版), 2017, 39(7):17−24. Chen K, Liu J Y, Jiang Y Y, et al. Analysis of morphological differences of the leaves of adult ginkgo trees in the non-flowering and non-fruiting period[J]. Journal of Southwest University (Natural Science Edition), 2017, 39(7): 17−24.
[27] 许馨文, 肖娟, 田茂洁, 等. 青杨无性系雌雄植株幼苗的形态及叶绿素含量差异[J]. 光谱实验室, 2009, 26(6):1646−1649. Xu X W, Xiao J, Tian M J, et al. Differences in morphology and chlorophyll content between male and female clonal seedlings of Populus cathayana[J]. Chinese Journal of Spectroscopy Laboratory, 2009, 26(6): 1646−1649.
[28] Du Q Z, Xu B H, Gong C R, et al. Variation in growth, leaf, and wood property traits of Chinese white poplar (Populus tomentosa), a major industrial tree species in northern China[J]. Canadian Journal of Forest Research, 2014, 44(4): 326−339. doi: 10.1139/cjfr-2013-0416
[29] 康向阳, 王君. 杨树多倍体诱导技术研究[M]. 北京: 科学出版社, 2010. Kang X Y, Wang J. Poplar polyploid induction technology research[M]. Beijing: Science Press, 2010.
-
期刊类型引用(4)
1. 赵钰婷,陈冬瑶,杨柳,李晶楠,宁广亮,姜静. 白桦四倍体×紫雨桦二倍体杂交种子活力及杂种子代生长特性分析. 温带林业研究. 2025(01): 1-8 .  百度学术
百度学术
2. 马鸿文,任宇昕,龙羿辛,王楠,冯祥元,俞天泉,华晓琴,王君. 青黑杨杂种全同胞二倍体与三倍体长枝叶性状变异研究. 北京林业大学学报. 2024(01): 27-34 . 本站查看
3. 王舒扬,田力,周顺陶,储月娥,梅迪,袁佳秋,余延浩,洑香香. 多倍化对青钱柳叶形态、光合性能和次生代谢产物积累的影响. 林业科学. 2024(08): 120-131 . 百度学术
4. 卞瑶,耿丽妍,冯彦博,陈福燕,张雯,王宏伟. 不同进化历程下的异源四倍体小麦核型进化及其稳定性分析. 辽宁师范大学学报(自然科学版). 2023(03): 363-369 . 百度学术
其他类型引用(5)
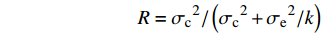

计量
- 文章访问数: 1329
- HTML全文浏览量: 375
- PDF下载量: 60
- 被引次数: 9
