
Canopy characteristics in gaps and its relationship with seedlings and saplings in a spruce-fir forest in the Changbai Mountain area of northeastern China
-
摘要:目的 林隙在森林更新循环中起着重要作用,对林隙内冠层结构和光因子及其与幼苗幼树的数量和生长指标之间的关系进行分析,以探究林隙冠层特征对幼苗幼树的影响,提出促进云冷杉林天然更新的措施,为云冷杉林可持续经营和资源可持续利用提供科学依据。方法 本研究于2019年8月对吉林省金沟岭林场长白山云冷杉异龄针阔混交林林分内48个林隙的冠层结构、光因子及幼苗幼树进行调查,采用Pearson相关性分析对不同大小林隙内冠层结构和光因子及其与幼苗幼树的关系进行分析。结果 (1)林隙冠层开度(CO)、叶面积指数(LAI)和总辐射(Tot)主要分布区间分别为12% ~ 20%、2 ~ 3和11% ~ 21%。(2)CO、LAI及光因子在不同大小林隙内差异显著(P < 0.05),CO、散射辐射(TDF)随林隙增大而增大,Tot和直接辐射随林隙增大先减小后增加。(3)各树种幼苗幼树株数随CO和Tot的增大先增加后减少,最适合幼苗幼树存活的CO和Tot区间分别为12% ~ 20%和11% ~ 21%。(4)林隙面积小于120 m2时最适合冷杉幼苗幼树的生长;小于90 m2时最适合色木槭幼苗幼树的生长;云杉幼苗幼树在小于30 m2及60 ~ 90 m2的林隙中生长最好;最适宜红松幼苗幼树生长的林隙面积为60 ~ 90 m2。结论 该林分主要冠层分布区间比较适宜4个树种更新幼苗幼树的生长,当CO和Tot超过一定范围后,幼苗幼树数量反而下降。因此,可以根据林分经营需要减少林隙内更新幼苗幼树的数量,或对林隙面积进行适当调节,为目的树种释放生长空间,促进其更新。Abstract:Objective The formation of gaps plays an important role in the forest renewal cycle. The relationship between structure and light factors of canopy in the gap and its relationship with the number and growth indicators of seedlings and saplings were analyzed. In order to explore the impact of characteristics of the gap canopy on seedlings and saplings, the measures to promote the natural renewal of spruce-fir forests were put forward to provide scientific basis for the sustainable management and sustainable use of spruce-fir forests.Method This study investigated the canopy structure, light factors, seedlings and saplings of 48 gaps in the spruce-fir heterogeneous mixed forest of Changbai Mountain in Jingouling Forest Farm, Jilin Province of northeastern China in August 2019. Pearson correlation analysis was used to analyze the canopy structure and light factors in gaps of different sizes and their relationship with seedlings and saplings.Result (1) The main distribution intervals of canopy openness (CO), leaf area index (LAI) and trans total radiation (Tot) were 12%−20%, 2−3 and 11%−21%, respectively. (2) CO, LAI and light factors were significantly different in gaps of different sizes (P < 0.05), CO and trans diffuse radiation (TDF) increased with the increase of forest gap, Tot and trans direct radiation (TDR) first decreased and then increased with the increase of gap. (3) The number of seedlings and saplings of each tree species increased first and then decreased with the increase of CO and Tot. The most suitable CO and Tot for the survival of seedlings and saplings were 12%−20% and 11%−21%, respectively. (4) The gap less than 120 m2 was most suitable for the growth of fir seedlings and saplings; while the gap less than 90 m2 was the most suitable for the growth of Acer seedlings and saplings; spruce seedlings and saplings grew best in gaps less than 30 m2 and 60−90 m2; the most suitable gap area for the growth of Korean pine seedlings and saplings was 60−90 m2.Conclusion The main canopy distribution interval of the forest is more suitable for the growth of seedlings and saplings of the four tree species, but when the CO and Tot exceed a certain range, the number of seedlings and saplings decreases. Therefore, the number of seedlings and saplings in the gap can be reduced according to the needs of forest management, or the area of gap can be appropriately adjusted to release growth space for the target tree species and promote its regeneration.
-
Keywords:
- gap /
- canopy structure /
- light factor /
- seedling and sapling
-
松栎柱锈菌(Cronartium orientale)是导致樟子松(Pinus sylvestris var. mongolica)、兴凯湖松(Pinus takahasii)和马尾松(Pinus massoniana)等2针松瘤锈病的森林有害生物,是一种转主寄生菌,其春孢子阶段寄生在松树枝干部[1-2]。当松树被松栎柱锈菌侵染致病后,其病原菌就会长期寄生在松树枝干部,并且病部肿瘤会逐年增大,给林分健康和林木生长造成极大危害[3]。黄酮类化合物具有较高的生物活性,是一种强抗氧化剂,能有效清除自由基从而延缓衰老和阻止细胞衰老、癌变,对人类心脑血管疾病也有很好的治疗和预防效果,具有较高的医疗和保健作用[4-8]。已有较多研究成果表明松科植物树皮与松针中黄酮类化合物含量丰富,具有较好的开发利用潜力[9-10]。国内学者如李石清等对南方红豆杉(Taxus chinensis var. mairei)枝叶总黄酮的研究、冯慧英等对巴山冷杉(Abies fargesii)总黄酮的研究、王飞等对青扦(Picea wilsonii)针叶总黄酮的研究等均表明其易提取且具有较强的抗氧化能力[11-13]。
担子菌门(Basidiomycota)的许多菌类代谢产物已经被开发利用,关于担子菌的研究已经成为当代生命科学、食品科学和医药领域的热点[14-15]。锈菌目(Uredinales)隶属于担子菌门,目前却尚未发现任何关于锈菌目真菌开发利用的研究,这也表明我们对于锈菌目真菌的潜在价值知之甚少。本研究选择了锈菌目中寄生在松科植物上的易于采集的松栎柱锈菌春孢子为研究对象,进行了松栎柱锈菌春孢子黄酮类化合物(以下简称CQF)的提取与体外抗氧化活性的研究,旨在为锈菌目菌物开发利用、森林有害生物开发利用和提升与拓宽森林有害生物防治理念提供科学依据。
2. 材料与方法
2.1 研究材料与预处理
松栎柱锈菌春孢子于2015年5月中旬采自大兴安岭地区,过80目筛后于45℃烘干至恒质量,密封冷藏。所用试剂盒均购自南京建成生物研究所;芦丁标准品,购自上海源叶生物科技有限公司;其他试剂均为国产分析纯;KQ-600B型超声波仪,昆山市超声波仪器有限公司;分析天平(感应量0.0001g);Gene Quant 1300型分光光度计,美国GE公司。
2.2 测定波长的选择与标准曲线的建立
精密称取芦丁标准品10.0mg,使用体积分数为75%的乙醇溶液配制成50mL 0.2mg/mL的标准溶液。取上述标准溶液稀释为质量浓度为0.04,0.06,0.08,0.10,0.12,0.14,0.16,0.18,0.20mg/mL的芦丁溶液,分别吸取5mL上述各质量浓度芦丁溶液,置于1,2,3,4,5,6,7,8,9号共9支25mL容量瓶中,分别加入5%亚硝酸钠(NaNO2)溶液1.0mL,摇匀后室温下静置6min,再分别加入10%硝酸铝(Al(NO3)3)溶液1.0mL,摇匀后室温下静置6min,再加入1mol/L的氢氧化钠(NaOH)溶液8mL,用75%乙醇溶液定容至25mL。显色15min后取5号容量瓶进行紫外全波长扫描,以确定吸收峰。用蒸馏水调零后在300~900nm范围内的扫描,截取其在400~650nm范围内的吸光度变化示意图,结果如图 1所示。可知经过本方法处理过的黄酮类化合物在500~520nm处出现了明显的吸收峰,因此判断本方法能够用于CQF的测定。为减小红移和蓝移对检测结果的影响,选择510nm作为测定波长。
不同质量浓度芦丁标准品溶液按照本方法操作后,在510nm处测定各质量浓度对应的吸光度A,以芦丁标准品质量浓度(mg/mL)为横坐标,吸光度A值为纵坐标,绘制标准曲线,结果如图 2所示。标准曲线的回归方程为y=2.5617x-0.001,回归方程的相关系数R2=0.9995,说明该检测方法可行,所得回归方程能够用来测定CQF质量浓度。
2.3 单因素试验
设定松栎柱锈菌春孢子黄酮类化合物的初始提取条件为:提取温度A=40℃、料液比B=1:40、提取时间C=1h、乙醇体积分数D=50%(v/v);每次试验所用春孢子粉200mg;超声频率40kHz,超声功率600W。将所得提取液5000r/min离心8min,吸取上层清液按照2.2的方法进行测定样品吸光值A。根据2.2中所得芦丁标准曲线:y=2.5617x-0.001推算CQF提取液中CQF的质量浓度,并通过式(1)计算CQF的得率。采用控制变量法依次考察该提取条件下上述4个单因子对CQF得率的影响。
T=NnVm×100% (1) 式中:T为CQF得率,%;N为根据标准曲线计算出的CQF的质量浓度,mg/mL;n为稀释倍数;V为提取液的体积,mL;m为供试春孢子质量,mg。
2.4 得率建模试验
在单因子考察试验基础上,根据中心组合实验(central composite design)原理使用软件Design-Expert 8.0.6对提取温度(A),料液比(B),提取时间(C),乙醇体积分数(D)这4个因子较优的水平进行了CQF得率的响应面建模试验。完成建模试验后使用建模试验数据建立CQF得率关于以上4个影响因素的二次多项式回归方程数学模型,对所得模型及各个影响因子进行方差分析。通过软件Design-Expert 8.0.6与所得数学模型得出最优CQF提取条件,并验证优化结果。建模试验各因素水平与实际值对应关系见表 1。
表 1 建模试验因子水平表Table 1. Factors and levels of flavonoid extraction experiment水平
Level影响因子Influencing factor 温度Temperature(A)/℃ 料液比Solid-liquid ratio(B)/(g·mL-1) 时间Time(C)/min 乙醇体积分数
Ethanol contentration(D)/%-2 35 1:100 20 50 -1 45 1:120 30 60 0 55 1:140 40 70 1 65 1:160 50 80 2 75 1:180 60 90 2.5 CQF抗氧化能力测定
除CQF的还原力与DPPH·清除能力之外,清除超氧阴离子(O2-·)活性、清除羟基自由基(·OH)活性和总抗氧化活性均采用从南京建成生物研究所购买的对应的试剂盒测定,并严格按照试剂盒使用说明执行。本文所有CQF体外抗氧化活性测定实验均以抗坏血酸(Vc)做阳性对照,用以评价CQF的体外抗氧化活性。
2.5.1 CQF还原力测定
用体积分数为50%的乙醇溶液将在建模试验中所收集的CQF溶液稀释为原来质量浓度的0.2、0.4、0.6、0.8和1.0倍,分别向装有1.0mL各质量浓度CQF溶液的试管中加入0.2mol/L的pH 6.6的磷酸缓冲溶液2.5mL,快速混匀后加入质量分数1%的铁氰化钾溶液2.5mL并在50℃下水浴20min。迅速冷却反应液并加入质量分数10%的三氯乙酸溶液2.5mL,充分混匀,3000r/min离心10min后取上清液3mL,依次加入3mL蒸馏水和1.0mL的0.1%的三氯化铁溶液,充分混匀后静置10min,于700nm波长下测定溶液吸光度。吸光度越大还原能力也越大,计算公式如(2)。
A700=A样品−A空白 (2) 2.5.2 DPPH·清除能力测定
用95%乙醇溶液配置DPPH·浓度为0.2mmol/L的乙醇溶液,置于冰箱中避光冷藏备用。测定方法:测定管用2mL DPPH·乙醇溶液中加入2mL各质量浓度的CQF样品溶液,CQF质量浓度设置梯度与2.5.1相同。对照管用95%乙醇代替DPPH·溶液;空白管用蒸馏水代替CQF溶液。混匀器混匀后避光反应30min。波长调至517nm处用0.5mL蒸馏水和2.5mL 95%的乙醇调零,然后测定反应液吸光值A,每个质量浓度设置3个平行实验,结果取平均值。DPPH·自由基的清除率通过公式(3)计算得出。
DPPH⋅清除率=A0−(A1−A2)A0×100% (3) 式中:A0为2mL DPPH·乙醇溶液+2mL超纯水的吸光值;A1为2mL DPPH·乙醇溶液+2mL样品溶液后的吸光值;A2为2mL样品溶液+2mL超纯水的吸光值。
3. 结果与分析
3.1 单因子试验
3.1.1 乙醇体积分数对CQF得率的影响
由图 3可知,在其他影响因子不变的条件下,乙醇体积分数在50%~90%范围内变化导致CQF的得率先逐渐增大,当乙醇体积分数增至70%的时候,得率最大,但是仍然不足0.5%。当乙醇体积分数超过70%以后,随着体积分数的增加导致CQF得率骤降并逐渐趋于稳定。黄酮类化合物难溶于水,易溶于有机溶剂,水和乙醇可以以任意比例互溶。在水中加入不同量的乙醇,溶液的极性也随之不同,因而对某种或某类物质的溶解能力也不相同。根据所得实验结果绘制的折线图可以看出乙醇体积分数为70%是较优水平。
![]() 图 3 乙醇体积分数对黄酮类化合物(CQF)得率的影响Figure 3. Effects of ethanol concentration on extraction rate of flavonoids
图 3 乙醇体积分数对黄酮类化合物(CQF)得率的影响Figure 3. Effects of ethanol concentration on extraction rate of flavonoids3.1.2 料液比对CQF得率的影响
根据图 4可以看出在其他条件不变的情况下,随着料液比的减小,CQF得率在不断升高,并逐渐趋于平稳,在料液比达到1:140以后,CQF得率增长幅度变小,得率增幅减小,因此选择1:140为较优水平。这一结果与CQF在50%乙醇溶液中的溶解度相关,当料液比较大时,CQF只有少部分溶于了乙醇溶剂,此时的提取液处于一种近饱和状态;随着提取液的增多,也就是料液比减小,未溶出的CQF部分也逐渐溶出,导致CQF的得率上升。当料液比减小到一定程度时,能溶于50%乙醇溶液的部分溶解殆尽,所以这时的得率逐渐趋于稳定,靠近极值。
![]() 图 4 料液比对CQF得率的影响Figure 4. Effects of solid-liquid ratio on extraction rate of flavonoids
图 4 料液比对CQF得率的影响Figure 4. Effects of solid-liquid ratio on extraction rate of flavonoids3.1.3 提取温度对CQF得率的影响图
在其他试验条件不变的条件下,考察了温度在15~75℃之间变化对CQF得率的影响,通过考察发现CQF得率先随着温度升高而增大,随着温度升高至65℃以后又逐渐降低。当提取液温度较低时,分子获得的能量相对较少,分子运动缓慢,有效提取CQF耗费的时间增长,这一点可以通过分子扩散现象很好地解释。但是,温度的变化同样会影响溶剂分子之间的作用力,从而影响溶剂溶解溶质的能力,导致CQF得率也随之发生变化。也就是说温度的变化同时影响着CQF分子运动速率与溶剂,因而CQF得率受此二者的交互作用影响,CQF得率随提取温度的变化如图 5所示。
![]() 图 5 提取温度对CQF提取率的影响Figure 5. Effects of extraction temperature on extraction rate of flavonoids
图 5 提取温度对CQF提取率的影响Figure 5. Effects of extraction temperature on extraction rate of flavonoids3.1.4 提取时间对CQF得率的影响
在其他试验因子不变的条件下,考察提取时间对CQF得率的影响,结果显示提取时间对CQF得率的影响微弱,时间在20~60min变化时,CQF得率的变化始终处在0.46%~0.468%之间。CQF分子溶解到提取液中需要一定的时间,所以在提取时间40min以内时,CQF得率随着时间增加而增高,随着提取时间的延长,CQF的得率反而逐渐开始降低。这或许是因为部分原生质体内的CQF分子进入了细胞壁,因为细胞壁本身就可以容纳黄酮类化合物,另一方面,松栎柱锈菌春孢子表面具有油球,对黄酮类化合物具有一定的溶解能力。除此之外,随着溶剂分子运动时间的增加,乙醇分子扩散增多引起溶剂中乙醇浓度降低,从而导致CQF提取率也降低。CQF得率随提取时间的变化如图 6所示。
![]() 图 6 提取时间对CQF提取率的影响Figure 6. Effects of extraction time on extraction rate of flavonoids
图 6 提取时间对CQF提取率的影响Figure 6. Effects of extraction time on extraction rate of flavonoids3.2 响应面法分析
3.2.1 建模试验设计及结果
使用软件Design-Expert 8.0.6对提取温度(A),料液比(B),提取时间(C),乙醇体积分数(D)这4个因子的较优水平进行CQF得率的响应面建模试验。试验因子水平设计如表 1所示,中心组合实验设计与结果见表 2。
表 2 中心组合实验设计与结果Table 2. Design and results of central composite design试验号
Run No.温度
Temperature(A)/℃料液比
Solid-liquid ratio(B)/(g·mL-1)时间
Time(C)/min乙醇体积分数
Ethanol concentration(D)/%得率
Yield(T)/%1 0 -2 0 0 1.6329 2 1 -1 -1 1 1.0071 3 -1 1 1 1 0.9619 4 1 1 1 -1 1.4116 5 2 0 0 0 1.2952 6 0 0 0 0 1.1149 7 -1 1 1 -1 1.1555 8 0 0 0 0 1.1586 9 1 -1 1 1 1.1196 10 0 0 0 0 1.0930 11 1 1 -1 -1 1.3928 12 0 0 0 2 0.7542 13 0 0 0 0 1.1477 14 -1 -1 1 1 0.9697 15 1 -1 -1 -1 1.2507 16 -1 -1 -1 -1 1.1524 17 -1 -1 -1 1 0.8713 18 1 1 1 1 1.1805 19 -1 1 -1 1 0.9993 20 0 2 0 0 1.1453 21 -1 -1 1 -1 1.1055 22 0 0 -2 0 1.2133 23 1 -1 1 -1 1.3959 24 0 0 2 0 0.1969 25 1 1 -1 1 1.0555 26 0 0 0 -2 1.1040 27 0 0 0 0 1.1258 28 -2 0 0 0 1.3675 29 0 0 0 0 1.1477 30 -1 1 -1 -1 1.2304 3.2.2 数学模型的建立与分析
以CQF的得率为响应值,通过使用软件Design-Expert 8.0.6对得率T与4个影响因子回归拟合之后,得到CQF关于4个影响因子的数学模型方程:
T=1.13+0.051A−0.019B−0.071C−0.11D+1.171e−3AB+0.029AC−0.015AD−0.017BC−3.513e−3BD+0.016CD+0.059A2+0.073B2−0.098C2−0.042D2 对该数学模型进行方差分析(表 3)可知该模型回归显著(P<0.05),失拟项不显著(P=0.149),说明未知因素对CQF得率影响较小,模型拟合效果良好。在我们设置的4因子水平梯度范围内,乙醇体积分数D对于CQF得率的影响显著(P<0.05),这与黄酮类化合物易溶于有机溶剂难溶于水的化学性质密不可分。由F值可得出各个影响因子对CQF得率的影响程度大小顺序为:乙醇体积分数(D)>提取时间(C)>提取温度(A)>料液比(B)。
表 3 数学模型方差分析Table 3. Parameter estimate of regression model and variance analysis方差来源
Variance source平方和Sum
f squares自由度
df均方
Mean squareF值
F valueP值
P value显著性
Significance模型Model 1.150 14 0.082 7.469 0.043 1 显著Significant 温度Temperature(A) 0.062 1 0.062 1.520 0.237 1 料液比Solid-liquid ratio(B) 0.009 1 0.009 0.210 0.650 1 时间Time(C) 0.120 1 0.120 2.900 0.109 1 乙醇体积分数Ethanol concentration(D) 0.290 1 0.290 7.010 0.018 3 AB 0.000 1 0.000 0.001 0.981 9 AC 0.013 1 0.013 0.320 0.577 2 AD 0.004 1 0.004 0.093 0.765 2 BC 0.005 1 0.005 0.120 0.736 6 BD 0.000 1 0.000 0.005 0.945 7 CD 0.004 1 0.004 0.100 0.756 6 A2 0.095 1 0.095 2.300 0.149 8 B2 0.150 1 0.150 3.580 0.078 1 C2 0.260 1 0.260 6.380 0.023 3 D2 0.048 1 0.048 1.160 0.297 5 残差Residual 0.620 15 0.041 失拟项Lack of fit 0.610 10 0.061 0.017 0.149 不显著Not significant 纯误差Pure error 0.003 5 0.001 总误差Total error 1.770 29 3.2.3 交互项对CQF得率的影响
响应面是以响应值与影响响应值的因子构建起了空间三维图形,可以让我们更直观的判断出两两不同影响因子交互作用对响应值的影响趋势以及响应值的变化范围。响应面图中颜色的变化表示响应值(CQF得率)的变化,颜色变化快,响应值变化也快,一般较低的响应值使用蓝色表示,较高的响应值用红色表示,其他颜色均为中间过渡颜色。同时,在一定方向和范围内响应面3D图形中坡度越大表示响应值变化的越快,即对CQF得率的影响更为显著。这一点在等高线图形中也有很好的体现,当某条线段在垂直其中一条轴的方向上移动时,在一定范围内与它所相交的等高线越多,则说明在该范围内某一因子对响应值的影响越显著。等高线表示在此条线上所有试验方案都能够得到相同的响应值,即虽然等高线所对应的试验条件不同,但是能够产生相同的实验结果。4个影响因子的两两之间交互作用对CQF得率的影响示意图见图 7。
![]() 图 7 各因素交互作用对CQF得率影响的响应面与等高线图Figure 7. Response surface and contour plots for the interaction effects of various factors on extraction rate of CQF
图 7 各因素交互作用对CQF得率影响的响应面与等高线图Figure 7. Response surface and contour plots for the interaction effects of various factors on extraction rate of CQF3.2.4 最佳提取工艺及验证
根据在3.2.2中对模型进行的方差分析,已经得知我们拟合出的回归方程模型显著,失拟项不显著,说明该方程是适合用于CQF得率与提取工艺的数学模型,所以我们可以借助于该方程确定最佳的CQF提取工艺。经软件Design-Expert 8.0.6分析,得知模型预测的最佳提取工艺为:提取温度65℃,料液比1:120,提取时间38min,提取液乙醇体积分数60%。在此条件下有对该优化结果进行了3次验证实验,3次验证实验所得出的CQF得率为1.382%,实际得率与在该条件下模型所预测的得率1.416%相差2.46%,这表明我们优化出来的CQF提取条件真实可靠,能够应用于实践。
3.3 抗氧化活性分析
3.3.1 CQF的还原力测定
还原力是评价物质抗氧化能力的重要指标之一,其原理是具有较强还原力的物质将Fe3+还原为Fe2+,Fe2+在酸性环境中能与FeCl3产生有颜色的普鲁士蓝。在该检测体系下物质的还原力越强,对应的吸光值也越大。根据图 8可以看出,CQF与Vc的还原力与质量浓度均呈现很好的线性关系,拟合度均超过0.998。在图示浓度范围内,CQF和Vc的质量浓度与还原力均表现出较好的量效关系,相同质量浓度条件下CQF的还原力更强,而且CQF质量浓度与还原力的线性方程斜率更大,量效关系更为明显。在图示范围内,不论是量效关系还是还原力,CQF都强于Vc,说明CQF具备很好的还原力。
3.3.2 CQF对DPPH·的清除能力
在图 9中,根据受试物质量浓度与DPPH·清除率的散点图,我们可以看出Vc对DPPH·的清除能力较CQF强,并迅速逼近清除率最大值。在散点图的基础上,可以得出CQF与Vc清除DPPH·的IC50分别为0.0492mg/mL和0.0335mg/mL,也就是说对CQF对DPPH·清除能力较Vc差。但是随着CQF质量浓度的逐步增大,CQF对DPPH·清除能力也逐步增强,量效关系越来越明显。对于该种人工合成自由基,CQF的反应不够灵敏。
![]() 图 9 受试物清除DPPH·能力的比较Figure 9. Comparison of DPPH· radical scavenging capacity of CQF and Vc
图 9 受试物清除DPPH·能力的比较Figure 9. Comparison of DPPH· radical scavenging capacity of CQF and Vc3.3.3 CQF总抗氧化能力
生物活性物质总抗氧化能力是指活性物质清除各种自由基的有效总和。虽然已有多种测定抗氧化活性的方法,但这些方法只是从某个角度来衡量天然抗氧化物的活性。事实上,抗氧化作用涉及到多种作用机制,它涉及一系列的问题,目前尚未形成标准方法[16]。本实验测定总抗氧化能力采用的南京建成生物研究所试剂盒原理为FRAP法,在该种检测方法下,由图 10可以看出CQF与Vc的总抗氧化能力相当。根据散点图经计算得知,CQF的半抑制率IC50为0.0303mg/mL,而Vc的半抑制率为0.0240mg/mL,可以看出Vc的总抗氧化能力较CQF强,但是CQF与Vc趋势线在0.0654mg/mL处相交。整体来看,两种物质达到最大清除率都不需要较大浓度,量效关系相近,其总抗氧化能力并不具有很大的差异。所以说,CQF的总抗氧化能力还是非常理想的。
3.3.4 CQF对·OH的清除能力
羟基自由基(·OH)属于一种强氧化剂(E0=2.80V,3.06V),氧化能力远大于一般氧化剂,是机体中最活跃的一种活性氧分子,具有很强的破坏性。它几乎可以和活细胞中的任何生物分子发生反应而造成损伤,而且反应速度极快[17]。因此,清除机体内过多的·OH具有非常重要的生物学意义。通过与Vc的对比测试,我们可以看出Vc在质量浓度0.016 ~0.080mg/mL范围内呈现线性变化规律,但对·OH较强的清除能力远低于CQF。由图 11可以非常明显的看出Vc与CQF在·OH清除能力上的差距。虽然Vc具有很强的还原性,不仅能够直接清除·OH,还能够阻断Fenton反应,从而减少体系中的·OH,但是CQF对·OH的敏感程度与清除能力依然远胜Vc。经测定,CQF对·OH清除能力的IC50仅为0.0129mg/mL,因为Vc对·OH的清除能力远小于CQF,故未予测定与计算其对·OH的半抑制率。
3.3.5 CQF对O2-·的清除能力
超氧阴离子O2-·经过一系列反应可以生成其他氧自由基,是大多数氧自由基的母体,能够攻击细胞DNA,具有较强的毒性,所以清除生物机体内过多的O2-·具有重要意义。理论上来讲,CQF能够提供电子从而有效清除自由基,但本课题在测定CQF清除O2-·能力时却发现了截然相反的实验结果。由图 12可知,两种受试物对清除O2-·都具有一定的量效关系,但是测定结果不在同一象限内。这也就是说在该中检测体系下受试物CQF非但没有清除O2-·,反而促进了O2-·的产生。所使用试剂盒检测原理为模拟机体中黄嘌呤与黄嘌呤氧化酶反应系统,黄嘌呤在黄嘌呤酶的催化下转化为尿酸,在此过程中能够产生O2-·,然后通过衡量O2-·的量来推算加入反应体系的物质时清除还是产生O2-·。黄酮类化合物作为电子提供物质,不会产生O2-·,但是CQF提取液是未经过除杂的,一些会对该反应体系产生影响的物质可能一并被提取了,比如说嘌呤或黄嘌呤氧化酶以及一些自氧化物质,这些物质生成O2-·的能力远超黄酮类物质清除O2-·的能力,所以才产生了如图 12所示的测定结果[18-20]。这一问题没有得到有效解决,所以尚不清楚导致该检测结果的具体原因,需要在后续工作中进一步研究导致该结果的具体原因与解决方法。黄嘌呤酶氧化法目前是测定O2-·最完善最普遍的方法,但是这种方法应该对待测样品设置一定的要求或合适的处理条件以排除干扰因子,需要我们进一步完善这种测定方法。
4. 结论与讨论
从微生物孢子中提取黄酮类化合物的例子尚不多见,在此背景下,本研究采用控制变量法逐个考察了能够影响CQF得率的4个因子。然后使用软件Design-Expert 8.0.6设计了建模试验,完成建模试验之后利用可靠的CQF得率数学模型得到了适用的最优CQF提取条件:提取温度65℃,料液比1:120,提取时间38min,提取液乙醇体积分数60%,在该提取条件下CQF得率为1.382%。虽然松栎柱锈菌春孢子具有厚且坚硬的几丁质细胞壁,能够有效阻止细胞内含物的流失以抵抗不良环境因素,但是我们使用一般植物总黄酮提取的方法并辅以超声振荡也提取到了CQF,说明醇提取法同样能够用于其他孢子总黄酮的提取并且达到了预期的CQF得率。总黄酮得率既取决于提取方法,也取决于实验材料总黄酮含量。一般来说针叶树实验材料总黄酮得率高于菌物但不足5%。闫琳娜通过对落叶松(Larix gmelinii)松针黄酮的研究发现落叶松松针总黄酮含量较高且在不同提取条件下得率都能达到6%以上[21];孙琼优化了在菌物中黄酮类化合物含量相对较高的杏鲍菇(Pleurotus eryngii)黄酮类化合物的提取条件,但是得率仍然不足3%[22]。同时我们也看到了黄嘌呤酶氧化法在实际应用中暴露出来的问题,没能有效测定出CQF清除O2-·的能力,需要进一步明确并排除干扰因子或者改进O2-·检测方法。
黄晓冬等对阔叶树种白桦(Betula platyphylla)活体寄生真菌桦褐孔菌(Inonotus obliquus)总黄酮的研究表明其具有较强的体内外抗氧化活性[23],Boonsong等研究了5种食用菌总酚与类黄酮的提取与抗氧化活性,发现乙醇溶液更适合菌物总酚的提取,其提取物均表现出较强的抗氧化活性,然而很多的食用菌不是活体寄生或不含有黄酮类物质[24]。整体来看,对于针叶树总黄酮的研究成果与热情远胜担子菌,而活体寄生的担子菌总黄酮的研究很鲜见。本研究所选研究对象将针叶树与担子菌门菌物总黄酮的研究很好地有机结合,旨在为锈菌目菌物开发利用、森林有害生物开发利用和转变森林有害生物防治理念提供科学依据。迄今为止,人类对锈菌的认知还很不完善,尚未看到国内外任何关于锈菌开发利用的报道。本研究首次完成了CQF的提取与体外抗氧化活性测定工作,发现CQF具有很强的还原力,还原力较Vc强;其次CQF的总抗氧化能力和DPPH·清除能力与Vc相当;CQF对·OH具有非常好的清除作用,这一能力远胜Vc,与Treml等对黄酮类化合物清除·OH能力的研究现状综述相一致[26]。体外试验结果为体内试验提供了一定的科学依据,这表明CQF具有开发利用的潜在价值。
长久以来,人类对于有害生物的认识是不全面的,往往只是单纯的从治理的角度去对待,总是尝试使用各种各样的手段降低或控制有害生物的种群数量。森林有害生物确实为林业生产活动和森林健康产生了很多负面的影响,如果说发现并证实某种有害生物有其独特的利用价值,那么它的某种功能或价值就能在一定程度上抵偿它所造成的损失,从而引导我们转变传统的有害生物防治理念。
-
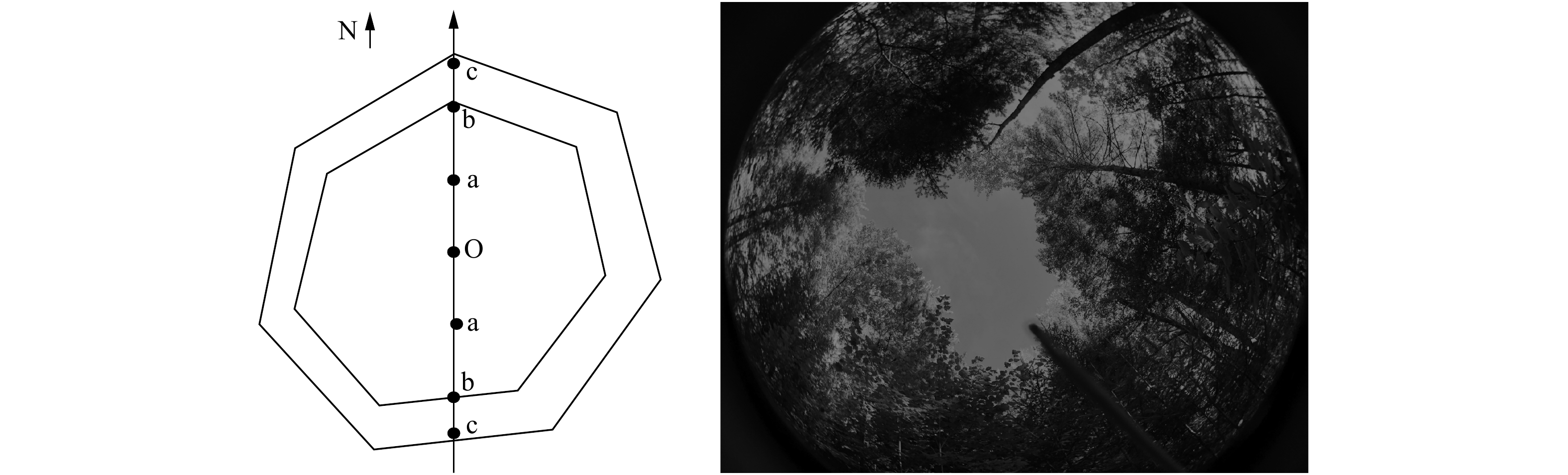
![]()
图 1 冠层结构和光因子数据采集
O. 林隙中心;a. 中心与冠缘连线中点;b. 林冠边缘;c. 林冠下。O represents gap center; a represents midpoint from the gap center to the canopy edge; b represents canopy edge; c represents under the canopy.
Figure 1. Data collection of canopy structure and light factors
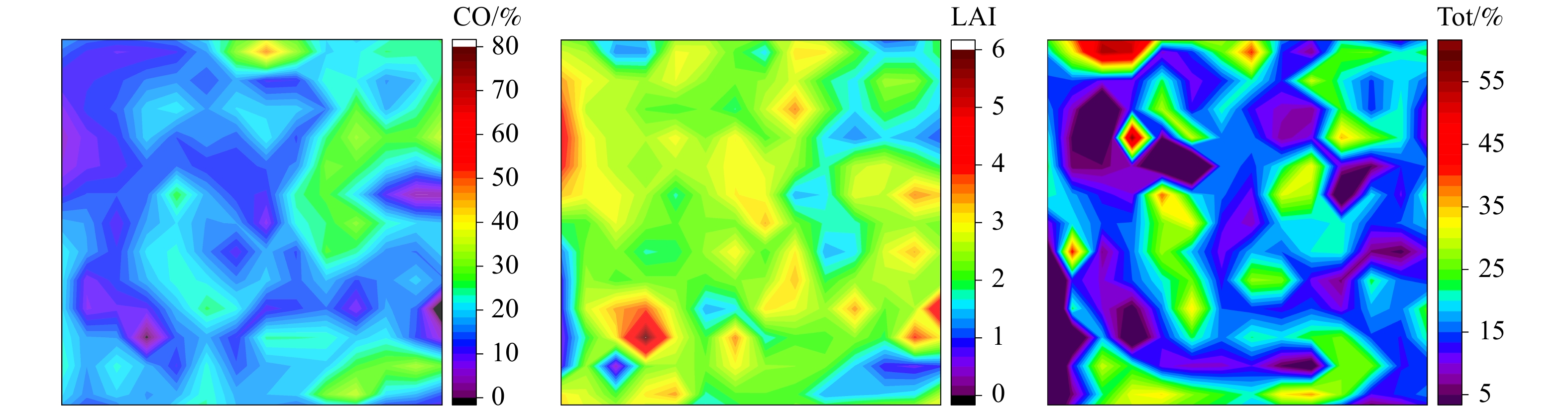
![]()
图 3 冠层因子样本率堆积图
CO(Ⅰ [8%, 12%)、Ⅱ [12%, 16%)、Ⅲ [16%, 20%)、Ⅳ [20%, 24%)、Ⅴ [24%, 28%)、Ⅵ [28%, 32%)、Ⅶ [32%, 36%));LAI(Ⅰ [1, 1.5)、Ⅱ [1.5, 2)、Ⅲ [2, 2.5)、Ⅳ [2.5, 3)、Ⅴ [3, 3.5)、Ⅵ [3.5, 4)、Ⅶ [4, 4.5));Tot(Ⅰ [6%, 11%)、Ⅱ [11%, 16%)、Ⅲ [16%, 21%)、Ⅳ [21%, 26%)、Ⅴ [26%, 31%)、Ⅵ [31%, 36%)、Ⅶ [36%, 41%));TDR(Ⅰ [0, 8%)、Ⅱ [8%, 16%)、Ⅲ [16%, 24%)、Ⅳ [24%, 32%)、Ⅴ [32%, 40%)、Ⅵ [40%, 48%)、Ⅶ [48%, 56%));TDF(Ⅰ [9%, 15%)、Ⅱ [15%, 21%)、Ⅲ [21%, 27%)、Ⅳ [27%, 33%)、Ⅴ [33%, 39%)、Ⅵ [39%, 45%)、Ⅶ [45%, 51%))
Figure 3. Sample rate accumulation diagram of canopy factors
![]()
图 4 冠层开度和总辐射对幼苗幼树株数的影响
Figure 4. Effects of CO and Tot on number of seedlings and saplings
表 1 林隙边缘木特征
Table 1 Characteristic of the trees in gap edge
树种 Tree species 数量 Quantity/% 平均胸径 Average DBH/cm 平均高 Average height/m 平均冠幅 Average crown width/m 冷杉 Abies nephrolepis 39.84 23.26 17.39 4.04 云杉 Picea koraiensis 5.42 25.17 16.58 4.59 红松 Pinus koraiensis 23.31 27.06 18.64 4.65 色木槭 Acer mono 7.32 18.97 17.29 3.84 白桦 Betula platyphylla 2.98 31.97 20.84 4.91 枫桦 Betula costata 0.27 5.40 7.80 2.18 紫椴 Tilia amurensis 20.86 13.70 12.60 3.47  下载: 导出CSV
下载: 导出CSV
表 2 林隙内主要幼苗幼树特征
Table 2 Characteristic of main seedlings and saplings in gaps
树种 Tree species 数量 Quantity/% 平均地径 Average ground diameter/mm 平均高 Average height /m 平均冠幅 Average crown width/m 冷杉 Abies nephrolepis 68.23 19.63 1.16 0.80 云杉 Picea koraiensis 9.40 17.59 1.26 0.36 红松 Pinus koraiensis 2.66 14.88 1.19 0.81 色木槭 Acer mono 19.71 8.35 0.64 0.58
下载: 导出CSV
表 3 冠层因子基本信息
Table 3 Basic information of canopy factors
冠层因子
Canopy factor最小值
Min. value最大值
Max. value平均值
Mean平均偏差
Average deviation标准偏差
Standard deviationCO/% 8.61 35.01 17.26 0.038 0 0.047 3 LAI 1.30 4.15 2.37 0.400 8 0.499 4 Tot/% 6.41 40.61 16.52 0.047 7 0.060 2 TDR/% 0.24 51.15 11.73 0.073 1 0.094 3 TDF/% 9.30 48.52 21.30 0.059 9 0.074 3 注:CO.冠层开度;LAI.叶面积指数;Tot.总辐射;TDR.直接辐射;TDF.散射辐射。下同。Notes: CO represents canopy openness; LAI represents leaf area index; Tot represents trans total radiation; TDR represents trans direct radiation; TDF represents trans diffuse radiation. The same below.
下载: 导出CSV
表 4 不同大小林隙内冠层因子变化
Table 4 Change of canopy factors in different size gaps
林隙面积 Gap size/m2 CO/% LAI Tot/% TDR/% TDF/% < 30 15.66 ± 0.39A 2.55 ± 0.05A 15.25 ± 0.62A 13.51 ± 1.23AC 17.00 ± 0.53A 30 ~ 60 15.86 ± 0.49AB 2.47 ± 0.06A 14.81 ± 0.59A 9.76 ± 0.84B 19.95 ± 0.71B 60 ~ 90 16.21 ± 0.70AB 2.45 ± 0.07AB 14.86 ± 0.77A 8.92 ± 1.29B 20.70 ± 1.04B 90 ~ 120 18.17 ± 0.73B 2.21 ± 0.06BC 18.35 ± 1.09B 10.65 ± 1.57ABC 24.13 ± 1.08C 120 ~ 150 21.60 ± 1.07C 1.92 ± 0.06CD 20.25 ± 1.03BC 12.58 ± 1.80ABC 29.86 ± 1.66D ≥ 150 22.71 ± 1.03C 1.90 ± 0.08D 22.76 ± 1.44C 15.05 ± 2.39C 30.47 ± 1.45D 注:表中数值表示平均值 ± 标准误差;不同大写字母表示不同林隙面积等级之间差异显著(P < 0.05)。Notes: values are presented as the mean ± standard error. Different uppercase letters indicate significant differences among gap size classes (P < 0.05).
下载: 导出CSV
表 5 光因子与冷杉和色木槭地径、树高及冠幅的相关性
Table 5 Correlations between light factors and ground diameter (DGH), tree height and crown width (CW) of Abies nephrolepis and Acer mono
林隙面积
Area of gap/m2光因子
Light factor/%冷杉地径
DGH of Abies nephrolepis冷杉树高
Tree height of Abies nephrolepis冷杉冠幅
CW of Abies nephrolepis色木槭地径
DGH of Acer mono色木槭树高
Tree height of Acer mono色木槭冠幅
CW of Acer mono< 30 TDR 0.231** 0.241** 0.135** 0.280** 0.425** 0.116 TDF 0.112* 0.141** 0.049 −0.177* −0.042 0.114 Tot 0.205** 0.234** 0.107* 0.040 0.204* 0.138 30 ~ 60 TDR −0.093* −0.106* −0.082 −0.005 −0.044 0.003 TDF 0.039 0.116* 0.110* −0.132* −0.116 −0.102 Tot 0.131 0.112 0.222 −0.091 −0.112 −0.081 60 ~ 90 TDR 0.249** 0.332** 0.178** 0.411** −0.037 −0.069 TDF −0.074 −0.027 −0.095* −0.240** 0.086 0.049 Tot 0.086* 0.184** 0.022 −0.131 0.099 0.058 90 ~ 120 TDR −0.111* −0.154** −0.098* −0.242* −0.145 −0.084 TDF 0.277** 0.299** 0.240** −0.152 −0.187 −0.272* Tot 0.021 0.076 0.043 −0.396** −0.301** −0.282* 120 ~ 150 TDR −0.088 −0.152** −0.082 −0.346** −0.482** −0.455** TDF −0.274** −0.380** −0.357** −0.317** −0.435** −0.407** Tot −0.180** −0.270** −0.213** −0.345** −0.478** −0.450** ≥ 150 TDR −0.302** −0.348** −0.236** −0.026 −0.227** −0.075 TDF −0.043 −0.088** −0.053 0.001 −0.068 0.010 Tot −0.308** −0.379** −0.252** −0.019 −0.230** −0.048 注:*代表在0.05水平下的差异显著,**代表在0.01水平下的差异显著。Notes: * represents significant difference at the 0.05 level, ** represents significant difference at the 0.01 level.
下载: 导出CSV
表 6 光因子与云杉和红松地径、树高及冠幅的相关性
Table 6 Correlations between light factors and DGH, tree height and CW of Picea koraiensis and Pinus koraiensis
林隙面积
Area of gap/m2光因子
Light factor/%云杉地径
DGH of Picea koraiensis云杉树高
Tree height of Picea koraiensis云杉冠幅
CW of Picea koraiensis红松地径
DGH of Pinus koraiensis红松树高
Tree height of Pinus koraiensis红松冠幅
CW of Pinus koraiensis< 30 TDR 0.382** 0.307* 0.068 −0.015 −0.080 0.047 TDF −0.079 −0.092 0.123 −0.248 −0.018 −0.157 Tot 0.164 0.115 0.129 −0.269 −0.038 −0.156 30 ~ 60 TDR −0.217 −0.279* −0.166 −0.359 −0.239 −0.406* TDF −0.183 −0.135 −0.054 0.260 0.158 0.146 Tot −0.222 −0.239* −0.124 0.005 −0.005 −0.131 60 ~ 90 TDR 0.086 0.268** 0.016 0.585** 0.587** 0.296 TDF −0.161 −0.006 −0.159 −0.115 0.007 0.063 Tot −0.115 0.151 −0.159 0.195 0.301 0.217 90 ~ 120 TDR −0.219 −0.248* −0.145 0.107 −0.128 −0.063 TDF 0.229* 0.232* 0.124 −0.034 0.066 0.001 Tot −0.132 −0.164 −0.102 0.113 −0.117 −0.081 120 ~ 150 TDR 0.094 −0.142 0.131 −0.281 −0.280 0.051 TDF −0.29 −0.526** −0.338 −0.449 −0.387 −0.052 Tot −0.044 −0.298 −0.036 −0.359 −0.333 0.010 ≥ 150 TDR −0.273** −0.325** −0.211* −0.554** −0.532** −0.458** TDF −0.155 −0.227* −0.255** 0.678** 0.646** 0.577** Tot −0.325** −0.416** −0.341** 0.239 0.224 0.217
下载: 导出CSV
-
[1] Watt A S. Pattern and process in the plant community[J]. Journal of Ecology, 1947, 35(1/2): 1−22. doi: 10.2307/2256497
[2] 郑云峰, 尹准生, 唐孝甲. 树种天然更新影响因素研究[J]. 华东森林经理, 2020, 34(2):1−4. doi: 10.3969/j.issn.1004-7743.2020.02.001 Zheng Y F, Yin Z S, Tang X J. Study on influencing factors of natural regeneration of tree species[J]. East China Forest Management, 2020, 34(2): 1−4. doi: 10.3969/j.issn.1004-7743.2020.02.001
[3] He Z S, Wang L J, Jiang L. Effect of microenvironment on species distribution patterns in the regeneration layer of forest gaps and non-gaps in a subtropical natural forest, China[J]. Forests, 2019, 10(2): 90−102. doi: 10.3390/f10020090
[4] Zhang T, Yan Q L, Wang J. Restoring temperate secondary forests by promoting sprout regeneration: effects of gap size and within-gap position on the photosynthesis and growth of stump sprouts with contrasting shade tolerance[J]. Forest Ecology and Management, 2018, 429: 267−277. doi: 10.1016/j.foreco.2018.07.025
[5] Moghimian N, Habashi H, Kooch Y. Influence of wind throw events on soil carbon sequestration and fertility status at local scales: a case study in Hyrcanian forest[J]. European Journal of Experimental Biology, 2013, 3: 160−167.
[6] Vitousek P M, Denslow J S. Nitrogen and phosphorus availability in treefall gaps of a lowland tropical rainforest[J]. Journal of Ecology, 1986, 74(4): 1167−1178. doi: 10.2307/2260241
[7] Kostrakiewicz G K. The impact of time of gap origin on microsite conditions and seedling recruitment in Molinietum caeruleae meadows[J]. International Journal of Conservation Science, 2015, 6(1): 111−124.
[8] Meer P J V D, Dignan P, Saveneh A G. Effect of gap size on seedling establishment, growth and survival at three years in mountain ash (Eucalyptus regnans F. Muell.) forest in Victoria, Australia[J]. Forest Ecology and Management, 1999, 117(1/3): 33−42.
[9] Zhu J J, Matsuzaki T, Lee F Q, et al. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest[J]. Forest Ecology and Management, 2003, 182(1): 339−354.
[10] Wang Z B, Yang H J, Dong B Q, et al. Effects of canopy gap size on growth and spatial patterns of Chinese pine (Pinus tabulaeformis) regeneration[J]. Forest Ecology and Management, 2017, 385: 46−56. doi: 10.1016/j.foreco.2016.11.022
[11] 胡振宇, 孙楠, 梁晓东. 长白落叶松人工林不同林隙间伐对林下更新生长的影响[J]. 林业科技, 2016, 41(1):11−13. doi: 10.3969/j.issn.1001-9499.2016.01.004 Hu Z Y, Sun N, Liang X D. Effects of different gap thinning on undergrowth of larch plantation in Changbai Mountain[J]. Forestry Science and Technology, 2016, 41(1): 11−13. doi: 10.3969/j.issn.1001-9499.2016.01.004
[12] Huth F, Wagner S. Gap structure and establishment of silver birch regeneration (Betula pendula Roth.) in Norway spruce stands (Picea abies L. Karst.)[J]. Forest Ecology and Management, 2006, 229(1/2/3): 314−324.
[13] 李谭宝, 李淑静, 王彩云. 黄龙山白皮松林林隙物种多样性动态[J]. 西北林学院学报, 2015, 30(4):66−72. doi: 10.3969/j.issn.1001-7461.2015.04.11 Li T B, Li S J, Wang C Y. Gap dynamics of species diversity in Pinus bungeana forest in Huanglong Mountain[J]. Journal of Northwest Forestry University, 2015, 30(4): 66−72. doi: 10.3969/j.issn.1001-7461.2015.04.11
[14] 王永强, 蔡燕茹, 曾焕忱, 等. 不同林冠开度下亚热带林下植物的组成和多样性[J]. 西北农林科技大学学报(自然科学版), 2016, 44(5):64−72. Wang Y Q, Cai Y R, Zeng H C, et al. Composition and diversity of understory plant species in subtropical forests under different canopy openness[J]. Journal of Northwest A&F University (Natural Science Edition), 2016, 44(5): 64−72.
[15] Zerbe S. Restoration of natural broad-leaved woodland in Central Europe on sites with coniferous forest plantations[J]. Forest Ecology and Management, 2002, 167(1): 27−42.
[16] 陈圣宾, 宋爱琴, 李振基. 森林幼苗更新对光环境异质性的响应研究进展[J]. 应用生态学报, 2005, 16(2):365−370. doi: 10.3321/j.issn:1001-9332.2005.02.034 Chen S B, Song A Q, Li Z J. Research advance in response of forest seedling regeneration to light environmental heterogeneity[J]. Chinese Journal of Applied Ecology, 2005, 16(2): 365−370. doi: 10.3321/j.issn:1001-9332.2005.02.034
[17] Runkle J R. Patterns of disturbance in some old-growth mesic forests of eastern North America[J]. Ecology, 1983, 64(4): 623.
[18] 罗桂生, 马履一, 贾忠奎, 等. 油松人工林林隙天然更新及与环境相关性分析[J]. 北京林业大学学报, 2019, 41(9):59−68. Luo G S, Ma L Y, Jia Z K, et al. Correlation analysis between natural regeneration and environment in canopy gap of Chinese pine (Pinus tabuliformis) plantation[J]. Journal of Beijing Forestry University, 2019, 41(9): 59−68.
[19] 蔡杨新, 黄梅珍, 许鲁东, 等. 闽粤栲天然林种群数量与结构的林隙边缘效应[J]. 四川农业大学学报, 2017, 35(1):31−36. Cai Y X, Huang M Z, Xu L D, et al. Gap edge effects on population size and structure in Castanopis fissa natural forest[J]. Journal of Sichuan Agricultural University, 2017, 35(1): 31−36.
[20] Macfarlane C. Classification method of mixed pixels does not affect canopy metrics from digital images of forest overstorey[J]. Agricultural and Forest Meteorology, 2011, 151(7): 833−840. doi: 10.1016/j.agrformet.2011.01.019
[21] 刘志理, 金光泽. 小兴安岭白桦次生林叶面积指数的估测[J]. 生态学报, 2013, 33(8):2505−2513. doi: 10.5846/stxb201201120065 Liu Z L, Jin G Z. Estimation of leaf area index of secondary Betula platyphylla forest in Xiaoxing’ an Mountains[J]. Acta Ecology Sinica, 2013, 33(8): 2505−2513. doi: 10.5846/stxb201201120065
[22] 马泽清, 刘琪璟, 曾慧卿, 等. 南方人工林叶面积指数的摄影测量[J]. 生态学报, 2008, 28(5):1971−1980. doi: 10.3321/j.issn:1000-0933.2008.05.011 Ma Z Q, Liu Q J, Zeng H Q, et al. Estimation of leaf area index of planted forests in subtropical China by photogrammetry[J]. Acta Ecology Sinica, 2008, 28(5): 1971−1980. doi: 10.3321/j.issn:1000-0933.2008.05.011
[23] Gardiner E S, Hodges J D. Growth and biomass distribution of cherry bark oak (Quercus pagoda Raf.) seedlings as influenced by light availability[J]. Forest Ecology and Management, 1998, 108: 127−134. doi: 10.1016/S0378-1127(98)00220-5
[24] 班宏娜. 樟子松人工林树冠层光分布规律及对生长影响的研究[D]. 哈尔滨: 东北林业大学, 2010. Ban H N. The research on canopy light distribution law and growth in fluence on Pinus sylverstris artificial stand [D]. Harbin: Northeast Forestry University, 2010.
[25] 方怡然, 潘澜, 薛立. 冰雪灾害后的杉木人工林冠层结构与林下光照及土壤生化特性的关系[J]. 生态环境学报, 2018, 27(4):609−616. Fang Y R, Pan L, Xue L. Relationships between canopy structure and understory light and soil biochemical property in a Cunninghamia lanceolata stand suffering from ice-snow damage[J]. Ecology and Environmental Sciences, 2018, 27(4): 609−616.
[26] 刘少冲, 段文标, 陈立新. 小兴安岭阔叶红松林不同大小林隙光照时空分布特征[J]. 东北林业大学学报, 2014, 42(8):46−51. doi: 10.3969/j.issn.1000-5382.2014.08.010 Liu S C, Duan W B, Chen L X. Spatiotemporal distribution characteristics of light in different size gaps in the mixed broad-leaved Korean pine forest in Xiaoxing’an Mountains[J]. Journal of Northeast Forestry University, 2014, 42(8): 46−51. doi: 10.3969/j.issn.1000-5382.2014.08.010
[27] Grubb P J, Bellingham P J, Kohyama T S, et al. Disturbance regimes, gap-demanding trees and seed mass related to tree height in warm temperate rain forests worldwide[J]. Biological Reviews, 2013, 88(3): 701−744. doi: 10.1111/brv.12029
[28] 区余端, 苏志尧. 粤北山地常绿阔叶林自然干扰后冠层结构空间异质性动态[J]. 植物科学学报, 2012, 30(3):223−229. Qu Y D, Su Z Y. Spatial heterogeneity dynamics of canopy structure in a montane evergreen broadleaved forest following a natural disturbance in north Guangdong[J]. Piant Science Journal, 2012, 30(3): 223−229.
[29] 陈龙斌, 孙昆, 张旭, 等. 林隙干扰对森林生态系统的影响[J]. 应用生态学报, 2021, 32(2):701−710. Chen L B, Sun K, Zhang X, et al. Effects of forest gap disturbance on forest ecosystem[J]. Chinese Journal of Applied Ecology, 2021, 32(2): 701−710.
[30] Coble A P, Cavaleri M A. Light drives vertical gradients of leaf morphology in a sugar maple (Acer saccharum) forest[J]. Tree Physiology, 2014, 34(2): 146−158. doi: 10.1093/treephys/tpt126
[31] 李兵兵, 秦琰, 刘亚茜, 等. 燕山山地油松人工林林隙大小对更新的影响[J]. 林业科学, 2012, 48(6):147−151. doi: 10.11707/j.1001-7488.20120622 Li B B, Qin Y, Liu Y Q, et al. Effects of gap size on regeneration of Pinus tabulaeformis plantation in the Yanshan Mountain[J]. Scientia Silvae Sinicae, 2012, 48(6): 147−151. doi: 10.11707/j.1001-7488.20120622
[32] 罗大庆, 郭泉水, 薛会英, 等. 西藏色季拉山冷杉原始林林隙更新研究[J]. 林业科学研究, 2002, 15(5):564−569. doi: 10.3321/j.issn:1001-1498.2002.05.010 Luo D Q, Guo Q S, Xue H Y, et al. A research of gap regeneration of virgin fir forest in Mount Sejila in Tibet[J]. Forest Research, 2002, 15(5): 564−569. doi: 10.3321/j.issn:1001-1498.2002.05.010
[33] 周振钊, 范春楠, 郭忠玲, 等. 长白山红松阔叶林林隙及更新特征[J]. 北华大学学报(自然科学版), 2019, 20(2):161−167. Zhou Z Z, Fan C N, Guo Z L, et al. Gap and regeneration characteristics of Korean pine broad-leaved forest in Changbai Mountain[J]. Journal of Beihua University (Natural Science), 2019, 20(2): 161−167.
[34] Downey M, Valkonen S, Heikkinen J. Natural tree regeneration and vegetation dynamics across harvest gaps in Norway spruce dominated forests in southern Finland[J]. Canadian Journal of Forest Research, 2018, 48(5): 524−534. doi: 10.1139/cjfr-2017-0358














计量
- 文章访问数: 903
- HTML全文浏览量: 309
- PDF下载量: 69
