
Study on forest road of fireproof blockade functions based on PyroSim
-
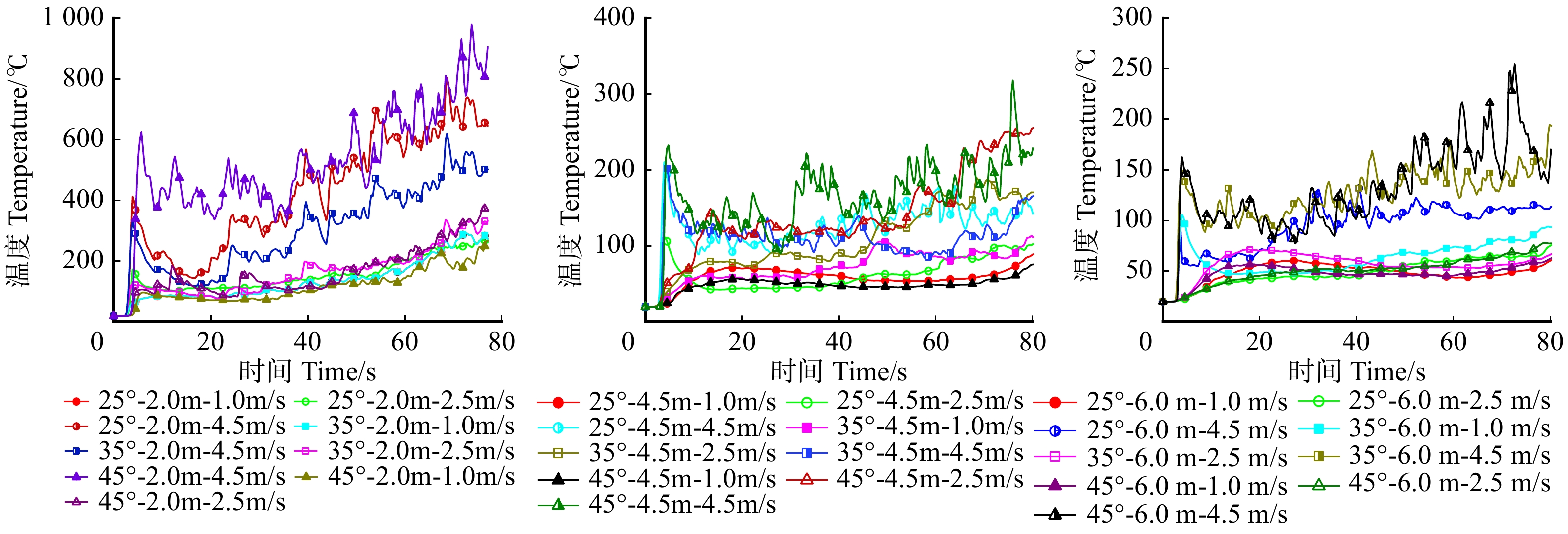
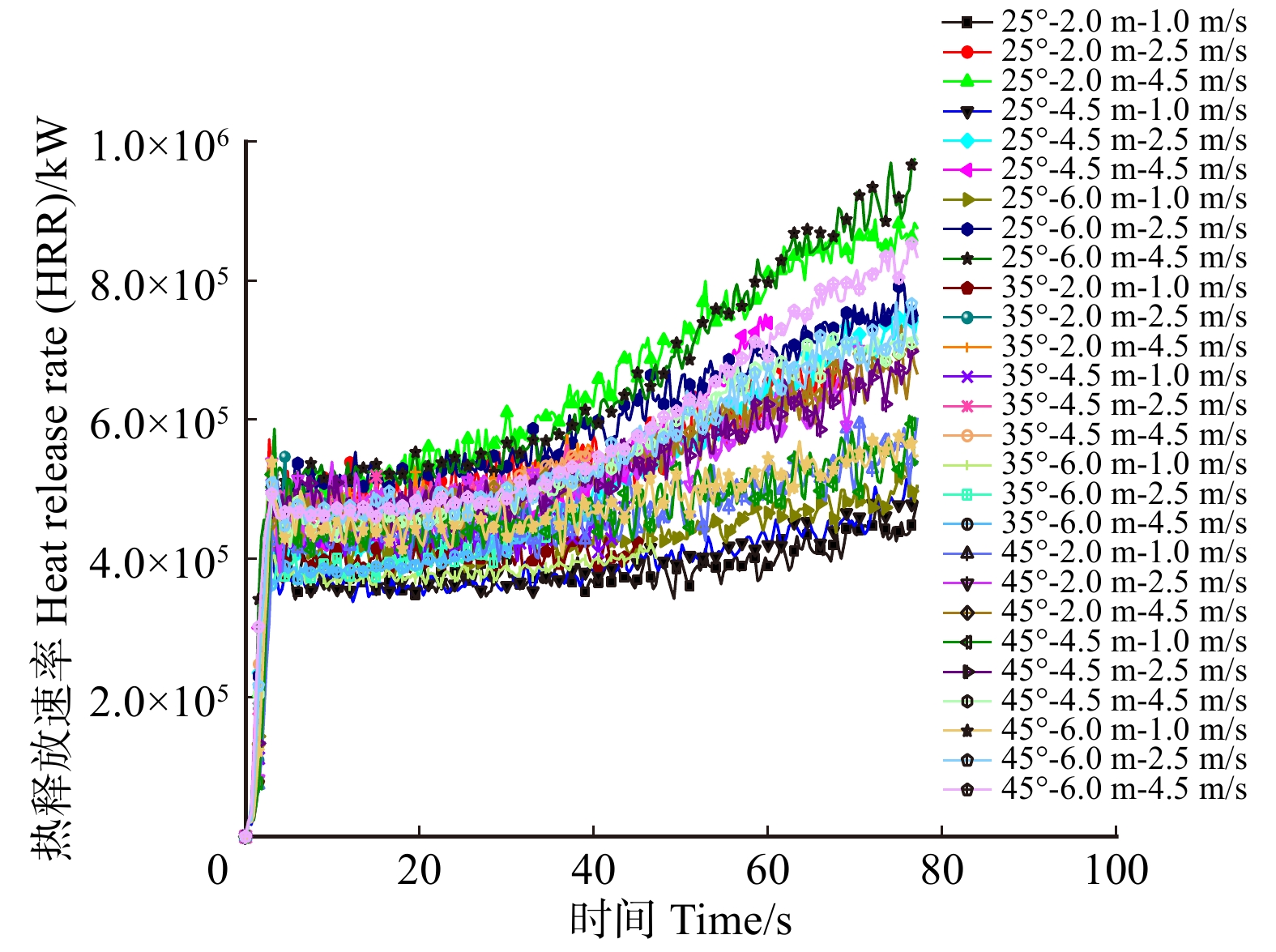
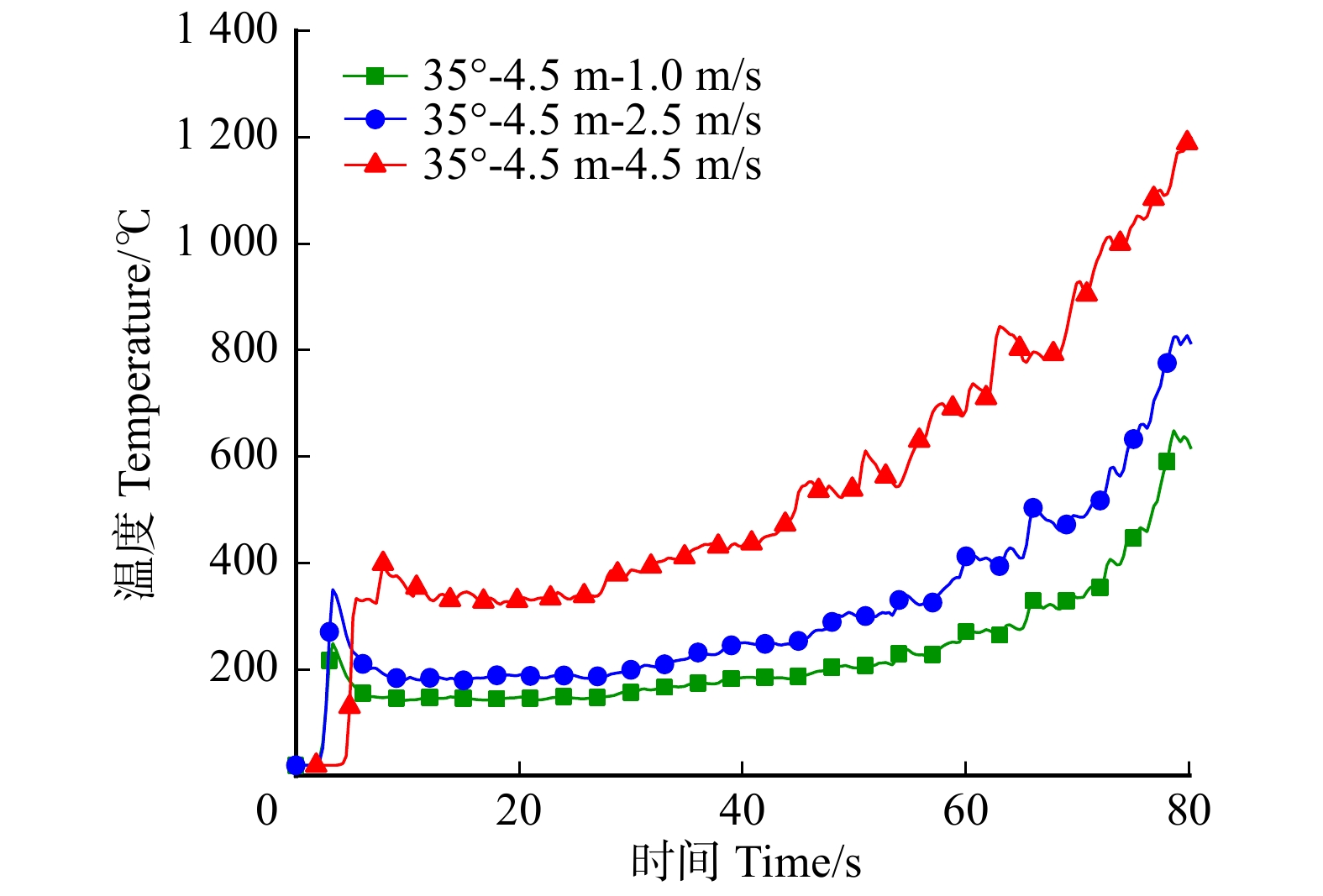
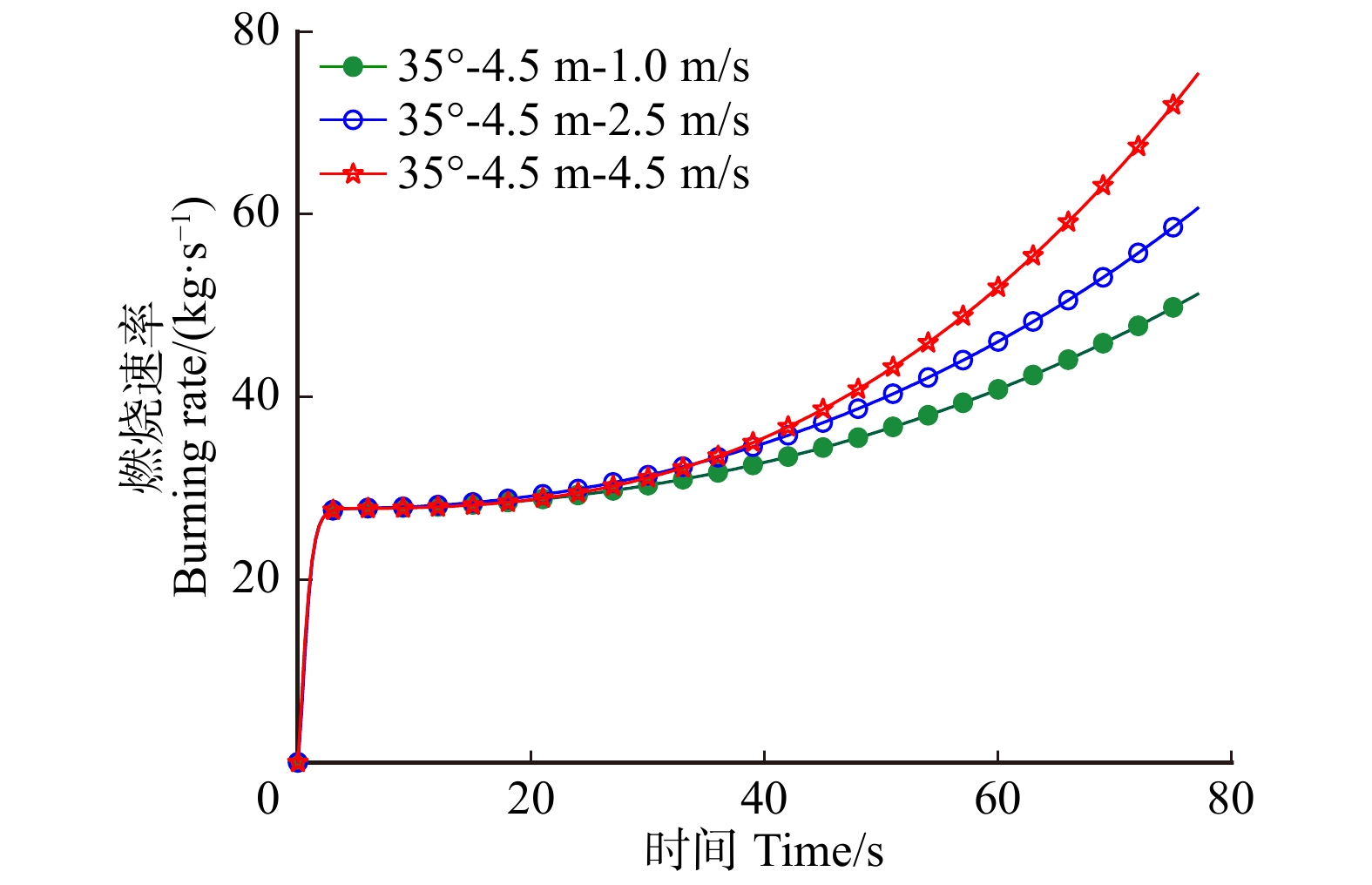
摘要:目的 林道网络及其宽度是林业生产、交通和旅游的需求,在发挥其最大经济效益的同时,兼顾林火巡护监测、应急救援,以及发生火灾时快速运送扑救人员和装备,阻隔森林火灾蔓延和发挥防火隔离功能。方法 运用火灾动态仿真模拟软件PyroSim分别以坡度25°、35°、45°,风速1、2.5、4.5 m/s,林道宽度2、4.5、6 m构建云南松林微观山体模型,施加长12 m、宽1 m、热释放速率4.6 × 104 kW/m2的带状火源,模拟中等强度林火研究不同林道宽度的防火阻隔功能。结果 林道宽度2 m无法满足防火隔离需求。林道宽度4.5 m的防火阻隔功能一般,当坡度超过35°、风速大于4.5 m/s时易失效。林道宽度6 m拥有较强的防火隔离功能,能较好阻隔中等强度的林火蔓延,可满足坡度小于45°、风速小于4.5 m/s的情况,当风速大于4.5 m/s时该林道宽度的隔离功能可能失效。中等强度的森林火灾火场中心温度在700 ~ 1 200 ℃左右,热释放功率可达3.0 × 105 ~ 9.0 × 105 kW,蔓延山坡10 m只需约1 min,可燃物燃烧速率60 s可达到40 kg/s以上。结论 云南省云南松林区道路宽度建议设置为6 m,能够取得较好的防火效益。常年处于高风速,坡度大于45°,树种高大的林区建议增加林道宽度至8 ~ 10 m。此外本研究应用PyroSim建立火场模型仿真模拟林道阻火功能是可行的,可为林区道路建设的合理设计、确定林道密度和宽度的阻火功能指标与经济效益相适应的最大限度阈值、降低森林火灾破坏力具有一定的指导意义。Abstract:Objective Forest road network and its width is the basic demands of forestry production, transportation, and tourism. While giving maximum economic benefits, its function on forest fire patrol monitoring, emergency rescue, rapid transport of firefighters and equipment when happening fires also should be taken into account, as well as the function of block forest fire spreading and forest fire isolation.Method The micro-mountains model of Pinus yunnanensis was constructed using fire dynamic simulation software PyroSim with parameters including slopes of 25°, 35° and 45°, wind speed of 1 , 2.5 and 4.5 m/s, and forest-road widths of 2, 4.5 and 6 m. And with the application of belt fire source with 12 m in length and 6 m in width, and heat release rate of 4.6 × 104 kW/m2, which simulated moderate-intensity forest-fire, we aimed to study fire-isolating functions at different forest-road widths.Result The forest road width of 2 m failed to meet the need for fire protection and isolation. And the one with 4.5 m achieved relatively poor fire-isolating function performance. And the one with more than 35° slope and greater than 4.5 m/s wind speed was easy to lose its function. And 6 m width road got a strong fire isolation function. Comparing with the one that can better isolate medium-intensity forest-fire spreading condition can meet the one with less than 4.5 m/s wind speed and less than 45° slope. When wind speed reached more than 4.5 m/s, the fireproof blockade functions of forest-road width of 6 m may be lost. Generally, the central temperature of moderate-intensity forest fire was about 700−1 200 ℃. And corresponding heat release rate power can reach 3.0 × 105 − 9.0 × 105 kW. And it only took about 1 min to spread 10 m on the hill. And the burning rate of combustibles can reach above 40 kg/s in the 60 s.Conclusion It is suggested to build the road with 6 m width in P. yunnanensis district in Yunnan Province of southwestern China, which can achieve better fire-prevention benefits. And road with width of 8−10 m is highly recommended in the forest area, where the stands with higher tree height all year round suffering high wind speed and slope greater than 45°. It also approve that the feasibility of simulating forest-road fire-isolating function by constructing the fire-site model with PyroSim, the results in this study have certain guiding significance on the rational designs of forest-road constructions, such as the determinations of maximum threshold values of fire prevention functions of forest-road with indexes of densities and widths, so can achieve certain economic benefits, as well as reductions of destructive powers of forest fires.
-
Keywords:
- Pinus yunnanensis /
- forest-road width /
- heat release rate /
- blockade function /
- PyroSim
-
多倍化是高等植物基因组进化的显著特征之一,大约有75%的被子植物和95%的蕨类植物在进化过程中发生过多倍化事件[1]。许多主要作物,包括小麦(Triticum aestivum,2n = 6x = 42)、土豆(Solanum tuberosum,2n = 4x = 48)、玉米(Zea mays,2n = 4x = 20)、咖啡豆(Coffea arabica,2n = 4x = 44)等,经过漫长的进化过程,最终生存下来的都是多倍体植株[2]。多倍体由于基因剂量效应,重复基因的表达情况也发生变化,导致植物在生长发育、形态生理、遗传适应性及对环境胁迫的耐性等方面与亲本二倍体有很大差异[3-5]。例如,相比二倍体,四倍体柑橘(Citrus limonia)叶片肥厚,茎干加粗,栅栏组织和海绵组织增长,表皮细胞体积增大,在高盐碱的逆境环境下,表现出更强的抗性[6-7];四倍体毛泡桐(Paulownia tomentosa)叶片面积、叶绿素含量、可溶性糖含量、可溶性蛋白质含量、净光合速率均大于二倍体,耐盐、耐寒、耐旱能力更强[8]。植物加倍后产生的新表型往往具有实际应用价值,如四倍体葡萄(Vitis vinifera)[9]、三倍体柑橘[10]、三倍体毛白杨(Populus tomentosa)[11]等许多品种都具有优于二倍体的性状,已经广泛应用于农林生产实践。
枣(Ziziphus jujuba)是鼠李科(Rhamnaceae)枣属(Ziziphus)植物,原产于中国,栽培历史悠久,枣果富含维生素和人体必需的氨基酸及多种微量元素[12],具有重要的营养和保健价值。酸枣(Ziziphus jujuba Mill. var. spinosa)作为枣的变种,果小、味酸,果仁可入药,其果实常用于制作果脯、果汁等,具有重要的经济价值[13]。此外,酸枣根系发达,耐寒、耐旱、耐贫瘠能力强[14],是重要的生态经济树种。近年来,枣树多倍体育种工作取得了显著的成果。Gu等[15]通过离体加倍获得了四倍体冬枣,相比二倍体,四倍体叶片变大变厚、颜色变深,保卫细胞内叶绿体增多,芽、茎变粗,花变大,花期延迟。刘孟军等[16]经秋水仙素诱变获得四倍体鲜食枣品种‘辰光’,具有果实大,果皮薄,果肉酥脆,早果速丰,枣头节间变短,叶缘锯齿增大,气孔密度减小,保卫细胞增大等特征。但目前尚未在分子水平上对这些变异特征进行深度探讨。
在前期工作中,本实验室以酸枣组培苗为试材进行秋水仙素离体诱导,成功获得了酸枣四倍体种质,并对二倍体和四倍体酸枣进行了表型特征的初步对比[17]。本研究以实验室原有的酸枣二倍体和四倍体组培苗为材料,对二者的生理指标进行测定,同时开展RNA-seq技术的转录组学研究,分析酸枣多倍体化导致的基因表达变化。通过该研究,有利于进一步了解基因组复制后分子水平的变异,深入理解酸枣四倍体性状变异,为今后关键调控基因的克隆及基因工程育种等工作提供参考。
1. 材料与方法
1.1 试验材料
以北京林业大学林木遗传育种国家工程实验室已有的酸枣二倍体及其四倍体组培苗为试验材料。组培室内温度维持在26 ℃,光周期为16 h光照/8 h黑暗。选取继代60 d的生长健壮的二倍体和四倍体酸枣组培苗,转接到生根培养基上,生根后30 d进行生理指标的测定及转录组测序。
分别随机选取二倍体和四倍体组培苗各5株,采集叶片测定叶绿素含量及叶片相对含水量,重复3次。同时,二倍体和四倍体组培苗分别随机选取5个植株,采集植物的叶片和茎,重复3次,取样后迅速放入液氮中磨成粉末,置于− 80 ℃冰箱保存,用于可溶性糖、可溶性蛋白含量测定以及转录组测序。
1.2 生理指标的测定
1.2.1 测定方法
用SPAD-502Plus仪测定叶绿素含量;用饱和称重法测定植物的叶片相对含水量[18];采用蒽酮比色法测定可溶性总糖含量[19];采用考马斯亮蓝染色法测定可溶性蛋白质含量[20]。
1.2.2 数据分析
使用Excel 2010进行数据录入及处理,用SPSS软件对数据进行独立样本t检验(P < 0.05)。
1.3 RNA提取
使用上海天根生化有限公司(Qiagen China, Shanghai, China)生产的RNeasy plant Mini kits试剂盒提取RNA,提取过程按说明书中的操作流程进行。用天根公司生产的RNase-Free DNaseSet对提取的RNA进行纯化。用琼脂糖电泳对提取出来的RNA完整性进行检测,紫外灯下观察是否具有清晰的条带。在进行转录组测序之前,对提取的RNA质量进一步评估,计算A260/A280是否介于1.8 ~ 2.0之间,28S/18S是否介于1.6 ~ 2.0之间,再分别用NanoDrop 2000分析仪(DE, USA)和Agilent 2100生物芯片分析系统检测RNA浓度和完整性,RNA检测合格之后用于后续分析。
1.4 转录组测序
委托北京百迈客生物科技有限公司(Biomarker Technology, Beijing, China)用Illunima HiSeq 2000测序系统进行cDNA文库构建及转录组测序(RNA-seq)。获得raw data后,去除测序接头及引物序列,并将低质量的序列过滤掉(Q30 ≥ 93.96%),得到clean data。利用HISAT2软件(http://ccb.jhu.edu/software/hisat2/index.shtml)将clean data比对到枣树参考基因组(https://www.ncbi.nlm.nih.gov/genome/15586?genome_assembly_id=219393),得到mapped data,进行插入片段长度检验、随机性检验等文库质量评估。
采用NCBI(https://www.ncbi.nlm.nih.gov/)、GO Ontology(http://geneontology.org/)、KEGG(https://www.kegg.jp/)、KOG(http://www.ncbi.nlm.nih.gov/KOG/)、Pfam(https://pfam.xfam.org/)、Swiss-Prot(https://www.uniprot.org/)数据库对基因组功能进行注释。基于所选参考基因组序列,使用StringTie(https://ccb.jhu.edu/software/stringtie/index.shtml)软件对mapped reads进行拼接,并与原有的基因组注释信息进行比较,寻找原来未被注释的转录区,过滤掉编码过短肽链(少于50个氨基酸残基)或只包含单个外显子的序列,发掘该物种的新基因,从而补充和完善原有的基因组注释信息。
1.5 差异基因的鉴定及生物信息学分析
应用DESeq2软件(http://www.bioconductor.org/packages/release/bioc/html/DESeq.htm)进行样品组间的差异表达分析,将差异倍数(FC) > 1.5且错误发现率(FDR) < 0.05作为筛选差异表达基因(DEG)的标准。对筛选出的DEG进行GO Ontology和KEGG Pathway分类,并应用超几何检验对注释到的不同的GO(P < 0.05)和KEGG条目(P < 0.05)进行富集分析。用FPKM值(Fragments per kilobase of transcript per million fragments mapped)作为衡量基因表达水平的指标,R语言包绘制表达量聚类热图。
2. 结果与分析
2.1 二倍体及其同源四倍体酸枣生理指标对比
植物染色体加倍后,细胞核内的遗传物质相应增加,一些营养物质或其他成分含量显著提高。由表1可知,四倍体酸枣可溶性糖、可溶性蛋白质、叶绿素含量显著高于二倍体,可溶性糖含量是二倍体的2倍;可溶性蛋白质比二倍体高50.1%;叶绿素含量比二倍体高27.2%。但是,四倍体植株叶片相对含水量与二倍体没有显著差异。
表 1 二倍体和四倍体酸枣不同生理指标的比较Table 1. Comparison of different physiological index between diploid and its autotetraploid sour jujube样品
Sample可溶性糖
Soluble sugar可溶性蛋白
Soluble protein叶绿素
Chlorophyll叶片相对含水量
Leaf relative water content二倍体酸枣
Diploid sour jujube25.03 ± 4.73a 4.13 ± 0.63a 29.17 ± 3.02a 0.52 ± 0.01a 四倍体酸枣
Autotetraploid sour jujube41.15 ± 4.45b 6.20 ± 0.62b 37.10 ± 2.82b 0.57 ± 0.01a 注:表内数值表示方式为“平均值 ± 标准差”,同列不同字母表示差异显著(P < 0.05)。Notes: values in table were “mean ± SD”, different letters in the same column represent significant differences (P < 0.05). 2.2 测序结果统计
为进一步理解二倍体酸枣加倍后变异的分子机制,对二倍体和四倍体酸枣共6个样本进行RNA-seq,获得平均7.92 Gb clean reads,Q30均在94%以上,将这些数据比对到枣树基因组,比对效率在82%以上,表明测序结果良好(表2)。
表 2 测序结果统计Table 2. Statistics of sequencing results样品 Sample Clean reads Clean bases Q30/% GC 含量 GC content/% 比对率 Mapped ratio/% D1 25 635 963 7 620 864 926 94.57 45.34 83.31 D2 27 771 165 8 268 560 788 94.43 45.1 83.67 D3 29 668 511 8 501 884 148 94.62 45.41 83.38 T1 27 572 679 7 659 547 606 94.21 45.19 82.92 T2 27 039 014 7 346 405 010 94.52 45.46 83.27 T3 29 816 038 8 096 301 012 94.53 45.32 82.52 注:D1、D2、D3分别为二倍体酸枣的3个重复;T1、T2、T3分别为四倍体酸枣的3个重复。Notes: D1, D2 and D3 are three biological duplicates of diploid sour jujube; T1, T2 and T3 are three biological duplicates of autotetraploid sour jujube. 通过对原有的枣树基因组进行补充和完善,共发掘2 536个新基因。将这些新基因与GO、KEGG、KOG、Pfam、Swiss-Prot、Nr数据库进行序列比对,获得新基因的注释信息(图1)。
2.3 二倍体及其同源四倍体酸枣差异表达基因分析
对二倍体和四倍体酸枣的差异表达基因进行分析。结果表明,二倍体与四倍体酸枣共有1 329个基因差异表达(FC > 1.5, FDR < 0.05),其中四倍体酸枣有636个基因表达量高于二倍体,表达量倍数介于1.5 ~ 18.2之间;693个基因表达量低于二倍体,表达量倍数介于1.5 ~ 11.8之间(图2)。
![]() 图 2 二倍体与四倍体酸枣差异表达基因统计红色的点代表四倍体相对于二倍体表达量上调的基因,绿色的点代表表达量下调的基因,黑色的点代表没有差异的基因;FC代表差异倍数;FDR代表错误发现率。The red point indicates up-regulated genes in autotetraploid than that in diploid, green point indicates down-regulated genes and black point indicates unchanged genes; FC represents fold change; FDR represents false discovery rate.Figure 2. Statistics of differentially expressed genes in diploid and autotetraploid sour jujube
图 2 二倍体与四倍体酸枣差异表达基因统计红色的点代表四倍体相对于二倍体表达量上调的基因,绿色的点代表表达量下调的基因,黑色的点代表没有差异的基因;FC代表差异倍数;FDR代表错误发现率。The red point indicates up-regulated genes in autotetraploid than that in diploid, green point indicates down-regulated genes and black point indicates unchanged genes; FC represents fold change; FDR represents false discovery rate.Figure 2. Statistics of differentially expressed genes in diploid and autotetraploid sour jujube2.3.1 差异表达基因GO功能分类与富集分析
为了分析二倍体与四倍体差异基因的生物学功能,分别从生物学过程、细胞组分和分子功能3个方面对注释到GO Ontology数据库中的1 064个差异基因进行功能分类(表3)。在生物学过程分类中,参与细胞过程、单一生物过程和代谢过程的差异基因数目最多,分别为873、851和829个,其次是胁迫应答过程,为753个;在细胞组分分类中,细胞部位和细胞差异基因最多,分别为878和864个,其次是细胞器为714个。在分子功能分类中,参与催化活性的差异基因最多,为655个,其次是参与结合功能的差异基因为620个。
表 3 二倍体与四倍体酸枣差异表达基因的GO分类及富集分析Table 3. GO classification and enrichment analysis of differentially expressed genes in diploid and autotetraploid sour jujubeGO一级分类
GO classify 1GO ID GO二级分类
GO classify 2所有基因
All gene差异基因
Different gene富集P值
Enriched P value生物学过程
Biological processGO:0009987 细胞过程 Cellular process 15 527 873 0.725 1 GO:0008152 代谢过程 Metabolic process 14 536 829 0.983 6 GO:0044699 单一生物过程 Single-organism process 14 286 851 0.004 2 GO:0050896 胁迫应答 Response to stimulus 11 883 753 0.000 0 GO:0065007 生物调控 Biological regulation 9 802 603 0.021 6 GO:0032502 发育过程 Developmental process 8 154 492 0.003 7 GO:0071840 细胞组分或生物合成
Cellular component organization or biogenesis7 166 386 0.373 7 GO:0051179 定位 Localization 6 533 378 0.054 4 GO:0032501 多细胞生物过程 Multicellular organismal process 6 469 372 0.090 1 GO:0000003 生殖 Reproduction 5 573 331 0.070 8 GO:0022414 生殖过程 Reproductive process 5 536 332 0.065 2 GO:0051704 多生物过程 Multi-organism process 4 643 249 0.002 4 GO:0023052 信号传导 Signaling 3 681 237 0.184 7 GO:0040007 生长 Growth 2 639 142 0.016 5 GO:0002376 免疫系统过程 Immune system process 2 446 131 0.001 1 GO:0098754 解毒 Detoxification 550 49 GO:0048511 节律过程 Rhythmic process 294 26 0.064 5 GO:0022610 生物附着 Biological adhesion 134 7 0.413 8 GO:0040011 运动 Locomotion 73 4 0.295 6 GO:0001906 细胞杀伤 Cell killing 15 2 0.556 6 细胞组分
Cellular componentGO:0044464 细胞部位 Cell part 16 088 878 0.000 0 GO:0005623 细胞 Cell 15 666 864 0.713 8 GO:0043226 细胞器 Organelle 12 958 714 0.659 2 GO:0016020 细胞膜 Membrane 8 588 489 0.740 6 GO:0044422 细胞器部位 Organelle part 5 560 302 0.938 5 GO:0044425 细胞膜部位 Membrane part 2 828 163 0.948 7 GO:0032991 高分子复合物 Macromolecular complex 2 285 81 0.999 8 GO:0030054 细胞连接点 Cell junction 2 259 154 0.438 3 GO:0005576 胞外区 Extracellular region 2 091 173 0.000 2 GO:0031974 膜封闭腔 Membrane-enclosed lumen 563 21 0.284 9 GO:0099080 超分子复合物 Supramolecular complex 154 14 GO:0044421 胞外区部位 Extracellular region part 132 11 0.428 4 GO:0009295 拟核 Nucleoid 45 4 0.058 4 分子功能
Molecular functionGO:0005488 结合 Binding 11 517 620 0.983 3 GO:0003824 催化活性 Catalytic activity 10 913 655 0.587 2 GO:0005215 转运活性 Transporter activity 1 496 91 0.801 3 GO:0001071 核酸结合转录因子活性
Nucleic acid binding transcription factor activity1 062 76 0.327 1 GO:0004871 信号传导活性 Signal transducer activity 782 23 0.216 1 GO:0060089 分子传感器活动 Molecular transducer activity 627 21 0.216 1 GO:0005198 结构分子活动 Structural molecule activity 437 16 0.914 3 GO:0009055 电子载体活动 Electron carrier activity 350 27 0.040 0 GO:0098772 分子功能调节剂 Molecular function regulator 285 23 GO:0016209 抗氧化活性 Antioxidant activity 179 15 0.489 6 GO:0000988 转录因子活性,蛋白结合
Transcription factor activity, protein binding78 3 0.763 7 GO:0045735 营养库活动 Nutrient reservoir activity 45 1 0.154 2 利用KS检验,以富集P值小于0.05作为评价标准,在二倍体与四倍体差异表达基因中满足此条件的GO条目即为GO功能显著富集。在生物过程分类中显著富集了包括胁迫应答过程、生物调控、发育过程、生长过程和免疫系统过程等7个条目;在细胞组分类别中显著富集的GO条目有2个,包括细胞部位和胞外区组成;在分子功能类别中,显著富集的条目只有1个,为电子载体活动。结果说明,二倍体与四倍体酸枣在生长发育、细胞代谢过程、逆境胁迫抗性等方面存在差异。
2.3.2 差异表达基因KEGG Pathway富集分析
二倍体与四倍体酸枣共有244个差异表达基因注释到特定的KEGG 通路中,这些通路可以分为细胞过程、环境信息加工、遗传信息加工、代谢和生物系统5类,其中在代谢中注释到的差异基因数目最多,占差异基因总注释数目的73.8%(图3)。有13条通路在KEGG Pathway中显著富集(P < 0.05),(图4),主要包括氨基酸代谢、碳水化合物代谢、信号传导、能量代谢等。碳水化合物代谢通路中,参与淀粉和蔗糖代谢途径的差异表达基因数目最多,占注释的差异表达基因总数的10.66%;信号传导通路中,参与植物激素信号转导途径的差异基因有18个,占注释的差异表达基因总数的7.38%;在氨基酸代谢通路中,丙氨酸,天门冬氨酸和谷氨酸代谢、精氨酸和脯氨酸代谢以及酪氨酸代谢3个途径差异基因数目分别占注释的差异基因总数的2.5%、3.3%和3.7%;在能量代谢通路中,光合天线蛋白途径中注释的差异基因占注释差异基因总数的1.2%。此外,相比二倍体植株,四倍体植株中表达量增加的差异基因显著富集在淀粉与蔗糖代谢(P < 0.000 9)、植物激素信号传导(P < 0.004 0)等通路中。以上结果表明,二倍体与四倍体酸枣在碳水化合物和氨基酸代谢、信号传递以及能量代谢途径中存在差异,其中,与淀粉和蔗糖代谢、植物激素传导途径相关的基因在四倍体中显著上调。
![]() 图 3 二倍体与四倍体酸枣差异表达基因KEGG分类Figure 3. KEGG classification of differentially expressed genes in diploid and autotetraploid sour jujube
图 3 二倍体与四倍体酸枣差异表达基因KEGG分类Figure 3. KEGG classification of differentially expressed genes in diploid and autotetraploid sour jujube![]() 图 4 二倍体与四倍体酸枣差异表达基因显著富集的KEGG通路Figure 4. KEGG pathway with significantly enriched differentially expressed genes in diploid and autotetraploid sour jujube
图 4 二倍体与四倍体酸枣差异表达基因显著富集的KEGG通路Figure 4. KEGG pathway with significantly enriched differentially expressed genes in diploid and autotetraploid sour jujube2.3.3 差异基因表达量分析
为进一步解析二倍体及其同源四倍体可溶性物质的差异,对与糖和蛋白质转运、代谢相关的16个基因进行差异表达分析(图5)。结果发现,与糖代谢相关的关键基因,如编码β-呋喃果糖苷酶的INV(CCG007195)基因、编码β-葡萄糖苷酶的BGLU47(CCG000062)基因、编码果胶酯酶的PME35(CCG011493)基因、编码葡糖醛酸异构酶的GAE6(CCG027883)基因、编码α-海藻糖磷酸合成酶的TPS9(CCG011925)基因和编码蔗糖磷酸合成酶的SPS2(CCG011942)基因等均上调表达,差异表达倍数介于1.5 ~ 2.5之间。与氨基酸转运和代谢相关的基因,例如编码丝氨酸乙酰转移酶的SAT1(CCG028964)基因、编码精氨琥珀酸合酶(NewGene3586)和磷酸甘油酸脱氢酶PGDH3(CCG002072)等的基因在四倍体酸枣中上调表达,表达量为二倍体的1.7 ~ 2.0倍。
![]() 图 5 二倍体与四倍体差异基因表达量热图D1、D2、D3分别为二倍体酸枣的3个生物学重复;T1,T2,T3分别为四倍体酸枣的3个生物学重复。方块中不同的颜色表示基于FPKM值的基因表达水平;每行的数据分别进行标准化;蓝色表示在四倍体中表达量下调,红色表示在四倍体中表达量上调。D1, D2 and D3 are the three duplicates of diploid sour jujube; T1, T2 and T3 were the three duplicates of autotetraploid sour jujube. Different colors indicate different levels of gene expression based on the FPKM values; the data in each row were normalized and compared, separately; blue indicates down-regulation in autotetraploid, and red indicates up-regulation.Figure 5. Heat map of differentially expressed genes in the diploid and autotetraploid sour jujube
图 5 二倍体与四倍体差异基因表达量热图D1、D2、D3分别为二倍体酸枣的3个生物学重复;T1,T2,T3分别为四倍体酸枣的3个生物学重复。方块中不同的颜色表示基于FPKM值的基因表达水平;每行的数据分别进行标准化;蓝色表示在四倍体中表达量下调,红色表示在四倍体中表达量上调。D1, D2 and D3 are the three duplicates of diploid sour jujube; T1, T2 and T3 were the three duplicates of autotetraploid sour jujube. Different colors indicate different levels of gene expression based on the FPKM values; the data in each row were normalized and compared, separately; blue indicates down-regulation in autotetraploid, and red indicates up-regulation.Figure 5. Heat map of differentially expressed genes in the diploid and autotetraploid sour jujube对与内源激素相关的23个差异基因表达量进行比较分析(图5),结果发现,在二倍体植株中,编码油菜素内酯生物合成相关蛋白(CYP)的基因(CCG015397、CCG007876、CCG007877)的表达量是四倍体的1.8 ~ 2.1倍;编码乙烯不敏感蛋白的基因EIN2(CCG001434)的表达量是四倍体的1.5倍。在四倍体植株中,与诱导、响应和转运生长素相关的基因,如ARG7(Addgene2586)、ARF9(CCG010227)、GH3.6(CCG025779)、IAA26(CCG025671、CCG025726)、LAX3(CCG001581)、X15(CCG013493)的表达量是二倍体的1.5 ~ 2.1倍。说明酸枣四倍体在表型上呈现的变异特征可能与这些关键基因的差异表达有关。
2.4 二倍体及其同源四倍体酸枣的差异转录因子分析
转录因子参与了许多生物学进程,在调控下游功能基因方面起重要作用。为了解酸枣加倍后的转录因子变异,对二倍体和四倍体酸枣进行了转录因子功能预测,发现与转录因子相关的差异基因共158个(FC > 1.5, FDR < 0.05),包含14个编码转录抑制子的基因,58个蛋白激酶相关基因和86个编码转录因子的基因,共编码35种转录因子(图6)。编码AP2/ERF-ERF转录因子的基因数目最多,其次为HB-HD-ZIP、LOB和MYB转录因子。四倍体植株中表达量高于二倍体的编码转录因子的基因有102个,属于28种转录因子,其中,编码MYB转录因子的基因最多。
![]() 图 6 编码转录因子的差异基因数目统计Figure 6. Statistics of the number of differentially expressed genes encoding transcription factors
图 6 编码转录因子的差异基因数目统计Figure 6. Statistics of the number of differentially expressed genes encoding transcription factors3. 讨 论
3.1 二倍体与四倍体生理特征差异
四倍体通常较二倍体具有更大的器官、细胞、更多的次生代谢物含量[21],这使得四倍体成为植物育种专家增加植物产量的重要手段之一。本研究中发现四倍体酸枣的叶绿素含量、可溶性糖和可溶性蛋白含量均显著高于二倍体。这与白桦(Betula platyphylla)[22]和刺槐(Robinia pseudoacacia)[23]基因组加倍后植株的生理表现相同。这些生理指标的差异可能是四倍体植株形态异于二倍体的重要原因之一,叶绿素含量高可能导致四倍体叶色更深,而可溶性糖和可溶性蛋白含量高为植株提供更多的能源物质,从而导致四倍体叶片更大、茎更粗。进一步说明,叶片细胞中的叶绿素含量、可溶性糖含量和可溶性蛋白质含量均可作为初步鉴定四倍体酸枣的辅助指标。但是,研究中发现叶片相对含水量在二倍体与四倍体酸枣中没有显著差异,这与四倍体连翘(Forsythia suspensa)[24]、四倍体甜瓜(Cucumis melo)[25]的研究结果表现一致。
3.2 二倍体与四倍体差异基因分析
植物多倍化后引起表型及生理的变异,这主要是由于基因表达改变引起的。大量研究发现植物中基因表达发生微小的变化都可能引起表型的变异[26]。高通量测序技术已经成为多倍体植物优良性状解析的常用策略[27],本研究中我们利用RNA-seq技术对酸枣二倍体和四倍体进行转录组学分析,测序数据分析结果表明测序结果良好。基因差异表达是四倍体酸枣变异的分子基础。因此,我们进一步对二倍体与四倍体酸枣的差异表达基因进行分析,二者共有1 329个基因差异表达,通过GO Ontology分析表明在胁迫应答、生物调控、发育、生长过程和免疫系统7个过程中差异基因富集,说明四倍体在生长发育、抗性方面与二倍体存在差异。KEGG通路分析发现差异基因显著富集在碳水化合物和氨基酸代谢、信号传递以及能量代谢方面,其中在淀粉与蔗糖代谢、植物激素传导通路中显著上调表达。这些差异基因在四倍体中的表达量升高可能参与了可溶性糖和内源生长素的积累过程,为四倍体提供更多能源物质,从而增强植株生长势。这一结果与杨树三倍体研究中发现的主要参与碳水化合物代谢、细胞生长的基因上调使其在生长表现上优于二倍体的结果相同[28]。
3.3 二倍体与四倍体差异基因功能分析
酸枣经基因组加倍后,表现为合成更多的可溶性糖和可溶性蛋白,这可能与众多关键基因在四倍体中的差异表达有关。前人研究表明,GAE6催化拟南芥半乳糖醛酸的合成,促进果胶多糖的形成[29];SPS控制光合产物向蔗糖和淀粉的分配,在蔗糖的形成过程中起正向调节作用[30]。在本研究中GAE6和SPS2在四倍体酸枣中上调表达,可能是导致四倍体植株体内可溶性糖含量增加的关键基因。此外,BGLU47、TPS9和SPS2编码糖代谢过程中的关键酶,可以通过调节植物体内渗透胁迫进而增强植物承受逆境胁迫的能力[31]。本研究对这些关键调控基因分析发现,此类基因在四倍体中具有较高的表达量,推测四倍体酸枣可能具有较强的渗透调节能力,具体性状还需要进一步的实验研究。植物磷酸丝氨酸途径在植物发育和次生代谢产物的生物合成中具有重要作用,PGDH3编码催化这一途径的PGDH酶,与色氨酸的生物合成呈正相关[32]。本研究中四倍体内PGDH3表达量显著高于二倍体,可能是导致四倍体酸枣体内可溶性蛋白含量较高的原因之一。
植物生长发育与内源激素的调节有密不可分的联系,内源激素的水平会显著影响细胞分化、糖合成与代谢等生理过程[33]。青杨(Populus cathayana)三倍体中参与生长素、细胞分裂素、油菜素内酯等激素合成与转导的基因相比二倍体显著上调表达导致三倍体生长势增强[34];桑树(Morus alba)四倍体与二倍体的差异基因参与了生长素和乙烯等激素的合成及传导过程,使四倍体较二倍体具有更大的生长优势[35]。在本研究中,四倍体与二倍体有18个差异基因显著富集在植物激素信号传导通路中,其中12个基因在四倍体中表达量较高,包括与生长素诱导相关的基因ARG7(SAUR家族)、GH3.6(GH3家族)、X15(SAUR家族),生长素响应基因IAA26(AUX/IAA家族),编码转运蛋白的基因LAX3(AUX1家族)以及生长素应答因子ARF9(ARF家族)等(图7),这些基因不仅正调控IAA的生物合成,还参与了激素的信号转导过程[36-38]。6个基因在四倍体中表达量较低,包括细胞色素P450单加氧酶(CYP)和乙烯不敏感蛋白(EIN)等,CYP参与甾醇催化形成油菜素甾醇的过程,从而促进叶片伸展和茎的伸长生长[39];EIN2是正调控乙烯信号传导通路的中心元件,与器官的衰老、成熟有关[40-41]。因此,编码内源激素的差异基因表达量的变化可能是导致四倍体生长表型变异的重要原因之一,对此还需要进一步实验验证。
![]() 图 7 与内源生长素相关的表达通路方块内的红色背景代表基因在四倍体中上调表达,蓝色背景代表基因在四倍体中既有上调表达又有下调表达。The red background in the square indicates up-regulated expression of genes in autotetraploid than that of diploid, while the blue background indicates up-regulated and down-regulated expressed genes.Figure 7. Expression pathway related to endogenous auxin
图 7 与内源生长素相关的表达通路方块内的红色背景代表基因在四倍体中上调表达,蓝色背景代表基因在四倍体中既有上调表达又有下调表达。The red background in the square indicates up-regulated expression of genes in autotetraploid than that of diploid, while the blue background indicates up-regulated and down-regulated expressed genes.Figure 7. Expression pathway related to endogenous auxin3.4 二倍体与四倍体转录因子差异分析
转录因子通过与下游基因相互作用,激活或抑制其他基因的转录,或者与其他转录因子互作调节基因的表达,从而引起一系列的应答反应,改变植物性状[42]。MYB转录因子广泛参与了植物的生长发育与形态建成[43]、次生代谢[44]和环境胁迫应答[45]等过程。在本研究中,我们分析二倍体和四倍体酸枣的差异转录因子,发现四倍体植株中编码MYB转录因子的基因在所有转录因子类型中上调表达数目最多。这一结果与菘蓝(Isatis indigotica)四倍体研究中发现的MYB在根、茎、叶中表达量高于二倍体的结果相一致[46],因此,四倍体在表型和生理方面的变异特征可能与编码MYB转录因子的基因上调表达有关。此外,MYB转录因子的差异表达还导致了菘蓝四倍体抗性增强,说明酸枣四倍体在对环境胁迫耐受性方面可能高于二倍体。因此,今后应该进一步对酸枣四倍体的抗性进行研究。
多倍体植物在基因组复制后含有新的基因调控网络,完整的RNA-seq数据对于理解新的调控网络至关重要。本研究在检测生理指标的基础上,对酸枣不同倍性植株进行转录组测序并结合性状分析了差异表达基因的功能,为深入理解酸枣四倍体的变异性状提供参考,通过进一步的研究筛选候选基因,为枣树关键调控基因的克隆及基因工程育种提供分子基础。
-
![]()
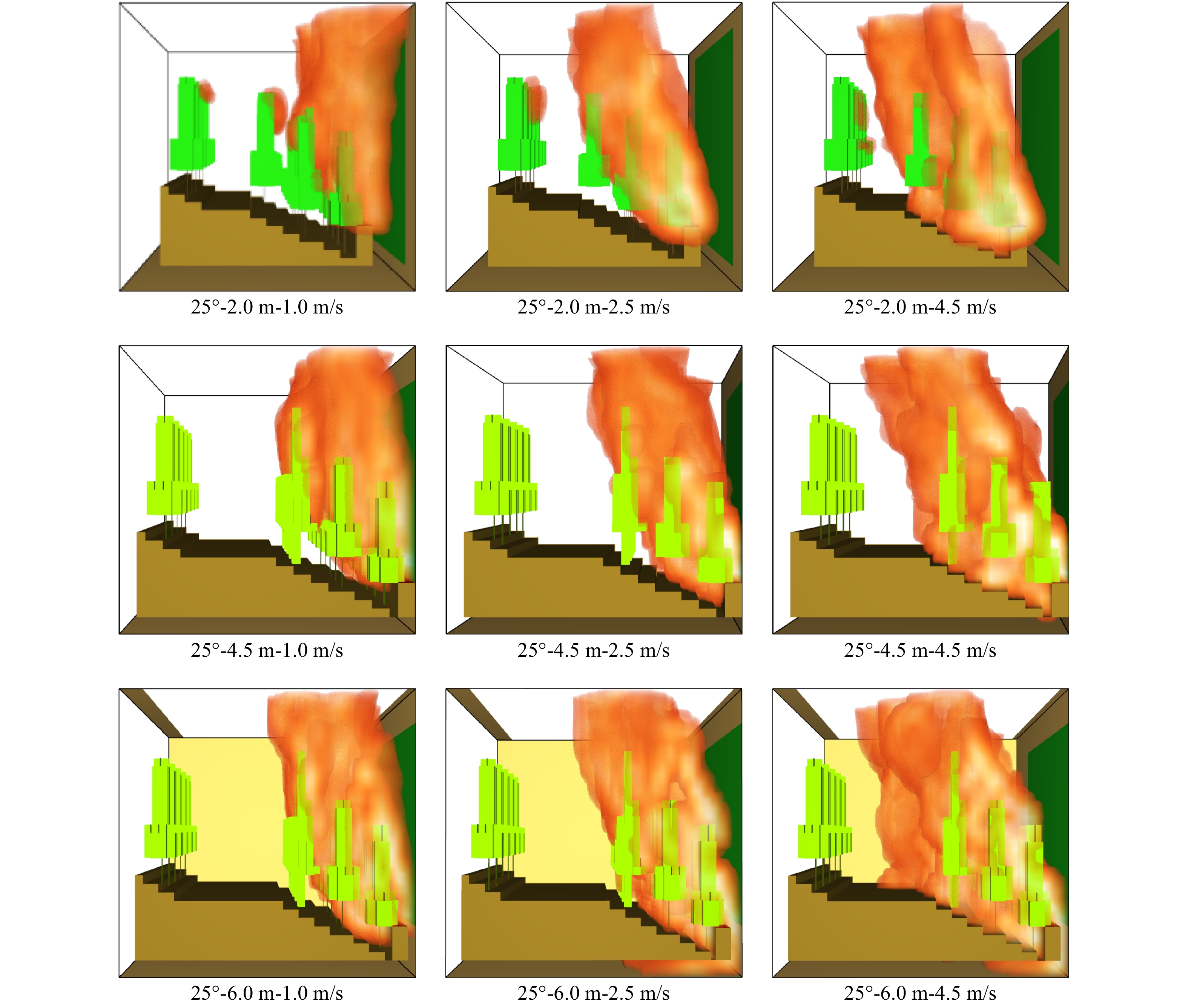
图 4 坡度25°时不同林道宽度对不同风速条件下林火蔓延的阻隔模拟
Figure 4. Blockade simulation on various widths of forest roads to fire spread under different wind speeds at slope of 25°
![]()
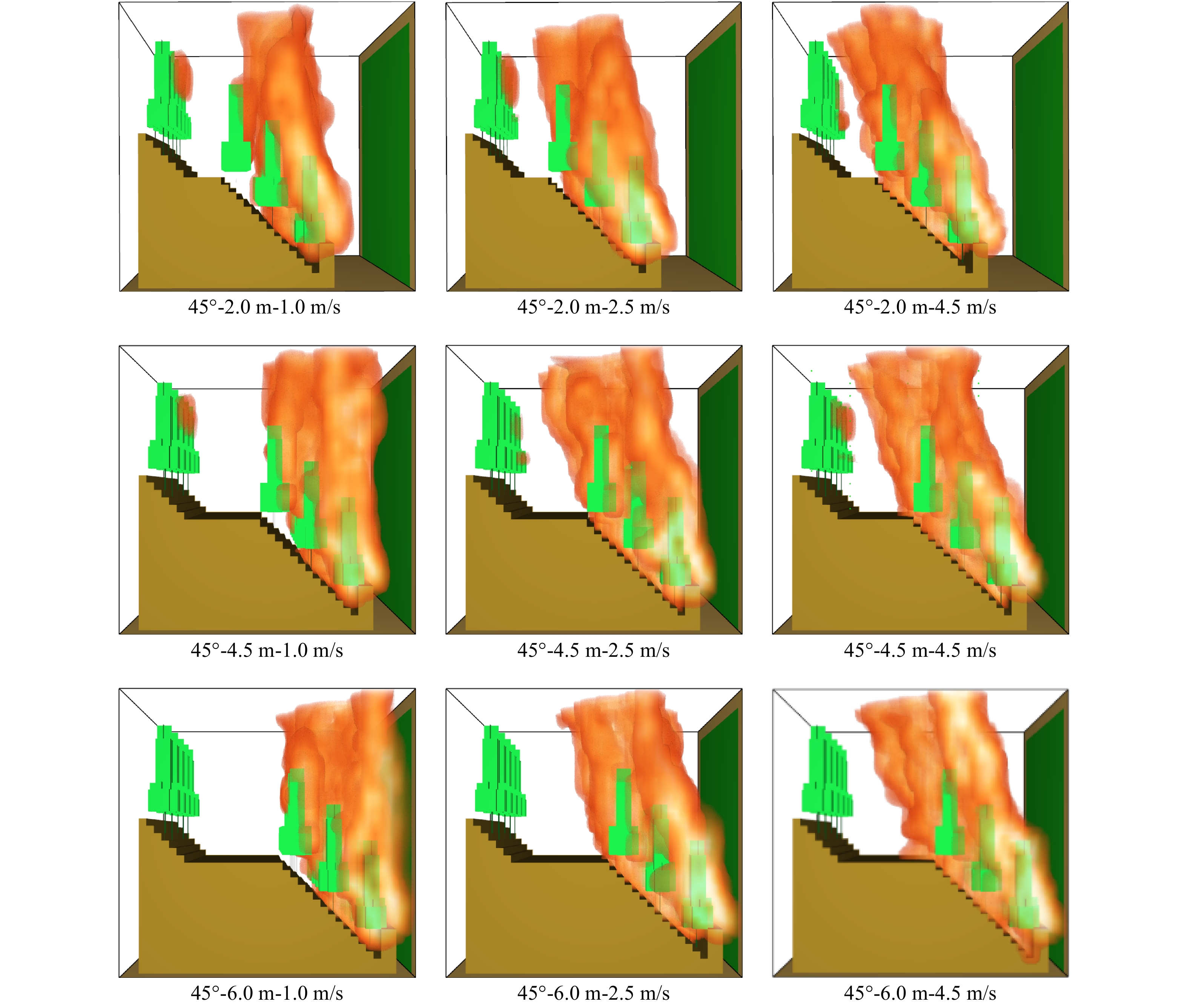
图 5 坡度35°时不林道宽度对不同风速条件下林火蔓延的阻隔
Figure 5. Blockade simulation on various widths of forest roads to fire spread under different wind speeds at slope of 35°
![]()
图 6 坡度45°时林道宽度对不同风速条件下林火蔓延的阻隔
Figure 6. Blockade simulation on various widths of forest roads to fire spread under different wind speeds at slope of 45°
表 1 4种标准t2火
Table 1 Four kinds of t2 standard fire
火灾增长类型
Fire growth type增长系数
Growth coefficient/(kW·s−2)达到1 MW需要时间
Time of reaching 1 MW/s典型可燃材料
Typical combustible material超快速 Ultra-fast speed 0.187 800 75 油池火、易燃的装饰家具、轻的窗帘
Pool fire, flammable decorative furniture, light curtains快速 Fast speed 0.046 890 150 装满东西的邮袋、塑料泡沫、叠放的木架
A mail bag full of things, plastic foam, stacked wooden shelves中速 Medium speed 0.011 270 300 棉与聚酯纤维弹簧床垫、木制办公桌
Cotton and polyester spring mattress, wooden desk慢速 Slow speed 0.002 931 600 厚重木制品
Heavy woodwork 下载: 导出CSV
下载: 导出CSV
表 2 云南松可燃物参数
Table 2 Forest fuel parameters of P. yunnanensis
参数
Parameter燃烧热
Combustion heat/
(kJ·g−1)参考温度
Reference
temperature/℃密实度
Density/
(kg·m−3)反应热
Reaction heat/
(kJ·g−1)热传导率
Heat conductivity/
(W·km−1)比热容
Specific heat capacity/
(J·kg−1)树干 Trunk 8 260 900 3 0.28 3.0 树叶 Leaf 13.28 260 15 1.5 0.05 3.2
下载: 导出CSV
表 3 模型试运行参数
Table 3 Model testing running parameters
工况
Working condition网格
Mesh/m3火源面积
Fire source area/m2热释放速率
Heat release rate/(kW·m−2)蔓延速度
Spreading rate/(m·min−1)1 0.5 1 4.464 × 104 0.2 2 0.5 12 2.0 × 104 1.0 3 0.5 12 4.6 × 104 8.0
下载: 导出CSV
表 4 模拟工况表
Table 4 Simulated working conditions
工况号
Working condition No.类别
Class工况号
Working condition No.类别
Class工况号
Working condition No.类别
Class工况号
Working condition No.类别
Class1 25°-2.0 m-1.0 m/s 8 25°-6.0 m-2.5 m/s 15 35°-4.5 m-4.5 m/s 22 45°-4.5 m-1.0 m/s 2 25°-2.0 m-2.5 m/s 9 25°-6.0 m-4.5 m/s 16 35°-6.0 m-1.0 m/s 23 45°-4.5 m-2.5 m/s 3 25°-2.0 m-4.5 m/s 10 35°-2.0 m-1.0 m/s 17 35°-6.0 m-2.5 m/s 24 45°-4.5 m-4.5 m/s 4 25°-4.5 m-1.0 m/s 11 35°-2.0 m-2.5 m/s 18 35°-6.0 m-4.5 m/s 25 45°-6.0 m-1.0 m/s 5 25°-4.5 m-2.5 m/s 12 35°-2.0 m-4.5 m/s 19 45°-2.0 m-1.0 m/s 26 45°-6.0 m-2.5 m/s 6 25°-4.5 m-4.5 m/s 13 35°-4.5 m-1.0 m/s 20 45°-2.0 m-2.5 m/s 27 45°-6.0 m-4.5 m/s 7 25°-6.0 m-1.0 m/s 14 35°-4.5 m-2.5 m/s 21 45°-2.0 m-4.5 m/s 注:各工况以坡度(°)-路宽(m)-风速(m/s)标记。Notes: working condition is marked by slope (°)-road width (m)-wind speed (m/s).
下载: 导出CSV
-
[1] 叶兵. 国内外森林防火技术及其发展趋势[D]. 北京: 中国林业科学研究院, 2000. Ye B. The present situation and development tendency of domestic and abroad technical level of forest fire prevention[D]. Beijing: China Academy of Forestry Research Institute of Forestry, 2000.
[2] Picchio P, Pignatti G, Marchi E, et al. The application of two approaches using GIS technology implementation in forest road network planning in an Italian mountain setting[J]. Forests, 2018, 9(5): 277. doi: 10.3390/f9050277
[3] Buday E, Akay A E. Evaluation of forest road network planning in landslide sensitive areas by GIS-based multi-criteria decision making approaches in Ihsangazi Watershed, northern Turkey[J]. Šumarski List, 2019, 143(7−8): 325−336.
[4] Akay O A, Akgul M, Demir M, et al. Analysis of factors associated with the amount of forest road reconstruction activity in Turkey: autoregressive distributed lag modelling approach[J]. Forest Ecology and Management, 2020, 458: 117800.
[5] Keramati A, Lu P, Sobhani A, et al. Impact of forest road maintenance policies on log transportation cost, routing, and carbon-emission trade-offs: oregon case study[J]. Journal of Transportation Engineering, Part A: Systems, 2020, 146(5): 04020028. doi: 10.1061/JTEPBS.0000335
[6] Çalışkan E, Karahalil U. Evaluation of forest road network and determining wood extraction system using GIS: a case study in Anbardağ planning unit[J]. Sumarski List, 2017, 141(3−4): 163−171. doi: 10.31298/sl.141.3-4.6
[7] Petković V, Potočnik I. Planning forest road network in natural forest areas: a case study in northern bosnia and herzegovina[J]. Croatian Journal of Forest Engineering, 2018, 39(1): 45−56.
[8] Tan J. Planning a forest road network by a spatial data handling-network routing system[J]. Acta Forestalia Fennical, 1992, 227: 26−41.
[9] Hrůza P, Mikita T, Tyagur N, et al. Detecting forest road wearing course damage using different methods of remote sensing[J]. Remote Sensing, 2018, 10(4): 492. doi: 10.3390/rs10040492
[10] Fallahchai M M, Haghverdi K, Mojam M S, et al. Ecological effects of forest roads on plant species diversity in Caspian forests of Iran[J]. Acta Ecologica Sinica, 2018, 38(3): 255−261. doi: 10.1016/j.chnaes.2017.08.002
[11] Andrea L, Cristiano F, Fabio F, et al. Forest road planning, construction and maintenance to improve forest fire fighting: a review[J]. Croatian Journal of Forest Engineering, 2019, 40(1): 207−219.
[12] Schöning D R, Bachmann D A, Allgöwer D B. GIS-based framework for wildfire risk assessment final report MINerve2[R]. Zurich:University of Zurich,1997.
[13] 邱荣祖, 方金武, 詹正宜, 等. 林道网理论研究的历史回顾与展望[J]. 福建林学院学报, 2000, 20(4):370−374. doi: 10.3969/j.issn.1001-389X.2000.04.022 Qiu R Z, Fang J W, Zhan Z Y, et a1. A review and prospect for the research of forest road network[J]. Journal of Fujian College of Forestry, 2000, 20(4): 370−374. doi: 10.3969/j.issn.1001-389X.2000.04.022
[14] 罗康. 基于GIS技术的北京市森林防火资源布局的分析[D]. 株洲: 中南林学院, 2003. Luo K. GIS-based layout analysis of forest fire protection in Beijing[D]. Zhuzhou: Central South Forestry College, 2003.
[15] 王国华. 森林地面防火资源调控技术研究[D]. 株洲: 中南林业科技大学, 2009. Wang G H. The technology research of ground-based resources for forest fire control and adjust[D]. Zhuzhou: Central South University of Forestry and Technology, 2009.
[16] 邱荣祖, 邓志华. 林道网的森林火灾防治效益评价[J]. 森林防火, 1998(4):21−22. Qiu R Z, Deng Z H. Evaluation on forest road network of forest fire prevention benefit[J]. Forest Fire Prevention, 1998(4): 21−22.
[17] 张国红. 我国森林防火阻隔系统建设现状与思考[J]. 林业资源管理, 2008(4):15−18. doi: 10.3969/j.issn.1002-6622.2008.04.005 Zhang G H. Present situation and thoughts on the development of the forest fire prevention and obstruction system in China[J]. Forest Resources Management, 2008(4): 15−18. doi: 10.3969/j.issn.1002-6622.2008.04.005
[18] Sun R Y, Jenkins M A, Krueger S K, et al. An evaluation of fire-plume properties simulated with the fire dynamics simulator (FDS) and the Clark coupled wildfire model[J]. Canadian Journal of Forest Research, 2006, 36(11): 2894−2908. doi: 10.1139/x06-138
[19] 尚超, 王克印, 黄海英, 等. 基于的火灾树木建模研究[J]. 消防科学与技术, 2013, 32(9):1030−1033. doi: 10.3969/j.issn.1009-0029.2013.09.030 Shang C, Wang K Y, Huang H Y, et al. Forest trees in fire model research based on the Pyrosim[J]. Fire Science and Tecnnology, 2013, 32(9): 1030−1033. doi: 10.3969/j.issn.1009-0029.2013.09.030
[20] 关祥毅, 刘少刚. 樟子松树冠火燃烧场数学模型的研究[EB/OL]. 北京: 中国科技论文在线(2013−01−25) [2019−09−25]. http://www.paper.edu.cn/releasepaper/content/201301-1040. Guan X Y, Liu S G. Research on the mathematical model of mongolian Scots pine crown fire burning area[EB/OL]. Beijing: Sciencepaper Online (2013−01−25) [2019−09−25]. http://www.paper.edu.cn/releasepaper/content/201301-1040.
[21] 辛喆, 王顺喜, 云峰, 等. 基于火灾模拟软件的草原火灾蔓延规律数值分析[J]. 农业工程学报, 2012, 8(10):54−57. Xin Z, Wang S X, Yun F, et al. Numerical analysis on spreading laws of grassland fire based on fire dynamics simulator (FDS)[J]. Transactions of the Chinese Society of Agricultural Engineering, 2012, 8(10): 54−57.
[22] 李世友. 滇中森林可燃物燃烧性及林火行为研究[D]. 北京: 中国林业科学研究院, 2014. Li S Y. Study on the combustibility of forest fuels and fire behavior in central Yunnan Province[D]. Beijing: China Academy of Forestry Research Institute of Forestry, 2014.
[23] Morvan D, Hoffman C, Rego F, et a1. Numerical simulation of the interaction between two fire fronts in grassland and shrubland[J]. Fire Safety Journal, 2011, 46(8): 469−479. doi: 10.1016/j.firesaf.2011.07.008
[24] Overholt K J, Cabrera J, Kurzawski A, et a1. Characterization of fuel properties and fire spread rates for little bluestem grass[J]. Fire Technology, 2014, 50(1): 9−38. doi: 10.1007/s10694-012-0266-9
[25] Miloua H. Fire behavior characteristics in a pine needle fuel bed in northwest Africa[J]. Journal of Forestry Research, 2019, 30(3): 959−967. doi: 10.1007/s11676-018-0676-8
[26] Mell W E, Manzello S L, Maranghides A. Numerical modeling of fire spread through trees and shrubs[J]. Forest Ecology and Management, 2006, 234(1): S82.
[27] Mell W E, Jenkins M A, Gould J, et a1. A physics-based approach to modeling grassland fires[J]. International Journal of Wildland Fire, 2007, 16(1): 1−22. doi: 10.1071/WF06002
[28] 程远平. 火灾过程中火源热释放速率模型及其实验测试方法[J]. 火灾科学, 2002, 11(2):70−74. doi: 10.3969/j.issn.1004-5309.2002.02.002 Cheng Y P. The model and experimental testing method of heat release rate of fuel during the development of fire[J]. Fire Safety Science, 2002, 11(2): 70−74. doi: 10.3969/j.issn.1004-5309.2002.02.002
[29] Jones W W, Forney G P, Peacock R D, et a1. A technical reference for CFAST: an engineering tool for estimating fire and smoke transport [R]. Gaithersburg: Building and Fire Research Laboratory, 2000.
[30] Jones W W, Peacock R D, Forney G P, et al. Verification and validation of CFAST: a model of fire growth and smoke spread[R]. Washington: National Institute of Standards and Technology, 2004.
[31] Nelson H E. An engineering analysis of the early stages of fire development-the fire at the dupont plaza hotel and casino, December 31, 1986[R/OL].(1987−05−29)[2019−01−22]. https://doi.org/10.6028/NBS.IR.87−3560.
[32] 中华人民共和国林业部.林区公路路线设计规范: LYJ 113—1992 [S].北京: 中国标准出版社, 1997. National Forestry Administration. Design specification of road for forest highway:LYJ 113−1992[S]. Beijing: Standards Press of China,1993.
[33] 张志斌, 杨莹, 张小平, 等. 我国西南地区风速变化及其影响因素[J]. 生态学报, 2014, 34(2):471−481. Zhang Z B, Yang Y, Zhang X P, et a1. Wind speed changes and its influencing factors in southwestern China[J]. Acta Ecologica Sinica, 2014, 34(2): 471−481.
[34] 王秋华. 森林火灾燃烧过程中的火行为研究[D]. 北京: 中国林业科学研究院, 2010. Wang Q H. Study on fire behaviors in forest burning[D]. Beijing: Chinese Academy of Forestry, 2010.
-
期刊类型引用(8)
1. 刘丽,王永康,苏万龙,赵爱玲,任海燕,薛晓芳,石美娟,李登科. 中国酸枣种质资源收集保存、鉴定评价及筛选利用进展. 西北农业学报. 2024(02): 191-200 .  百度学术
百度学术
2. Meng Li,Chenxing Zhang,Lu Hou,Xinru Liu,Hanqing Zhao,Xiaoming Pang,Wenhao Bo,Yingyue Li. Differences in leaf cuticular wax induced by whole-genome duplication in autotetraploid sour jujube. Horticultural Plant Journal. 2024(01): 66-76 . 必应学术
3. 杨亚杰,李昱樱,申状状,陈天,荣二花,吴玉香. 草棉不同倍性材料叶片转录组差异表达分析. 作物学报. 2022(11): 2733-2748 . 百度学术
4. 宋文龙,葛卉,刘兴菊,梁海永. 金叶榆二倍体及其同源四倍体的生理特征与转录组差异分析. 河北农业大学学报. 2022(04): 86-92 . 百度学术
5. 张秀珍,宋文龙,梁海永,王志彬. 四倍体白榆叶片形态特征和光合特性研究. 林业科技. 2022(06): 1-4 . 百度学术
6. 张子璇,谭明谱,王将,张成才,罗丽娜,向增旭. 茅苍术二倍体和同源四倍体叶片转录组比较. 植物资源与环境学报. 2021(04): 41-49 . 百度学术
7. 高真,张成才,罗丽娜,张子璇,王将,向增旭. 茅苍术二倍体及其同源四倍体的生理特征与转录组差异分析. 南京农业大学学报. 2020(06): 1024-1032 . 百度学术
8. 张成才,高真,罗丽娜,梁晖辉,向增旭. 霍山石斛同源四倍体与二倍体活性成分及转录组比较分析. 中国中药杂志. 2020(23): 5669-5676 . 百度学术
其他类型引用(14)










计量
- 文章访问数: 1890
- HTML全文浏览量: 735
- PDF下载量: 128
- 被引次数: 22
