
Biomass allocation and its allometric growth of Pinus yunnanensis seedlings of different families
-
摘要:目的 旨在探明不同家系云南松苗木器官生物量分配格局及其异速生长现象,了解苗木的个体发育规律及适应策略。方法 对10个家系310株云南松苗木的生长及生物量相关指标进行调查,利用单因素方差分析比较不同家系苗木的生物量及其分配差异,并采用标准化主轴分析法对其异速生长关系进行分析。结果 (1)不同家系苗木生物量的积累与分配存在差异,但各器官生物量的分配均表现为叶 > 茎 > 根。苗木个体越小,叶生物量占比越大,随着个体大小的增加,倾向于将更多的生物量分配到茎。(2)苗木器官生物量间及器官生物量与个体大小间的异速生长关系在不同家系中不尽相同,总体表现为等速生长。(3)地上生物量与根生物量间及茎生物量与根生物量间具有共同的异速生长指数,分别为1.054和1.209。不同家系苗木既存在等速生长又具有异速生长现象,异速生长关系并不一致。结论 异速生长关系在不同的家系中并不唯一,既有趋同适应又存在差异,反应了苗木的生长与适应策略。Abstract:Objective This paper aims to reveal the biomass allocation and allometric relationships from different families, and to analyze their ontogenetic law and adaptive strategy of seedlings.Method The growth and biomass indexes of 310 Pinus yunnanensis seedlings from 10 families were investigated. One-way ANOVA was used to compare the differences in biomass and biomass allocation of seedlings from different families, and the allometric relationship was analyzed by standardized major axis analysis.Result (1) The biomass accumulation and allocation were different among varied families, the biomass allocation in each organ was ordered by leaf > stem > root, the biomass allocation of seedlings was constrained by its individual size, and the smaller the individual size was, the larger the leaf biomass ratio was, while the larger the individual size was, the more the stem biomass ratio was. (2) The allometric growth relationships within the plant organ biomass and organ biomass to individual size were quite discrepant, but the growth relationships were isometric overall. (3) The biomass pair aboveground and root biomass, stem and root biomass of seedlings exhibited a common allometric scaling exponent index of 1.054 and 1.209, respectively. The seedlings of different families had both isometric and allometric relationships.Conclusion There is no consistency allometric growth relationship among different families, but it represents the growth and adaptation strategies of seedlings.
-
Keywords:
- Pinus yunnanensis /
- biomass allocation /
- allometric growth
-
生物量分配格局反映了植物的生活史对策,具有重要的生态学意义和进化意义[1-3]。植物在生长发育过程中,会不断调整生长与生物量的分配来适应变化的环境。在短时间内,可能仅表现为根、叶、茎不同器官相对生长速率的差异[4],而长时间内,植物对某一特定环境的适应,会使其形态发生可塑性改变,进而影响植物群落结构,对植物生长产生深远的影响[5-6]。生物量分配模式的量化分析是植物生态学和植物进化史研究的重要组成部分,对于指导农林业生产实践具有重要意义,因此对生物量分配进行量化分析十分有必要。
异速生长是指生物体某两个性状的相对生长速率不同的现象,由物种遗传特性决定,还受环境因子影响[7-8]。从异速生长的角度来讲,植物的资源分配是个体大小依赖的过程,也就是说,异速生长定量地描述了植物生长和资源分配之间的关系[3, 7]。因此,研究植物的异速生长关系及其变化对了解林木的个体发育规律、各器官生长速率及植物的适应性响应机制十分必要。前人对植物的异速生长现象做了大量研究,其中,有研究指出异速生长指数是变化的,且小个体植物的异速生长指数接近1。如Enquist等[9]研究指出,小树苗代谢速率和个体大小之间的异速生长指数为1,随着植物的生长分形网络结构得到完善,异速生长指数变为3/4。Cheng等[10]研究也表明小树苗的异速生长指数为1,较大植物个体为0.82。另外,不同物种的异速生长指数不同,Chen等[11]对两年生苗研究指出,不同物种的异速生长指数不同,甚至同一物种的个体在不同条件下的异速生长指数也有差异。那么对于同一物种不同家系的云南松(Pinus yunnanensis)苗木,异速生长现象又会存在什么变化?根据以上研究,我们有理由推测不同家系云南松苗木的异速生长现象可能会存在差异。
云南松是西南地区荒山造林的先锋树种和主要的用材树种,也是云南省主要的经济树种,具有较高的经济价值和生态效益[12]。之前对异速生长现象的研究主要涉及生境响应、繁殖策略及种间及种内竞争等方面,其中又以密度效应、叶内性状变化研究居多[13-16]。目前,对云南松异速生长的研究只是基于不同器官[17]、不同地理种源[18],而对其不同家系内异速生长现象的研究涉猎较少,我们对云南松苗木不同家系的异速生长现象理解尚不完全。因此,本文以不同家系云南松苗木为研究对象,探明其异速生长现象。这有利于揭示不同家系云南松苗木的各器官生长速率以及生活史策略,同时也可丰富云南松异速生长现象研究的内容。
1. 研究区概况与研究方法
1.1 研究区概况
试验地设置在西南林业大学苗圃(102°45′41″E、25°04′00″N),海拔1 945 m,属北亚热带半湿润高原季风气候,年平均气温14.7 ℃,绝对最低温−9 ℃,绝对最高温32.5 ℃。年降水量700 ~ 1 100 mm,全年降水量在时间分布上明显地分为干、湿两季,5—10月为雨季,降水量占全年的85%左右,11月至次年4月为干季,降水量仅占全年的15%左右,年平均相对湿度68.2%。全年无霜期近年来均在240 d以上,全年晴天较多,日照数年均2 445.6 h,日照率56%。终年太阳投射角度大,年均总辐射量达543.24 kJ/cm2。
1.2 种子采集与育苗管理
1.2.1 种子采集
2014年12月在云南省昆明市宜良县禄丰村林场从10株优良母树上采集不同家系云南松的种子。采集地位于103°00′02″ ~ 103°16′06″E,24°26′11″ ~ 24°30′08″N,海拔为1 270 ~ 2 583 m,属亚热带季风气候,年均气温17.5 ℃,绝对最高温31.5 ℃,绝对最低温−5 ℃,年均降水量995.3 mm,年相对湿度68%左右。采集的林分为云南松天然林,混生少量华山松(Pinus armandii) 、滇油杉(Keteleeria evelyniana) 。采种母树的平均胸径约22 cm,树高约13 m,林冠下有极少量灌木,林地有云南松凋落的针叶覆盖,厚0.5 ~ 3.0 cm,林分郁闭度在0.6以上。所选采种母树为树干通直圆满、结实状况良好且无病虫害、林龄25年以上的单株。采种母树间距100 m左右,采集树冠中上部饱满的当年生成熟球果,每个优树采集20个以上,采集后按照家系进行归类编号,10个家系的编号分别为2008、2017、2020、2024、2033、2034、2036、2048、2050、2206。将分类编号完备的球果带回实验室晾晒、风干,待球果开裂后,按单株取出种子,并去除杂质及废种,用于后期播种观测研究。
1.2.2 育苗管理
2015年3月将云南松10个家系的种子在25 ℃温水中浸泡24 h,播种前用0.5%高锰酸钾喷洒床面,进行土壤消毒。用点播的方式播种于苗床,苗床规格为1 m × 30 m,株行距为5 cm × 10 cm,苗期试验采用单因素完全随机区组设计,共3个区组,每个区组10个小区,每个小区播40粒种子。播种后,盖薄膜小拱棚,不定期浇水,待苗出齐后,逐渐间苗,之后进行露天培育,及时除去杂草,适时浇水。苗期均采用相同的常规方法进行管理,保证圃内环境条件基本一致。
1.3 指标测定
2016年12月,用整株收获法挖取苗圃中10个家系云南松苗木,各家系苗木(2008 ~ 2206)的样本量分别为33、33、35、25、26、27、37、30、36、28,共计310株。然后用游标卡尺测量每株苗木的地径(D),直尺测定每株苗木苗高(H),分别精确至0.01 mm、0.01 cm。用水小心冲洗根系泥土,保持根系完整,放在阴凉处风干。用电子天平称量每株苗木根、茎、叶的鲜质量,分别装入标记好的纸袋中,在105 ℃的烘箱中杀青30 min后,调至80 ℃进行烘干处理直至质量恒定,分别测量根(地下部分,RB)、茎(SB)、叶(LB)的生物量,精确至0.001 g。并分别计算地上生物量(AGB)、总生物量(TB)、各器官生物量分配比例、地上生物量分配比例等。
1.4 数据分析
在确定异速生长关系前,明确对个体大小的定义。尽管株高、胸径和种子大小等一些指标均可表征植物个体大小。但多数研究者认为生物量是更为本质或更为准确的一个表征指标,其中又以总生物量最具代表性[19-22],因此用总生物量(TB)表征个体大小。异速生长关系可表示为Y = βXα,其中,Y是某种生物学特征或者功能,β是标准化常数,X指个体大小(总生物量,TB),α是异速生长指数,α = 1为等速关系,α ≠ 1为异速关系。确定异速生长参数时,须将幂函数线性转化为log2Y = log2β + α·log2X。采用标准主轴回归分析(standardized major axis,SMA)获取异速生长方程的参数估计[14, 18]。计算异速生长方程的斜率α,并比较斜率之间以及各斜率与1.0的差异性。若斜率间无显著差异,给出共同斜率,则需要进一步比较它们的截距是否相同,并进一步进行共轴漂移检验,即判断它们是否具有同一的线性拟合轴,采用Wald方法检验不同家系苗木沿共同主轴位移差异的显著性,计算位移量。上述异速生长指数(α)和截距(log2β)的计算、截距漂移检验等均采用R软件的smatr包进行。所有统计检验的显著水平均采用α = 0.05[16]。株高 ~ 地径的异速生长关系常用弹性相似模型、几何相似模型、压力相似模型描述,相应的异速生长指数分别为2/3、1、1/2,根据异速生长指数置信区间是否包含2/3、1和1/2判断株高 ~ 地径的异速生长关系符合哪种模型[23]。
2. 结果与分析
2.1 不同家系苗木生物量的积累与分配
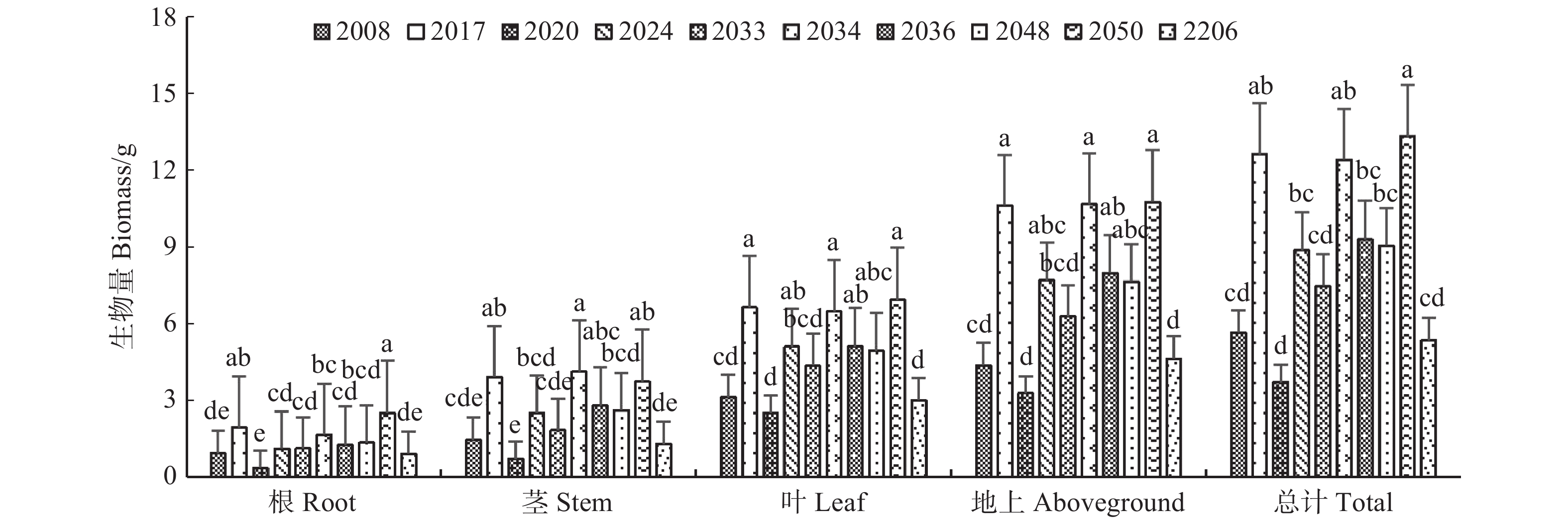
云南松苗木各器官生物量积累与分配在不同家系间差异显著(P < 0.05)(图1、图2)。在生物量积累方面,家系2017、2034、2050的总生物量较大,即个体较大,家系2008、2020、2206相对较小。茎的生物量积累量最大的为家系2034,而根、叶、地上生物量积累量均表现为家系2050最大,2020最小。生物量分配比表现为:根的生物量分配比为家系2050最大,2020最小,地上生物量反之;茎的生物量分配比为2034最大,2020最小,叶生物量分配反之。不同家系云南松苗木生物量的积累与分配比总体表现为:叶生物量 > 茎生物量 > 根生物量,地上部分 > 根(地下部分)。综合看来,云南松苗木将较多的生物量投资和分配到叶和茎上,而投资分配给根的生物量较少,其生长主要侧重于地上部分生长,干物质主要储存在叶、茎中。苗木个体越小,叶所占的生物量比例越大,反之亦然。
![]() 图 1 不同家系云南松苗木生物量积累量生物量积累量的值为平均值± 标准误。不同小写字母表示家系间差异显著(P < 0.05)。下同。Biomass accumulation value was mean ± standard error. Different lowercase letters indicate significant differences among varied families (P < 0.05). The same below.Figure 1. Biomass accumulation of P. yunnanensis seedlings of different families
图 1 不同家系云南松苗木生物量积累量生物量积累量的值为平均值± 标准误。不同小写字母表示家系间差异显著(P < 0.05)。下同。Biomass accumulation value was mean ± standard error. Different lowercase letters indicate significant differences among varied families (P < 0.05). The same below.Figure 1. Biomass accumulation of P. yunnanensis seedlings of different families![]() 图 2 不同家系云南松苗木生物量分配比例Figure 2. Biomass allocation ratios of P. yunnanensis seedlings of different families
图 2 不同家系云南松苗木生物量分配比例Figure 2. Biomass allocation ratios of P. yunnanensis seedlings of different families2.2 形态指标及形态指标与个体大小间的异速生长
在不区分家系的情况下(表1),苗高 ~ 地径及地径 ~ 个体大小之间的异速生长指数均显著小于1.0(P < 0.05),表现为异速生长。这表明在不考虑家系差异的情况下,云南松苗木生物量的累积速率大于地径生长速率和苗高生长速率,但苗高 ~ 地径间的异速生长关系不符合压力相似模型、几何相似模型、弹性相似模型中的任何一个。
表 1 云南松苗木不同性状间的异速生长分析Table 1. Allometric growth analysis of P. yunnanensis seedlings with different traits性状
Trait决定系数
Determination coefficient显著性
Significance (P)斜率
Slope95%置信区间
95% confidence interval截距
Intercept95%置信区间
95% confidence intervalF值
F valueP−1.0 H ~ D 0.253 0.000 0.962 0.873 ~ 1.060 0.198 0.110 ~ 0.624 0.430 0.000 D ~ TB 0.666 0.000 0.524 0.491 ~ 0.559 0.503 0.473 ~ 0.533 442.845 0.000 LB ~ RB 0.755 0.000 0.996 0.942 ~ 1.052 0.563 0.541 ~ 0.585 0.026 0.873 LB ~ SB 0.757 0.000 0.854 0.808 ~ 0.903 0.380 0.356 ~ 0.403 31.786 0.000 SB ~ RB 0.771 0.000 1.165 1.105 ~ 1.230 0.215 0.190 ~ 0.239 31.794 0.000 AGB ~ RB 0.817 0.000 1.002 0.955 ~ 1.051 0.735 0.716 ~ 0.753 0.004 0.948 RB ~ TB 0.870 0.000 1.016 0.976 ~ 1.058 −0.825 −0.862 ~ 0.789 0.603 0.438 SB ~ TB 0.898 0.000 1.184 1.143 ~ 1.227 −0.747 0.803 ~ 1.000 87.296 0.000 LB ~ TB 0.948 0.000 1.011 0.986 ~ 1.038 −0.259 −0.282 ~ 0.236 0.767 0.382 AGB ~ TB 0.994 0.000 1.018 1.009 ~ 1.027 −0.092 −0.100 ~ 0.084 15.722 0.000 注:P−1.0表示斜率与理论值1.0的差异显著性。H、D、RB、SB、LB、AGB、TB分别为苗高、地径、根生物量、茎生物量、叶生物量、地上生物量、总生物量(个体大小)。下同。
Notes: P−1.0 indicates significant difference between the estimated model slope and theoretical value 1.0; H, D, RB, SB, LB, AGB, TB are seedling height, ground diameter, root biomass, stem biomass, leaf biomass, aboveground biomass and total biomass (individual size), respectively. The same below.就不同家系而言,苗高 ~ 地径及地径 ~ 个体大小间总体上具有显著或极显著的异速生长关系(P < 0.05),异速生长指数在不同家系间差异显著(P < 0.05)(表2)。苗高 ~ 地径的回归斜率在0.678 ~ 1.483间波动。其中家系2020、2036、2048的异速生长指数与1.0差异显著,表现为异速生长。其余家系的异速生长指数与1.0均无显著差异,表现为等速生长。在表现为异速生长的家系中,2020、2036为显著小于1.0的异速生长,符合弹性相似模型。而2048为显著大于1.0的异速生长关系,说明家系2020、2036地径的生长速率要大于苗高的生长速率,而家系2048则反之。地径 ~ 个体大小间的斜率在0.354 ~ 0.832范围内变化。家系2008表现为等速生长,其余家系均表现为异速生长,即对于绝大多数家系而言,地径的生长速率小于生物量的累积速率。
表 2 不同家系云南松苗木形态指标及形态指标与个体大小间的异速生长分析Table 2. Allometric growth analysis of morphological index and the relationship between morphological index and individual size of P. yunnanensis seedlings性状
Trait家系
Family决定系数
Determination coefficientP 斜率
Slope95%置信区间
95% confidence intervalF值
F valueP−1.0 类型
TypeH ~ D 2008 0.002 0.825 0.728b 0.509 ~ 1.042 3.227 0.082 I 2017 0.403 0.000 0.919b 0.695 ~ 1.215 0.372 0.546 I 2020 0.465 0.000 0.770b 0.596 ~ 0.995 4.300 0.046 A 2024 0.193 0.028 1.168ab 0.800 ~ 1.705 0.690 0.415 I 2033 0.138 0.062 1.006ab 0.686 ~ 1.474 0.001 0.975 I 2034 0.266 0.006 0.941ab 0.666 ~ 1.330 0.125 0.726 I 2036 0.474 0.000 0.678b 0.530 ~ 0.867 10.595 0.003 A 2048 0.359 0.000 1.483a 1.093 ~ 2.013 7.142 0.012 A 2050 0.366 0.000 0.990ab 0.753 ~ 1.303 0.005 0.944 I 2206 0.215 0.013 1.043ab 0.735 ~ 1.480 0.058 0.812 I D ~ TB 2008 0.406 0.000 0.832a 0.630 ~ 1.100 1.785 0.191 I 2017 0.577 0.000 0.354b 0.280 ~ 0.448 111.926 0.000 A 2020 0.753 0.000 0.506b 0.425 ~ 0.603 72.119 0.000 A 2024 0.555 0.000 0.442b 0.333 ~ 0.587 42.892 0.000 A 2033 0.754 0.000 0.473b 0.385 ~ 0.582 65.659 0.000 A 2034 0.884 0.000 0.406b 0.353 ~ 0.466 229.341 0.000 A 2036 0.826 0.000 0.485b 0.420 ~ 0.559 125.305 0.000 A 2048 0.712 0.000 0.432b 0.351 ~ 0.531 86.261 0.000 A 2050 0.862 0.000 0.410b 0.360 ~ 0.466 253.696 0.000 A 2206 0.588 0.000 0.514b 0.398 ~ 0.663 32.407 0.000 A 注:不同小写字母表示家系间差异显著 (P < 0.05)。A表示异速生长关系,I表示等速生长关系。下同。Notes: Different lowercase letters indicate significant differences among varied families (P < 0.05). A means allometric growth relationship, I indicates isometric growth relationship. The same below. 2.3 器官生物量间的异速生长
在不区分家系的情况下(表1),叶 ~ 根、地上 ~ 根间的异速生长指数与1.0无显著差异,为等速生长。而茎 ~ 根、叶 ~ 茎间的异速生长指数与1.0具有显著差异(P < 0.05),表现为异速生长。其中,LB ~ SB为显著小于1.0的异速生长,茎 ~ 根表现为显著大于1.0的异速生长。这表明茎生物量的累积速率大于叶、根生物量的累积速率,且与家系无关。
就不同家系而言,云南松苗木各器官生物量间存在极显著的异速生长关系(P < 0.01)(表3)。叶与根、茎之间的异速生长指数在不同家系间差异显著(P < 0.05),但茎 ~ 根、地上 ~ 根间无显著差异。叶 ~ 根间除家系2024、2033、2048与1.0有显著差异,表现为异速生长外,其余家系与1.0无显著差异,表现为等速生长。在存在异速生长的家系中,2024和2033表现为显著大于1.0的异速生长,2048反之。即家系2024和2033叶生物量的累积速率大于根生物量的累积速率,家系2048则相反。叶 ~ 茎间家系2017、2036、2048、2050与1.0具有显著差异,表现为异速生长,其余家系与1.0均无显著,表现为异速生长。SB ~ RB之间家系2017、2020、2036、2048、2050与1.0具有显著差异,其余家系与1.0无显著差异,且均为显著大于1.0的异速生长,说明茎生物量的累积速率大于根生物量。地上 ~ 根之间除家系2024、2033与1.0具有显著差异外,其余家系与1.0无显著差异。由此可以看出,对于各器官生物量间的异速生长关系,绝大多数家系表现为等速生长。
表 3 不同家系云南松苗木各器官生物量间的异速生长分析Table 3. Analysis of allometric growth of each organ of P. yunnanensis seedlings among different families性状
Trait家系
Family决定系数
Determination coefficientP 斜率
Slope95%置信区间
95% confidence intervalF值
F valueP−1.0 类型
TypeLB ~ RB 2008 0.729 0.000 1.017b 0.842 ~ 1.230 0.034 0.856 I 2017 0.814 0.000 1.033b 0.883 ~ 1.210 0.178 0.676 I 2020 0.801 0.000 1.168b 0.998 ~ 1.366 4.023 0.053 I 2024 0.538 0.000 1.658a 1.242 ~ 2.215 13.856 0.001 A 2033 0.808 0.000 1.298ab 1.080 ~ 1.559 8.690 0.007 A 2034 0.850 0.000 1.090b 0.930 ~ 1.277 1.237 0.277 I 2036 0.666 0.000 1.109b 0.911 ~ 1.351 1.128 0.295 I 2048 0.900 0.000 0.857b 0.759 ~ 0.969 6.670 0.015 A 2050 0.854 0.000 0.894b 0.783 ~ 1.021 2.946 0.095 I 2206 0.639 0.000 1.027b 0.808 ~ 1.305 0.050 0.826 I LB ~ SB 2008 0.756 0.000 0.902b 0.753 ~ 1.079 1.369 0.251 I 2017 0.725 0.000 0.774b 0.639 ~ 0.937 7.557 0.010 A 2020 0.849 0.000 1.010ab 0.880 ~ 1.158 0.020 0.889 I 2024 0.643 0.000 1.238a 0.959 ~ 1.600 2.973 0.098 I 2033 0.688 0.000 1.092ab 0.865 ~ 1.379 0.600 0.446 I 2034 0.872 0.000 0.995ab 0.859 ~ 1.152 0.006 0.941 I 2036 0.583 0.000 0.794b 0.637 ~ 0.989 4.554 0.040 A 2048 0.879 0.000 0.656b 0.574 ~ 0.750 43.564 0.000 A 2050 0.892 0.000 0.733b 0.654 ~ 0.822 31.413 0.000 A 2206 0.649 0.000 1.061ab 0.837 ~ 1.344 0.256 0.617 I SB ~ RB 2008 0.652 0.000 1.128a 0.910 ~ 1.398 1.303 0.262 I 2017 0.714 0.000 1.335a 1.099 ~ 1.622 9.301 0.005 A 2020 0.833 0.000 1.157a 1.001 ~ 1.336 4.207 0.048 A 2024 0.500 0.000 1.340a 0.992 ~ 1.810 4.051 0.056 I 2033 0.699 0.000 1.188a 0.945 ~ 1.494 2.386 0.136 I 2034 0.786 0.000 1.096a 0.907 ~ 1.324 0.977 0.333 I 2036 0.654 0.000 1.397a 1.143 ~ 1.707 11.746 0.002 A 2048 0.862 0.000 1.307a 1.132 ~ 1.508 14.864 0.001 A 2050 0.807 0.000 1.220a 1.048 ~ 1.421 7.064 0.012 A 2206 0.728 0.000 0.968a 0.786 ~ 1.193 0.101 0.753 I AGB ~ RB 2008 0.746 0.000 1.013a 0.843 ~ 1.217 0.020 0.889 I 2017 0.851 0.000 1.049a 0.911 ~ 1.207 0.474 0.496 I 2020 0.833 0.000 1.142a 0.989 ~ 1.320 3.528 0.069 I 2024 0.589 0.000 1.416a 1.077 ~ 1.860 7.041 0.014 A 2033 0.837 0.000 1.199a 1.012 ~ 1.421 4.902 0.037 A 2034 0.862 0.000 1.070a 0.919 ~ 1.241 0.829 0.371 I 2036 0.769 0.000 1.102a 0.935 ~ 1.299 1.423 0.241 I 2048 0.907 0.000 0.974a 0.866 ~ 1.096 0.203 0.656 I 2050 0.862 0.000 0.977a 0.858 ~ 1.111 0.139 0.712 I 2206 0.748 0.000 0.947a 0.774 ~ 1.157 0.311 0.582 I 不同家系的茎 ~ 根、地上 ~ 根之间具有共同斜率1.209和1.054,均显著大于理论值1.0(P < 0.05)。在共同斜率下,茎 ~ 根、地上 ~ 根的截距、截距漂移在不同家系间具有显著差异,具体表现为:家系2050沿共同主轴的位移量最大(0.759、1.221),2020最小(−0.845、−0.107)(表4)。这表明当根生物量一定时,即有限资源下,家系2050对茎、地上生物量的投入相对较大。
表 4 不同家系云南松苗木茎 ~ 根、地上 ~ 根生物量沿共同主轴的位移Table 4. Shift of biomass pair SB − RB and AGB − RB of seedlings along common axis of P. yunnanensis seedlings among different families家系
FamilySB ~ RB AGB ~ RB 截距
Intercpt95%置信区间
95% confidence interval漂移
Shift截距
Intercept95%置信区间
95% confidence interval漂移
Shift2008 0.172b 0.095 ~ 0.238 −0.007b 0.675b 0.619 ~ 0.726 0.520b 2017 0.191b 0.059 ~ 0.283 0.574ab 0.718b 0.657 ~ 0.781 1.052ab 2020 0.313ab 0.193 ~ 0.383 −0.845b 0.902a 0.851 ~ 1.039 −0.107b 2024 0.275ab 0.161 ~ 0.390 0.268b 0.761b 0.655 ~ 0.870 0.755b 2033 0.180b 0.103 ~ 0.256 0.161b 0.722b 0.667 ~ 0.779 0.706b 2034 0.337a 0.254 ~ 0.433 0.472ab 0.771b 0.701 ~ 0.840 0.889ab 2036 0.266b 0.182 ~ 0.327 0.408b 0.765b 0.716 ~ 0.808 0.888b 2048 0.190b 0.125 ~ 0.248 0.277b 0.735b 0.701 ~ 0.775 0.811b 2050 0.065b −0.021 ~ 0.144 0.759a 0.616b 0.582 ~ 0.694 1.221a 2206 0.187b 0.083 ~ 0.226 −0.146b 0.685b 0.603 ~ 0.737 0.394b 2.4 器官生物量与个体大小间的异速生长
在不区分家系的情况下(表1),根、叶与个体大小之间的异速指数与1.0无显著差异,表现为等速生长。茎、地上与个体大小的异速指数与1.0具有显著差异,表现为显著大于1.0的异速生长,表明在不考虑家系差异的情况下,云南松苗木茎、地上生物量的累积速率大于总生物量的累积速率。
就不同家系而言,各器官生物量与个体大小间具有极显著的异速生长关系(P < 0.01)(表5)。除根与个体大小在各家系间无显著差异外(P > 0.05),其余3对性状均在不同家系间存在显著差异(P < 0.05)。茎、叶与个体大小间,5个家系表现为异速生长,5个表现为等速生长。根与个体大小间,家系2024表现为异速生长,其余家系表现为等速生长。地上与个体大小间,家系2020、2024、2033、2036的异速生长指数与1.0存在显著差异,表现为异速生长,其余家系的异速生长指数与1.0均无显著差异,表现为等速生长。即根、地上与个体大小间,多数家系表现为等速生长。
表 5 不同家系云南松苗木器官生物量与个体大小间的异速生长分析Table 5. Analysis of allometric growth between each organ and individual size of P. yunnanensis seedlings among different families性状
Trait家系
Family决定系数
Determination coefficientP 斜率
Slope95%置信区间
95% confidence intervalF值
F valueP−1.0 类型
TypeRB 2008 0.823 0.000 1.015a 0.870 ~ 1.183 0.038 0.848 I 2017 0.892 0.000 0.974a 0.863 ~ 1.098 0.201 0.657 I 2020 0.868 0.000 0.901a 0.793 ~ 1.025 2.713 0.109 I 2024 0.703 0.000 0.785a 0.622 ~ 0.991 4.626 0.042 A 2033 0.877 0.000 0.876a 0.756 ~ 1.015 3.428 0.076 I 2034 0.895 0.000 0.953a 0.834 ~ 1.088 0.557 0.463 I 2036 0.818 0.000 0.933a 0.806 ~ 1.079 0.936 0.340 I 2048 0.935 0.000 1.029a 0.932 ~ 1.136 0.355 0.556 I 2050 0.910 0.000 1.034a 0.928 ~ 1.144 0.345 0.561 I 2206 0.819 0.000 1.069a 0.901 ~ 1.267 0.632 0.434 I SB 2008 0.874 0.000 1.145b 1.006 ~ 1.303 4.539 0.041 A 2017 0.851 0.000 1.300ab 1.129 ~ 1.497 14.690 0.001 A 2020 0.918 0.000 1.042b 0.942 ~ 1.153 0.692 0.412 I 2024 0.812 0.000 1.052b 0.873 ~ 1.267 0.316 0.580 I 2033 0.831 0.000 1.041b 0.876 ~ 1.237 0.229 0.636 I 2034 0.929 0.000 1.044b 0.936 ~ 1.165 0.652 0.427 I 2036 0.898 0.000 1.303ab 1.168 ~ 1.453 24.707 0.000 A 2048 0.947 0.000 1.345a 1.230 ~ 1.470 47.752 0.000 A 2050 0.941 0.000 1.258ab 1.155 ~ 1.369 30.720 0.000 A 2206 0.856 0.000 1.034b 0.888 ~ 1.205 0.206 0.654 I LB 2008 0.963 0.000 1.032b 0.962 ~ 1.107 0.851 0.363 I 2017 0.961 0.000 1.006b 0.936 ~ 1.081 0.031 0.861 I 2020 0.985 0.000 1.052b 1.007 ~ 1.099 5.614 0.024 A 2024 0.932 0.000 1.302a 1.164 ~ 1.457 24.195 0.000 A 2033 0.965 0.000 1.137b 1.051 ~ 1.230 11.446 0.002 A 2034 0.984 0.000 1.039b 0.987 ~ 1.093 2.294 0.142 I 2036 0.837 0.000 1.034b 0.901 ~ 1.188 0.246 0.623 I 2048 0.981 0.000 0.882b 0.837 ~ 0.930 23.824 0.000 A 2050 0.983 0.000 0.921b 0.880 ~ 0.965 13.080 0.001 A 2206 0.924 0.000 1.097b 0.982 ~ 1.226 2.942 0.098 I AGB 2008 0.991 0.000 1.028b 0.992 ~ 1.064 2.538 0.121 I 2017 0.996 0.000 1.021b 0.998 ~ 1.045 3.515 0.070 I 2020 0.997 0.000 1.030b 1.011 ~ 1.048 10.826 0.002 A 2024 0.984 0.000 1.111a 1.052 ~ 1.174 16.122 0.001 A 2033 0.996 0.000 1.051ab 1.025 ~ 1.078 16.631 0.000 A 2034 0.997 0.000 1.019b 0.998 ~ 1.041 3.466 0.074 I 2036 0.996 0.000 1.028b 1.006 ~ 1.050 6.757 0.014 A 2048 0.997 0.000 1.003b 0.983 ~ 1.023 0.084 0.774 I 2050 0.994 0.000 1.007b 0.980 ~ 1.034 0.249 0.621 I 2206 0.992 0.000 1.011b 0.976 ~ 1.048 0.424 0.521 I 3. 讨 论
3.1 生物量分配
植物生物量分配是遗传差异与环境共同作用的结果,是植物对环境适应的体现。本文研究结果显示,不同家系云南松苗木的生物量分配存在差异,在相对一致的生境与管理措施下,相同生长期内呈现不同的生物量分配格局,这可能与自身的遗传差异有关[24]。生物量异速分配理论认为,植物个体较小时,更注重叶资源的分配,随着个体增大,倾向于把资源分配到茎、根中,最终趋于平衡[25-26]。最优分配理论认为,植物在某一资源成为限制性资源时,会优先将代谢产物分配给可以获得限制性资源的器官。如光照受限时,生物量会优先分配到叶与茎中。养分或水分受限时,则优先分配到根[27-28]。本研究发现,不同家系云南松苗木各器官生物量的分配表现为叶 > 茎 > 根,且苗木个体越小,叶生物量占比越大,这与王晨等[29]的研究结果类似。从异速分配的角度来说,两年生苗木可能个体相对较小,因此将更多的资源分配给叶,以制造更多的有机物质来满足自身生长的需要。另外,苗圃内水分与养分相对充足,受限的因子主要是光照,苗木可能将较多的资源分配到叶与茎,以增强苗木对光照以及空间的竞争能力。何怀江等[28]对12个乔木树种的生物量进行研究,表明最优分配理论与异速生长理论彼此互补、并不独立。王晨等[29]也曾揭示异速分配理论和最优分配理论在解释毛竹(Phyllostachys edulis)生物量分配方面几乎是互为补充的,而非独立。就本文而言,两种理论似乎也是互补的,最优分配理论与异速分配理论可同时解释苗木的生物量分配格局。另外,生物量分配在一定程度上还受到密度的制约。密度会改变植物生物量的分配比例,但幼苗的生物量分配对密度的响应规律并不一致,一般认为由物种的基因型决定[30]。一些学者研究指出密度对生物量分配的影响并不大,如朱仕明等[31]对不同密度条件下乐昌含笑(Michelia chapensis)幼苗生物量进行研究,发现密度对乐昌含笑各器官生物量分配的影响不大。高平珍等[32]的研究表明,苦参(Sophotora flavescens)幼苗的各生物量分配指标在3种立竹密度试验下均无显著差异。冯银平等[33]的研究也表明种植密度对苜蓿(Medicago sativa)生物量分配并无显著影响,不同密度下苜蓿种群的根冠比、叶茎比之间均无显著差异。因此,密度对生物量分配的制约可能因物种而异,一些物种对密度效应可能并不敏感。鉴于上述理由,本文在对不同家系苗木生物量分配进行分析时,把不同家系苗木的样本量视为近似相等,并未过多阐释密度效应对生物量分配的影响。
3.2 异速生长关系
异速生长关系是指生物体的某些生物学特征(如个体形态指标)与个体大小(如质量)之间的幂函数关系[34-35]。树高 ~ 地径异速生长关系是树木面对不同环境压力采取的最普遍的生态对策,也是树木对环境变化最敏感的关系之一[36]。外界环境发生变化时,树木往往会通过物质和能量调控改变树高 ~ 地径的异速生长关系,从而调节形态结构,逐渐形成有利于自身生存的结构特征[37]。本文研究结果显示,不同家系云南松苗木苗高 ~ 地径间总体上表现为等速生长,符合几何相似模型。地径与个体大小间,绝大多数家系表现为显著小于1.0的异速生长。但是云南松苗木不同家系具有不同的异速生长指数,且这些异速指数不能同时符合弹性、应力及几何自相似模型,与胡波等[37]、赵广等[38]的研究结果相似。说明不同家系云南松苗木并没有完全一致的生长速率与分配速率,即植物属性间的异速关系具有一定的物种特异性。研究还发现在不考虑家系差异的情况下,生物量的累积速率大于地径的增长速率,这与江洪等[17]对云南松异速生长现象的初步研究结果类似。另外,云南松苗木地径生长速率大于苗高生长速率,这可能是与云南松苗木的“蹲苗”现象有关。1 ~ 3年生云南松苗木普遍存在蹲苗现象,这个阶段主要投资地径生长,导致地径的生长速率要快于苗高[17, 24]。
植物不同器官间的生长关系是普遍承认的生长规律,各器官的生长存在权衡关系[39]。本研究对云南松不同器官之间的异速关系进行研究发现,不同家系苗木各器官生物量间存在极显著的相关关系。叶与根、茎之间的回归斜率在不同家系间差异显著,但茎 ~ 根、地上 ~ 根之间无显著差异。总体而言,各器官生物量间绝大多数家系表现为等速生长。生态代谢理论提出了生物量在根、茎、叶之间分配的异速模型,预测对于非木本植物或缺乏次生组织的小个体木本植物(主要是指一年生幼苗),其根、茎、叶生物量间、根、茎、叶各器官与总生物量间均为等速生长关系[34-35, 40-41]。本文的研究结果符合生态代谢理论预测,各器官生物量间绝大多数家系表现为等速生长,表明生物量在三大器官间的分配关系出现了趋同进化。地上 ~ 根、茎 ~ 根之间具有共同的异速生长指数,这也是家系间趋同适应的表现。但不同家系苗木既存在等速生长又存在异速生长,异速生长关系不稳定,即没有出现完全一致的异速生长关系,这可能与遗传差异有关,每个家系苗木具有各自独特的生物学特性,致使生长类型不唯一[41]。
个体大小作为生物体的基本性状之一,其生长发育影响着形态特征、生长速率、代谢速率和繁殖能力等一系列的生物体性状,是植物适应能力大小的反应[42]。本文研究发现,各器官生物量与个体大小总体上表现为等速生长,符合理论预测。云南松不同家系各器官生物量与个体大小间具有极显著的相关关系(P < 0.01),说明个体大小显著影响着苗木生物量的分配,亦或生物量分配存在显著的个体发育漂变。其中茎与个体大小之间异速指数最大,表明随着个体大小的增加,会将更多的生物量分配到茎,反映了苗木灵活的生长与分配策略,这与生物量分配结果研究相吻合。除根生物量与个体大小在各家系间无显著差异外,其余器官与个体大小间在不同家系间存在显著差异,说明苗木在生长过程中并未过多的改变根生物量分配,根系是植物幼苗苗期主要的营养物质吸收、传送器官,在苗木的整个生长发育过程中起着重要作用,因此不会过多的改变根生物量分配。
4. 结 论
总之,由于苗木个体遗传特性的差异,云南松苗木的生长特性、生物量分配格局及异速生长关系在不同家系间存在较大差异,即异速关系具有物种特异性,但不同家系苗木器官特征间具有相同的生长速率与协同变化特征,在一定程度上说明了尽管物种进化中在发育、形态等方面出现了多样性分化,但长期生活在相同环境下,形成了相似的生物学特征,体现了苗木对环境的趋同适应;个体较小的苗木具有相对较高的叶生物量分配,个体较大的苗木具有较高的茎生物量分配,表明苗木以不同的生物量分配策略来响应个体大小的变化,这显示了其灵活的生长和资源分配策略。
-
![]()
图 1 不同家系云南松苗木生物量积累量
生物量积累量的值为平均值± 标准误。不同小写字母表示家系间差异显著(P < 0.05)。下同。Biomass accumulation value was mean ± standard error. Different lowercase letters indicate significant differences among varied families (P < 0.05). The same below.
Figure 1. Biomass accumulation of P. yunnanensis seedlings of different families
![]()
图 2 不同家系云南松苗木生物量分配比例
Figure 2. Biomass allocation ratios of P. yunnanensis seedlings of different families
表 1 云南松苗木不同性状间的异速生长分析
Table 1 Allometric growth analysis of P. yunnanensis seedlings with different traits
性状
Trait决定系数
Determination coefficient显著性
Significance (P)斜率
Slope95%置信区间
95% confidence interval截距
Intercept95%置信区间
95% confidence intervalF值
F valueP−1.0 H ~ D 0.253 0.000 0.962 0.873 ~ 1.060 0.198 0.110 ~ 0.624 0.430 0.000 D ~ TB 0.666 0.000 0.524 0.491 ~ 0.559 0.503 0.473 ~ 0.533 442.845 0.000 LB ~ RB 0.755 0.000 0.996 0.942 ~ 1.052 0.563 0.541 ~ 0.585 0.026 0.873 LB ~ SB 0.757 0.000 0.854 0.808 ~ 0.903 0.380 0.356 ~ 0.403 31.786 0.000 SB ~ RB 0.771 0.000 1.165 1.105 ~ 1.230 0.215 0.190 ~ 0.239 31.794 0.000 AGB ~ RB 0.817 0.000 1.002 0.955 ~ 1.051 0.735 0.716 ~ 0.753 0.004 0.948 RB ~ TB 0.870 0.000 1.016 0.976 ~ 1.058 −0.825 −0.862 ~ 0.789 0.603 0.438 SB ~ TB 0.898 0.000 1.184 1.143 ~ 1.227 −0.747 0.803 ~ 1.000 87.296 0.000 LB ~ TB 0.948 0.000 1.011 0.986 ~ 1.038 −0.259 −0.282 ~ 0.236 0.767 0.382 AGB ~ TB 0.994 0.000 1.018 1.009 ~ 1.027 −0.092 −0.100 ~ 0.084 15.722 0.000 注:P−1.0表示斜率与理论值1.0的差异显著性。H、D、RB、SB、LB、AGB、TB分别为苗高、地径、根生物量、茎生物量、叶生物量、地上生物量、总生物量(个体大小)。下同。
Notes: P−1.0 indicates significant difference between the estimated model slope and theoretical value 1.0; H, D, RB, SB, LB, AGB, TB are seedling height, ground diameter, root biomass, stem biomass, leaf biomass, aboveground biomass and total biomass (individual size), respectively. The same below. 下载: 导出CSV
下载: 导出CSV
表 2 不同家系云南松苗木形态指标及形态指标与个体大小间的异速生长分析
Table 2 Allometric growth analysis of morphological index and the relationship between morphological index and individual size of P. yunnanensis seedlings
性状
Trait家系
Family决定系数
Determination coefficientP 斜率
Slope95%置信区间
95% confidence intervalF值
F valueP−1.0 类型
TypeH ~ D 2008 0.002 0.825 0.728b 0.509 ~ 1.042 3.227 0.082 I 2017 0.403 0.000 0.919b 0.695 ~ 1.215 0.372 0.546 I 2020 0.465 0.000 0.770b 0.596 ~ 0.995 4.300 0.046 A 2024 0.193 0.028 1.168ab 0.800 ~ 1.705 0.690 0.415 I 2033 0.138 0.062 1.006ab 0.686 ~ 1.474 0.001 0.975 I 2034 0.266 0.006 0.941ab 0.666 ~ 1.330 0.125 0.726 I 2036 0.474 0.000 0.678b 0.530 ~ 0.867 10.595 0.003 A 2048 0.359 0.000 1.483a 1.093 ~ 2.013 7.142 0.012 A 2050 0.366 0.000 0.990ab 0.753 ~ 1.303 0.005 0.944 I 2206 0.215 0.013 1.043ab 0.735 ~ 1.480 0.058 0.812 I D ~ TB 2008 0.406 0.000 0.832a 0.630 ~ 1.100 1.785 0.191 I 2017 0.577 0.000 0.354b 0.280 ~ 0.448 111.926 0.000 A 2020 0.753 0.000 0.506b 0.425 ~ 0.603 72.119 0.000 A 2024 0.555 0.000 0.442b 0.333 ~ 0.587 42.892 0.000 A 2033 0.754 0.000 0.473b 0.385 ~ 0.582 65.659 0.000 A 2034 0.884 0.000 0.406b 0.353 ~ 0.466 229.341 0.000 A 2036 0.826 0.000 0.485b 0.420 ~ 0.559 125.305 0.000 A 2048 0.712 0.000 0.432b 0.351 ~ 0.531 86.261 0.000 A 2050 0.862 0.000 0.410b 0.360 ~ 0.466 253.696 0.000 A 2206 0.588 0.000 0.514b 0.398 ~ 0.663 32.407 0.000 A 注:不同小写字母表示家系间差异显著 (P < 0.05)。A表示异速生长关系,I表示等速生长关系。下同。Notes: Different lowercase letters indicate significant differences among varied families (P < 0.05). A means allometric growth relationship, I indicates isometric growth relationship. The same below.
下载: 导出CSV
表 3 不同家系云南松苗木各器官生物量间的异速生长分析
Table 3 Analysis of allometric growth of each organ of P. yunnanensis seedlings among different families
性状
Trait家系
Family决定系数
Determination coefficientP 斜率
Slope95%置信区间
95% confidence intervalF值
F valueP−1.0 类型
TypeLB ~ RB 2008 0.729 0.000 1.017b 0.842 ~ 1.230 0.034 0.856 I 2017 0.814 0.000 1.033b 0.883 ~ 1.210 0.178 0.676 I 2020 0.801 0.000 1.168b 0.998 ~ 1.366 4.023 0.053 I 2024 0.538 0.000 1.658a 1.242 ~ 2.215 13.856 0.001 A 2033 0.808 0.000 1.298ab 1.080 ~ 1.559 8.690 0.007 A 2034 0.850 0.000 1.090b 0.930 ~ 1.277 1.237 0.277 I 2036 0.666 0.000 1.109b 0.911 ~ 1.351 1.128 0.295 I 2048 0.900 0.000 0.857b 0.759 ~ 0.969 6.670 0.015 A 2050 0.854 0.000 0.894b 0.783 ~ 1.021 2.946 0.095 I 2206 0.639 0.000 1.027b 0.808 ~ 1.305 0.050 0.826 I LB ~ SB 2008 0.756 0.000 0.902b 0.753 ~ 1.079 1.369 0.251 I 2017 0.725 0.000 0.774b 0.639 ~ 0.937 7.557 0.010 A 2020 0.849 0.000 1.010ab 0.880 ~ 1.158 0.020 0.889 I 2024 0.643 0.000 1.238a 0.959 ~ 1.600 2.973 0.098 I 2033 0.688 0.000 1.092ab 0.865 ~ 1.379 0.600 0.446 I 2034 0.872 0.000 0.995ab 0.859 ~ 1.152 0.006 0.941 I 2036 0.583 0.000 0.794b 0.637 ~ 0.989 4.554 0.040 A 2048 0.879 0.000 0.656b 0.574 ~ 0.750 43.564 0.000 A 2050 0.892 0.000 0.733b 0.654 ~ 0.822 31.413 0.000 A 2206 0.649 0.000 1.061ab 0.837 ~ 1.344 0.256 0.617 I SB ~ RB 2008 0.652 0.000 1.128a 0.910 ~ 1.398 1.303 0.262 I 2017 0.714 0.000 1.335a 1.099 ~ 1.622 9.301 0.005 A 2020 0.833 0.000 1.157a 1.001 ~ 1.336 4.207 0.048 A 2024 0.500 0.000 1.340a 0.992 ~ 1.810 4.051 0.056 I 2033 0.699 0.000 1.188a 0.945 ~ 1.494 2.386 0.136 I 2034 0.786 0.000 1.096a 0.907 ~ 1.324 0.977 0.333 I 2036 0.654 0.000 1.397a 1.143 ~ 1.707 11.746 0.002 A 2048 0.862 0.000 1.307a 1.132 ~ 1.508 14.864 0.001 A 2050 0.807 0.000 1.220a 1.048 ~ 1.421 7.064 0.012 A 2206 0.728 0.000 0.968a 0.786 ~ 1.193 0.101 0.753 I AGB ~ RB 2008 0.746 0.000 1.013a 0.843 ~ 1.217 0.020 0.889 I 2017 0.851 0.000 1.049a 0.911 ~ 1.207 0.474 0.496 I 2020 0.833 0.000 1.142a 0.989 ~ 1.320 3.528 0.069 I 2024 0.589 0.000 1.416a 1.077 ~ 1.860 7.041 0.014 A 2033 0.837 0.000 1.199a 1.012 ~ 1.421 4.902 0.037 A 2034 0.862 0.000 1.070a 0.919 ~ 1.241 0.829 0.371 I 2036 0.769 0.000 1.102a 0.935 ~ 1.299 1.423 0.241 I 2048 0.907 0.000 0.974a 0.866 ~ 1.096 0.203 0.656 I 2050 0.862 0.000 0.977a 0.858 ~ 1.111 0.139 0.712 I 2206 0.748 0.000 0.947a 0.774 ~ 1.157 0.311 0.582 I
下载: 导出CSV
表 4 不同家系云南松苗木茎 ~ 根、地上 ~ 根生物量沿共同主轴的位移
Table 4 Shift of biomass pair SB − RB and AGB − RB of seedlings along common axis of P. yunnanensis seedlings among different families
家系
FamilySB ~ RB AGB ~ RB 截距
Intercpt95%置信区间
95% confidence interval漂移
Shift截距
Intercept95%置信区间
95% confidence interval漂移
Shift2008 0.172b 0.095 ~ 0.238 −0.007b 0.675b 0.619 ~ 0.726 0.520b 2017 0.191b 0.059 ~ 0.283 0.574ab 0.718b 0.657 ~ 0.781 1.052ab 2020 0.313ab 0.193 ~ 0.383 −0.845b 0.902a 0.851 ~ 1.039 −0.107b 2024 0.275ab 0.161 ~ 0.390 0.268b 0.761b 0.655 ~ 0.870 0.755b 2033 0.180b 0.103 ~ 0.256 0.161b 0.722b 0.667 ~ 0.779 0.706b 2034 0.337a 0.254 ~ 0.433 0.472ab 0.771b 0.701 ~ 0.840 0.889ab 2036 0.266b 0.182 ~ 0.327 0.408b 0.765b 0.716 ~ 0.808 0.888b 2048 0.190b 0.125 ~ 0.248 0.277b 0.735b 0.701 ~ 0.775 0.811b 2050 0.065b −0.021 ~ 0.144 0.759a 0.616b 0.582 ~ 0.694 1.221a 2206 0.187b 0.083 ~ 0.226 −0.146b 0.685b 0.603 ~ 0.737 0.394b
下载: 导出CSV
表 5 不同家系云南松苗木器官生物量与个体大小间的异速生长分析
Table 5 Analysis of allometric growth between each organ and individual size of P. yunnanensis seedlings among different families
性状
Trait家系
Family决定系数
Determination coefficientP 斜率
Slope95%置信区间
95% confidence intervalF值
F valueP−1.0 类型
TypeRB 2008 0.823 0.000 1.015a 0.870 ~ 1.183 0.038 0.848 I 2017 0.892 0.000 0.974a 0.863 ~ 1.098 0.201 0.657 I 2020 0.868 0.000 0.901a 0.793 ~ 1.025 2.713 0.109 I 2024 0.703 0.000 0.785a 0.622 ~ 0.991 4.626 0.042 A 2033 0.877 0.000 0.876a 0.756 ~ 1.015 3.428 0.076 I 2034 0.895 0.000 0.953a 0.834 ~ 1.088 0.557 0.463 I 2036 0.818 0.000 0.933a 0.806 ~ 1.079 0.936 0.340 I 2048 0.935 0.000 1.029a 0.932 ~ 1.136 0.355 0.556 I 2050 0.910 0.000 1.034a 0.928 ~ 1.144 0.345 0.561 I 2206 0.819 0.000 1.069a 0.901 ~ 1.267 0.632 0.434 I SB 2008 0.874 0.000 1.145b 1.006 ~ 1.303 4.539 0.041 A 2017 0.851 0.000 1.300ab 1.129 ~ 1.497 14.690 0.001 A 2020 0.918 0.000 1.042b 0.942 ~ 1.153 0.692 0.412 I 2024 0.812 0.000 1.052b 0.873 ~ 1.267 0.316 0.580 I 2033 0.831 0.000 1.041b 0.876 ~ 1.237 0.229 0.636 I 2034 0.929 0.000 1.044b 0.936 ~ 1.165 0.652 0.427 I 2036 0.898 0.000 1.303ab 1.168 ~ 1.453 24.707 0.000 A 2048 0.947 0.000 1.345a 1.230 ~ 1.470 47.752 0.000 A 2050 0.941 0.000 1.258ab 1.155 ~ 1.369 30.720 0.000 A 2206 0.856 0.000 1.034b 0.888 ~ 1.205 0.206 0.654 I LB 2008 0.963 0.000 1.032b 0.962 ~ 1.107 0.851 0.363 I 2017 0.961 0.000 1.006b 0.936 ~ 1.081 0.031 0.861 I 2020 0.985 0.000 1.052b 1.007 ~ 1.099 5.614 0.024 A 2024 0.932 0.000 1.302a 1.164 ~ 1.457 24.195 0.000 A 2033 0.965 0.000 1.137b 1.051 ~ 1.230 11.446 0.002 A 2034 0.984 0.000 1.039b 0.987 ~ 1.093 2.294 0.142 I 2036 0.837 0.000 1.034b 0.901 ~ 1.188 0.246 0.623 I 2048 0.981 0.000 0.882b 0.837 ~ 0.930 23.824 0.000 A 2050 0.983 0.000 0.921b 0.880 ~ 0.965 13.080 0.001 A 2206 0.924 0.000 1.097b 0.982 ~ 1.226 2.942 0.098 I AGB 2008 0.991 0.000 1.028b 0.992 ~ 1.064 2.538 0.121 I 2017 0.996 0.000 1.021b 0.998 ~ 1.045 3.515 0.070 I 2020 0.997 0.000 1.030b 1.011 ~ 1.048 10.826 0.002 A 2024 0.984 0.000 1.111a 1.052 ~ 1.174 16.122 0.001 A 2033 0.996 0.000 1.051ab 1.025 ~ 1.078 16.631 0.000 A 2034 0.997 0.000 1.019b 0.998 ~ 1.041 3.466 0.074 I 2036 0.996 0.000 1.028b 1.006 ~ 1.050 6.757 0.014 A 2048 0.997 0.000 1.003b 0.983 ~ 1.023 0.084 0.774 I 2050 0.994 0.000 1.007b 0.980 ~ 1.034 0.249 0.621 I 2206 0.992 0.000 1.011b 0.976 ~ 1.048 0.424 0.521 I
下载: 导出CSV
-
[1] Dovrat G, Meron E, Shachak M, et al. Plant size is related to biomass partitioning and stress resistance in water-limited annual plant communities[J]. Journal of Arid Environments, 2019, 165: 1−9. doi: 10.1016/j.jaridenv.2019.04.006
[2] Mccarthy M C, Enquist B J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation[J]. Functional Ecology, 2007, 21(4): 713−720. doi: 10.1111/j.1365-2435.2007.01276.x
[3] Mokany K, Raison R J, Prokushkin A S. Critical analysis of root: shoot ratios in terrestrial biomes[J]. Global Change Biology, 2010, 12(1): 84−96.
[4] Lacointe A. Carbon allocation among tree organs: a review of basic processes and representation infunctional-structural tree models[J]. Annals of Forest Science, 2000, 57(5): 521−533. doi: 10.1051/forest:2000139
[5] Jackson R B, Schenk H J, Jobbágy E G, et al. Belowground consequences of vegetation change and their treatment in models[J]. Ecological Applications, 2000, 10(2): 470−483. doi: 10.1890/1051-0761(2000)010[0470:BCOVCA]2.0.CO;2
[6] Baker T R, Malhi Y, Phillips O L, et al. The above-ground coarse wood productivity of 104 Neotropical forest plots[J]. Global Change Biology, 2004, 10(5): 563−591. doi: 10.1111/j.1529-8817.2003.00778.x
[7] Weiner J. Allocation, plasticity and allometry in plants[J]. Perspectives in Plant Ecology Evolution & Systematics, 2004, 6(4): 207−215.
[8] 韩文轩, 方精云. 幂指数异速生长机制模型综述[J]. 植物生态学报, 2008, 32(4):951−960. doi: 10.3773/j.issn.1005-264x.2008.04.025 Han W X, Fang J Y. Review on the mechanism models of allometric scaling laws: 3/4 vs. 2/3 power[J]. Chinese Journal of Plant Ecology, 2008, 32(4): 951−960. doi: 10.3773/j.issn.1005-264x.2008.04.025
[9] Enquist B J, Allen A P, Brown J H, et al. Biological scaling: does the exception prove the rule?[J]. Nature, 2007, 445: E9−E10. doi: 10.1038/nature05548
[10] Cheng D L, Li T, Zhong Q L, et al. Scaling relationship between tree respiration rates and biomass[J]. Biology Letters, 2010, 6(5): 715−717. doi: 10.1098/rsbl.2010.0070
[11] Chen X W, Li B L. Testing the allometric scaling relationships with seedlings of two tree species[J]. Acta Oecologica, 2003, 24(3): 125−129. doi: 10.1016/S1146-609X(03)00062-6
[12] 金振洲, 彭鉴. 云南松[M]. 昆明: 云南科技出版社. 2004. Jin Z Z, Peng J. Pinus yunnanensis[M]. Kunming: Yunnan Science and Technology Press, 2004.
[13] 刘玉芳, 陈双林, 郭子武, 等. 淹水对河竹鞭根系统生物量分配及异速生长模式的影响[J]. 林业科学研究, 2015, 28(4):502−507. doi: 10.3969/j.issn.1001-1498.2015.04.008 Liu Y F, Chen S L, Gou Z W, et al. Effect of waterlogging on biomass allocation and allometric pattern of rhizome and root system of phyllostachys rivalis[J]. Forest Research, 2015, 28(4): 502−507. doi: 10.3969/j.issn.1001-1498.2015.04.008
[14] 黄迎新, 宋彦涛, 范高华, 等. 灰绿藜形态性状与繁殖性状的异速关系[J]. 草地学报, 2015, 23(5):905−913. doi: 10.11733/j.issn.1007-0435.2015.05.002 Huang Y X, Song Y T, Fan G H, et al. Allometric relationships between morphological and reproductive traits of Chenopodium glaucum[J]. Acta Agrestia Sinica, 2015, 23(5): 905−913. doi: 10.11733/j.issn.1007-0435.2015.05.002
[15] 范高华, 崔桢, 张金伟, 等. 密度对尖头叶藜生物量分配格局及异速生长的影响[J]. 生态学报, 2017, 37(15):5080−5090. Fan G H, Cui Z, Zhang J W, et al. Effects of population density on the biomass allocationand allometric growth of Chenopodium acuminatum[J]. Acta Ecologica Sinica, 2017, 37(15): 5080−5090.
[16] 林华, 陈双林, 郭子武, 等. 苦竹叶片性状及其异速生长关系的密度效应[J]. 林业科学研究, 2017, 30(4):617−623. Lin H, Chen S L, Gou Z W, et al. Allometric relationship among leaf traits in different stand density of Pleioblastus amarus[J]. Forest Research, 2017, 30(4): 617−623.
[17] 江洪, 林鸿荣. 云南松异速生长现象的初步研究[J]. 林业科学, 1984, 20(1):80−83. Jiang H, Lin H R. A preliminary study on allometric growth of Pinus yunnanensis[J]. Scientia Silvae Sinicae, 1984, 20(1): 80−83.
[18] 李鑫, 李昆, 段安安, 等. 不同地理种源云南松幼苗生物量分配及其异速生长[J]. 北京林业大学学报, 2019, 41(4):41−50. Li X, Li K, Duan A A, et al. Biomass allocation and allometry of Pinus yunnanensis seedlings from different provenances[J]. Journal of Beijing Forestry University, 2019, 41(4): 41−50.
[19] 侯勤正, 叶广继, 马小兵, 等. 青藏高原不同生境下湿生扁蕾(Gentianopsis paludosa)个体大小依赖的繁殖分配[J]. 生态学报, 2016, 36(9):2686−2694. Hou Q Z, Ye G J, Ma X B, et al. Size-dependent reproductive allocation of Gentianopsis paludosa in different habitats of the Qinghai-Tibetan Plateau[J]. Acta Ecologica Sinica, 2016, 36(9): 2686−2694.
[20] 李利平, 安尼瓦尔·买买提, 努尔巴依·阿布都沙力克, 等. 新疆山地森林乔木和草地草本植物个体大小分布特征[J]. 生物多样性, 2017, 25(11):1202−1212. doi: 10.17520/biods.2016336 Li L P, Anwar M, Nurbay A, et al. Plant body size patterns of mountainous trees and grassland herbs in Xinjiang region, China[J]. Biodiversity Science, 2017, 25(11): 1202−1212. doi: 10.17520/biods.2016336
[21] 徐国瑞, 马克明. 土壤动物粒径谱研究进展[J]. 生态学报, 2017, 37(8):2506−2519. Xu G R, Ma K M. Advances in the body size spectra study of soil fauna[J]. Acta Ecologica Sinica, 2017, 37(8): 2506−2519.
[22] 邱东, 周桂玲, 刘同业. 3种棒果芥属植物生物量分配及异速生长分析[J]. 干旱地区农业研究, 2014, 32(6):215−220. doi: 10.7606/j.issn.1000-7601.2014.06.036 Qiu D, Zhou G L, Liu T Y. Analysis of biomass allocation and allometric growth of three Sterigmostemum species in Junggar Basin[J]. Agricultural Research in the Arid Areas, 2014, 32(6): 215−220. doi: 10.7606/j.issn.1000-7601.2014.06.036
[23] Niklas K J. Size-dependent variations in plant growth ratesand the“3/4-power rule”[J]. American Journal of Botany, 1994, 81(2): 134−144. doi: 10.1002/j.1537-2197.1994.tb15422.x
[24] 吕学辉, 魏巍, 陈诗, 等. 云南松优良家系超级苗选择研究[J]. 云南大学学报(自然科学版), 2012, 34(1):113−119. Lü X H, Wei W, Chen S, et al. Study on super seedling selection of superior families of Pinus yunnanensis Franch[J]. Journal of Yunnan University (Natural Sciences), 2012, 34(1): 113−119.
[25] Niklas K J, Enquist B J. Canonical rules for plant organ bio-mass partitioning and annual allocation[J]. American Journal Botany, 2002, 89(5): 812−819. doi: 10.3732/ajb.89.5.812
[26] Poorter H, Niklas K J, Reich P B, et al. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variationand environment control[J]. New Phytologist, 2012, 193(1): 30−50. doi: 10.1111/j.1469-8137.2011.03952.x
[27] Bloom A J, Chapin F S, Mooney I A. Resource limitation inplants-an economic analogy[J]. Annual Review of Ecology and Systematics, 1985, 16: 363−392. doi: 10.1146/annurev.es.16.110185.002051
[28] 何怀江, 叶尔江·拜克吐尔汉, 张春雨, 等. 吉林蛟河针阔混交林 12 个树种生物量分配规律[J]. 北京林业大学学报, 2016, 38(4):53−62. He H J, Yeerjiang B, Zhang C Y, et al. Biomass allocation of twelve tree species in coniferous and broad-leaved mixed forest in Jiaohe, Jilin Province, northeast China[J]. Journal of Beijing Forestry University, 2016, 38(4): 53−62.
[29] 王晨, 江泽慧, 郭起荣, 等. 毛竹地上器官的生物量分配及其随个体大小变化的规律[J]. 生态学杂志, 2014, 33(8):2019−2024. Wang C, Jiang Z H, Gou Q R, et al. Biomass allocation of aboveground components of Phyllostachys edulis and its variation with body size[J]. Chinese Journal of Ecology, 2014, 33(8): 2019−2024.
[30] Weiner J. Allocation, plasticity and allometry in plants[J]. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4): 207−215. doi: 10.1078/1433-8319-00083
[31] 朱仕明, 肖玲玲, 薛立, 等. 密度对乐昌含笑幼苗的生长和生物量的影响[J]. 中南林业科技大学学报, 2015, 35(8):77−80. Zhu S M, Xiao L L, Xue L, et al. Effects of planting density on growth and biomass of Michelia chapensis seedlings[J]. Journal of Central South University of Forestry & Technology, 2015, 35(8): 77−80.
[32] 高平珍, 陈双林, 郭子武, 等. 毛竹林下苦参和决明幼苗生长和生物量分配的立竹密度效应[J]. 生态学杂志, 2018, 37(3):861−868. Gao P Z, Chen S L, Guo Z W, et al. Growth and biomass allocation of Sophotora flavescens and Catsia tora seedlings beneath Moso bamboo forest in response to Moso bamboo density[J]. Chinese Journal of Ecology, 2018, 37(3): 861−868.
[33] 冯银平, 沈海花, 罗永开, 等. 种植密度对苜蓿生长及生物量的影响[J]. 植物生态学报, 2020, 44(3):248−256. doi: 10.17521/cjpe.2019.0157 Feng Y P, Shen H H, Luo Y K, et al. Effects of planting density on growth and biomass of Medicago sativa[J]. Chinese Journal of Plant Ecology, 2020, 44(3): 248−256. doi: 10.17521/cjpe.2019.0157
[34] West G B, Brown J H, Enquist B J. A general model for the origin of allometric scaling laws in biology[J]. Science, 1997, 276: 122−126. doi: 10.1126/science.276.5309.122
[35] Niklas K J. Plant allometry: is there a grand unifying theory?[J]. Biological Reviews, 2004, 79(4): 871−889. doi: 10.1017/S1464793104006499
[36] Wang X P, Fang J Y, Tang Z Y, et al. Climatic control of primary forest structure and DBH-height allometry in Northeast China[J]. Forest Ecology and Management, 2006, 234(1/3): 264−274.
[37] 胡波, 钟全林, 程栋梁, 等. 刨花楠树高与胸径异速生长的关系[J]. 沈阳大学学报(自然科学版), 2012, 24(3):9−14. Hu B, Zhong Q L, Cheng D L, et al. Relationship between machilus’ height and allometric growth of diameter at breast[J]. Journal of Shenyang University(Natural Science), 2012, 24(3): 9−14.
[38] 赵广, 韩学琴, 王雪梅, 等. 修枝对辣木株高-地径异速生长关系的影响[J]. 生态学杂志, 2018, 37(2):391−398. Zhao G, Han X Q, Wang X M, et al. Effects of pruning on allometric relationship between height and basal diameter of Moringa oleifera[J]. Chinese Journal of Ecology, 2018, 37(2): 391−398.
[39] Sun S, Jin D, Shi P. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship[J]. Annals of Botany, 2006, 97(1): 97−107. doi: 10.1093/aob/mcj004
[40] Enquist B J. Global allocation rules for patterns of biomass partitioning in seed plants[J]. Science, 2002, 295: 1517−1520. doi: 10.1126/science.1066360
[41] Huang Y X, Lechowicz M J, Zhou D W, et al. Evaluating general allometric models: interspecific and intraspecific data tell different stories due to interspecific variation in stem tissue density and leaf size[J]. Oecologia, 2016, 180: 671−684. doi: 10.1007/s00442-015-3497-x
[42] 刘左军, 杜国祯, 陈家宽. 不同生境下黄胄橐(Ligularia virgaureain)个体大小依赖的繁殖分配[J]. 植物生态学学报, 2002, 26(1):44−50. Liu Z J, Du G Z, Chen J K. Individual size-dependent reproductive distribution of Ligularia virgaureain in different habitats[J]. Chinese Journal of Plant Ecology, 2002, 26(1): 44−50.
-
期刊类型引用(3)
1. 毛林海,孔祥涛,梁璞,傅金和,许佳诺. 竹材物理力学性质影响因素研究进展. 世界竹藤通讯. 2024(02): 91-97 .  百度学术
百度学术
2. 贾舒予,王游,韦鹏练,马欣欣,吴谊民. 竹节结构及力学性能研究现状. 世界竹藤通讯. 2024(03): 90-99 . 百度学术
3. 曹释予,张翔,季加贵,江甜,周雨砚,王雪花. 截面形态对竹条弯曲性能的影响. 家具. 2023(05): 33-37+116 . 百度学术
其他类型引用(1)

计量
- 文章访问数: 1448
- HTML全文浏览量: 589
- PDF下载量: 216
- 被引次数: 4
