
Wind tunnel simulation of complex deformation law on retaining wall under aeolian sand environment
-
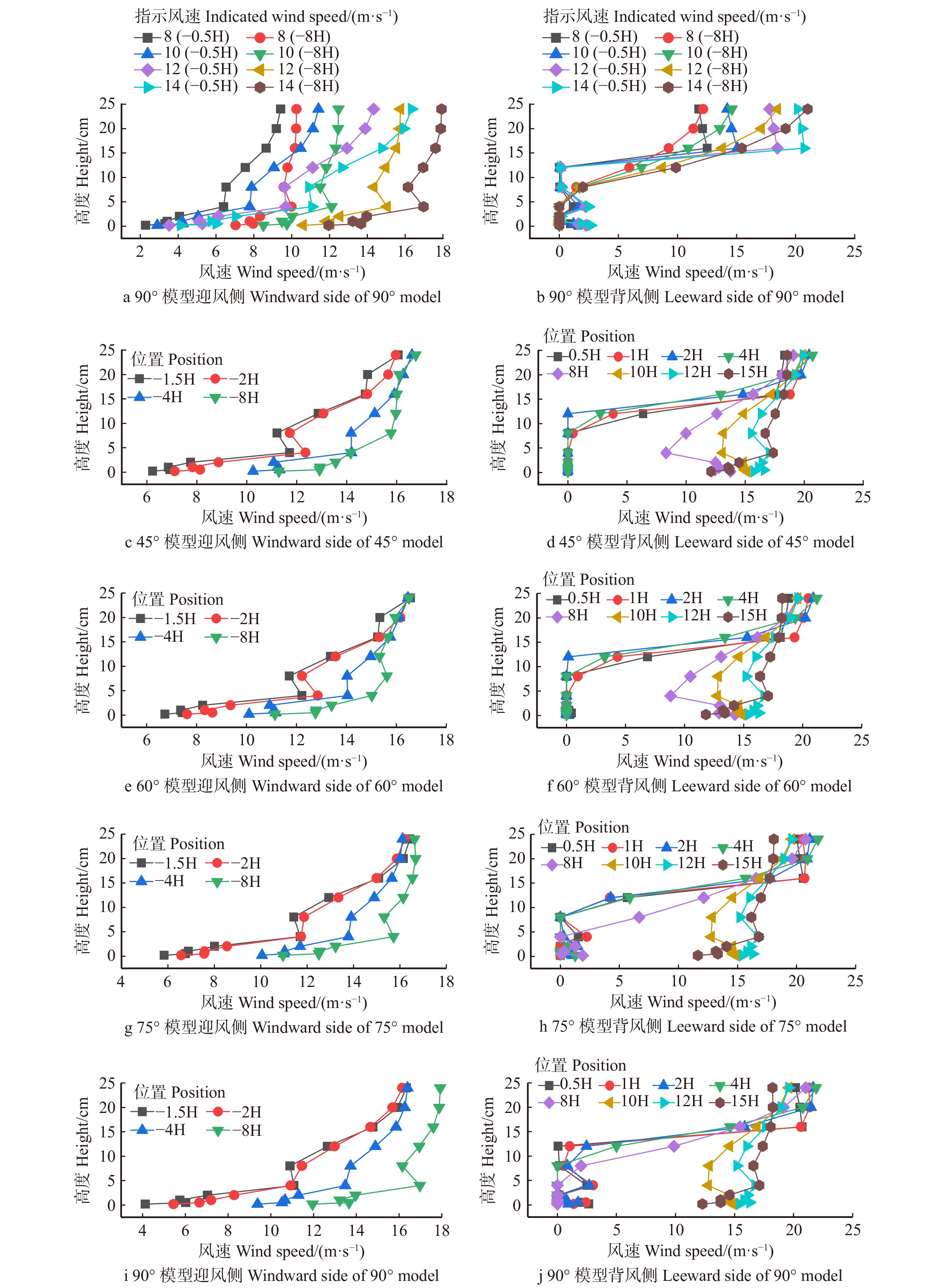
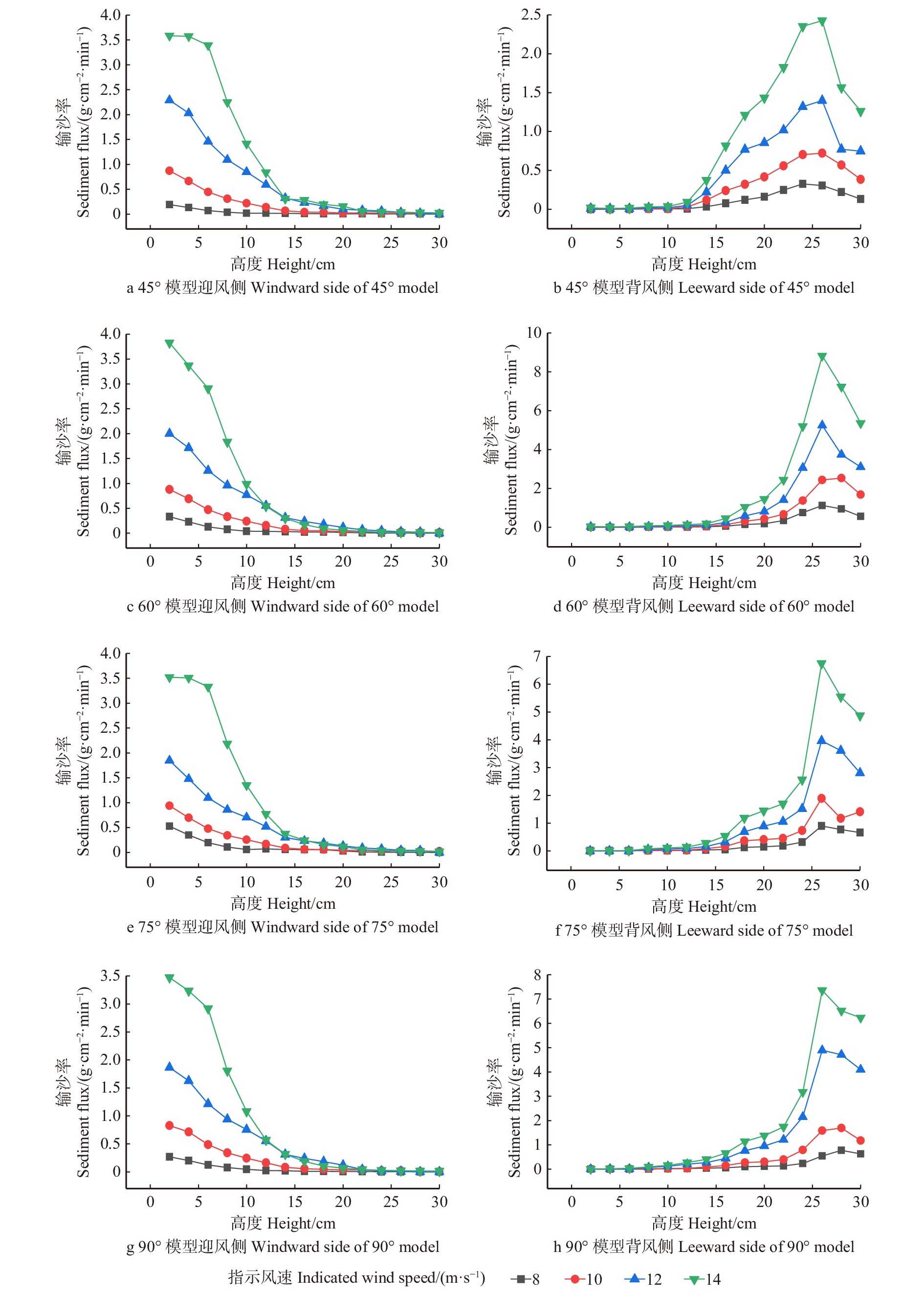
摘要:目的 风沙防护是干旱环境下人类基础设施建设和维护的关键工程问题之一,为探明不透风防风阻沙措施风沙运移的变化规律,筛选特殊风沙区域防护体系构建合适设置参数。方法 采用风洞模拟对不同风向夹角条件下防风挡沙墙的气流速度场、风沙流结构以及沙粒阻截率/输导率的变化规律进行研究。结果 (1)迎风侧模型4倍障高处始终为气流急剧变化的拐点,且不随着指示风速的变化而变化;当指示风速小于10 m/s,背风侧弱风区或静风区的范围随着指示风速的增大而增大,大于12 m/s则呈相反规律;同一指示风速作用下,随风向夹角的增大有效防护范围逐渐增加。(2)迎风侧积沙主要集中于近地表0 ~ 10 cm,平均占总输沙量的85.31%;45°、60°、75°与 90°4 种风向夹角条件下背风侧输沙量主要集中在20 ~ 30 cm高度范围内,分别占总输沙量的71.25%、88.75%、85.25%、86.00%。(3)背风侧0 ~ 10 cm高度层沙粒截留随指示风速的增加而增加,75°夹角时最大,平均为95.64%;10 ~ 30 cm高度层沙粒输导随着指示风速的增加有增大趋势,45°夹角时最大,平均为81.09%。结论 指示风速的变化对防风挡沙墙气流速度场变化规律影响不显著,而对背风侧弱风区或静风区的范围影响较大,75°时有效防护范围最佳;迎风侧输沙量随高度的增加逐渐减少,背风侧随高度的增大呈现出先增加后减少的变化趋势;作为防风阻沙措施风向夹角应设置为75°左右。Abstract:Objective Wind and sand protection is one of the key engineering issues in the construction and maintenance of human infrastructure in arid environment. This paper aims to find out the complex law of wind and sand migration, and to select the appropriate setting parameters of special wind and sand protection system.Method Wind tunnel simulation was used to study the variation of wind velocity field, sand flow structure and sand interception ratio/permeability under different wind direction angles.Result (1) The height of 4 times barrier on the windward side model was always the inflection point of the sharp change of the airflow, and did not change with the indicated wind speed. When the indicated wind speed was less than 10 m/s, the leeward area of weak wind or calm wind area increased with the indicated wind speed, and the opposite law was presented when the indicated wind speed was greater than 12 m/s. Under the action of the same indication wind speed, the effective protection range increased gradually with the increase of wind direction angle. (2) The windward sediment mainly concentrated at 0−10 cm near the surface, accounting for 85.31% of the total sediment transport. The leeward sediment transport under different wind direction angle measures mainly concentrated in the range of 20−30 cm height, accounting for 71.25%, 88.75%, 85.25% and 86.00% of the total sediment transport, respectively. (3) Sand interception in leeward layer at 0−10 cm height increased with the increase of indicated wind speed, and reached the maximum at the included angle of 75°, averaging 95.64%. The sand transport in the 10−30 cm height layer had an increasing trend with the increase of indicated wind speed, and the maximum was 81.09% on average when the included angle was 45°.Conclusion The variation of indicated wind speed has no significant influence on the variation law of the airflow velocity field of retaining wall, but has a greater influence on the leeward side of the weak wind area or the range of the calm wind area, and the effective protection range is the best at 75°. The sand transport on the windward side gradually decreases with the increase of height, while that on the leeward side climb up and then decline with the increase of height. The wind angle should be set at about 75° as a measure to prevent wind and sand.
-
蔗糖代谢在植物生长发育过程中发挥着重要的调控作用。作为植物中重要的碳源存储形式之一,蔗糖的合成和代谢过程倍受研究者们的关注。其中蔗糖合成过程主要由蔗糖磷酸合酶(sucrose phosphate synthase,SPS)和蔗糖磷酸化酶(sucrose phosphorylase,SPP)进行调控,而蔗糖合酶(sucrose synthase,SUS)调控蔗糖与UDP-葡萄糖和果糖间的可逆合成和分解过程,蔗糖转化酶(invertase,INV)则不可逆地调控蔗糖分解形成葡萄糖和果糖[1-2]。蔗糖转化酶作为蔗糖分解过程中的关键酶,按照其在植物细胞内分布部位的不同,可分为液泡转化酶(vacuolar invertase,VIN)、细胞壁转化酶(cell wall invertase,CWIN)和胞质转化酶(cytoplasmic invertase,CINV) 3类;按照其pH值的不同,细胞壁转化酶和液泡转化酶又被称为酸性转化酶(acid invertase,AIN),而胞质转化酶则被称为碱性/中性转化酶(alkaline/neutral invertase,NIN)[3]。因二者进化来源不一,碱性/中性转化酶与酸性转化酶的同源性较低[4],其中碱性/中性转化酶类属于糖基水解酶家族100,主要定位于叶绿体、线粒体和胞质溶胶内,与植物的生长发育和胁迫响应相关[5-7]。
前人对于拟南芥(Arabidopsis thaliana)的研究表明:碱性/中性转化酶在植物的生长发育过程中发挥着重要的调控作用[1]。在拟南芥中,碱性/中性转化酶基因家族共包含9个成员,其中AtCINV1(cytoplasmic invertase 1)调控拟南芥根形态的正常发育,抑制其表达会抑制主根生长和促进侧根的发育[8-9];AtINVE(alkaline/neutral invertase E)则参与拟南芥叶绿体的发育,抑制其表达不仅造成叶绿体发育畸形且提高了植株的氮同化速率[10];AtINVH(alkaline/neutral invertase H)则在拟南芥再生组织部位中具有特异地高表达,其突变体表现出茎缩短、开花时间延迟等特性,且其根部中的活性氧(ROS)水平急剧下降[11]。此外,在百脉根(Lotus japonicus)中,LjINV1(alkaline/neutral invertase 1)基因的突变会导致植株根和茎长度的缩短,且无花粉产生,影响植株的生殖发育[12]。在巴西橡胶树(Hevea brasiliensis)中,转化酶活性则与乳胶的产量紧密关联[13]。水稻(Oryza sativa)中OsCYT-INV1(cytoplasmic invertase 1)基因的突变则会导致根长的缩短和开花时间的延长以及不育[14]。在杂交山杨(Populus tremula × tremuloides)中,转化酶的活性则与纤维素合成相关,其活性的下降会导致纤维素含量的下降,影响植物的次生生长[15]。此外,碱性/中性转化酶的活性还与植物的胁迫防御相关。在拟南芥中AtINVA(alkaline/neutral invertase A)和AtINVG(alkaline/oneutral invertase G)与氧化防御相关基因的表达紧密相关,过表达AtINVA和AtINVG会抑制抗坏血酸过氧化物酶基因AtAPX2(oxidative stress-responsive ascorbate peroxidase 2)的表达[16]。在小麦(Triticum aestivum)中,渗透胁迫和低温处理则会引起叶片中转化酶活性的增加[17]。在枸橘(Poncirus trifoliata)中,过表达PtrA/NINV(alkaline/neutral invertase)可降低植株体内的活性氧水平并增强其光合作用效率,以应对冷、高盐和干旱等不利环境条件的影响[18]。在小麦中,Ta-A/N-Inv1(Alkaline/neutral invertase 1)的表达则与条锈病相关,其可能通过提高细胞内己糖水平从而降低光合作用,减少活性氧的产生,从而阻止叶片坏死[19]。
杨树(Populus spp.)因其木材性质优良、速生以及基因组结构简单,在木本植物中作为模式植物而具有重要的研究价值。随着毛果杨(Populus trichocarpa)基因组数据的公布,以杨树为代表植物开展对木本植物新陈代谢的研究显得尤为方便[20]。基于基因组数据,前人针对杨树的转化酶基因家族也已开展了相关分析,并对其成员在不同组织部位的表达模式进行了研究[5, 21],然而对于杨树中转化酶成员基因功能的研究却相对较少。本研究中,我们以毛白杨(Populus tomentosa)碱性/中性转化酶基因PtoNIN1为例,对其基因结构进行相关分析,并对其在不同组织部位的表达模式进行了探究;同时以模式植物拟南芥为例,对其进行过表达验证,初步探究PtoNIN1在植物生长发育过程中的作用,为研究杨树其他转化酶成员功能提供了借鉴,也为揭示杨树蔗糖代谢过程奠定了基础。
1. 材料和方法
1.1 植物材料
本研究中毛白杨根、茎、叶材料来源于实验室内部保存生长6个月的毛白杨优良无性系组培苗TC1521,同时参照Chen等[22-23]方法分别采集成年毛白杨植株8个不同发育时期下雌花芽和雄花芽材料,以及成年树的成熟叶。采用改良的CTAB法分别提取各样品的总RNA,通过Reverse Transcription System Ⅰ(Promega)系统反转录合成cDNA,用于PtoNIN1基因的克隆和组织特异性表达研究。
转基因受体材料为实验室保存的哥伦比亚生态型拟南芥。拟南芥种子经由5%次氯酸钠溶液(AR,Macklin)消毒15 min后移入1/2 MS培养基(添加20 g/L蔗糖和5.5 g/L琼脂),4 ℃冰箱低温储藏3 d后移至气候箱内培养。培养箱温度为(22 ± 1) ℃,采用16 h光照/8 h黑暗的长光照周期进行培养。培养7 d后移至含有蛭石的营养土内进行培养。
1.2 毛白杨同源PtoNIN1基因的克隆与测序
以毛果杨转化酶基因家族的同源基因PtrNIN1(Potri.008G101500)为索引,基于已公布的毛白杨基因组序列[24]设计并合成了克隆引物(表1)。以毛白杨组培苗叶片材料的cDNA为模版,进行PCR扩增。PCR反应体系为:2 × Phanta® Max Master Mix (Vazyme)10 μL,前后引物(1 μmol/L)各0.5 μL,模板cDNA(25 ~ 50 ng/μL)2 μL,ddH2O 0.8 μL,总体系为20 μL。PCR的反应条件为:预变性98 ℃,3 min;变性98 ℃,30 s;退火58 ℃,60 s;延伸72 ℃,1 min,共进行35个循环;再延伸72 ℃,5 min,最后12 ℃保存。PCR克隆后通过FastPure® Gel DNA Extraction Mini Kit (Vazyme)进行切胶回收。连接转化后送至公司进行测序,测序数据由北京睿博兴科生物科技有限公司提供。
表 1 本实验所用引物序列Table 1. Primers used in the study基因
Gene正向引物
Forward primer反向引物
Reverse primerC-PtoNIN1 ATGAACAGTAGTAGTTGTA TCAGACATGAATTTGAGATC O-PtoNIN1 AGAACACGGGGGACTCTTGACATGAACAGT
AGTAGTTGTATTGGTAGGGGAAATTCGAGCTGGTCACTCAGACATG
AATTTGAGATCTTGCPtoNIN1 AACAATGAAACCCTGTTGCAG CCGTATCTTGTGACCAACTACGG PtoActin GGATGGCTATATAGGTTGCAGGAA GAGATGCTGGGAAGGAATTGAAAG AtSUS2 AGGGTGTACCAAATCTCAT CATAGTGAAAGCTGTGTGG AtSUS3 GAGATACCGCAGGGAGAGTT CAGCATTTCAGTCTCAAGGG AtSUS4 CAAGTGTAAGCATGACCC AGGAACAGCTTGAGCCAG AtSPP2 AGAAGCTAGCAACTTCCCTGAG GCTAACCTTGTGTGGCCTCT AtSPS2 TGTTTCAACCTTGGCACAAA GTGGTTTCAACCCTCCTGAA AtEF1A4α TTGACCAGATCAACGAGCCCAAGA ACTCGTGGTGCATCTCAACAGACT 注:C-PtoNIN1为PtoNIN1基因CDS区克隆所用引物,O-PtoNIN1为PtoNIN1过表达载体构建所用引物,PtoNIN1为PtoNIN1基因表达量检测所用引物。Notes: C-PtoNIN1 is used for the gene colning of PtoNIN1, O-PtoNIN1 is used for the construction of over-expression vector, and PtoNIN1 is used for qRT-PCR. 1.3 毛白杨PtoNIN1基因的生物信息学分析
基于Phytozome数据库(https://phytozome-next.jgi.doe.gov/)的毛果杨基因组数据绘制PtoNIN1基因的染色体定位和基因结构,蛋白的理化性质采用在线软件ProtParam tool(https://web.expasy.org/ protparam/)进行分析,保守结构域分析采用在线软件InterProScan(http://www.ebi.ac.uk/interpro/search)和Pfam(http://pfam.xfam.org/ search)进行分析,二级结构分析则通过在线软件SOPMA(https://npsa-prabi.ibcp.fr/ cgi-bin/npsa_automat.pl?page=npsa_sopma.html)进行统计分析,蛋白三级结构采用软件SWISS-MODEL(https://swissmodel.expasy.org/)进行分析,跨膜结构域预测采用在线软件TMHMM 2.0(https://services.healthtech.dtu.dk/service.php?TM HMM-2.0)进行分析,信号肽的预测采用在线软件SignalP-5.0(https://services. healthtech.dtu.dk/service.php?SignalP-5.0)进行分析,亚细胞定位预测采用在线软件PSORT Prediction(http://psort1.hgc.jp/form.html)进行,分别利用在线软件NetPhos 3.1(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)和NetNGlyc 1.0(https://services.healthte ch.dtu.dk/service.php?NetNGlyc-1.0)对PtoNIN1蛋白的磷酸化位点和糖基化位点进行预测分析,多序列同源比对则采用软件DNAMAN(V9.0.1.116)进行。启动子区序列分析则采用在线软件PlantCARE(http://bioinformatics.psb.ugent. be/-webtools/plantcare/html/)进行分析。
为进一步分析PtoNIN1蛋白的进化关系,基于基因的氨基酸序列通过MEGA11.0构建系统进化树,采用Neighbor-Joining(NJ)算法的Poisson模型,Complete deletion模式进行建树,bootstrap值取1 000。其中,毛果杨包含12个成员,PtrNIN1(Potri.008G101500)、PtrNIN2(Potri.013G006600)、PtrNIN3(Potri.008G024100)、PtrNIN4(Potri.010G236100)、PtrNIN5(Potri.005G010800)、PtrNIN6(Potri.004G186500)、PtrNIN7(Potri.005G239400)、PtrNIN8(Potri.019G082000)、PtrNIN9(Potri.004G167500)、PtrNIN10(Potri.002G173600)、PtrNIN11(Potri.009G129000)、PtrNIN12(Potri.013G110800);拟南芥包含9个成员,AtCINV1(AT1G35580)、AtCINV2(AT4G09510)、AtINVA(AT1G56560)、AtINVB(AT4G34860)、AtINVC(AT3G06500)、AtINVD(AT1G22650)、AtINVE(AT5G22510)、AtINVF(AT1G72000)、AtINVH(AT3G05820);水稻包含8个成员,OsNIN1(LOC_Os03g20020)、OsNIN2(LOC_Os01g22900)、OsNIN3(LOC_Os02g32730)、OsNIN4(LOC_Os04g33490)、OsNIN5(LOC_Os02g03320)、OsNIN6(LOC_Os11g07440)、OsNIN7(LOC_Os04g35280)、OsNIN8(LOC_Os02g34560);其余蛋白成员依次为LbNI(KR026955)、DlNI(KP769773)、DoNI2(KY794404)、HbNIN1(GU573728)、HbNIN2(GU573727)、HbNIN3(KC577600)、HfCIN3(MT379656)、DcNI4(XM_020835477.2)、Ta-A/N-Inv1(AM295169)。其中毛果杨和水稻基因数据来自Phytozome数据库,拟南芥基因数据来自TAIR数据库(https://www.arabidopsis.org/),其他基因数据来自NCBI数据库(https://www.ncbi.nlm.nih.gov/)。同时利用在线软件MEME(https://meme-suite.org/meme/tools/meme)对不同物种间转化酶成员的保守序列元件进行预测分析。
1.4 毛白杨PtoNIN1基因的组织特异性表达分析
实时荧光定量PCR(qRT-PCR)引物(表1)使用Primer3在线软件(http://fokker.wi.mit.edu)进行设计,采用StepOne PlusTM Real-Time PCR System with Tower(ABI,USA)检测系统和ChamQ Universal SYBR qPCR Master Mix(Vazyme)进行qRT-PCR扩增检测,以PtoActin基因为内参对照[22],采用2−∆∆Ct法计算基因的相对表达量。毛果杨碱性/中性转化酶基因家族成员表达量数据来源于Phytozome数据库中已公布的转录组数据。
1.5 农杆菌介导的拟南芥遗传转化
以实验室所有的pCAMBIA1301载体质粒为骨架,采用无缝克隆试剂盒ClonExpress®ⅡOne Step Cloning Kit(c112-02,Vazyme)构建由35S启动子驱动的PtoNIN1过表达载体,无缝克隆引物如表1所示。载体经由测序验证后转化农杆菌感受态GV3101,通过农杆菌介导的花序浸染法获得拟南芥转基因植株。拟南芥转基因植株采用潮霉素进行筛选,潮霉素使用质量浓度为20 mg/L。
1.6 拟南芥转基因植株的表型鉴定
经由PCR验证后,对萌发后培养30 d的T3代转基因拟南芥植株进行表型测定,同时参照王静澄[25]的方法测定转基因拟南芥根、茎及叶中的果糖和蔗糖含量。采用葡萄糖测定试剂盒(A154-1-1,南京建成生物工程研究所)测定转基因植株中葡萄糖的含量,每个转基因株系分别选取5个单株作为生物学重复,以拟南芥野生型植株为对照,计算转基因植株中蔗糖、葡萄糖和果糖的相对含量。采用qRT-PCR的方法测定拟南芥内源蔗糖代谢相关基因的表达量变化,引物序列如表1所示,以拟南芥AtEF1A4α基因为内参对照[26],采用2−∆∆Ct法计算基因的相对表达量。
2. 结果与分析
2.1 毛白杨PtoNIN1基因的克隆与生物信息学分析
以毛果杨同源基因PtoNIN1和已公布的毛白杨基因组序列为参考,克隆并获得了毛白杨同源基因序列PtoNIN1。测序结果显示,毛白杨PtoNIN1基因的CDS区序列长度为2 073 bp,共编码690个氨基酸(图1a);染色体定位结果显示,PtoNIN1位于毛白杨第8号染色体,共包含6个外显子区和5个内含子区(图1b,c)。氨基酸理化性质分析结果显示:PtoNIN1蛋白分子量77.80 kDa,理论等电点为6.22,分子式为C3 474H5 376N948O1 027S32,不稳定系数38.58,为稳定性蛋白。亲疏水性分析结果显示,PtoNIN1蛋白的亲水性总平均值为−0.277,为亲水性蛋白。
![]() 图 1 毛白杨中PtoNIN1的序列(a)、染色体定位(b)和基因结构(c)分析Figure 1. Sequence (a), chromosomal location (b) and gene structure (c) of PtoNIN1 in Populus tomentosa
图 1 毛白杨中PtoNIN1的序列(a)、染色体定位(b)和基因结构(c)分析Figure 1. Sequence (a), chromosomal location (b) and gene structure (c) of PtoNIN1 in Populus tomentosa蛋白的二级结构分析结果显示:无规则卷曲在PtoNIN1蛋白中所占比例最高,为39.42%,其次为α螺旋(36.81%)、伸展链(18.26%)和β折叠(5.51%)(图2a)。三级结构分析结果显示:其可形成同型六聚体结构,与鱼腥藻(Anabaena spp.)中碱性转化酶InvA蛋白结构最为相近(图2b)。结构域分析结果显示:其含有糖苷酶超家族100(glycosyl-hydrolase-100 superfamily)的特征结构域,属于碱性和中性转化酶基因家族。亚细胞定位预测结果显示,其可能位于线粒体基质空间中。跨膜结构域和信号肽的预测结果显示,PtoNIN1蛋白不具备跨膜结构域和信号肽。磷酸化位点预测结果显示:PtoNIN1蛋白中存在着丝氨酸、苏氨酸和酪氨酸磷酸化位点,其中丝氨酸磷酸化位点占比最高(图2c)。糖基化位点预测结果显示,PtoNIN1蛋白的2号、37号、44号、63号、123号、147号、176号和189号位置上的天冬酰胺为潜在的糖基化氨基酸位点(图2d)。
![]() 图 2 毛白杨PtoNIN1蛋白的二级结构(a)、三级结构(b)、磷酸化(c)和糖基化(d)位点分析图a中紫色代表无规则卷曲,蓝色代表α螺旋,红色代表伸展链,绿色代表β折叠。In Fig. a, purple represents irregular curls, blue represents α-spiral, red represents extension chain, and green represents β-fold.Figure 2. Secondary (a) and tertiary (b) structure, phosphorylation (c) and glycosylation site (d) of PtoNIN1 in P. tomentosa
图 2 毛白杨PtoNIN1蛋白的二级结构(a)、三级结构(b)、磷酸化(c)和糖基化(d)位点分析图a中紫色代表无规则卷曲,蓝色代表α螺旋,红色代表伸展链,绿色代表β折叠。In Fig. a, purple represents irregular curls, blue represents α-spiral, red represents extension chain, and green represents β-fold.Figure 2. Secondary (a) and tertiary (b) structure, phosphorylation (c) and glycosylation site (d) of PtoNIN1 in P. tomentosa2.2 毛白杨PtoNIN1基因的多序列比对和系统进化分析
以Phytozome v13.0数据库中毛果杨基因组数据为参照,对毛白杨碱性/中性转化酶PtoNIN1与毛果杨的碱性/中性转化酶基因家族成员进行多序列比对分析(图3)。其中PtoNIN1与毛果杨中性转化酶基因家族中12个成员的氨基酸相似性依次为95.38%、69.13%、53.98%、54.48%、66.05%、55.07%、44.06%、42.46%、43.19%、40.84%、44.06%和42.32%。
![]() 图 3 PtoNIN1蛋白与毛果杨碱性/中性转化酶基因家族成员的多序列比对Figure 3. Multiple sequence alignment of PtoNIN1 with the alkaline/neutral invertase gene family in P. trichocarpa
图 3 PtoNIN1蛋白与毛果杨碱性/中性转化酶基因家族成员的多序列比对Figure 3. Multiple sequence alignment of PtoNIN1 with the alkaline/neutral invertase gene family in P. trichocarpa此外,为了进一步探究PtoNIN1的蛋白功能,我们联合了与其同源性较高的其他物种中研究较为透彻的碱性/中性转化酶蛋白构建了系统进化树(图4)。聚类分析结果显示:植物中性转化酶基因家族可分为2大亚类,一类主要定位于胞质内,包含AtCINV1、AtCINV2、Ta-A/N-Inv1、LbNI、HbNIN1/2等成员;另一类主要定位于细胞器内,按照其定位的细胞器类型又可分为2种类型,一类位于线粒体型(Ⅰ-Ⅰ),一类位于叶绿体型(Ⅰ-Ⅱ)。其中,PtoNIN1与毛果杨中同源蛋白PtrNIN1亲缘关系最近,与拟南芥中AtINVA/H/C、水稻中OsNIN1/2和铁皮石斛(Dendrobium officinale)中DoNI2处于同一分支,表明其类属于线粒体型转化酶成员,这也与亚细胞定位预测的结果相一致。MEME分析结果显示:植物碱性/中性转化酶家族中共含有11个不同的保守基序,除毛果杨中PtoNIN7、PtoNIN9、PtoNIN10和PtoNIN11、水稻中OsNIN5和OsNIN7以及AtINVB和LbNI外,其余碱性/中性转化酶成员中均含有这11个保守的基序。
![]() 图 4 植物碱性/中性转化酶基因家族的系统进化和保守基序分析Figure 4. Phylogenetic and conserved motif analysis of alkaline/neutral invertase gene families in plant
图 4 植物碱性/中性转化酶基因家族的系统进化和保守基序分析Figure 4. Phylogenetic and conserved motif analysis of alkaline/neutral invertase gene families in plant基于已公布的毛白杨基因组序列,我们以PtoNIN1基因上游3 kbp区域序列为例,对其启动子区顺式作用元件进行预测分析(图5)。其中,光响应元件(13个)是PtoNIN1中分布最为丰富的顺式作用元件,表明PtoNIN1是潜在的光调控基因。此外,干旱诱导(2个)、厌氧诱导(4个)、防御和胁迫响应(2个)以及赤霉素响应元件(2个)也分布于PtoNIN1的启动子区域。
![]() 图 5 PtoNIN1启动子区顺式作用元件分析Figure 5. Analysis of cis-acting elements in the promoter region of PtoNIN1
图 5 PtoNIN1启动子区顺式作用元件分析Figure 5. Analysis of cis-acting elements in the promoter region of PtoNIN12.3 毛白杨PtoNIN1基因的时空特异性表达模式分析
为了进一步探究PtoNIN1在毛白杨发育过程中的作用,我们以PtoActin基因为内参基因,对不同组织部位(根、茎、叶及成年树的成熟叶)和不同发育时期毛白杨雄花芽和雌花芽的表达模式进行分析。结果显示:PtoNIN1在毛白杨根中表达量较高,其次是成年树的成熟叶,在茎和叶中表达量则相对较低(图6a)。对不同发育时期下雌花芽和雄花芽的研究结果表明:PtoNIN1在雌雄花芽成花诱导期(时期S1)具有高表达,随后表达量出现明显下降,并在整个花芽发育时期均保持较为稳定的表达趋势(图6b,c)。
![]() 图 6 PtoNIN1的基因表达模式分析S1. 成花诱导期;S2. 花原基形成期;S3. 器官发生期;S4. 伸长期;S5. 孢子形成期;S6 ~ S7. 休眠期;S8. 小孢子发生期。S1, floral induction stage; S2, primordia formation stage; S3, organogenesis stage; S4, enlargement stage; S5, archespore formation stage; S6−S7, dormancy stages; S8, microsporogenesis stage.Figure 6. Gene expression pattern analysis of PtoNIN1
图 6 PtoNIN1的基因表达模式分析S1. 成花诱导期;S2. 花原基形成期;S3. 器官发生期;S4. 伸长期;S5. 孢子形成期;S6 ~ S7. 休眠期;S8. 小孢子发生期。S1, floral induction stage; S2, primordia formation stage; S3, organogenesis stage; S4, enlargement stage; S5, archespore formation stage; S6−S7, dormancy stages; S8, microsporogenesis stage.Figure 6. Gene expression pattern analysis of PtoNIN12.4 拟南芥中PtoNIN1基因的过表达分析
为进一步探究PtoNIN1的基因功能,我们以pCAMBIA1301载体质粒为骨架,为PtoNIN1构建过表达载体(图7a),并对模式植物拟南芥进行过表达研究。T3代植物经由PCR扩增验证后共获得6个转基因过表达株系(图7b)。
![]() 图 7 拟南芥过表达载体的构建(a)和转基因株系验证(b)图a中基因黑色区域部分为用于荧光定量检测和转基因株系验证所用片段;图b中1 ~ 6分别代表了PtoNIN1的6个表达株系,col则为拟南芥野生型,M为5000 bp的DNA Marker。In Fig. a, the black region of the gene is the fragment used for qRT-PCR and transgenic line validation; in Fig. b, the number of 1−6 represent the six overexpression lines of PtoNIN1, respectively, col represents the wild-type of arabidopsis, and M is a DNA Marker for 5000 bp.Figure 7. PtoNIN1-overexpression vector (a) and validation of transgenic lines (b) in arabidopsis
图 7 拟南芥过表达载体的构建(a)和转基因株系验证(b)图a中基因黑色区域部分为用于荧光定量检测和转基因株系验证所用片段;图b中1 ~ 6分别代表了PtoNIN1的6个表达株系,col则为拟南芥野生型,M为5000 bp的DNA Marker。In Fig. a, the black region of the gene is the fragment used for qRT-PCR and transgenic line validation; in Fig. b, the number of 1−6 represent the six overexpression lines of PtoNIN1, respectively, col represents the wild-type of arabidopsis, and M is a DNA Marker for 5000 bp.Figure 7. PtoNIN1-overexpression vector (a) and validation of transgenic lines (b) in arabidopsis拟南芥中过表达PtoNIN1显著提高了植株中生物量的积累。与野生型相比,转基因株系中仅Pro35S:PtoNIN1/col-2的株高出现显著提升,其余株系与野生型相比并无明显差异(图8a,b)。然而与野生型相比,转基因株系中Pro35S:PtoNIN1/col-2和Pro35S:PtoNIN1/col-4的茎及角果鲜质量显著高于野生型,Pro35S:PtoNIN1/col-2、Pro35S:PtoNIN1/col-3和Pro35S:PtoNIN1/col-4的莲座叶鲜质量显著高于野生型,表明PtoNIN1在拟南芥中的过表达可能影响了植株生物量的积累(图8b,c)。对表型差异明显的转基因株系Pro35S:PtoNIN1/col-2/3/4中蔗糖、葡萄糖和果糖的测定分析结果表明:PtoNIN1的过表达并未影响莲座叶中蔗糖、葡萄糖和果糖的含量;然而与野生型相比,转基因株系茎及角果中蔗糖的含量显著提升,而Pro35S:PtoNIN1/col-4中果糖含量相比于野生型出现显著下降,其余株系中果糖和葡萄糖的含量相对于野生型并未产生显著差异(图8d ~ f)。
![]() 图 8 拟南芥及其转基因株系的表型分析图a中左侧为培养40 d的拟南芥野生型植株,右侧为培养40 d的Pro35S:PtoNIN1/col-2转基因过表达植株;图b ~ g中横坐标col表示野生型,1 ~ 6分别表示转基因株系Pro35S:PtoNIN1/col-1/2/3/4/5/6。其中图e ~ g中黑色柱子代表莲座叶部位的糖含量比较,灰色柱子代表茎及角果部位的糖含量比较;**代表在P < 0.01下差异显著,*代表在P < 0.05下差异显著。下同。In Fig. a, the left side shows the wild-type plants in arabidopsis cultured for 40 d, and the right side shows the transgenic plants of Pro35S: PtoNIN1/col-2 cultured for 40 d; in Fig. b−g, col represents the wild-type plants in arabidopsis, and the number of 1−6 represent the transgenic plants of Pro35S: PtoNIN1/col-1/2/3/4/5/6, respectively; thereinto, the black column in Fig. e−g represents the sugar content in the rosette leaves, and the gray column represents the sugar content in the stems and siliques. ** means significant difference at P < 0.01 level, * means significant difference at P < 0.05 level. The same below.Figure 8. Phenotypic analysis of arabidopsis and its transgenic lines
图 8 拟南芥及其转基因株系的表型分析图a中左侧为培养40 d的拟南芥野生型植株,右侧为培养40 d的Pro35S:PtoNIN1/col-2转基因过表达植株;图b ~ g中横坐标col表示野生型,1 ~ 6分别表示转基因株系Pro35S:PtoNIN1/col-1/2/3/4/5/6。其中图e ~ g中黑色柱子代表莲座叶部位的糖含量比较,灰色柱子代表茎及角果部位的糖含量比较;**代表在P < 0.01下差异显著,*代表在P < 0.05下差异显著。下同。In Fig. a, the left side shows the wild-type plants in arabidopsis cultured for 40 d, and the right side shows the transgenic plants of Pro35S: PtoNIN1/col-2 cultured for 40 d; in Fig. b−g, col represents the wild-type plants in arabidopsis, and the number of 1−6 represent the transgenic plants of Pro35S: PtoNIN1/col-1/2/3/4/5/6, respectively; thereinto, the black column in Fig. e−g represents the sugar content in the rosette leaves, and the gray column represents the sugar content in the stems and siliques. ** means significant difference at P < 0.01 level, * means significant difference at P < 0.05 level. The same below.Figure 8. Phenotypic analysis of arabidopsis and its transgenic lines此外,以3个表型最为突出的拟南芥转基因株系为例,我们也对外源基因PtoNIN1及拟南芥内源蔗糖代谢相关基因的表达量变化进行了分析。其中外源基因PtoNIN1的表达量与拟南芥表型具有显著相关关系,表型最为显著的Pro35S:PtoNIN1/col-2株系中外源PtoNIN1的表达量相对较高(图9a)。参照前人研究[26-28],我们分别选取了野生型拟南芥中表达量相对较高的蔗糖合酶成员(AtSUS2-4)、蔗糖磷酸合酶成员(AtSPS2)和蔗糖磷酸化酶成员(AtSPP2)对拟南芥转基因株系中蔗糖代谢过程的变化进行分析。相比于野生型,拟南芥叶片、茎及角果中的AtSUS2-4、AtSPS2和AtSPP2的表达量均出现了显著上升,且相比于叶片,茎及角果中基因表达量的上升更为显著(图9b ~ f)。
![]() 图 9 拟南芥及其转基因株系的基因表达模式分析黑色柱子代表莲座叶部位中基因表达模式分析,灰色柱子代表茎及角果部位的基因表达模式分析,2 ~ 4分别表示转基因株系Pro35S:PtoNIN1/col-2/3/4。The black column represents the gene expression patterns in the rosette leaves, and the gray column represents the gene expression patterns in the stems and siliques, and the number of 2−4 represent the transgenic plants of Pro35S: PtoNIN1/col-2/3/4, respectively.Figure 9. Gene expression patterns of arabidopsis and its transgenic lines
图 9 拟南芥及其转基因株系的基因表达模式分析黑色柱子代表莲座叶部位中基因表达模式分析,灰色柱子代表茎及角果部位的基因表达模式分析,2 ~ 4分别表示转基因株系Pro35S:PtoNIN1/col-2/3/4。The black column represents the gene expression patterns in the rosette leaves, and the gray column represents the gene expression patterns in the stems and siliques, and the number of 2−4 represent the transgenic plants of Pro35S: PtoNIN1/col-2/3/4, respectively.Figure 9. Gene expression patterns of arabidopsis and its transgenic lines3. 讨 论
蔗糖代谢作为植物生长发育过程中的重要代谢过程,可通过分解蔗糖为植物提供生长发育所需的能源,另一方面蔗糖分解所形成的单糖分子又可作为信号分子调控植物的生长发育过程[1]。转化酶作为植物蔗糖分解过程中2类关键酶之一,在植物的生长发育、信号转导、碳源分配和外界环境刺激响应等方面发挥了重要作用[29]。然而,前人的研究多集中于酸性转化酶家族,碱性/中性转化酶因其稳定性差、活性低而在前期被研究者们认为其仅起到酸性转化酶功能的补充作用[30]。随着测序数据的公布和基因组学的不断发展,越来越多研究者意识到碱性/中性转化酶家族具有其独特的功能,且在水稻(Oryza sativa)[31]、玉米(Zea mays)[32]、棉花(Gossypium barbadense)[33]、柑橘(Citrus reticulata)[34]、葡萄(Vitis vinifera)[35]、梨(Pyrus bretschneideri)[36]和苹果(Malus pumila)[37]等多个植物物种中开展了基因家族的遗传进化分析。然而对于中性转化酶基因功能的研究却相对较少,且多集中于模式植物拟南芥[4, 21, 25]和一些草本植物,如水稻[14]、小麦[19]、枸杞[38]等,木本植物中的研究仍十分匮乏。
杨树作为木本植物中的模式植物,随着其基因组数据的公布,研究者们也针对其中性转化酶家族开展了相关研究[5],然而对于杨树碱性/中性转化酶基因功能的研究却鲜有报道。本研究中我们以毛果杨中PtrNIN1序列为索引,对已公布的毛白杨基因组序列进行同源比对分析,从中获得了毛白杨同源基因序列PtoNIN1。对其进行生物信息学分析显示:其具有碱性/中性转化酶家族特征结构域,且包含12个保守的氨基酸基序,这与前人关于杨树碱性/中性转化酶基因家族的研究相一致[5]。对其进行三级结构预测分析显示:其与鱼腥藻中InvA结构极为类似,该基因是植物碱性/中性转化酶基因家族的潜在祖先之一,可特异识别蔗糖的α-1, 2-糖苷键[39]。同源序列比对和进化树分析均表明PtoNIN1与PtrNIN1具有高度相似性,表明碱性/中性转化酶在杨属不同物种间具有保守性。此外,PtoNIN1在进化上与AtINVA/H/C、OsNIN1和DoNI2处于同一分支,该类蛋白均定位于线粒体内,这也与亚细胞定位预测的结果相一致。在拟南芥中,AtINVA/C在植物的根、茎和花均具有明显的表达[40],OsNIN1在水稻的各个组织部位均具有显著的表达[31],DoNI2在铁皮石斛中根、茎、叶也被检测出具有较高的表达量[41],表明线粒体定位的转化酶成员在植株各个组织部位的发育中都具有重要的功能。本研究中PtoNIN1在毛白杨的根、茎、叶及成熟叶同样被检测出具有较高的表达量,表明了其在毛白杨生长发育过程中的重要性,然而不同于草莓(Fragaria × ananassa)[42]和水稻[43]中的研究,PtoNIN1在根中的表达量最高,高于成熟叶。此外,AtINVA/C/H在拟南芥中被证实参与了植物的花发育过程,其突变体均延迟了植物的开花时间[11, 40]。本研究中我们对PtoNIN1在毛白杨雌雄花序发育过程中的作用进行了探究,PtoNIN1在雌雄花序的发育过程中具有相似的表达趋势,其在成花诱导期具有显著的高表达,随后保持了一个相对稳定的表达趋势,我们推测PtoNIN1可能对于雌雄花芽的诱导和发育过程的维持具有重要作用。启动子区分析结果表明PtoNIN1受光照、厌氧环境、干旱、防御和胁迫等环境信号调控,此外赤霉素响应元件也存在于PtoNIN1的启动子区,与木薯 (Manihot esculenta)中MeNINV1[44]相类似。
此外,为进一步探究PtoNIN1的生物学功能,我们以模式植物拟南芥为例对其进行了过表达研究。相比于野生型,转基因植株的莲座叶、茎及角果的鲜质量均得到了显著提升。前人对拟南芥中PtoNIN1同源基因AtINVA/C/H的研究已表明,该类型基因的突变会造成了植株茎的缩短[11, 40],然而在拟南芥中过表达PtoNIN1却并未显著增强其植株高度(除Pro35S:PtoNIN1/col-2外),我们推测这可能是由于PtoNIN1和拟南芥中的同源基因发生了功能冗余。转化酶是蔗糖分解过程中的关键酶,我们同样对野生型和转基因植株间的可溶性糖含量进行了分析比较,其中PtoNIN1的过表达显著增强了茎及角果中蔗糖含量的积累,而葡萄糖和果糖的含量却并未发生明显变化,我们推测PtoNIN1可能增强了植物整体的蔗糖代谢过程,蔗糖代谢相关基因表达量验证的结果也证实了这一结论,蔗糖合酶、蔗糖磷酸化酶和蔗糖磷酸磷酸化酶相关基因的表达量在转基因株系中都得到了显著提升。
本研究通过同源克隆的方式从毛白杨基因组数据中获取碱性/中性转化酶成员PtoNIN1,剖析了PtoNIN1的基因和蛋白序列结构特征,揭示了PtoNIN1在不同器官组织和不同雌雄花芽发育时期的表达模式,同时利用模式植物拟南芥异源过表达初步探究了PtoNIN1的基因功能,为进一步深入研究杨树碱性/中性转化酶基因的遗传调控机制,揭示杨树蔗糖代谢过程奠定了前期基础。
-
![]()
图 2 气流速度场测点分布示意图
H为模型高度,取值10 cm。H is model height, 10 cm.
Figure 2. Layout diagram measuring points of the airflow velocity field
![]()
表 1 不同模型风洞内雷诺数值
Table 1 Reynolds values of different models in wind tunnels
模型高度 Model height/cm 平均风速 Mean wind speed/(m·s−1) 温度 Temperature/℃ 气压 Air pressure/hPa 雷诺数 Reynolds number 5 8 ~ 14 13 855 3.87 × 105 ~ 6.78 × 105 10 8 ~ 14 13 854 3.87 × 105 ~ 6.78 × 105 20 8 ~ 14 15 860 3.84 × 105 ~ 6.72 × 105 30 8 ~ 14 13 858 3.89 × 105 ~ 6.79 × 105  下载: 导出CSV
下载: 导出CSV
表 2 防风挡沙墙风沙流结构变化
Table 2 Wind-sand flow structure changes of retaining wall
模型与风向夹角
Angle of model and wind
direction angle/(°)指示风速
Indicated wind
speed/(m·s−1)QF/
(g·cm−2·min−1)QB/
(g·cm−2·min−1)Q0 ~ 10/QF/% Q0 ~ 10/QB/% Q10 ~ 20/QB/% Q20 ~ 30/QB/% 45 8 0.50 1.65 0.90 0.01 0.24 0.75 10 2.88 4.10 0.87 0.01 0.28 0.72 12 9.32 7.72 0.83 0.01 0.31 0.68 14 16.18 13.47 0.88 0.01 0.29 0.70 60 8 0.94 4.23 0.85 0.01 0.11 0.88 10 3.07 9.76 0.85 0.01 0.10 0.89 12 8.26 18.63 0.81 0.01 0.10 0.89 14 14.18 32.62 0.91 0.01 0.10 0.89 75 8 1.53 3.25 0.81 0.01 0.12 0.87 10 3.29 6.74 0.82 0.01 0.15 0.84 12 7.59 15.22 0.79 0.01 0.14 0.85 14 15.71 25.21 0.88 0.01 0.14 0.85 90 8 0.80 2.71 0.89 0.01 0.13 0.86 10 3.06 6.56 0.85 0.01 0.13 0.86 12 7.87 19.93 0.81 0.01 0.13 0.86 14 13.87 29.19 0.90 0.01 0.13 0.86 注:QF为模型迎风侧3H处输沙总量;QB为模型背风侧3H处输沙总量。Notes: QF is total amount of sediment transport at 3H on the windward side of the model, QB is total amount of sediment transport at 3H on the leeward side of the model.
下载: 导出CSV
表 3 防风挡沙墙阻截率/输导率
Table 3 Rejection rate/conducting rate of retaining wall
% 风速
Wind speed/(m·s−1)近地面高度层
Near ground height level/cm模型与风向夹角 Angle of model and wind direction/(°) 45 60 75 90 8 0 ~ 10 −97.34 −95.15 −97.25 −94.44 10 ~ 20 +88.14 +71.31 +30.73 +80.68 20 ~ 30 +99.04 +99.56 +99.56 +99.29 10 0 ~ 10 −98.62 −97.12 −98.75 −95.85 10 ~ 20 +73.43 +62.88 +59.86 +56.37 20 ~ 30 +97.27 +98.81 +98.81 +97.22 12 0 ~ 10 −99.14 −97.31 −99.03 −96.58 10 ~ 20 +41.55 +25.41 +36.30 +46.70 20 ~ 30 +96.23 +98.99 +99.00 +98.13 14 0 ~ 10 −99.21 −97.73 −99.54 −97.49 10 ~ 20 +54.89 +64.87 +53.62 +67.83 20 ~ 30 +98.00 +99.66 +99.66 +99.24 注:−为模型截留率,+为模型输导率。Notes: − is the model rejection rate, + is the model conducting rate.
下载: 导出CSV
-
[1] Xie S B, Qu J J, Pang Y J. Dynamic wind differences in the formation of sand hazards at high and low altitude railway sections[J]. Journal of Wind Engineering & Industrial Aerodynamics, 2017, 169: 39−46.
[2] Zhao W Z, Hu G L, Zhang Z H, et al. Shielding effect of oasis-protection systems composed of various forms of wind break on sand fixation in an arid region: a case study in the Hexi Corridor, northwest China[J]. Ecological Engineering, 2008, 33(2): 119−125. doi: 10.1016/j.ecoleng.2008.02.010
[3] 屈建军, 刘贤万, 雷加强, 等. 尼龙网栅栏防沙效应的风洞模拟实验[J]. 中国沙漠, 2001(3):62−66. doi: 10.3321/j.issn:1000-694X.2001.01.012 Qu J J, Liu X W, Lei J Q, et al. Simulation experiments on sand-arresting effect of nylon net fence in wind tunnel[J]. Journal of Desert Research, 2001(3): 62−66. doi: 10.3321/j.issn:1000-694X.2001.01.012
[4] Xiao J H, Yao Z Y, Qu J J. Influence of Golmud-Lhasa section of Qinghai-Tibet Railway on blown sand transport[J]. Chinese Geographical Science, 2015, 25: 39−50. doi: 10.1007/s11769-014-0722-1
[5] Zhang C L, Li Q, Zhou N, et al. Field observations of wind profiles and sand fluxes above the windward slope of a sand dune before and after the establishment of semi-buried straw checkerboard barriers[J]. Aeolian Research, 2016, 20: 59−70. doi: 10.1016/j.aeolia.2015.11.003
[6] Zhang S, Ding G D, Yu M H, et al. Effect of straw checkerboards on wind proofing, sand fixation, and ecological restoration in shifting sandy land[J/OL]. Environmental Research and Public Health, 2018, 15(10): 2184 [2018−10−06]. https://doi.org/10.3390/ijerph15102184.
[7] Xu X L, Zhang K L, Kong Y P, et al. Effectiveness of erosion control measures along the Qinghai-Tibet Highway, Tibet Plateau, China[J]. Transportation Research Part D: Transport and Environment, 2006, 11(4): 302−309. doi: 10.1016/j.trd.2006.06.001
[8] Dong Z, Li H L, Wang J, et al. Wind tunnel test on effect of controlling windand deposited sand of geogrid sand-barrier[J]. Journal of Soil and Water Conservation, 2007, 5: 35−39.
[9] 程建军, 蒋富强, 杨印海, 等. 戈壁铁路沿线风沙灾害特征与挡风沙措施及功效研究[J]. 中国铁道科学, 2010, 31(5):15−20. Cheng J J, Jiang F Q, Yang Y H, et al. Study on the hazard characteristics of the drifting sand along the railway in Gobi area and the efficacy of the control engineering measures[J]. China Railway Science, 2010, 31(5): 15−20.
[10] Zhang K, Qu J, Liao K, et al. Damage by wind-blown sand and its control along Qinghai-Tibet railway in China[J]. Aeolian Research, 2010, 1: 143−146. doi: 10.1016/j.aeolia.2009.10.001
[11] Cheng J J, Xue C X. The sand-damage-prevention engineering system for the railway in the desert region of the Qinghai-Tibet Plateau[J]. Journal of Wind Engineering and Industrial Aerodynamics, 2014, 125: 30−37. doi: 10.1016/j.jweia.2013.11.016
[12] 韩致文, 王涛, 董治宝, 等. 风沙危害防治的主要工程措施及其机理[J]. 地理科学进展, 2004(1):13−21. doi: 10.11820/dlkxjz.2004.01.002 Han Z W, Wang T, Dong Z B, et al. Main engineering measurements and mechanism of blown sand hazard control[J]. Progress in Geography, 2004(1): 13−21. doi: 10.11820/dlkxjz.2004.01.002
[13] Zhang C L, Zou X Y, Pan X H, et al. Near-surface airflow field and aerodynamic characteristics of the railway-protection system in the Shapotou region and their significance[J]. Journal of Arid Environments, 2007, 71: 169−187. doi: 10.1016/j.jaridenv.2007.03.006
[14] Li B, Sherman D J. Aerodynamics and morphodynamics of sand fences: a review[J]. Aeolian Research, 2015, 17: 33−48. doi: 10.1016/j.aeolia.2014.11.005
[15] Luca B, Davide F, Aandrea L G. Solid barriers for windblown sand mitigation: aerodynamic behavior and conceptual design guidelines[J]. Journal of Wind Engineering & Industrial Aerodynamics, 2018, 173: 79−90.
[16] 屈建军, 喻文波, 秦晓波. HDPE功能性固沙障防风效应试验[J]. 中国沙漠, 2014, 34(5):1185−1193. Qu J J, Yu W B, Qin X B. Wind-protecting efficiency of HDPE functional sand-fixing barriers[J]. Journal of Desert Research, 2014, 34(5): 1185−1193.
[17] 何志辉, 李生宇, 王海峰, 等. 塔克拉玛干沙漠4种结构尼龙阻沙网的防风阻沙效益对比[J]. 干旱区研究, 2014, 31(2):369−374. He Z H, Li S Y, Wang H F, et al. Comparison of wind prevention and sand blocking effects of 4 types of sand blocking nylon nets in the central Taklimakan Desert[J]. Arid Zone Research, 2014, 31(2): 369−374.
[18] 李凯崇, 石龙, 孔令伟, 等. 兰新高铁沿线不同挡沙措施防护效果评价[J]. 铁道工程学报, 2017, 34(3):11−14, 21. doi: 10.3969/j.issn.1006-2106.2017.03.003 Li K C, Shi L, Kong L W, et al. Protection effect of different kinds of sand-barriers along Lanzhou-Xinjiang High-Speed Railway[J]. Journal of Railway Engineering Society, 2017, 34(3): 11−14, 21. doi: 10.3969/j.issn.1006-2106.2017.03.003
[19] 辛文栋. 风区高速铁路路基沙害防治研究与设计[J]. 铁道工程学报, 2015, 32(10):61−65, 72. doi: 10.3969/j.issn.1006-2106.2015.10.011 Xin W D. Sand disaster prevention research and design for high speed railway subgrade in the wind zone[J]. Journal of Railway Engineering Society, 2015, 32(10): 61−65, 72. doi: 10.3969/j.issn.1006-2106.2015.10.011
[20] 袁鑫鑫, 王海峰, 雷加强, 等. 不同间距双排尼龙阻沙网防风效应的风洞模拟[J]. 中国沙漠, 2016, 36(5):1238−1246. Yuan X X, Wang H F, Lei J Q, et al. Wind tunnel simulation of windbreak effect of double-row nylon net fence with different interval[J]. Journal of Desert Research, 2016, 36(5): 1238−1246.
[21] Baines W. Effects of velocity distribution on wind loads and flow patterns on buildings [C]// Proceedings of the symposium on wind effects on buildings and structures. Teddington: National Physical Laboratories, 1963: 198−225.
[22] Good M C, Joubert P N. The form drag of two-dimensional bluff-plates immersed in turbulent boundary layers[J]. Journal of Fluid Mechanics, 1968, 31(3): 547−582. doi: 10.1017/S0022112068000327
[23] Luca B, Marko H, Lorenzo R. Windblown sand along railway infrastructures: a review of challenges and mitigation measures[J]. Journal of Wind Engineering & Industrial Aerodynamics, 2018, 177: 340−365.
[24] 张克存, 屈建军, 牛清河, 等. 青藏铁路沿线砾石方格固沙机理风洞模拟研究[J]. 地球科学进展, 2010, 25(3):284−289. Zhang K C, Qu J J, Niu Q H, et al. Simulative research on the mechanism of rocky checkerboard sand barriers along Qinghai-Tibet Railway in wind tunnel[J]. Advances in Earth Science, 2010, 25(3): 284−289.
[25] 韩致文, 陈广庭, 胡英娣, 等. 塔里木沙漠公路防沙体系建设几个问题的探讨[J]. 干旱区资源与环境, 2000, 14(2):35−40. doi: 10.3969/j.issn.1003-7578.2000.02.007 Han Z W, Chen G T, Hu Y T, et al. Discussion on problems about construction of sand-controlling systems along desert highway in Tarim Basin, Taklimakan Desert[J]. Journal of Arid Land Resources and Environment, 2000, 14(2): 35−40. doi: 10.3969/j.issn.1003-7578.2000.02.007
[26] 吴望一. 流体力学[M]. 北京: 北京大学出版社, 1982. Wu W Y. Hydromechanics[M]. Beijing: Peking University Press, 1982.
[27] Liu H Y, Hou Z F, Chen Z, et al. Effects of standing stubble on the interception of soil erosion particles[J]. Land Degradation & Development, 2019, 30(3): 1−9.
[28] Mcewan I K, Willetts B B. Adaptation of the near-surface wind to the development of sand transport[J]. Journal of Fluid Mechanics, 1993, 252: 99−101. doi: 10.1017/S0022112093003684
[29] Liu X P, Dong Z B. Experimental investigation of concentration profile of a blowing sand cloud[J]. Geomorphology, 2004, 60(3): 371−381.
[30] Dong Z B, Wang H T, Zhang X H, et al. Height profile of particle concentration in an aeolian saltating cloud: a wind tunnel investigation by PIV MSD[J]. Geophysical Research Letters, 2004, 30(19): 1−4.
[31] 李凯崇, 薛春晓, 刘贺业, 等. 不同类型挡沙措施风沙防护机理的风洞实验研究[J]. 铁道工程学报, 2015, 32(1):17−21. doi: 10.3969/j.issn.1006-2106.2015.01.004 Li K C, Xue C X, Liu H Y, et al. Wind tunnel test on sand-preventing mechanism of different kinds of sand-barriers[J]. Journal of Railway Engineering Society, 2015, 32(1): 17−21. doi: 10.3969/j.issn.1006-2106.2015.01.004
[32] 左合君. 临策铁路防沙明洞防风阻沙机理及对风沙环境的影响[D]. 呼和浩特: 内蒙古农业大学, 2013. Zuo H J. The impact of opencut tunnel on wind and sand resistance and its around environment in Lin-Ce Railway[D]. Hohhot: Inner Mongolia Agricultural University, 2013.
-
期刊类型引用(4)
1. 杨桦,李祥乾,王帆,方睿,杨伟. 长足大竹象信息素结合蛋白CbuqPBP2互作蛋白的筛选与验证. 西北农林科技大学学报(自然科学版). 2024(01): 87-97 .  百度学术
百度学术
2. 万超,张月,胡莉,伍炳华,袁媛. 茉莉花JsMYB305转录因子的原核表达及蛋白纯化. 福建农业学报. 2022(02): 164-169 . 百度学术
3. 武建颖,张燕,孙贺贺,赵玉兰,董金皋,申珅,郝志敏. 玉米大斑病菌蛋白激酶A催化亚基StPKA-C1/C2的表达与互作蛋白筛选. 农业生物技术学报. 2022(10): 1976-1986 . 百度学术
4. 胡莉,万超,张蕖,陈清西,伍炳华,袁媛. 茉莉花JsMYB305转录因子互作蛋白的筛选及验证. 福建农业学报. 2022(09): 1135-1144 . 百度学术
其他类型引用(9)











计量
- 文章访问数: 1210
- HTML全文浏览量: 392
- PDF下载量: 33
- 被引次数: 13
