
Seedling dynamics and environmental driving factors of coniferous and broadleaved mixed forest in Jiaohe, Jilin Province of northeastern China
-
摘要:目的 本文以吉林蛟河不同发育阶段针阔混交林为研究对象,对比分析幼苗密度的核密度估计曲线年际变化规律,探讨土壤因子的边际变化和土壤因子的分布变化对幼苗密度分布动态的相对影响。方法 在420 m × 520 m的中龄林样地和500 m × 840 m的成熟林样地中系统布设幼苗调查样方。利用分位数回归和反事实分解法检验环境因子在不同分位数水平上对幼苗密度的边际影响,进而明确幼苗密度在不同分位点上呈现不均衡变化的主导因素。结果 幼苗密度的核密度估计曲线呈峰值向左偏移、长尾向右侧延伸的正偏态分布。在θ = 0.90的高分位点,中龄林样地中幼苗密度变化值为−5.9,成熟林样地幼苗密度变化值为−2.6。中龄林样地在θ = 0.75时幼苗密度变化值为5.8,成熟林样地在θ = 0.50和θ = 0.75时幼苗密度变化值均为2。在高分位点上幼苗密度的更大变化反映了右单尾概率分布的不均衡性。在不同估计分位点上系数效应和协变量效应对幼苗密度分布变化的相对作用大小不同。中龄林样地中系数效应在全部估计分位点上都具有很高的解释量;成熟林样地中在θ = 0.50分位点上,协变量效应对幼苗密度变化具有89%的解释量,在其他的估计分位点上系数效应的解释量更高。因此,土壤因子分布变化对幼苗密度分布变化影响的相对作用更大,而土壤因子边际变化则是导致幼苗密度不均衡变化的主要因素。土壤中速效氮、速效磷、速效钾分布变化对每个估计分位点上幼苗密度变化的解释量大多不足30%,而土壤含水量和土壤pH值对幼苗密度变化具有相对更大的影响。结论 幼苗密度的年际变化在不同分位点上是不均衡的,在高分位点上的变化尤为明显。土壤因子的边际变化和土壤因子的分布变化共同决定了幼苗的存活动态。Abstract:Objective In this paper, taking the coniferous and broadleaved mixed forest at different development stages in Jiaohe, Jilin Province of northeastern China as research object, the interannual variation of kernel density estimation curve of seedling density was compared and analyzed. The relative effects of marginal changes of soil factors and distribution changes of soil factors on the dynamics of seedling density distribution were discussed.Method Seedling investigation sample plots were systematically arranged in 420 m × 520 m half-matured forest (HF) sample plots and 500 m × 840 m mature forest (MF) sample plots. Quantile regression and counterfactual decomposition were used to test the marginal effects of environmental factors on seedling density at different quantile levels, and then to identify the leading factors resulting in the unequal change of seedling density.Result The kernel density estimation curve of seedling density showed a positive skewed distribution with peak value shifting to the left and long tail extending to the right. At the high quantile of θ = 0.90, the change value of seedling density was −5.9 in HF sample plots and −2.6 in MF sample plots. The change value of seedling density in HF sample plot was 5.8 when θ = 0.75, and that in MF sample plot was 2 when θ = 0.50 and θ = 0.75. The greater change of seedling density at high quantile reflects the inequality of probability distribution of the right single tail. The relative effects of coefficient effect and covariant effect on seedling density distribution were different in varied estimated quantiles. The coefficient effect in HF sample plots had a high explanation on all estimated quantiles; in MF sample plots, the covariant effect had 89% explanation for the change of seedling density at θ = 0.50 quantile. In other estimated quantiles, the explanation of coefficient effect was higher. Therefore, the relative effect of distribution of soil factors on the distribution of seedling density was greater, and the marginal change of soil factors was the main factor leading to unequal change of seedling density. Most of the changes in the distribution of available nitrogen, available phosphorus and available potassium in soil explained less than 30% of the changes in seedling density at each estimated quantile, while soil water content and soil pH had a relatively greater effect on the change of seedling density.Conclusion The interannual variation of seedling density is unequal in different quantiles, especially in the high quantile. The marginal change of soil factors and the distribution of soil factors determine the survival dynamics of seedlings.
-
天然更新是森林生态系统中资源的再生产,指利用林木自身繁殖和恢复能力在林地或干扰迹地上形成新一代幼林的过程[1]。幼苗的定居和生长发育决定了森林群落演替方向和植被恢复过程[2]。植物种子从生产、扩散、萌发、幼苗定居和生长到衰老死亡,每个阶段都受到各种生物因子(如食草动物、人类活动、病原体等)和环境因子(如光照条件、土壤养分等)影响[3]。幼苗更新是植物生活史中最脆弱的时期,对环境变化反应非常敏感[4]。同时幼苗阶段也是植物生活史的重要阶段,对于森林物种更新乃至群落演替都十分重要[5]。因此,深入了解幼苗更新的限制因子是森林生态系统经营和生物多样性保护的基础[6]。
幼苗更新以及与幼苗相关的生物多样性维持机制研究是生态学研究的热点[7]。幼苗动态研究主要关注少数几个稳态影响变量,或者仅关注个别变量对幼苗更新的影响。赵雪等研究了地形因子对阔叶红松林主要树种幼苗更新的影响[8]。Lebrija-Trejos等认为幼苗存活率与同种幼苗邻体密度之间具有显著负相关关系[9]。Moran等发现夏季高温对幼苗存活具有负向影响[10]。上述研究主要关注于地形、温度以及负密度制约对幼苗更新的影响。林地土壤是幼苗生长的基质环境,土壤类型、土层厚度、养分含量以及空间分布特征等都直接作用于森林更新过程[11-13]。刘帅等发现长白山次生针阔混交林幼苗存活率与土壤含水量和土壤有效氮等显著正相关[14]。Wright等基于连续11年土壤养分添加实验发现土壤养分对幼苗生长具有强烈影响[15]。但该研究主要探究单一养分添加对苗木生长状况的影响,对幼苗更新及幼苗密度分布变化的影响因素尚未涉及。
本文旨在分析幼苗密度的核密度估计曲线年际变化规律,探讨环境因子的边际变化和环境因子的分布变化对幼苗密度分布动态的相对影响。利用分位数回归和反事实分解法检验环境因子在不同分位点上对幼苗密度的边际影响,进而分析环境因子分布的变化在幼苗密度不均衡变化中的相对作用,为温带森林生物多样性维持机制提供理论依据。
1. 研究区概况与研究方法
1.1 研究区概况与样地设置
研究地点位于吉林省林业实验区国有林保护中心(43°51′00″~44°04′48″N、127°34′48″~127°51′00″E,海拔330 ~ 1 176 m)。该区域属长白山系张广才岭西南余脉,具有受季风影响的温带大陆性气候,年平均气温为3.8 ℃, 年平均降水量为700 ~ 800 mm。7月份温度最高,月平均气温为21.7 ℃;1月份温度最低,月平均气温为–18.6 ℃。土壤类型主要为暗棕色森林土,土层厚度在20 ~ 100 cm之间。植被类型属于长白山植物区系,主要林分类型为次生针阔混交林[16]。
2009年夏季在研究区内中龄林和成熟林各建立一个大面积固定监测样地(表1)。中龄林样地大小为420 m × 520 m,海拔范围为468 ~ 519 m,胸高断面积排名前5的树种为胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandschurica)、红松(Pinus koraiensis)、色木槭(Acer mono)、春榆(Ulmus davidiana var. japonica)。成熟林样地大小为500 m ×840 m,海拔范围为459 ~ 517 m,胸高断面积排名前5的树种为色木槭、千金榆(Carpinus cordata)、红松、胡桃楸、水曲柳。
表 1 中龄林幼苗的物种组成动态Table 1. Quantitative composition of seedling species in half-matured forest (HF)树种
Tree species2012年苗数
Seedling number
in 20122013年苗数
Seedling number
in 20132012—2013年
新生苗数
Recruitment number
in 2012−20132012—2013年
死亡苗数
Dead seedling number
in 2012−20132012—2013年
死亡率
Mortality rate in
2012−2013/%2012年
重要值
Importance value
(IV) in 20122013年
重要值
IV in 2013水曲柳
Fraxinus mandschurica677 311 218 584 86.3 52.96 31.00 沙松
Abies holophylla311 45 10 276 88.7 17.87 4.72 色木槭
Acer mono118 446 409 81 68.6 13.78 38.22 白牛槭
Acer mandshuricum55 369 353 39 70.9 7.87 33.43 胡桃楸
Juglans mandshurica46 7 7 46 100.0 5.26 1.02 红松
Pinus koraiensis28 5 5 28 100.0 2.19 0.57 春榆
Ulmus davidiana var.
japonica27 32 9 4 14.8 2.28 3.60 簇毛槭
Acer barbinerve23 10 10 23 100.0 1.75 1.13 青杨
Populus cathayana23 36 22 9 39.1 1.24 1.60 紫椴
Tilia amurensis10 39 39 10 100.0 1.01 3.61 裂叶榆
Ulmus laciniata4 2 0 2 50.0 0.53 0.33 黄檗
Phellodendron amurense3 0 0 3 100.0 0.37 0.00 青楷槭
Acer tegmentosum3 23 23 3 100.0 0.37 2.51 稠李
Padus racemosa1 0 0 1 100.0 0.16 0.00 山杨
Populus davidiana1 0 0 1 100.0 0.16 0.00 鼠李
Rhamnus davurica1 0 0 1 100.0 0.16 0.00 糠椴
Tilia mandshurica0 1 1 0 0.0 0.00 0.16 千金榆
Carpinus cordata0 5 5 0 0.0 0.00 0.82 合计 Total 1 331 1 331 1 111 1 111 83.5 1.2 幼苗及环境因子调查方法
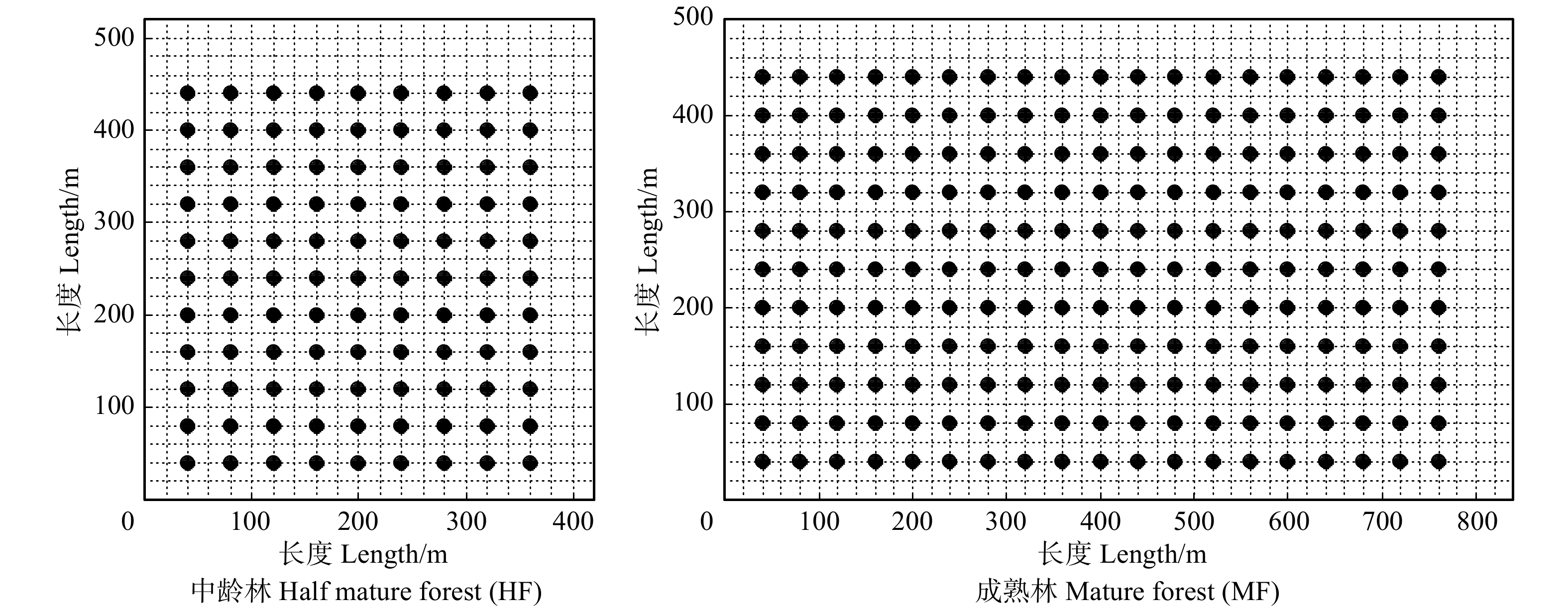
采取机械布点法在每个样地中隔40 m布设一个幼苗监测点,在中龄林设置监测点99个、成熟林209个(图1)。每个监测点分别设置1个0.5 m × 0.5 m种子承接盘和4个1 m × 1 m的幼苗调查样方,幼苗样方位于承接盘四周距离承接盘边缘1 m处,为了便于长期观测将幼苗样方的4个角用PVC管标记。本文以2012、2013年连续两年的幼苗监测数据为基础,每年7—8月份调查高度在30 cm以下幼苗的种类、高度、基径、年龄、存活状态以及相对坐标,并挂牌标记。高度为地面到幼苗最高顶芽的垂直距离,利用钢卷尺测量(精度为0.1 cm);基径为距离地面1 cm处的幼苗基部直径,利用游标卡尺测量(精度为0.01 cm);年龄分为一年生幼苗和多年生幼苗。本文中调查对象为野生实生苗,是指由种子萌发而长成的苗木,一般具有根系发达、生活力强盛、可塑性大、寿命长等特点。
分别于2012年8月和2013年8月,在承接盘周围的土壤表层(0 ~ 10 cm)各随机采集2份500 g的土样,分别测定其有机质、全氮、全磷、全钾、速效氮、速效磷、速效钾含量和pH值,两份取样和测量结果求平均值。同时用手持土壤水分测定仪测量承接盘及其对应幼苗样方表层土壤水分含量,取5次测量的平均值作为该承接盘所在观测点的土壤含水量。
幼苗重要值的计算:幼苗重要值 = (相对多度 + 相对频度 + 相对盖度)/3,式中:多度 = 样方内物种的个体数;频度 = (某物种出现的样方数/总样方数) × 100%;盖度 = (幼苗地上部分投影面积/样方面积) × 100%。
1.3 分位数回归及反事实分解
分位数回归是通过最小化残差绝对值的加权和来估计模型各参数的一种回归分析方法,传统线性回归模型描述了因变量的条件均值分布受自变量的影响过程。最小二乘法是估计传统线性回归系数的最基本方法。但在实际的经济生活中,当数据中存在严重的异方差,或后尾、尖峰情况时,分位数回归能够弥补普通最小二乘法(OLS)在回归分析中的这些缺陷。分位数回归方程是估计概率分布所有部分变化率的线性模型的扩展[17]。分位数回归更全面地反映了数据信息,可以观察到因变量分布的尾部,从而弥补最小二乘法只能估计因变量分布中心趋势的不足。本文采用该方法来探究环境因子对不同分位数水平上幼苗密度分布的影响,分位数回归系数向量反映了不同分位数水平(θ)上环境因子对幼苗密度的边际影响。利用基于分位数分布系统的反事实分解技术[18],将环境因子的边际变化和环境因子的分布变化对幼苗密度分布动态的影响分解开来。
首先构造2012年和2013年的给定自变量z的条件幼苗密度分布,再构造特定环境因子的分布与2012年相同,其他环境因子分布与2013年相一致时幼苗密度的反事实密度函数。将幼苗密度的变化分解成系数效应和协变量效应两个作用。系数效应是由分位数回归方程中估计系数的变化引起的,可用反事实密度函数与2012年幼苗密度的边际密度函数计算的基尼系数之差计算,表示由环境因子的边际变化导致的幼苗密度的不均衡变化。协变量效应是协变量变化带来的影响,可以通过2013年幼苗密度的边际密度函数与构造的反事实密度函数计算的基尼系数之差计算,表示环境因子的分布变化导致的幼苗密度出现的不均衡变化。数据分析通过R软件中McSpatial包来计算[19]。
2. 结果与分析
2.1 幼苗组成及数量特征
中龄林观测到2 662株实生苗,隶属于11科11属18种,其中水曲柳、色木槭、白牛槭和沙松的幼苗最多。2012—2013年新增幼苗1 620株,2012年观测的幼苗到2013年仅存活220株,死亡率高达83.5%(表1)。成熟林观测到8 026株实生苗,隶属于12科14属18种,其中水曲柳、色木槭、白牛槭、沙松和糠椴的幼苗最多。2012—2013年新增幼苗3 190株,2012年观测的幼苗到2013年仅存活530株,死亡率高达87.7%(表2)。
表 2 成熟林幼苗的物种组成动态Table 2. Quantitative composition of seedling species in mature forest (MF)树种
Tree species2012年苗数
Seedling number
in 20122013年苗数
Seedling number
in 20132012—2013年
新生苗
Recruitment number
in 2012−20132012—2013年
死亡苗数
Dead seedling number
in 2012−20132012—2013年
死亡率
Mortality rate in
2012−2013/%2012年
重要值
IV in 20122013年
重要值
IV in 2013水曲柳
Fraxinus mandschurica1403 537 219 1085 77.3 40.39 24.32 沙松
Abies holophylla1391 41 3 1353 97.3 29.25 2.11 色木槭
Acer mono538 1481 1402 459 85.3 22.40 50.23 白牛槭
Acer mandshuricum143 750 724 117 81.8 8.42 30.06 糠椴
Tilia mandshurica118 104 104 118 100.0 5.38 5.23 红松
Pinus koraiensis89 30 27 86 96.6 4.38 1.54 暴马丁香
Syringa reticulata264 0 0 264 100.0 3.96 0.00 紫椴
Tilia amurensis80 435 420 65 81.3 3.80 14.94 簇毛槭
Acer barbinerve131 45 30 116 88.5 3.73 2.28 裂叶榆
Ulmus laciniata29 2 0 27 93.1 1.71 0.15 胡桃楸
Juglans mandshurica27 8 5 24 88.9 1.69 0.59 千金榆
Carpinus cordata26 87 84 23 88.5 1.56 5.12 春榆
Ulmus davidiana var.
japonica24 47 27 4 16.7 1.36 2.78 青楷槭
Acer tegmentosum23 130 128 21 91.3 1.10 7.13 蒙古栎
Quercus mongolica10 3 2 9 90.0 0.71 0.22 柠筋槭
Acer triflorum6 20 15 1 16.7 0.43 1.35 毛榛
Corylus mandshurica3 0 0 3 100.0 0.09 0.00 黄檗
Phellodendron amurense1 0 0 1 100.0 0.07 0.00 合计 Total 4 306 3 720 3 190 3 776 87.7 2.2 幼苗密度动态及驱动因子
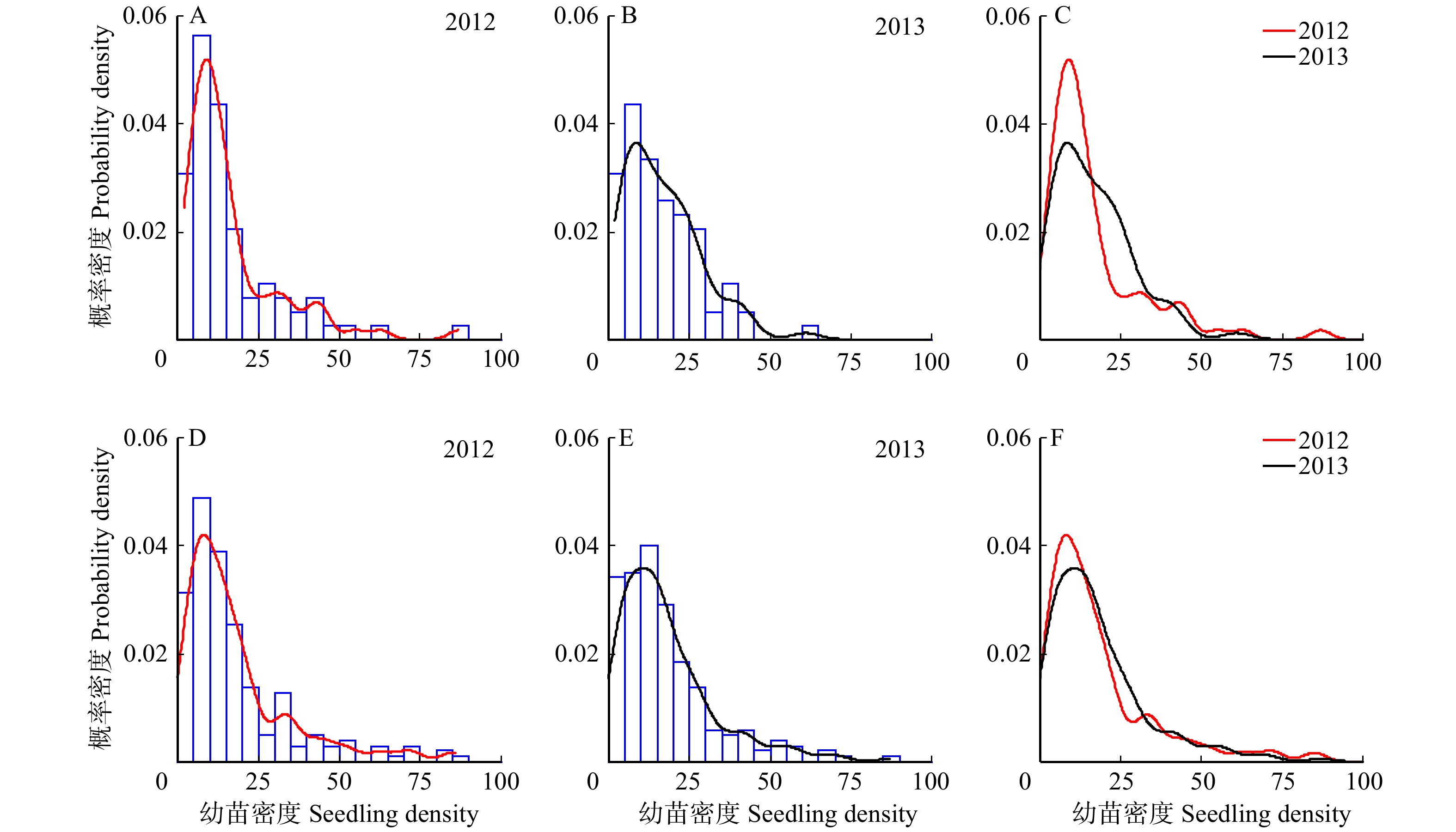
幼苗密度的概率分布在中龄林和成熟林样地中呈现类似的变化趋势。幼苗密度的核密度估计曲线呈峰值向左偏移、长尾向右侧延伸的正偏态分布。与2012年相比,2013年在低幼苗密度处的核密度估计值下降,在中等密度处的核密度估计值增大(图2)。在θ = 0.90的高分位点,中龄林样地中幼苗密度变化值为−5.9,成熟林样地幼苗密度变化值为−2.6。中龄林样地在θ = 0.75时幼苗密度变化值为5.8,成熟林样地在θ = 0.50和θ = 0.75时幼苗密度变化值均为2(表3)。这种在高分位点上幼苗密度的更大变化反映了右单尾概率分布的不均衡性。
![]() 图 2 幼苗密度的核密度估计值的年际变化A ~ C为中龄林林地,D ~ F为成熟林林地。A−C represent HF sample plot, D−F represent MF sample plot.Figure 2. Comparison of kernel density estimates of number distribution between 2012 and 2013表 3 不同分位点上幼苗密度分布变化的反事实分解Table 3. Decomposing the changes in kernel density of number at each estimated quantiles
图 2 幼苗密度的核密度估计值的年际变化A ~ C为中龄林林地,D ~ F为成熟林林地。A−C represent HF sample plot, D−F represent MF sample plot.Figure 2. Comparison of kernel density estimates of number distribution between 2012 and 2013表 3 不同分位点上幼苗密度分布变化的反事实分解Table 3. Decomposing the changes in kernel density of number at each estimated quantiles样地
Sample plot分位点
Quantile (θ)边际分布
Marginal distribution解释比例
Explained proportion单个协变量对变化量的解释比例
Proportion of overall changes explained by an individual covariate2012 2013 变化量
Change协变量效应
Covariate
effect系数效应
Coefficient
effect土壤含水量
Soil moisture
content土壤酸碱度
Soil pH碱解氮
Alkali hydrolyzed
nitrogen速效磷
Available
phosphorus速效钾
Available
potassium中龄林样地
HF sample plot0.10 4.0 4.7 0.7 0.10 0.90 0.06 0.03 0.01 0.25 0.25 7.3 7.4 0.1 0.04 0.96 0.67 0.44 0.25 0.50 11.5 15.0 3.5 0.03 0.97 0.30 0.02 0.75 18.0 23.8 5.8 0.13 0.86 0.10 0.08 0.90 37.2 31.3 −5.9 1.00 0.24 0.28 成熟林样地
MF sample plot0.10 3.0 4.0 1.0 0.04 0.96 0.33 0.73 0.30 0.04 0.25 7.0 7.2 0.2 0.07 0.93 0.34 0.98 0.57 0.49 0.04 0.50 12.0 14.0 2.0 0.89 0.11 0.31 0.75 22.0 24.0 2.0 0.04 0.96 0.27 0.48 0.98 0.68 0.07 0.90 42.2 39.6 −2.6 0.47 0.53 0.56 在不同估计分位点上系数效应和协变量效应对幼苗密度分布变化的相对作用大小不同。中龄林样地中系数效应在全部估计分位点上都具有很高的解释比例;成熟林样地中在θ = 0.50分位点上,协变量效应对幼苗密度变化具有89%的解释比例,在其他的估计分位点上系数效应的解释量更高。因此,土壤因子分布变化对幼苗密度分布变化影响的相对作用更大,而土壤因子边际变化则是导致幼苗密度不均衡变化的主要因素(图3,图4)。协变量作用之间相互抵消会导致协变量总体效应不明显,进一步分解单个协变量的作用发现,土壤中速效氮、速效磷、速效钾分布变化对幼苗密度动态的贡献较小,对每个估计分位点上幼苗密度变化的解释量大多不足30%;土壤含水量和土壤pH值对幼苗密度变化具有更高的解释量,作用效应在低分位点上表现尤为突出(表3)。
![]() 图 3 中龄林样地中幼苗密度动态与环境变量的分位数回归Figure 3. Quantile regression between seedling dynamics and environmental variables in HF sample plots
图 3 中龄林样地中幼苗密度动态与环境变量的分位数回归Figure 3. Quantile regression between seedling dynamics and environmental variables in HF sample plots![]() 图 4 成熟林样地中幼苗密度动态与环境变量的分位数回归Figure 4. Quantile regression between seedling dynamics and environmental variables in MF sample plots
图 4 成熟林样地中幼苗密度动态与环境变量的分位数回归Figure 4. Quantile regression between seedling dynamics and environmental variables in MF sample plots3. 结论与讨论
幼苗种类与乔木层树种具有较高的一致性,绝大部分幼苗为主林层的优势树种。幼苗物种组成的年际变化不明显,但不同树种幼苗密度的年际变化差异较大。比如水曲柳、沙松和红松的幼苗密度大幅度减少,而白牛槭和色木槭的幼苗密度大幅度增加。这与种子结实丰歉年[20]导致的种源限制密切相关。水曲柳和红松在观测年份结实很少甚至不结实,此时水曲柳及红松的新苗生数远低于死亡苗数;而白牛槭和色木槭观测年内大量结实,新生苗多于死亡苗。一些乔木种的繁殖体易受自然天敌(病原体和植食性动物)侵染或取食[21],种子发芽率或幼苗成活率较低,也会形成幼苗密度的迅速下降。除此之外,由于生物和环境因素波动导致的幼苗大量出生及大量死亡是决定幼苗密度动态的重要原因[22]。
土壤水分条件是影响森林生产力的主要环境因子[23-25],也是影响幼苗存活与生长的重要限制因子之一[26-27]。以往研究表明,疏花水柏枝(Myricaria laxiflora)幼苗的生长与土壤含水量的变化存在显著相关[28]。天然更新的幼苗由于根系较浅,对土壤水分含量的变化十分敏感。自然界中干旱或者降水都会对幼苗的早期存活及后续生长产生较大影响。本文中土壤含水量对中龄林和成熟林幼苗密度年际动态的影响高度一致,都表现为在低分位点的影响明显大于高分位点,因此土壤含水量变化是导致幼苗密度动态呈现不均衡的重要因子。李晗等研究发现过低或过高的土壤含水量都不利于幼苗生长,当土壤相对含水量为55% ~ 65%时光合速率最高,有利于幼苗的生长[29]。王鑫等发现土壤水分含量较高时幼苗数量会出现下降的趋势,而过低的土壤含水量也会对幼苗造成干旱胁迫影响[30],这些结果都在一定程度上支持了本文的结论。
幼苗密度的动态变化也受到土壤pH值的影响,酸性环境会增加幼苗根系质膜受损程度、减少叶绿素含量,引起光合作用减弱,进而抑制幼苗存活[31]。Dyderski等发现较低的pH值有利于云杉林更新[32],意味着弱酸性土壤环境更有利于幼苗的存活与更新。本文中土壤pH值在两个研究样地中的作用不一致,对于中龄林样地在分位点θ = 0.90高分位点上的解释量24%;对于成熟林样地在θ = 0.10和θ = 0.25低分位点上对幼苗密度的解释量在70%以上。幼苗更新密度也与土壤养分条件有关,望天树(Parashorea chinensis)幼苗更新密度与土壤氮含量和速效钾含量都呈正相关关系[33]。土壤养分条件的变化能够直接影响幼苗的存活与生长发育,幼苗根茎叶生物量分配和养分含量均与土壤养分显著正相关[34]。以往研究显示幼苗密度受土壤碱解氮和速效磷含量影响[35];不同密度的幼苗对土壤中氮磷元素的养分吸收率并不一致,当幼苗的氮磷元素吸收率减小时其密度也随之增大[36]。本文土壤养分在不同分位点上对幼苗密度的影响不同,也是导致幼苗密度边际分布出现不均衡变化的主要原因。
-
![]()
图 2 幼苗密度的核密度估计值的年际变化
A ~ C为中龄林林地,D ~ F为成熟林林地。A−C represent HF sample plot, D−F represent MF sample plot.
Figure 2. Comparison of kernel density estimates of number distribution between 2012 and 2013
![]()
图 3 中龄林样地中幼苗密度动态与环境变量的分位数回归
Figure 3. Quantile regression between seedling dynamics and environmental variables in HF sample plots
![]()
图 4 成熟林样地中幼苗密度动态与环境变量的分位数回归
Figure 4. Quantile regression between seedling dynamics and environmental variables in MF sample plots
表 1 中龄林幼苗的物种组成动态
Table 1 Quantitative composition of seedling species in half-matured forest (HF)
树种
Tree species2012年苗数
Seedling number
in 20122013年苗数
Seedling number
in 20132012—2013年
新生苗数
Recruitment number
in 2012−20132012—2013年
死亡苗数
Dead seedling number
in 2012−20132012—2013年
死亡率
Mortality rate in
2012−2013/%2012年
重要值
Importance value
(IV) in 20122013年
重要值
IV in 2013水曲柳
Fraxinus mandschurica677 311 218 584 86.3 52.96 31.00 沙松
Abies holophylla311 45 10 276 88.7 17.87 4.72 色木槭
Acer mono118 446 409 81 68.6 13.78 38.22 白牛槭
Acer mandshuricum55 369 353 39 70.9 7.87 33.43 胡桃楸
Juglans mandshurica46 7 7 46 100.0 5.26 1.02 红松
Pinus koraiensis28 5 5 28 100.0 2.19 0.57 春榆
Ulmus davidiana var.
japonica27 32 9 4 14.8 2.28 3.60 簇毛槭
Acer barbinerve23 10 10 23 100.0 1.75 1.13 青杨
Populus cathayana23 36 22 9 39.1 1.24 1.60 紫椴
Tilia amurensis10 39 39 10 100.0 1.01 3.61 裂叶榆
Ulmus laciniata4 2 0 2 50.0 0.53 0.33 黄檗
Phellodendron amurense3 0 0 3 100.0 0.37 0.00 青楷槭
Acer tegmentosum3 23 23 3 100.0 0.37 2.51 稠李
Padus racemosa1 0 0 1 100.0 0.16 0.00 山杨
Populus davidiana1 0 0 1 100.0 0.16 0.00 鼠李
Rhamnus davurica1 0 0 1 100.0 0.16 0.00 糠椴
Tilia mandshurica0 1 1 0 0.0 0.00 0.16 千金榆
Carpinus cordata0 5 5 0 0.0 0.00 0.82 合计 Total 1 331 1 331 1 111 1 111 83.5  下载: 导出CSV
下载: 导出CSV
表 2 成熟林幼苗的物种组成动态
Table 2 Quantitative composition of seedling species in mature forest (MF)
树种
Tree species2012年苗数
Seedling number
in 20122013年苗数
Seedling number
in 20132012—2013年
新生苗
Recruitment number
in 2012−20132012—2013年
死亡苗数
Dead seedling number
in 2012−20132012—2013年
死亡率
Mortality rate in
2012−2013/%2012年
重要值
IV in 20122013年
重要值
IV in 2013水曲柳
Fraxinus mandschurica1403 537 219 1085 77.3 40.39 24.32 沙松
Abies holophylla1391 41 3 1353 97.3 29.25 2.11 色木槭
Acer mono538 1481 1402 459 85.3 22.40 50.23 白牛槭
Acer mandshuricum143 750 724 117 81.8 8.42 30.06 糠椴
Tilia mandshurica118 104 104 118 100.0 5.38 5.23 红松
Pinus koraiensis89 30 27 86 96.6 4.38 1.54 暴马丁香
Syringa reticulata264 0 0 264 100.0 3.96 0.00 紫椴
Tilia amurensis80 435 420 65 81.3 3.80 14.94 簇毛槭
Acer barbinerve131 45 30 116 88.5 3.73 2.28 裂叶榆
Ulmus laciniata29 2 0 27 93.1 1.71 0.15 胡桃楸
Juglans mandshurica27 8 5 24 88.9 1.69 0.59 千金榆
Carpinus cordata26 87 84 23 88.5 1.56 5.12 春榆
Ulmus davidiana var.
japonica24 47 27 4 16.7 1.36 2.78 青楷槭
Acer tegmentosum23 130 128 21 91.3 1.10 7.13 蒙古栎
Quercus mongolica10 3 2 9 90.0 0.71 0.22 柠筋槭
Acer triflorum6 20 15 1 16.7 0.43 1.35 毛榛
Corylus mandshurica3 0 0 3 100.0 0.09 0.00 黄檗
Phellodendron amurense1 0 0 1 100.0 0.07 0.00 合计 Total 4 306 3 720 3 190 3 776 87.7
下载: 导出CSV
表 3 不同分位点上幼苗密度分布变化的反事实分解
Table 3 Decomposing the changes in kernel density of number at each estimated quantiles
样地
Sample plot分位点
Quantile (θ)边际分布
Marginal distribution解释比例
Explained proportion单个协变量对变化量的解释比例
Proportion of overall changes explained by an individual covariate2012 2013 变化量
Change协变量效应
Covariate
effect系数效应
Coefficient
effect土壤含水量
Soil moisture
content土壤酸碱度
Soil pH碱解氮
Alkali hydrolyzed
nitrogen速效磷
Available
phosphorus速效钾
Available
potassium中龄林样地
HF sample plot0.10 4.0 4.7 0.7 0.10 0.90 0.06 0.03 0.01 0.25 0.25 7.3 7.4 0.1 0.04 0.96 0.67 0.44 0.25 0.50 11.5 15.0 3.5 0.03 0.97 0.30 0.02 0.75 18.0 23.8 5.8 0.13 0.86 0.10 0.08 0.90 37.2 31.3 −5.9 1.00 0.24 0.28 成熟林样地
MF sample plot0.10 3.0 4.0 1.0 0.04 0.96 0.33 0.73 0.30 0.04 0.25 7.0 7.2 0.2 0.07 0.93 0.34 0.98 0.57 0.49 0.04 0.50 12.0 14.0 2.0 0.89 0.11 0.31 0.75 22.0 24.0 2.0 0.04 0.96 0.27 0.48 0.98 0.68 0.07 0.90 42.2 39.6 −2.6 0.47 0.53 0.56
下载: 导出CSV
-
[1] 韩有志, 王政权. 森林更新与空间异质性[J]. 应用生态学报, 2002, 13(5):615−619. doi: 10.3321/j.issn:1001-9332.2002.05.024 Han Y Z, Wang Z Q. Spatial heterogeneity and forest regeneration[J]. Chinese Journal of Applied Ecology, 2002, 13(5): 615−619. doi: 10.3321/j.issn:1001-9332.2002.05.024
[2] 潘春柳, 赖家业, 黎向东, 等. 单性木兰种子雨与天然更新的初步调查[J]. 生态学杂志, 2008, 27(12):2235−2239. Pan C L, Lai J Y, Li X D, et al. Seed rain and natural regeneration of Kmeria septentrionalis in Mulun of Guangxi[J]. Chinese Journal of Ecology, 2008, 27(12): 2235−2239.
[3] Cairns S C. Introduction to population ecology[J]. Austral Ecology, 2006, 31(7): 907−908. doi: 10.1111/j.1442-9993.2006.01674.x
[4] 彭闪江, 黄忠良, 彭少麟, 等. 植物天然更新过程中种子和幼苗死亡的影响因素[J]. 广西植物, 2004, 24(2):113−121. doi: 10.3969/j.issn.1000-3142.2004.02.004 Peng S J, Huang Z L, Peng S L, et al. Factors influencing mortality of seed and seedling in plant nature regeneration process[J]. Guihaia, 2004, 24(2): 113−121. doi: 10.3969/j.issn.1000-3142.2004.02.004
[5] Poorter L. Are species adapted to their regeneration niche, adult niche, or both[J]. American Naturalist, 2007, 169(4): 433−442. doi: 10.1086/512045
[6] 张健, 李步杭, 白雪娇, 等. 长白山阔叶红松林乔木树种幼苗组成及其年际动态[J]. 生物多样性, 2009, 17(4):385−396. doi: 10.3724/SP.J.1003.2009.09102 Zhang J, Li B H, Bai X J, et al. Composition and interannual dynamics of tree seedlings in broad-leaved Korean pine (Pinus koraiensis) mixed forest in Changbai Mountain[J]. Biodiversity Science, 2009, 17(4): 385−396. doi: 10.3724/SP.J.1003.2009.09102
[7] 姚杰, 闫琰, 张春雨, 等. 吉林蛟河针阔混交林乔木幼苗组成与月际动态[J]. 植物生态学报, 2015, 39(7):717−725. doi: 10.17521/cjpe.2015.0068 Yao J, Yan Y, Zhang C Y, et al. Composition and monthly dynamics of tree seedlings in a coniferous and broad-leaved mixed forest in Jiaohe, Jilin Province, China[J]. Chinese Journal of Plant Ecology, 2015, 39(7): 717−725. doi: 10.17521/cjpe.2015.0068
[8] 赵雪, 刘妍妍, 金光泽. 地形对阔叶红松林幼苗更新的影响[J]. 应用生态学报, 2013, 24(11):3035−3042. Zhao X, Liu Y Y, Jin G Z. Effects of topography on seedling regeneration in a mixed broadleaved-Korean pine forest in Xiaoxing’an Mountains, Northeast China[J]. Chinese Journal of Applied Ecology, 2013, 24(11): 3035−3042.
[9] Lebrija-Trejos E, Wright S J, Hernández A, et al. Does relatedness matter? Phylogenetic density-dependent survival of seedlings in a tropical forest[J]. Ecology, 2014, 95: 940−951. doi: 10.1890/13-0623.1
[10] Moran E V, Das A J, Keeley J E, et al. Negative impacts of summer heat on Sierra Nevada tree seedlings[J]. Ecosphere, 2019, 10(6): 1−22.
[11] 陈永富. 森林天然更新障碍机制研究进展[J]. 世界林业研究, 2012, 25(2):41−45. Chen Y F. Research progress of natural regeneration barrier of forest[J]. World Forestry Research, 2012, 25(2): 41−45.
[12] 康冰, 刘世荣, 蔡道雄, 等. 南亚热带不同植被恢复模式下土壤理化性质[J]. 应用生态学报, 2010, 21(10):2479−2486. Kang B, Liu S R, Cai D X, et al. Soil physical and chemical characteristics under different vegetation restoration patterns in China south subtropical area[J]. Chinese Journal of Applied Ecology, 2010, 21(10): 2479−2486.
[13] 苏松锦, 刘金福, 何中声, 等. 格氏栲天然林土壤养分空间异质性[J]. 生态学报, 2012, 32(18):5673−5682. doi: 10.5846/stxb201108041146 Su S J, Liu J F, He Z S, et al. The spatial heterogeneity of soil nutrients in a mid-subtropical Castanopsis kawakamii natural forest[J]. Acta Ecologica Sinica, 2012, 32(18): 5673−5682. doi: 10.5846/stxb201108041146
[14] 刘帅, 廖嘉星, 肖翠, 等. 长白山次生针阔混交林乔木幼苗存活的影响因素分析[J]. 植物生态学报, 2016, 40(7):711−722. doi: 10.17521/cjpe.2015.0366 Liu S, Liao J X, Xiao C, et al. Effects of biotic neighbors and habitat heterogeneity on tree seedling survival in a secondary mixed conifer and broad-leaved forest in Changbai Mountain[J]. Chinese Journal of Plant Ecology, 2016, 40(7): 711−722. doi: 10.17521/cjpe.2015.0366
[15] Wright S J, Yavitt J B, Wurzburger N, et al. Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest[J]. Ecology, 2011, 92(8): 1616−1625. doi: 10.1890/10-1558.1
[16] 闫琰, 张新娜, 姚杰, 等. 吉林蛟河不同演替阶段针阔混交林乔木幼苗数量组成及其时间动态[J]. 植物生态学报, 2016, 40(2):127−139. doi: 10.17521/cjpe.2015.0365 Yan Y, Zhang X N, Yao J, et al. Composition and temporal dynamics of tree seedlings at different successional stages of conifer and broad-leaved mixed forests in Jiaohe, Jilin Province, China[J]. Chinese Journal of Plant Ecology, 2016, 40(2): 127−139. doi: 10.17521/cjpe.2015.0365
[17] Koenker R, Bassett G. Regression quantiles[J]. Econometrica, 1978, 46: 33−50. doi: 10.2307/1913643
[18] Machado J A F, Mata J. Counterfactual decomposition of changes in wage distributions using quantile regression[J]. Journal of Applied Econometrics, 2005, 20: 445−465. doi: 10.1002/jae.788
[19] Mcmillen D. McSpatial: nonparametric spatial data analysis [Z/OL]. R package version 2.0. 2013 [2020−09−11]. http://CRANR-project.org/package=McSpatial.
[20] 樊后保, 臧润国, 李德志. 蒙古栎种群天然更新的研究[J]. 生态学杂志, 1996, 15(3):15−20. Fan H B, Zang R G, Li D Z. Natural regeneration of Mongolian oak population[J]. Chinese Journal of Ecology, 1996, 15(3): 15−20.
[21] Yan Y, Zhang C Y, Wang Y X, et al. Drivers of seedling survival in a temperate forest and their relative importance at three stages of succession[J]. Ecology and Evolution, 2015, 5: 4287−4299. doi: 10.1002/ece3.1688
[22] 尹华军, 程新颖, 赖挺, 等. 川西亚高山65年人工云杉林种子雨、种子库和幼苗定居研究[J]. 植物生态学报, 2011, 35(1):35−44. doi: 10.3724/SP.J.1258.2011.00035 Yin H J, Cheng X Y, Lai T, et al. Seed rain, soil seed bank and seedling regeneration in a 65-year Picea asperata plantation in subalpine coniferous, western Sichuan, China[J]. Chinese Journal of Plant Ecology, 2011, 35(1): 35−44. doi: 10.3724/SP.J.1258.2011.00035
[23] 郝彦宾, 王艳芬, 崔骁勇. 干旱胁迫降低了内蒙古羊草草原的碳累积[J]. 植物生态学报, 2010, 34(8):898−906. doi: 10.3773/j.issn.1005-264x.2010.08.002 Hao Y B, Wang Y F, Cui X Y. Drought stress reduces the carbon accumulation of the Leymus chinensis steppe in Inner Mongolia, China[J]. Chinese Journal of Plant Ecology, 2010, 34(8): 898−906. doi: 10.3773/j.issn.1005-264x.2010.08.002
[24] Tao F L, Hayashi Y, Zhang Z, et al. Global warming, rice production, and water use in China: developing a probabilistic assessment[J]. Agricultural and Forest Meteorology, 2008, 148(1): 94−110. doi: 10.1016/j.agrformet.2007.09.012
[25] 卢丽萍, 程丛兰, 刘伟东, 等. 30年来我国农业气象灾害对农业生产的影响及其空间分布特征[J]. 生态环境学报, 2009, 18(4):1573−1578. doi: 10.3969/j.issn.1674-5906.2009.04.062 Lu L P, Cheng C L, Liu W D, et al. Effect of the agricultural meteorological disasters on agricultural production and its spatial distribution characteristics during the last 30 years in China[J]. Ecology and Environmental Sciences, 2009, 18(4): 1573−1578. doi: 10.3969/j.issn.1674-5906.2009.04.062
[26] Morgan J M. Osmoregulation and water stress in higher plants[J]. Annual Review of Plant Physiology, 1984, 35(1): 299−319. doi: 10.1146/annurev.pp.35.060184.001503
[27] Hare P D, Cress W A, van Staden J. Dissecting the roles of osmolyte accumulation during stress[J]. Plant, Cell and Environment, 1998, 21(6): 535−553. doi: 10.1046/j.1365-3040.1998.00309.x
[28] 官守鹏, 陈芳清, 吕坤, 等. 疏花水柏枝幼苗生物量与构件对模拟土壤地下水位变化的响应[J]. 植物科学学报, 2019, 37(4):485−494. Guan S P, Chen F Q, Lü K, et al. Responses of biomass and components of Myricaria laxiflora seedlings to simulated soil water level changes[J]. Plant Science Journal, 2019, 37(4): 485−494.
[29] 李晗, 杨福孙, 李昌珍, 等. 不同土壤含水量下槟榔幼苗形态和生理特性[J]. 热带作物学报, 2020, 41(6):1132−1137. doi: 10.3969/j.issn.1000-2561.2020.06.009 Li H, Yang F S, Li C Z, et al. Morphological and physiological characteristics of Areca catechu L. seedlings under different soil moisture contents[J]. Chinese Journal of Tropical Crops, 2020, 41(6): 1132−1137. doi: 10.3969/j.issn.1000-2561.2020.06.009
[30] 王鑫, 余新晓, 贾国栋, 等. 不同土壤水分条件下侧柏幼苗的生理活动及氮素分配策略[J]. 水土保持学报, 2020, 34(3):311−317. Wang X, Yu X X, Jia G D, et al. Physiological activities and nitrogen allocation strategies of Platyccladus orientalis seedlings under different soil water conditions[J]. Journal of Soil and Water Conservation, 2020, 34(3): 311−317.
[31] 周思婕, 王平, 张敏, 等. 酸胁迫对马尾松幼苗生长及根际铝形态的影响[J]. 应用与环境生物学报, 2019, 25(6):1292−1300. Zhou S J, Wang P, Zhang M, et al. Effects of acid stress on growth and aluminum speciation in the rhizosphere of Masson pine (Pinus massoniana L.) seedlings[J]. Chinese Journal of Applied and Environmental Biology, 2019, 25(6): 1292−1300.
[32] Dyderski M, Gazda A, Hachułka M, et al. Impacts of soil conditions and light availability on natural regeneration of Norway spruce Picea abies (L.) H. Karst. in low-elevation mountain forests[J]. Annals of Forest Science, 2018, 75(4): 91. doi: 10.1007/s13595-018-0775-x
[33] 刘奎, 黄宝榴, 陈凯, 等. 望天树天然林幼苗更新及生长与环境因子的关系[J]. 湖南师范大学自然科学学报, 2018, 41(4):47−53. Liu K, Huang B L, Chen K, et al. Relationship between seedling regeneration, growth and environmental factors in Parashorea chinensis natural forest[J]. Journal of Natural Science of Hunan Normal University, 2018, 41(4): 47−53.
[34] 吴富勤, 张新军, 申仕康, 等. 土壤养分与水分对猪血木幼苗生长及生理特性的影响[J]. 广东农业科学, 2015, 20:45−51. doi: 10.3969/j.issn.1004-874X.2015.06.009 Wu F Q, Zhang X J, Shen S K, et al. Effect of soil nutrition and water content on the seedling growth and physiological characteristics of Euryodendron excelsum H. T. change[J]. Guangdong Agricultural Sciences, 2015, 20: 45−51. doi: 10.3969/j.issn.1004-874X.2015.06.009
[35] 任学敏, 杨改河, 秦晓威, 等. 巴山冷杉-牛皮桦混交林乔木更新及土壤化学性质对更新的影响[J]. 林业科学, 2012, 48(1):1−6. doi: 10.11707/j.1001-7488.20120101 Ren X M, Yang G H, Qin X W, et al. Tree regeneration in an Abies fargesii-Betula utilis mixed forest and effects of the soil chemical properties on regeneration[J]. Scientia Silvae Sinicae, 2012, 48(1): 1−6. doi: 10.11707/j.1001-7488.20120101
[36] 黄婷, 张莹, 陈建忠, 等. 育苗密度对千年桐幼苗N、P利用和生物量积累的影响[J]. 西南林业大学学报, 2013, 33(5):17−23. doi: 10.3969/j.issn.2095-1914.2013.05.003 Huang T, Zhang Y, Chen J Z, et al. Effect of seedling density on nitrogen and phosphorus utilization and biomass accumulation of Aleurites montana seedlings[J]. Journal of Southwest Forestry University, 2013, 33(5): 17−23. doi: 10.3969/j.issn.2095-1914.2013.05.003
-
期刊类型引用(2)
1. 张萌,范秀华. 吉林蛟河针阔混交林红松和紫椴个体生长的影响因素. 北京林业大学学报. 2024(09): 26-34 .  本站查看
本站查看
2. 贺翔宇,历胤男,苑飞,周新江. 三岔子林业局针阔混交林林分生长模型的构建. 防护林科技. 2022(06): 34-36 . 百度学术
其他类型引用(4)


计量
- 文章访问数: 985
- HTML全文浏览量: 379
- PDF下载量: 86
- 被引次数: 6
