
Spatio-temporal expression of candidate genes PeCFL1 and PeCFL2 in seed hair development of poplar
-
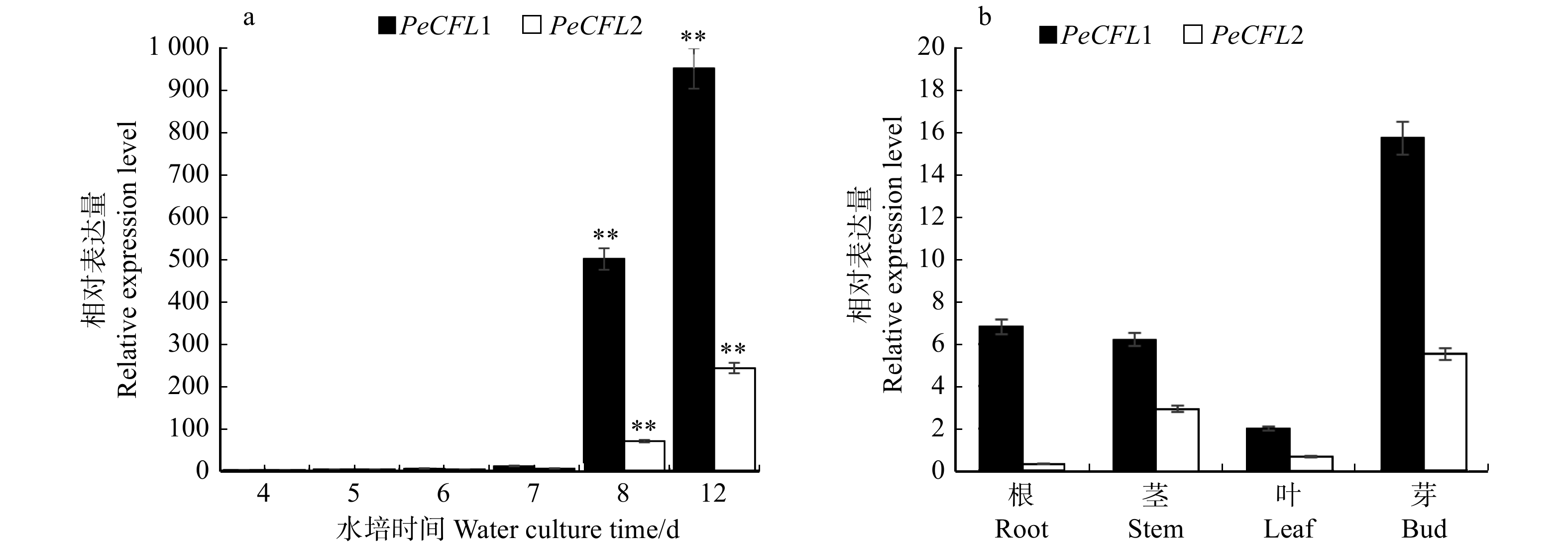
摘要:目的 杨絮是由杨树子房胎座表皮细胞发育而来的种毛,近年来已成为我国北方城市环境问题之一。目前对杨树种毛发育基因的研究还不深入,本研究将2个在杨树子房发育过程中差异性表达、且基因注释均与棉纤维发育相关的基因(PeCFL1和PeCFL2)作为种毛发育调控的候选基因,探究二者在欧美杨‘渤丰3号’中表达的时空特异性,为深入研究该基因在杨絮发育过程中的调控作用及基因工程手段改良杨树品种奠定基础。方法 取‘渤丰3号’杨越冬花枝,采集水培4、5、6、7、8、12 d的雌花序,进行固定、石蜡包埋、切片,观察子房发育及种毛的形态发生过程。采用实时定量PCR检测PeCFL1和PeCFL2基因在‘渤丰3号’杨雌花序发育过程中以及在根、茎、叶等营养器官中的表达模式。利用原位杂交技术检测PeCFL1和PeCFL2基因在杨树花器官中的组织表达特异性,揭示杨絮发育调控相关候选基因PeCFL1和PeCFL2的时空表达模式。结果 ‘渤丰3号’杨雌花枝水培12 d后,子房胎座底部出现纤维状结构,杨絮开始形成。PeCFL1和PeCFL2在‘渤丰3号’杨根、茎、叶及雌花枝腋芽中微量表达,在水培4 ~ 7 d的雌花序中表达量较少,水培8 d后表达量开始显著升高,12 d时表达量继续大幅上升,此时的石蜡切片可见子房胎座底部纤维状结构。原位杂交结果显示,PeCFL1和PeCFL2基因在杨树子房的子房壁和胎座部位表达。结论 种毛发育调控相关候选基因PeCFL1和PeCFL2在‘渤丰3号’杨雌花子房胎座底部出现纤维状结构时表达显著升高,并在胎座底部纤维状结构和子房壁中特异性表达,说明其与种毛发育调控密切相关,可作为基因工程改良杨树飞絮的目标基因。Abstract:Objective Poplar seed hair is produced by the development of epidermal cells of the placenta at the base of ovary and it has become one of the environmental problems in northern cities of China in recent years. At present, the research on poplar seed hair is not in-depth. In this study, two genes (PeCFL1 and PeCFL2), which are differentially expressed during the development of poplar ovary and related to cotton fiber development, were selected as candidate genes for the regulation of seed hair development. We studied the spatio-temporal specificity of PeCFL1 and PeCFL2 genes expressed in Populus × euramericana ‘Bofeng 3’ to lay a foundation for further study on the regulation of the two genes during the development of poplar seed hair and improvement of poplar varieties by genetic engineering.Method After 4, 5, 6, 7, 8, 12 days of hydroponic culture, the female inflorescences of Populus × euramericana ‘Bofeng 3’ were collected, fixed, paraffin embedded and sliced to observe the ovary development and the morphogenesis of poplar seed hair. Real time quantitative PCR was used to detect the expression patterns of PeCFL1 and PeCFL2 genes during the development of female inflorescence and in vegetative organs such as roots, stems and leaves of ‘Bofeng 3’ poplar. The tissue expression specificity of PeCFL1 and PeCFL2 genes in poplar flower organs was detected by in situ hybridization, and the temporal and spatial expression patterns of candidate genes PeCFL1 and PeCFL2 related to the regulation of poplar floc development were revealed.Result After 12 days of hydroponic culture, fibrous structure appeared at the bottom of ovary placentation and poplar seed hair began to form. PeCFL1 and PeCFL2 were expressed in roots, stems, leaves and axillary buds of female flower branches of ‘Bofeng 3’ poplar. For the female inflorescence of hydroponic culture, a small amount of PeCFL1 and PeCFL2 were expressed from the 4th to the 7th day, and the expression levels began to increase significantly after the 8th day. Then the expression levels continued to increase significantly on the 12th day, at the same time, the fibrous structure at the bottom of placenta could be seen in paraffin section. The results of in situ hybridization showed that PeCFL1 and PeCFL2 genes were expressed in ovary wall and placenta of poplar ovary.Conclusion The expression of PeCFL1 and PeCFL2 is significantly increased when the fibrous structure appeared at the bottom of the placentation in female flowers of Populus × euramericana ‘Bofeng 3’, and they are specifically expressed in the fibrous structure at the bottom of the placenta and the ovary wall, indicating that they are closely related to the regulation of seed hair development, and can be used as target genes for genetic engineering improvement of poplar seed hair.
-
Keywords:
- Populus × euramericana ‘Bofeng 3’ /
- poplar seed hair /
- PeCFL1 /
- PeCFL2 /
- expression analysis
-
21世纪以来,优美整洁的城镇景观和人居环境逐渐成为日本国家形象的代名词,提升着日本在国际环境领域的软实力[1]。然而,在工业化和城镇化高速发展的过程中,日本也曾出现城镇无序扩张、生态环境破坏、景观杂乱无章等问题,并导致民众对生活环境的归属感、认同感缺失[2]。20世纪70年代起,日本地方行政单位开始重视城镇景观的公共性[3]。90年代日本经济发展渐趋成熟,人们对于美好生活环境的向往愈发明确,并自发通过各种民间组织的形式参与其中。日本社会逐渐形成政府−企业−民间的三元结构,多方协同参与城镇景观建设和管理工作,在补充政府服务机能的同时,形成社会−经济−环境效益循环机制[4]。
目前国内很多城市在建设花卉小镇、特色小镇的过程中遇到了发展运营的瓶颈问题,其中一部分原因在于社会和经济的可持续性问题。小镇建设过程和运营过程中参与主体过于单一,未能形成有效的多方参与和制约机制,导致经济效益偏颇、社会效益不足,从而难以达到长期稳定的可持续发展状态。本文从多方参与的研究视角,对日本花城建设运动的多方参与模式进行梳理分析,通过聚类分析对活动的参与主体和内容进行分类总结,希望能够为我国新型城镇化建设、人居景观环境提升的多方参与途径等方面提供一些参考和借鉴。
1. 花城建设运动
1990年,以大阪国际花绿博览会为契机,民间非营利组织(non profit organization,NPO,以下简称为“民间组织”)“日本花の会”(日本花之会,以下简称为“花之会”)发起城镇景观营造运动——“花のまちづくり”(花城建设)。运动秉承博览会“自然与人类共生”的基本理念,呼吁社会各界通过花卉植物创造人与人和谐相处、花与绿相得益彰的人居环境。运动在开展初期便受到许多地方行政单位、民间组织和企业的广泛呼应,至今已连续展开近30年,活动理念逐渐渗透到社会各界,成为全日本范围内举办时间长、影响范围广、受各级行政单位支持的人居景观环境营建活动之一[5]。
运动的发展理念倡导从创造宜居生活环境、培养环保生活方式、促进地区振兴发展的3个视角出发,由个人、民间组织、地方行政单位、企业等多方主体相互协助,通过花卉植物积极进行城镇人居环境建设。同时,其发展理念还强调花城建设是基于“花卉的社会性”开展的公众参与活动。倡导参与者在营造花卉景观的过程中,不应局限于环境和美学价值,更需要将花城建设作为人−社会−花卉之间的交互平台,充分重视活动参与过程中的环境教育、人际交往、社区营造、地区振兴等社会价值[5]。
2. 多方力量支撑花城建设运动发展
2.1 民间组织作为主导力量
花之会成立于1962年,是日本排名第一、世界排名第二的重化工业器材制造公司“株式会社小松制作所”设立的财团法人,并于2009年获得认证成为公益财团法人。财团法人是出于共同目的捐助财产集合而成立的社会组织,是民间非营利组织的一种,相当于基金会。而公益财团法人是获得日本行政厅认定的财团法人,通过进行出于公益目的所进行的事业项目可以享受免税、减税等优惠政策。相反,进行非公益盈利事业则需负担高额的税费。因此,相比财团法人时期,其活动的公益性要求更加严格,如进行《公益法人认定法》规定公益事业以外的营利项目需要缴纳高额税费。
成立50年来,花之会秉承“通过花卉为人们的内心带来宁静”的创立理念,进行樱花苗木捐赠、景观环境建设、古树名木调查保护等社会环境公益活动。与其他发达国家一样,日本的NPO组织承担着完善政府服务的职能,是提升国家软实力的重要途径[6]。花之会运动推广过程中,肩负动员社会力量、协调政府民众的功能,是花城建设运动理念普及、实施推进的主导力量。
2.2 城市事件创造发展机遇
1989年,花之会在大阪国际花绿博览会的影响之下,开始关注城镇人居景观环境建设问题,并发表《欧洲和日本花城建设研究》。园艺博览会聚集了丰富的自然资源和园艺产业,传递着大量植物花卉魅力和相关资讯[7],促使日本民众的绿化意识和对花卉植物的关心重视空前高涨。因此,1990年花之会乘借博览会的事件性话题和社会影响力,正式揭开花城建设运动的序幕,在持续宣传普及博览会基本理念的基础上,提升社会大众对生活环境保护建设的自发性意识,助推城镇绿色空间和人居景观环境的建设工作。
2.3 年度活动保持参与热度
1991年起,花之会为了保持运动的社会关注度和花之会为了保持建设活动的社会关注度和影响范围,在政府和相关民间组织的支持下,每年在不同城镇轮流举办“全国花城建设年会”,由多方主体共同参与,相互交流讨论当下花城建设的热点话题和发展方向。此外,年会每年面向社会广泛征集活动事例(图1),协同政府对优秀建设活动进行表彰(“花のまちづくりコンクール”,花城建设大赛),包括花城建设大奖、农林水产大臣奖、国土交通大臣奖等主要奖项。评判标准主要关注活动在人居环境、生态保护、环境友好、地域特色、花卉特色、公众活动、地区活力等7个方面(图2)。诸如园博会这类大型城市事件结束之后,花之会通过举办年度活动继续激发公众的参与积极性,实现花城建设运动的持续性开展和普及。
2.4 社会责任提供稳定支撑
此外,支援花之会至今仍是小松制作所实现企业社会责任(corporate social responsibility,CSR)的环节之一,为花城建设活动的开展提供资金支撑。CSR可分为经济、环境和社会3大方面,其中环境方面主要体现为环境经营[8],是指在减少地球环境负荷、进行社会贡献的同时,将自发型环境保护作为企业经营战略的一部分,以提升企业的软性竞争力。环境经营理念在日本最早出现于1991年日本经济团体联合会制定《经团联地球环境宪章》中,提出为实现经济、社会可持续发展,企业应将环境策略作为存续发展的必要条件,自发进行环境保护、人居环境建设、宣传教育、民间活动、政策优化等工作。目前,许多日本企业都已将CSR 环境经营作为社会贡献和经营战略的重要部分。
花之会成立当初正值日本环境公害问题最严重的时期,在环保重压之下,重化工企业开始采取环境保护治理措施[9]。发展至今,环境经营已经从出于经济目的进行公害防治等被动环保行为,转向关注生态、人居、社会、国家等诸多层面问题的主动复合型社会行为[10]。值得一提的是,诸如花之会这类独立的民间非营利组织,并不简单等同于帮助企业实现CRS环境经营、享受税收优惠或提升企业形象的工具。相反,企业捐赠作为活动持续稳定的资金渠道之一[11],聚集其他企业、个人和民间组织的资助,支撑NPO实现改善人居环境、辅助政府工作、协调民众意愿等目标,有效发挥环境、社会和经济多重复合价值。
3. 多方参与主体的变化趋势
花城建设运动的参与主体主要分为市级、村级等行政单位、民间组织、个人和企业4种类型,伴随运动理念逐渐渗透到社会民众的生活之中,加之近30年间日本社会经济的发展变化,运动的参与主体也逐渐产生转变。由于个人也能够作为花城建设的活动主体,很难彻底掌握全日本所有的参与情况,因此将官方发布花城建设大赛的参与和获奖名单作为参考依据,对参与主体进行整理,梳理其变化趋势。
3.1 参与主体由个人转向民间组织
花城建设运动开展至今已持续了29年,花城建设年会和大赛到2018年共连续举办28届。花城建设大赛的累计报名40 015件,参与者数量整体呈增长态势(图3)。从不同主体的参与情况来看,民间组织参与最多且数量持续稳步增长,累计参与数量达到整体的59.89%。个人参与数量虽有所下降,但仍占总累计参与数量的31.62%。其次为企业(5.40%)和行政单位(3.09%),参与数量变化基本持平。
![]() 图 3 花城建设参与数量变化(1991—2018年)Figure 3. Changes in the number of floral town construction participations (1991−2018)
图 3 花城建设参与数量变化(1991—2018年)Figure 3. Changes in the number of floral town construction participations (1991−2018)花城建设运动在前期主要以政府和企业为主导力量,通过政府主导的宣传推广扩大活动影响力。随着博览会效应逐渐降温,社会关注度和活动量趋于平稳,开始进入将近20年的稳定沉淀期。当社会的公众参与机制趋于成熟,花城建设思想和公众的环境创造意识逐渐形成普遍认识之后,民间力量开始成为活动主体,且个人参与者逐渐聚集发展成民间组织,辅助政府推进人居景观环境建设工作。
3.2 获奖主体由数量平均转向民间组织
从累计640件的获奖主体来看,起初的15年间4类参与主体的获奖数量差距不大(图4)。此后,民间组织一跃成为最主要的获奖主体,且累计获奖数量最多,占整体的40.16%,但是其平均获奖比率仅有1.07%。个人获奖者的数量虽有波动但基本持平,占整体的26.25%,获奖比率也仅为1.33%。企业(17.34%)和行政单位(16.25%)的累积获奖数量最少,且近10年来略微下降,但获奖比率分别高达8.41%和5.13%。由此可见,花城建设运动对与行政单位和企业参与的鼓励程度高于民间组织和个人,但民间组织和个人的参与基数却远远高于另外两类。从侧面反映出,日本在景观环境等软实力方面的推动主力已由政府转向民间,形成民间力量主导人居环境建设的社会发展趋势。
![]() 图 4 花城建设获奖数量变化(1991—2018年)Figure 4. Changes in the number of floral town construction awards (1991−2018)
图 4 花城建设获奖数量变化(1991—2018年)Figure 4. Changes in the number of floral town construction awards (1991−2018)4. 花城建设的多方参与模式分析
由于花之会仅公开2003年以后的获奖活动资料,因此以官方公布的16届共计339条资料为依据,删除重复项整合得到279个样本。为了得出活动主体的主要花城建设模式,收集样本基本信息,整理得出环境、社会、经济方面的活动内容共计22项。将上述22项名义型变量转换为0-1变量,进行数量化处理并导入SPSS软件进行聚类分析。聚类过程使用Ward法和平方欧式距离,先将集合中每个样本自成一类,在进行类别合并时计算重心间方差,按照离差平方和增加幅度最小的顺序逐级合并[12]。Ward法具有同类离差平方和较小、类间偏差平方和较大,且使得聚类导致的类内离差平方和增量最小的特点。以平方欧式距离作为样本间距离,两样本之间的距离是两个样本在每个变量上的相应值之差的平方和,个体之间的距离越小越相似,距离越大则区别越明显。最后,以组间距离最大、分组大于3组为基准切割树状图,得到3个分组[13](图5)。
![]() 图 5 花城建设活动参与特征聚类分析Figure 5. Clustering analysis on participation features of floral town construction activities
图 5 花城建设活动参与特征聚类分析Figure 5. Clustering analysis on participation features of floral town construction activities4.1 个人、企业为主体的社会贡献型
以个人或企业为活动主体的社会贡献型花城建设模式,共包含84个样本,其中个人占71.43%、企业占20.24%、民间组织占8.33%。此类型主要表现为利用私有设施进行花坛、花卉装饰等建造活动,以实现促进地区交流、改善社区环境等作用(图5)。
4.1.1 个人公共环境意识
以个人为主体的花城建设活动,主要源于个人的社会环境意识、社会公益思想或兴趣爱好等原因,由活动主体独立出资自发进行花坛、花田、花卉装饰等景观建设。形成一定规模之后,个人逐渐与政府、民间组织、其他居民或外界专家等建立合作关系,受到政府或团体的部分活动资金补助。同时,在公众媒体的宣传及传播之下,个人的建设活动逐渐受到社会关注,起到传播花城建设的理念及思想等作用,而且通过社交媒体扩大宣传,随着参观游客的增加扩大经济收益,最终往往能够聚集更多拥有相同理念的个人形成民间非营利组织,在成员出资、捐款和政府经济补助之下进一步扩大活动的影响力[14-15](图6)。
4.1.2 企业社会环境贡献
企业为主体的花城建设活动,主要体现为企业应对社会和环境发展趋势进行花卉景观建设。由企业员工参与城镇公共景观建设,或联合政府、民间组织、居民等组织环境教育和公众活动。企业通过协助打造地域特色景观、参与城镇人居环境营造,在进行社会环境贡献的同时,履行其社会责任、提升社会信任度,塑造积极的企业形象和品牌特色,形成通过社会和环境效益达到提高经济效益的有机循环。并与当地政府、企业、居民、民间组织协同配合,开展市民园艺知识指导等社会公益活动,与相关各方主体形成协助、信赖关系,最终通过花卉景观实现企业经营体系优化(图7)。此外,根据相关政策规定,通过资助相关民间组织活动能够获得相应的税收减免优惠。例如,企业通过向国家认证的民间公共团体地区建设项目进行10万日元以上的资助,可以获得资助金额30%的税收减免[16]。而企业捐赠也作为民间活动的稳定资金渠道之一,支撑NPO实现改善人居环境、辅助政府工作、协调民众意愿等目标,有效发挥NPO活动的环境、社会和经济多重复合价值。
4.2 民间组织、政府为主体的公共活动型
以民间组织或地方行政单位为活动主体的公众活动型模式,共包含161个样本,其中民间组织占69.57%、行政单位占17.39%,此外还包含少量个人和企业主体。此类型主要表现为利用花坛、公共设施周围绿地、道路绿化等公共绿地,组织市民活动、地区交流和社区环境改善活动,通过景观建设促进地区经济活力发展(图5)。运营模式多为民间组织和地方行政单位相互协作,通过组织居民公共活动进行景观改造,根据活动的主导主体可分为民间组织主导和政府主导两种类型。
4.2.1 民间组织主导举行居民活动
以民间组织为主体的花城建设,主要体现为拥有相同景观环境意识的居民自发组成民间组织,成员之间共享一定的生态价值观和行动方略,依此实施具体的花城建设活动。其活动内容多为协助当地政府进行公共景观建设,在多方参与支持之下举办赏花节、园艺科普讲座等活动,并在当地政府的指导下协调民间景观活动与政府城镇景观建设之间的关系。通过参与体验的方式唤起更多居民的景观环境和公益贡献意识(图8)。民间组织的活动经费由成员的会费、社会和企业捐款、政府补助、财团资助、相关收入、融资和资本运作等部分组成[17]。国家、都道府县、市町村等各级政府都分别设有相应的补助规定,例如国家补助的金额上限高、申请难度大,地方政府的补助金额少、批准率高。而民间财团或基金则会公开招募特定方向的民间组织,对通过审核的组织进行资助[18]。
4.2.2 行政单位引导支持公众活动
由行政单位主导的花城建设活动,主要由政府号召民间组织和当地居民参加,在专家和学者或专业机构的指导下形成合作关系。建设活动过程中,由政府出资引导公众一起参与方案制定和活动组织,积极倡导居民进行环境志愿者、家庭垃圾交换花苗等活动;或者由政府协同当地居民团体、企业、自治会组成花城建设实施委员会,资助民间力量实施景观建设工作。一方面,政府部门持续收集活动的反馈信息,完善活动内容和合作方式,形成政府与民间协助互补的良性循环[19](图9);另一方面,政府动员民间力量协同互助形成花城建设的合作体系,通过有限的资金实现更大的社会和景观效益。
![]() 图 9 花城建设的地方行政单位参与模式Figure 9. Local government participation model of floral town construction
图 9 花城建设的地方行政单位参与模式Figure 9. Local government participation model of floral town construction4.3 学校团体为主体的环境教育型
以学校为主体的环境教育型模式包含34个样本,其中民间组织占94.12%、行政单位占5.88%。主要是在学校的出资和组织之下由学生建设管理校内外的花坛,自然环境课程与实践体验相结合。同时,周边居民和家长协助学校,逐步培养学生的感知参与意欲、理解行动能力、社会环境意识和判断表达能力[20](图10)。其主要目的在于培养学生动手能力和环境知识,形成感知自然、热爱环境的生态意识,增强人际交往能力等方面,通过环境教育带动长期的社会、环境和经济效益。活动的经费主要由学校负担,也可以向地方政府申请教育资助。一些县、市通过政府教育预算向学校提供环境教育专项资助,学校通过申请活动支出经费用于青少年环境教育,如鸟取县的“环境教育推进事业”,学校可申请活动支出经费,2017年度共计支出1 210万日元用于青少年环境教育。
5. 花城建设多方参与模式总结与借鉴
花城建设运动以城市事件为契机由民间非营利组织创设,在政府、居民、其他组织和企业的支持下得以持续开展。通过对16届279个样本的活动主体和内容进行聚类分析,得出个人和企业进行社会贡献、民间组织和行政单位合作组织公共活动、学校团体进行环境教育3种主要参与模式(图11)。
在我国,目前与1990年大阪国际花绿博览会同为国际园艺生产者协会(AIPH)批准、国际展览局(BIE)认可的A1级园艺博览会,有1999年昆明和2019年北京延庆举办的世界园艺博览会。此外,还有国家住建部联同地方政府举办的中国国际园林博览会,以及省级园博会等。诸如此类博览会的选址多位于城市建设边界或以外,大部分园区作为永久性城市绿地予以保留,少部分区域则作为社会公共服务区利用,对于城市发展建设来说,在增加绿地空间、改善公共环境等方面具有积极意义。
园艺博览会的影响并不仅仅局限于园区周边城镇,还应当承担更广域、更长远的作用,即通过城市事件向更多民众传递绿色生活理念,带动生态文明思想的快速渗透。正如日本花城建设,借助大阪国际花绿博览会带动全民参与城镇的人居环境建设,促进社会各界进行环境贡献。本文从城镇人居环境建设角度出发,从动员社会力量、多方主体协同、多方利益耦合3个方面,对多方参与途径下日本的花城建设模式进行了总结与借鉴,旨在为我国新型城镇化建设、人居景观环境提升过程中的公众参与方式提供一些参考。
(1)从社会资本向社会力量扩充,保障公利与私利的协调分配。民间力量既是花城建设活动的发源,也是实际建设的参与主体,通过城市事件激发社会参与积极性,在引入社会资本的基础上有效动员社会力量,充实了花城建设的人力资源。我国目前的小城镇规划建设过程中,以采用政府和社会资本合作(PPP)模式、租赁集体土地使用和经营权的方式为主流,当地居民作为城镇建设的主体地位较低。由此一来,在引入社会资本的过程中,难以避免企业出于利益考虑重视投资回报而忽视环境建设,同时企业的经济实力直接影响环境建设质量,甚至关乎城镇是否能真正建成落地。另一方面,当地居民从建设运营环节剥离,其福祉需求难以得到反馈和体现,公众参与缺少规范化途径和正确引导,最终导致原住民的边缘化问题。因此,在规划建设中需要由当地政府支持政策创新,平衡公共利益与企业盈利间的利益分配。通过专项补助、公众活动等方式,引入从社会资本到社会力量的全方位资源,拓展公共参与的机制和渠道,有效落实以人为本的规划建设目标、补充政府的环境服务职能。
(2)从单方付出向多方合作转变,构建政府引导、协作有序的主体协同关系。花城建设活动的主体包含政府、企业、居民、民间非营利组织、学校等多元主体,不同主体间的出发点虽各不相同,但通过博览会思想的持续普及,民间形成以改善人居环境的目标共识,利用私有或公有土地进行着不同类型的环境营造活动。我国在重视城镇产业和经济发展的同时,也需要回归到城镇作为居民生活空间的本质,切实将为居民提供美丽宜居环境作为建设首要目的之一。在现行政府主导的行政管理体制特点基础上,有效引导企业进行社会环境贡献,正确指导居民参与公共环境建设,并且可以通过引入民间组织、科研院所作为桥梁和纽带,沟通协调不同方面的意见、需求和利益矛盾。居民之间、居民与政府间沟通,则可以借鉴目前一些社区、新农村建设中采用的网格党支部等网格化管理机制,构筑“城镇—街道—楼院—楼栋”的管理层级,由基层调动基层的参与积极性,用民主方式反馈并提升居民的切身人居环境利益。最终形成多方共建、合作协助的伙伴关系,实现咨询—反馈—共识—协助—监督的良性协同机制,激发不同主体从不同出发点积极融入到同一个环境建设目标之中。
(3)从利益诉求向利益贡献升华,实现多方主体利益耦合的可持续发展模式。花城建设活动中,企业和民间组织分别通过环境营造达到促进盈利或提升地区经济活性化的经济效益,但多方主体更多地将社会、环境效益和公益贡献作为花城建设的主要目的,发挥植物景观的间接经济效益,促进地区的经济活力,这也是博览会精神得以渗透普及的体现。但是,目前国内许多小城镇、特色小镇建设PPP项目的公益性难以得到保障,其原因可以认为是由于社会公众非契约方的特点,公众利益对企业行为不具约束力。然而,实现公共价值对PPP模式下的城镇可持续发展具有不可替代的意义,公众利益的体现、当地居民的参与能够间接带动城镇经济效益提升。因此,我国在城镇建设过程中,需要通过不断完善、磨合,建立公益贡献、公众监督、社会参与的鼓励协调机制,提升多方参与主体对人居环境建设和环境公共意识的认知度与判断力,达成高层次的生态价值观共识。通过从城镇环境寻求利益诉求向主动进行利益贡献的转变与升华,形成多方主体多元利益耦合协调,最终实现以环境带动社会和经济发展的绿色可持续发展模式。
-
![]()
图 1 水培‘渤丰3号’杨雌花序的采样时期
a ~ f分别为水培第4、5、6、7、8、12 天采集的雌花序。a−f represent the female inflorescences collected on the 4th, 5th, 6th, 7th, 8th, 12th day after water culture, respectively.
Figure 1. Collection time of female inflorescences of Populus × euramericana ‘Bofeng 3’ after water culture
![]()
图 2 ‘渤丰3号’杨子房切片
a ~ e分别代表水培后第4、5、6、7、8 天采集的花序样本的切片观测结果。f、g为水培后12 d样本观测结果。Ovu、PlH、Pl分别代表胚珠、胎座毛、胎座。比例尺为200 μm。 a−e represent observations of samples collected on the 4th, 5th, 6th, 7th, 8th day after water culture, respectively. f,g are samples observed on the 12th day after water culture. Ovu, ovule; PlH, placenta hair; Pl, placenta. Bar in each photo represents 200 μm.
Figure 2. Ovary section of Populus × euramericana ‘Bofeng 3’
![]()
图 3 PeCFL1和PeCFL2在‘渤丰3号’杨不同水培时间雌花序(a)及营养器官(b)中的表达检测
**表示样本在P < 0.01水平有显著性差异。** represents significant difference at P < 0.01 level.
Figure 3. Expression of PeCFL1 and PeCFL2 in female inflorescence with different hydroponic time (a) andvegetative organs (b) of Populus × euramericana ‘Bofeng 3’
![]()
图 4 ‘渤丰3号’杨子房中PeCFL1和PeCFL2基因的原位表达情况
a为反义探针杂交结果,b为正义探针阴性对照杂交结果。a图绿色和b图红色表示杂交信号区域。a is the ovary hybridized with antisense probes; b is the ovary hybridized with sense probes (control). Green in a and red in b indicate areas of hybridization signal.
Figure 4. In situ expression of PeCFL1 and PeCFL2 in ovary of Populus × euramericana ‘Bofeng 3’
表 1 qRT-PCR引物
Table 1 Primers used in qRT-PCR analysis
基因
Gene产物长度
Product length/bp引物序列
Primer sequence用途
UsageActin 195 F:5′-AAACTGTAATGGTCCTCCCTCCG-3′ 内参
Internal referenceR:5′-GCATCATCACAATCACTCTCCGA-3′ q-PeCFL1 161 F:5′-GAGAGCACGTGGAAGACGAT-3′ qRT-PCR R:5′-AGCTAGCTTCTCTGGCTTGC-3′ q-PeCFL2 262 F:5′-CTGGAGACCACGTGGAAGAC-3′ R:5′-ACGCCTCTACTCGCTTGTTC-3′  下载: 导出CSV
下载: 导出CSV
表 2 RNA探针序列
Table 2 RNA probe sequences
基因
Gene序列
Sequence荧光标记 Fluorescence
labelingPeCFL1 5′-CAAGAGCAACAAGUACCCUUGCCAUCAUCA-3′ CY5标记(红色,阴性对照)
CY5 labeling (red, negative control)5′-UGAUGAUGGCAAGGGUACUUGUUGCUCUUG-3′ FAM标记(绿色)
FAM labeling (green)PeCFL2 5′-GGAUCAGGGAAACUCAGGAAAGAACCUUCUCCG-3′ CY5标记(红色,阴性对照)
CY5 labeling (red, negative control)5′-CGGAGAAGGUUCUUUCCUGAGUUUCCCUGAUCC-3′ FAM标记(绿色)
FAM labeling (green)
下载: 导出CSV
-
[1] Bai S, Wu H, Zhang J, et al. Genome assembly of salicaceae Populus deltoides (eastern cottonwood) I-69 based on nanopore sequencing and Hi-C technologies[J]. Journal of Heredity, 2021, 112(3): 303−310. doi: 10.1093/jhered/esab010
[2] Lin Y C, Wang J, Delhomme N, et al. Functional and evolutionary genomic inferences in Populus through genome and population sequencing of American and European aspen[J]. PNAS, 2018, 115(46): E10970−E10978.
[3] Liu Y J, Wang X R, Zeng Q Y. De novo assembly of white poplar genome and genetic diversity of white poplar population in Irtysh River Basin in China[J]. Science China Life Sciences, 2019, 62(5): 609−618. doi: 10.1007/s11427-018-9455-2
[4] Qiu D, Bai S, Ma J, et al. The genome of Populus alba × Populus tremula var. glandulosa clone 84K[J]. DNA Research, 2019, 26(5): 423−431. doi: 10.1093/dnares/dsz020
[5] Tuskan G A, Difazio S, Jansson S, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray)[J]. Science, 2006, 313: 1596−1604. doi: 10.1126/science.1128691
[6] Wu H, Yao D, Chen Y, et al. De novo genome assembly of Populus simonii further supports that Populus simonii and Populus trichocarpa belong to different sections[J]. G3 (Bethesda), 2020, 10(2): 455−466. doi: 10.1534/g3.119.400913
[7] Yang W, Wang K, Zhang J, et al. The draft genome sequence of a desert tree Populus pruinosa[J]. Gigascience, 2017, 6(9): 1−7. doi: 10.1093/gigascience/gix066
[8] Zhang Z, Chen Y, Zhang J, et al. Improved genome assembly provides new insights into genome evolution in a desert poplar (Populus euphratica)[J/OL]. Molecular Ecology Resources, 2020, 20(3): 3142[2020−11−01]. https://doi.org/10.1111/1755-0998.13142.
[9] Polle A, Janz D, Teichmann T, et al. Poplar genetic engineering: promoting desirable wood characteristics and pest resistance[J]. Applied Microbiology and Biotechnology, 2013, 97(13): 5669−5679. doi: 10.1007/s00253-013-4940-8
[10] Zhang J, Gao F, Jia H, et al. Molecular response of poplar to single and combined ozone and drought[J]. Science of the Total Environment, 2019, 655(10): 1364−1375.
[11] Radojčić R I, de Marco A, Proietti C, et al. Poplar response to cadmium and lead soil contamination[J]. Ecotoxicology and Environmental Safety, 2017, 144: 482−489. doi: 10.1016/j.ecoenv.2017.06.011
[12] Zhou X, Ren S, Lu M, et al. Preliminary study of cell wall structure and its mechanical properties of C3H and HCT RNAi transgenic poplar sapling[J/OL]. Scientific Reports, 2018, 8(1): 10508[2020−11−01]. https://doi.org/10.1038/s41598-018-28675-5.
[13] Li S, Zhang Y, Ding C, et al. Proline-rich protein gene PdPRP regulates secondary wall formation in poplar[J]. Journal of Plant Physiology, 2019, 233: 58−72. doi: 10.1016/j.jplph.2018.12.007
[14] Kaul R B. Reproductive sructure and organogenesis in a cttonwood, Populus deltoides (Salicaceae)[J]. International Journal of Plant Sciences, 1995, 156: 172−180. doi: 10.1086/297238
[15] Hsu C Y, Liu Y, Luthe D S, et al. Poplar FT2 shortens the juvenile phase and promotes seasonal flowering[J]. Plant Cell, 2006, 18(8): 1846−1861. doi: 10.1105/tpc.106.041038
[16] Ramos-Sánchez J M, Triozzi P M, Alique D, et al. LHY2 integrates night-length information to determine timing of poplar photoperiodic growth[J]. Current Biology, 2019, 29(14): 2402−2406.e4. doi: 10.1016/j.cub.2019.06.003
[17] 董源. 毛白杨胚胎学观察. 胚珠、胚囊的构造、受精作用和胚的发育[J]. 北京林学院学报, 1984(1): 83−94. Dong Y. Some embryological observations on Populus tomentosa Carr. about the structure of ovule and embryo sac, the process of fertilization and development of embryo[J]. Journal of Beijing Forestry University, 1984(1): 83−94.
[18] 朱大保. 毛白杨有性生殖能力的研究[J]. 北京林业大学学报, 1991, 12(1): 1−9. Zhu D B. Research on the reproduction capacity of P. tomentosa[J]. Journal of Beijing Forestry University, 1991, 12(1): 1−9.
[19] 张志毅, 黄智慧, 张东芳, 等. 毛白杨标本园无性系开花结实的研究[J]. 北京林业大学学报, 1992, 14(增刊3): 43−51. Zhang Z Y, Huang Z H, Zhang D F, et al. An investigation on flowering and bearing of clones in the arboretum of Populus tomentosa[J]. Journal of Beijing Forestry University, 1992, 14(Suppl.3): 43−51.
[20] 张志毅, 于雪松, 朱之梯. 三倍体毛白杨有性生殖能力的研究[J]. 北京林业大学学报, 2000, 22(6): 1−4. doi: 10.3321/j.issn:1000-1522.2000.06.001 Zhang Z Y, Yu X S, Zhu Z T. Sexual reproduction of hybrid triploids in Populus tomentosa[J]. Journal of Beijing Forestry University, 2000, 22(6): 1−4. doi: 10.3321/j.issn:1000-1522.2000.06.001
[21] 安新民, 徐昌杰, 张上隆, 等. 应用滤纸吸附一法和改进的亚克隆方法快速筛选甜橙细胞壁转化酶基因(CS-CW)[J]. 细胞生物学, 2003, 25(1): 59−62. An X M, Xu C J, Zhang S L, et al. Rapid screening of sweet orange cell wall invertase gene (CS-CW) by filter paper adsorption method and improved subcloning method[J]. Cell Biology, 2003, 25(1): 59−62.
[22] Wang R, Zhu L, Zhang Y, et al. Genome-wide analysis of poplar NF-YB gene family and identified PtNF-YB1 important in regulate flowering timing in transgenic plants[J]. BMC Plant Biology, 2019, 19(1): 251. doi: 10.1186/s12870-019-1863-2
[23] 王静澄. 毛白杨种毛形成中的蔗糖代谢与关键基因表达[D]. 北京: 北京林业大学, 2010: 25. Wang J C. Sucrose metabolism and expression profiles of regulated genes during the occurring of seed hairs of Populus tomentosa [D]. Beijing: Beijing Forestry University, 2010: 25.
[24] Campo E. Flower morphogenesis and embryogeny in a bisexual Populus deltoides[J]. Giornale Botanico Italiano, 1963, 70: 212−219. doi: 10.1080/11263506309430203
[25] Li W D, Xu H, Cheng X F, et al. The behaviour of pollen tubes on the stigma in the intersectional crosses in Populus and its relation to seed-setting[J]. Chinese Journal of Botany, 1991, 3(2): 102−109.
[26] 李文钿, 马丰山. 木本植物有性杂交生殖生物学图谱[M]. 北京: 科学出版社, 2011: 15−16. Li W T, Ma F S. Sexual hybrid reproductive Biology atlas of woody plants[M]. Beijing: Science Press, 2011: 15−16.
[27] 中国科学院植物研究所. 中国高等植物图鉴(第一册)[M]. 北京: 科学出版社, 1972: 351. Institute of Botany, Chinese Academy of Sciences. Atlas of higher plants in China (Vol. 1) [M]. Beijing: Science Press, 1972: 351.
[28] Ye M, Chen Z, Su X, et al. Study of seed hair growth in Populus tomentosa, an important character of female floral bud development[J/OL]. BMC Genomics, 2014, 15(1): 475[2020−11−01]. https://doi.org/10.1186/1471-2164-15-475.
[29] Bhosale R, Maere S, de Veylder L. Endoreplication as a potential driver of cell wall modifications[J]. Current Opinion in Plant Biology, 2019, 51: 58−65. doi: 10.1016/j.pbi.2019.04.003
[30] Meinhardt H, Gierer A. Application of a theory of biological pattern formation based on lateral inhibition[J]. Journal of Cell Science, 1974, 15(2): 321−346. doi: 10.1242/jcs.15.2.321
[31] Koch A J, Meinhardt H. Biological pattern formation: from basic mechanisms to complex structures[J]. Reviews of Modern Physics, 1994, 66: 1481−1507. doi: 10.1103/RevModPhys.66.1481
[32] Bouyer D, Geier F, Kragler F, et al. Two-dimensional patterning by a trapping/depletion mechanism: the role of TTG1 and GL3 in Arabidopsis trichome formation[J/OL]. PLoS Biology, 2008, 6(6): e141[2020−11−09]. https://doi.org/10.1371/journal.pbio.0060141.
[33] Pattanaik S, Patra B, Singh S K, et al. An overview of the gene regulatory network controlling trichome development in the model plant, Arabidopsis[J/OL]. Frontiers in Plant Science, 2014, 5(5): 259[2020−11−10]. https://doi.org/10.3389/fpls.2014.00259.
[34] 王成. 城市森林建设中的植源性污染[J]. 生态学杂志, 2003, 22(3): 32−37. doi: 10.3321/j.issn:1000-4890.2003.03.007 Wang C. Plant-caused pollution in urban forest development[J]. Chinese Journal of Ecology, 2003, 22(3): 32−37. doi: 10.3321/j.issn:1000-4890.2003.03.007
[35] 王成. 城市花粉、飞絮飞毛等植源性污染特征及其防治[J]. 中国城市林业, 2018, 16(1): 1−6. Wang C. Characteristics of plant-caused pollution and its control measures in urban area[J]. China Urban Forestry, 2018, 16(1): 1−6.
[36] 李正理. 植物组织制片学[M]. 北京: 北京大学出版社, 1996: 71−72. Li Z L. Plant tissue film making[M]. Beijing: Peking University Press, 1996: 71−72.
[37] Sun J, Niu Q W, Tarkowski P, et al. The Arabidopsis AtIPT8/PGA22 gene encodes an isopentenyl transferase that is involved in de novo cytokinin biosynthesis[J]. Plant Physiology, 2003, 131(1): 167−176. doi: 10.1104/pp.011494
[38] Guo W, Zhao J, Li X, et al. A soybean β-expansin gene GmEXPB2 intrinsically involved in root system architecture responses to abiotic stresses[J]. Plant Journal, 2011, 66(3): 541−552. doi: 10.1111/j.1365-313X.2011.04511.x
[39] Yoo M J, Wendel J F. Comparative evolutionary and developmental dynamics of the cotton (Gossypium hirsutum) fiber transcriptome[J/OL]. PLoS Genetics, 2014, 10(1): e1004073[2020−10−20]. https://doi.org/10.1371/journal.pgen.1004073.
[40] Fang L, Wang Q, Hu Y, et al. Genomic analyses in cotton identify signatures of selection and loci associated with fiber quality and yield traits[J]. Nature Genetics, 2017, 49(7): 1089−1098. doi: 10.1038/ng.3887
[41] Wu M, Li L, Liu G, et al. Differentially expressed genes between two groups of backcross inbred lines differing in fiber length developed from Upland × Pima cotton[J]. Molecular Biology Reports, 2019, 46(1): 1199−1212. doi: 10.1007/s11033-019-04589-x
[42] Zhou X, Hu W, Li B, et al. Proteomic profiling of cotton fiber developmental transition from cell elongation to secondary wall deposition[J]. Acta Biochimica et Biophysica Sinica, 2019, 51(11): 1168−1177.
[43] Lu H, Klocko A L, Brunner A M, et al. RNA interference suppression of AGAMOUS and SEEDSTICK alters floral organ identity and impairs floral organ determinacy, ovule differentiation, and seed-hair development in Populus[J]. The New phytologist, 2019, 222(2): 923−937. doi: 10.1111/nph.15648
[44] Xue L, Wu H, Chen Y, et al. Evidences for a role of two Y-specific genes in sex determination in Populus deltoides[J/OL]. Nature Communications, 2020, 11(1): 5893[2020−11−03]. https://doi.org/10.1038/s41467-020-19559-2.
-
期刊类型引用(2)
1. 尹强,阿布力克木·托合提. 多方参与风貌协调区住宅建筑设计的系统论研究. 城市建筑. 2022(21): 91-95 .  百度学术
百度学术
2. 袁奇峰,李婷,李晟霖,曾丽蓉,李刚. 破解“孤城之困”:佛山西站枢纽新城城市设计探析. 规划师. 2022(11): 147-153 . 百度学术
其他类型引用(0)














计量
- 文章访问数: 846
- HTML全文浏览量: 223
- PDF下载量: 108
- 被引次数: 2
