
Structure and dynamic characteristics of Betula albo-sinensis populations in two regions in the Qinling Mountains of northwestern China
-
摘要:目的 红桦是我国特有的桦木科高大落叶乔木,也是我国暖温带森林主要建群种之一,具有观赏、经济和生态价值。通过对秦岭两地区红桦种群结构与动态特征研究,阐明红桦种群年龄结构特征及种群数量动态变化,建立种群发展预测模型,揭示红桦种群自然更新障碍原因,为今后红桦种群的保护和恢复提供理论依据。方法 对陕西省宁陕县火地塘林场和甘肃省礼县洮坪林场红桦种群调查与统计,绘制红桦种群静态生命表,分析种群年龄结构及数量动态变化,建立种群发展预测模型,同时结合种群动态数量化法和数量时间序列分析法揭示红桦种群的发展趋势。结果 两地种群年龄结构均在Ⅲ龄级时个体数最多,种群主要由幼龄个体组成;两地种群均是前期先衰退后增长,中期短暂衰退后增长,在Ⅵ ~ Ⅶ 龄级急剧衰退;洮坪种群比火地塘种群更早进入衰退期,其种群更易受到环境因子的影响;两地红桦种群的存活曲线都趋向于Deevey-Ⅱ 型。结论 火地塘和洮坪两地红桦种群的年龄结构均呈不规则的金字塔型,种群发展主要靠中老龄个体维持;两地红桦种群现阶段都为增长型种群,且都对生境因子具有较高的敏感性,洮坪种群尤为明显;在未来两地红桦种群均不同程度地缺乏幼苗,自然更新较困难。针对此问题,提出保护和恢复红桦种群的策略:人工培育幼苗移植到种群的林窗内;对幼龄林采用“3年5次”的抚育方式,对中龄林采用整枝、修剪、除灌等方式进行抚育;对于郁闭度达0.9以上的林分,适当间伐,每公顷保留1 500株左右,提高种群的自然更新能力。Abstract:Objective Betula albo-sinensis, a tall deciduous tree of the Betulaceae, unique to our country, is one of the main constructive species of warm temperate forests in China, with ornamental, economic as well as ecological values. Through the research on the structure and dynamic characteristics of red birch populations in two regions in the Qinling Mountains of northwestern China, clarifying the age structure characteristics and population dynamic changes, establishing the population development prediction model, and revealing the reason for the natural regeneration obstacle, we provided a theoretical basis for protection and restoration.Method The two B. albo-sinensis populations in Huoditang Forest Farm (Ningshan County, Shaanxi Province) and Taoping Forest Farm (Li County, Gansu Province) were investigated and counted. Then the static life table was drawn, the population age structure and quantitative dynamic changes were analyzed, and a population development prediction model was established. Meanwhile, the development trend was revealed by the population dynamics quantification method and quantitative time series analysis method.Result The age structure of the populations in the two regions was the most individuals at age Ⅲ, and the populations were mainly composed of young individuals. They both first declined and then increased in the early stage, then increased after a short decline in the middle term, and sharply dropped off at the age of Ⅵ−Ⅶ . The Taoping population was more susceptive to environmental factors, entered the decline period earlier than the Huoditang population. Their survival curves tended to be the Deevey-Ⅱ type.Conclusion The age structure of the B. albo-sinensis populations in Huoditang and Taoping both presents irregular pyramids, and the population development is mainly maintained by middle-aged and elderly individuals. They are both growing populations at the current stage and highly dependent on habitat factors, especially obvious for Taoping. In the future, natural regeneration would be untoward for them because of lacking seedlings to various degrees. In response to these problems, several strategies to protect and restore: artificially cultivate seedlings and transplant them into the gaps of the populations; adopt the “5 times in 3 years” tending method for young forests, and the pruning, pruning, mowing, removing irrigation and the like for middle-aged forests. As for stands with a canopy density of more than 0.9, appropriate thinning is required to keep about 1 500 plants per hectare to improve the natural regeneration capacity of the populations.
-
栓皮栎(Quercus variabilis)又名软木栎,是壳斗科栎属落叶乔木,木材坚韧耐磨,纹理直,耐水湿,结构略粗,是30个主要工业用材树种之一[1]。蒙古栎(Quercus mongolica)属壳斗科栎属,落叶乔木,是国家二级珍贵树种,也是中国东北林区中主要的次生林树种。栎类在生态建设中也发挥着重要的作用。
对插穗进行黄化处理可使插穗幼化,也是一种复幼的方法,可有效促进植物生根,许多难生根树种经过黄化处理后能够生根[2]。裴东[3]在核桃(Juglans regia)无性繁殖中采用埋干黄化催芽嫩枝扦插生根的方法,扦插成活率大大提高,且操作方法比常规的繁殖方法简易。有关黄化嫩枝扦插在栎类树种上的应用未见报道。
栎树体内含有酚类等阻碍生根物质,属扦插极难生根树种,采用生长调节剂使难生根树种长出不定根是无性繁殖扦插中最常用的技术手段,使用一定浓度的植物生长调节剂处理插穗可以提高穗内某些酶活性,调节内源激素的含量,加快细胞代谢,从而促进插穗生根[4]。有关栓皮栎和蒙古栎扦插成功的报道较少[5]。闫文涛等[6]研究了不同生长调节剂对蒙古栎嫩枝扦插生根的影响,研究结果表明:1 000 mg/L的IBA处理10 s,同时和1 000 mg/L的IBA+NAA处理10 s,可有效促进愈伤组织发育,但生根率仅为3.3%。刘家财等[7]通过对弗吉尼亚栎(Quercus virginiana)的引种与扦插繁殖生产试验研究,发现采用1 000 mg/L的IBA速蘸处理的中部枝条生根效果最佳,其成活率达75.0%。吴幸连[8]对大叶栎(Quercus griffithii)扦插繁殖技术的研究结果表明:未木质化的嫩枝以 550 mg/L的IBA + 500 mg/L的ABT-1号处理最好,平均生根率高达80.3%,平均生根数达7.8条/株。吕秀立等[9]对苏玛栎(Quercus shumardii)无性繁殖的研究表明:1 ~ 3年生实生母树上的穗条有一定的扦插繁殖潜力,以0.5 mg/L的IBA浸泡1 h处理苏玛栎插穗效果最佳,生根率达到24.2%;从4 ~ 13年生苏玛栎母树上采穗扦插,并进行相同处理后则不能生根,基部腐烂且无愈伤组织产生。胡婉仪等[10]在栓皮栎、麻栎(Quercus acutissima)、小叶栎(Quercus chenii)、石栎(Lithocarpus glaber)扦插简报中写到:1年生栓皮栎生根率为56%,2年生栓皮栎生根率为44%,1年生麻栎生根率为56%,1年生小叶栎生根率31%,1年生石栎生根率为32%,但这些试验材料仅使用了生根粉处理。从以上可以看出,绝大多数的栎树扦插生根还不理想,严重影响了优良种质、优良家系和优良无性系栎树的无性扩繁和推广。目前,已有的研究仅限于未经过实施黄化处理的嫩枝所进行的扦插试验,缺乏对其枝条进行黄化处理后的扦插研究。
本研究在实生苗上部搭建遮阳网,进行遮荫,获得黄化的繁殖材料,在此基础上进行扦插试验,以期研究黄化处理对两种栎树嫩枝扦插生根的影响。为了探索黄化处理栓皮栎、蒙古栎枝条后的扦插生根效果,提高生根率,本试验从栎树枝条黄化处理、不同生长调节剂种类及其不同质量浓度3个方面,对栓皮栎、蒙古栎扦插育苗生根影响进行研究,以期筛选出科学合理的最佳处理组合,总结栓皮栎、蒙古栎扦插育苗技术,为其扦插育苗提供技术支撑和科学依据[11]。
1. 材料与方法
1.1 材 料
试验于2018年7月—2018年9月,在河北省易县洪崖山国有林场管理局七里亭示范场苗圃塑料大棚进行,以当年生栓皮栎和蒙古栎实生苗为材料,在实生苗上部搭建遮阳网,进行遮荫,对苗木进行黄化处理,黄化处理10 d后采集生长势良好,有黄化特征的且无病虫害的当年生嫩枝,修剪成长7 ~ 8 cm的插穗,每个插穗预留2 ~ 3片叶,每片叶剪去2/3,预留3个以上腋芽,上切口在距上端芽1 cm处平切,下切口距下端芽0.5 cm处斜切。扦插基质为颗粒状珍珠岩(直径为3 mm),并将珍珠岩装入营养杯(直径为10 cm)中备用。在带有遮阳网的塑料大棚中进行扦插。
1.2 试验设计
本试验以选育的栓皮栎、蒙古栎优良种源当年生实生苗半木质化黄化嫩枝为材料,以未黄化实生苗半木质化嫩枝为对照,选用IBA、NAA 这2种植物生长调节剂,分别设置3 000、2 000、1 500、750、600和400 mg/L共6个质量浓度,以清水处理为对照,共14个处理(包括对照),每个处理48株插穗,3次重复,共2 016株插穗。
1.3 扦插与管理
将栓皮栎和蒙古栎插穗基部1 ~ 2 cm浸入到不同质量浓度的IBA、NAA溶液中,分别浸泡30 s,进行扦插处理,插入深度为1/3插穗。插穗扦插前将珍珠岩基质用0.1%高锰酸钾溶液消毒,1 h后浇透水,扦插后略喷水至叶片不滴水为宜,每天多次喷水保湿至叶片不滴水,保持棚内空气相对湿度在80%以上,并适当遮荫。插后22 d,统计存活插穗数、总根条数、根径粗度、根长度,用电子游标卡尺测量根径粗度,并计算生根率、总生根条数、根长和根径。
1.4 数据分析
扦插22 d,调查生根率、根径粗度、总根条数、根长度等指标,采用隶属函数模糊综合评价法,对栎树不同品种不同质量浓度NAA、IBA下的单株根数、单株根长、单株根径进行分析,即依据公式生根率(%) = (生根株数/供试总数) × 100%计算得分值,然后根据平均得分值进行排序[12]。隶属函数值
U(Xj) 计算公式为:U(Xj)=(Xj−Xmin 式中:Xj为指标测定值,Xmin、Xmax为所有处理某一指标的最小值和最大值。将各处理不同指标的隶属值进行累加求其平均值,平均值越大扦插生根效果越好[13]。
2. 结果与分析
2.1 不同质量浓度NAA、IBA对栓皮栎嫩枝扦插生根的影响
2.1.1 NAA对栓皮栎黄化和未黄化嫩枝扦插生根的影响
本试验条件下,栓皮栎黄化和未黄化嫩枝扦插属于皮生根类型,NAA各质量浓度处理的栓皮栎黄化嫩枝扦插生根指标均优于对照。当质量浓度达到1 500 mg/L时,生根指标水平均达到最高,其中黄化处理的生根率、平均生根数、平均根长、平均根径分别为90.0%、(3.70 ± 0.29)条、(3.40 ± 0.41)cm、(0.20 ± 0.04)mm,比未黄化的分别高出约15.0%、2.20 条、2.20 cm、0.04 mm。当质量浓度为400 mg/L时,黄化插穗平均根长为(1.20 ± 0.50)cm;当质量浓度为600、400 mg/L时,未黄化插穗生根率为0(表1、图1、图2)。其中,用清水处理的插穗3 d后叶片开始发黄,7 d后开始出现枯萎现象,10 d后死亡,生根率为0。用NAA处理的黄化插穗18 d时可在插穗基部观察到愈伤组织比未黄化插穗提前1 d,在扦插第24 d时黄化比未黄化的各项生根指标都高,其中黄化插穗最长不定根约4.50 cm,比未黄化插穗长3 cm,但根系较白,侧根较少。黄化与未黄化插穗生根指标均随着NAA质量浓度的升高而出现先升高后降低的趋势,随着NAA质量浓度增加到2 000 mg/L时,黄化与未黄化插穗各项生根指标水平均开始下降,当NAA质量浓度增加到3 000 mg/L时,黄化与未黄化生根率均为0。4个生根指标综合比较表明:在低质量浓度时,随NAA质量浓度的增加,对栓皮栎黄化与未黄化插穗生根有促进作用,当NAA质量浓度达1 500 mg/L时,黄化与未黄化插穗生根效果最佳,是较适宜的生长调节剂浓度。
表 1 不同质量浓度NAA对栓皮栎黄化和未黄化嫩枝扦插生根的影响Table 1. Effects of different mass concentrations of NAA on the rooting of etiolated and non-etiolated shoots of Quercus variabilis处理条件
Treatment
condition黄化嫩枝
生根率
Rooting rate of etiolated shoots/%未黄化嫩枝
生根率
Rooting rate of non-etiolated shoots/%黄化嫩枝平均
生根数
Average rooting number of etiolated shoots未黄化嫩枝平均
生根数
Average rooting number of non-etiolated shoots黄化嫩枝
平均根长
Mean root length of etiolated shoots/cm未黄化嫩枝
平均根长
Mean root length of non-etiolated shoots/cm黄化嫩枝
平均根径
Mean root diameter of etiolated shoots/mm未黄化嫩枝
平均根径
Mean root diameter of non-etiolated shoots/mm清水对照
Water control0 0 0 0 0 0 0 0 3 000 mg/L NAA 0 0 0 0 0 0 0 0 2 000 mg/L NAA 20.0 16.0 1.00 ± 0.20 1.00 ± 0.1 1.40 ± 0.30 1.00 ± 0.12 0.20 ± 0.08 0.15 ± 0.09 1 500 mg/L NAA 90.0 75.0 3.70 ± 0.29 1.50 ± 0.25 3.40 ± 0.41 1.20 ± 0.31 0.20 ± 0.04 0.16 ± 0.03 750 mg/L NAA 25.0 17.0 3.00 ± 0.30 1.10 ± 0.33 2.50 ± 0.60 1.10 ± 0.40 0.20 ± 0.06 0.13 ± 0.04 600 mg/L NAA 2.0 0 1.50 ± 0.10 0 1.20 ± 0.80 0 0.20 ± 0.07 0 400 mg/L NAA 5.0 0 2.50 ± 0.50 0 1.20 ± 0.50 0 0.20 ± 0.05 0 ![]() 图 1 不同质量浓度NAA对栓皮栎黄化嫩枝扦插生根的影响Figure 1. Effects of different mass concentrations of NAA on the rooting of etiolated Quercus variabilis shoots
图 1 不同质量浓度NAA对栓皮栎黄化嫩枝扦插生根的影响Figure 1. Effects of different mass concentrations of NAA on the rooting of etiolated Quercus variabilis shoots![]() 图 2 不同质量浓度NAA对栓皮栎未黄化嫩枝扦插生根的影响Figure 2. Effects of different mass concentrations of NAA on the rooting of non-etiolated Quercus variabilis shoots
图 2 不同质量浓度NAA对栓皮栎未黄化嫩枝扦插生根的影响Figure 2. Effects of different mass concentrations of NAA on the rooting of non-etiolated Quercus variabilis shoots2.1.2 IBA对栓皮栎黄化和未黄化嫩枝扦插生根的影响
本试验条件下,栓皮栎黄化和未黄化嫩枝扦插均属于皮生根类型,IBA各质量浓度处理的栓皮栎黄化嫩枝扦插生根均优于对照,当质量浓度达到1 500 mg/L时,生根指标水平达到最高,其中黄化处理的生根率、平均生根数、平均根长、平均根径分别为100.0%、(5.20 ± 0.23)条、(5.20 ± 0.40) cm、(0.16 ± 0.02) mm,比未黄化分别高出约20.0%、1.1条、0.80 cm、0.02 mm(表2、图3、图4)。其中,用清水处理的插穗3 d后叶片开始发黄,7 d后开始出现枯萎现象,10 d后死亡,生根率为0。用IBA处理的黄化插穗17 d时可在插穗下部观察到乳白嫩根,比未黄化的提前了1 d,22 d后最长不定根约5.20 cm,比未黄化的长了0.80 cm,但根系较细长呈褐色,侧根较多。黄化和未黄化嫩枝生根指标随着IBA质量浓度的升高呈现先升高后降低的趋势。随着IBA质量浓度增加到2 000 mg/L时,各项指标水平开始下降,当IBA质量浓度增加到3 000 mg/L,生根率为0。4个生根指标综合比较表明:在低质量浓度时,随IBA质量浓度的增加,对栓皮栎黄化嫩枝插穗生根有促进作用,当IBA质量浓度为1 500 mg/L时,生根效果最佳,是较适宜的生长调节剂浓度。
表 2 不同质量浓度IBA对栓皮栎黄化和未黄化嫩枝扦插生根的影响Table 2. Effects of different mass concentrations of IBA on rooting of etiolated and non-etiolated shoots of Quercus variabilis处理条件
Treatment
condition黄化嫩枝
生根率
Rooting rate of etiolated shoots/%未黄化嫩枝
生根率
Rooting rate of non-etiolated shoots/%黄化嫩枝平均
生根数
Average rooting number of etiolated shoots未黄化嫩枝平均
生根数
Average rooting number of non-etiolated shoots黄化嫩枝
平均根长
Mean root length of etiolated shoots/cm未黄化嫩枝
平均根长
Mean root length of non-etiolated shoots/cm黄化嫩枝
平均根径
Mean root diameter of etiolated shoots/mm未黄化嫩枝
平均根径
Mean root diameter of non-etiolated shoots/mm清水对照
Water control0 0 0 0 0 0 0 0 3 000 mg/L NAA 0 0 0 0 0 0 0 0 2 000 mg/L NAA 12.5 0 2.90 ± 0.50 0 2.90 ± 0.20 0 0.12 ± 0.02 0 1 500 mg/L NAA 100.0 80.0 5.20 ± 0.23 4.10 ± 0.20 5.20 ± 0.40 4.40 ± 0.40 0.16 ± 0.02 0.14 ± 0.02 750 mg/L NAA 54.0 46.0 4.40 ± 0.34 3.20 ± 0.80 4.40 ± 0.20 3.80 ± 0.30 0.15 ± 0.04 0.14 ± 0.03 600 mg/L NAA 11.0 8.0 2.10 ± 0.50 2.90 ± 0.40 2.80 ± 0.80 1.60 ± 0.20 0.15 ± 0.10 0.13 ± 0.05 400 mg/L NAA 10.0 5.0 2.20 ± 0.70 2.20 ± 0.90 3.90 ± 0.60 2.60 ± 0.94 0.13 ± 0.03 0.13 ± 0.01 ![]() 图 3 不同质量浓度IBA对栓皮栎黄化嫩枝扦插生根的影响Figure 3. Effects of different mass concentrations of IBA on the rooting of etiolated Quercus variabilis shoots
图 3 不同质量浓度IBA对栓皮栎黄化嫩枝扦插生根的影响Figure 3. Effects of different mass concentrations of IBA on the rooting of etiolated Quercus variabilis shoots![]() 图 4 不同质量浓度IBA对栓皮栎未黄化嫩枝扦插生根的影响Figure 4. Effects of different mass concentrations of IBA on the rooting of non-etiolated Quercus variabilis shoots
图 4 不同质量浓度IBA对栓皮栎未黄化嫩枝扦插生根的影响Figure 4. Effects of different mass concentrations of IBA on the rooting of non-etiolated Quercus variabilis shoots2.2 不同质量浓度NAA、IBA对蒙古栎嫩枝扦插生根的影响
2.2.1 不同质量浓度NAA对蒙古栎黄化和未黄化嫩枝扦插生根的影响
本试验条件下,蒙古栎黄化和未黄化插穗扦插属于皮生根类型,NAA各质量浓度处理的蒙古栎黄化插穗扦插生根指标均优于对照。当质量浓度达到1 500 mg/L时,生根指标达到最高,其中黄化处理的生根率、平均生根数、平均根长、平均根径分别为43.0%、(5.50 ± 0.20)条、(3.70 ± 0.60) cm、(0.30 ± 0.04) mm,比未黄嫩枝分别高出约13.0%、3.20条、1.10 cm、0.10 mm(表3)。其中,用清水处理的插穗3 d后叶片开始发黄,7 d后开始出现枯萎现象,10 d后死亡,生根率为0(图5、图6)。用NAA处理的黄化插穗17 d时,可在插穗基部观察到愈伤组织比未黄化插穗提前了2 d;22 d时,黄化插穗最长不定根长为3.91 cm,比未黄化插穗长1.17 cm,但根系较白,侧根较少。生根指标均随着NAA质量浓度的升高呈现先升高后降低的趋势。在NAA低质量浓度(400 mg/L)处理下,对蒙古栎黄化插穗扦插生根促进效果不明显;当质量浓度达到1 500 mg/L时,明显高于其他质量浓度下的对应指标;随着NAA质量浓度增加到2 000 mg/L时,各项指标开始下降;当NAA质量浓度增加到3 000 mg/L,生根率为0。当NAA质量浓度为3 000、2 000、600、400 mg/L时,未黄化插穗生根率为0。4个生根指标综合比较表明:在低质量浓度时,随NAA质量浓度的增加,对蒙古栎黄化和未黄化插穗生根均有促进作用;当NAA质量浓度为1 500 mg/L时,生根效果最佳,是较适宜的生长调节剂浓度。
表 3 不同质量浓度NAA对蒙古栎黄化和未黄化嫩枝扦插生根的影响Table 3. Effects of different mass concentrations of NAA on rooting of etiolated and non-etiolated shoots of Quercus mongolica处理条件
Treatment
condition黄化嫩枝
生根率
Rooting rate of etiolated shoots/%未黄化嫩枝
生根率
Rooting rate of non-etiolated shoots/%黄化嫩枝平均
生根数
Average rooting number of etiolated shoots未黄化嫩枝
平均生根数
Average rooting number of non-etiolated shoots黄化嫩枝
平均根长
Mean root length of etiolated shoots/cm未黄化嫩枝
平均根长
Mean root length of non-etiolated shoots/cm黄化嫩枝
平均根径
Mean root diameter of etiolated shoots/mm未黄化嫩枝
平均根径
Mean root diameter of non-etiolated shoots/mm清水对照
Water control0 0 0 0 0 0 0 0 3 000 mg/L NAA 0 0 0 0 0 0 0 0 2 000 mg/L NAA 11.0 0 1.30 ± 0.70 0 2.20 ± 0.20 0 0.20 ± 0.02 0 1 500 mg/L NAA 43.0 30.0 5.50 ± 0.20 2.30 ± 0.10 3.70 ± 0.60 2.60 ± 0.10 0.30 ± 0.04 0.20 ± 0.01 750 mg/L NAA 27.0 5.0 4.10 ± 0.40 1.90 ± 0.40 3.60 ± 0.20 1.60 ± 0.40 0.20 ± 0.03 0.20 ± 0.02 600 mg/L NAA 12.0 0 3.10 ± 0.30 0 3.00 ± 0.18 0 0.20 ± 0.06 0 400 mg/L NAA 5.0 0 1.20 ± 0.50 0 1.10 ± 0.50 0 0.19 ± 0.07 0 ![]() 图 5 不同质量浓度NAA对蒙古栎黄化嫩枝扦插生根的影响Figure 5. Effects of different mass concentrations of NAA on the rooting of etiolated Quercus mongolica shoots
图 5 不同质量浓度NAA对蒙古栎黄化嫩枝扦插生根的影响Figure 5. Effects of different mass concentrations of NAA on the rooting of etiolated Quercus mongolica shoots![]() 图 6 不同质量浓度NAA对蒙古栎未黄化嫩枝扦插生根的影响Figure 6. Effects of different mass concentrations of NAA on the rooting of non-etiolated Quercus mongolica shoots
图 6 不同质量浓度NAA对蒙古栎未黄化嫩枝扦插生根的影响Figure 6. Effects of different mass concentrations of NAA on the rooting of non-etiolated Quercus mongolica shoots2.2.2 不同质量浓度IBA对蒙古栎黄化和未黄嫩枝扦插生根的影响
本试验条件下,蒙古栎黄化和未黄化扦插属于皮生根类型,IBA各质量浓度处理的蒙古栎黄化嫩枝扦插生根均优于对照。其中,用清水处理的插穗3 d后叶片开始发黄,7 d后开始出现枯萎现象,10 d后死亡,生根率为0。用IBA处理的黄化插穗15 d时可在插穗下部观察到乳白嫩根,22 d后最长不定根约6.82 cm(图7),但根系较细长呈褐色,侧根较多。当质量浓度达到1 500 mg/L时,生根指标水平达到最高,其中黄化处理的生根率、平均生根数、平均根长、平均根径分别为84.6%、(4.80 ± 0.60)条、(6.30 ± 0.60)cm、(0.15 ± 0.01)mm,比未黄化的分别高出约14.6%、3.00条、1.70 cm、0.01 mm(表4)。生根指标均随着IBA质量浓度的升高出现先升高后降低的趋势。随着IBA质量浓度增加到2 000 mg/L时,各项指标开始下降;当IBA质量浓度增加到3 000 mg/L,生根率为0(图8)。4个生根指标综合比较表明:在低质量浓度时,随IBA质量浓度的增加,对蒙古栎黄化和未黄化插穗生根有促进作用;当IBA质量浓度为1 500 mg/L时,生根效果最佳,是较适宜的生长调节剂浓度。
表 4 不同质量浓度IBA对蒙古栎黄化和未黄化嫩枝扦插生根的影响Table 4. Effects of different mass concentrations of IBA on rooting of etiolated and non-etiolated shoots of Quercus mongolica处理条件
Treatment
condition黄化嫩枝
生根率
Rooting rate of etiolated shoots/%未黄化嫩枝
生根率
Rooting rate of non-etiolated shoots/%黄化嫩枝平均
生根数
Average rooting number of etiolated shoots未黄化嫩枝
平均生根数
Average rooting number of non-etiolated shoots黄化嫩枝
平均根长
Mean root length of etiolated shoots/cm未黄化嫩枝
平均根长
Mean root length of non-etiolated shoots/cm黄化嫩枝
平均根径
Mean root diameter of etiolated shoots/mm未黄化嫩枝
平均根径
Mean root diameter of non-etiolated shoots/mm清水对照
Water control0 0 0 0 0 0 0 0 3 000 mg/L NAA 0 0 0 0 0 0 0 0 2 000 mg/L NAA 20.0 16.0 2.80 ± 0.26 1.0 ± 0.20 3.80 ± 0.80 2.50 ± 0.30 0.13 ± 0.05 0.13 ± 0.02 1 500 mg/L NAA 84.6 70.0 4.80 ± 0.60 1.80 ± 0.10 6.30 ± 0.60 4.60 ± 0.50 0.15 ± 0.01 0.14 ± 0.03 750 mg/L NAA 10.0 8.0 2.50 ± 0.20 1.50 ± 0.30 3.60 ± 0.50 3.20 ± 0.40 0.14 ± 0.09 0.13 ± 0.06 600 mg/L NAA 6.0 5.0 2.00 ± 0.10 1.30 ± 0.80 3.50 ± 0.30 1.10 ± 0.20 0.11 ± 0.06 0.13 ± 0.08 400 mg/L NAA 9.0 4.5 2.00 ± 0.70 1.24 ± 0.50 3.50 ± 0.40 1.00 ± 0.10 0.13 ± 0.03 0.02 ± 0.04 ![]() 图 7 不同质量浓度IBA对蒙古栎黄化嫩枝扦插生根的影响Figure 7. Effects of different mass concentrations of IBA on the rooting of etiolated Quercus mongolica shoots
图 7 不同质量浓度IBA对蒙古栎黄化嫩枝扦插生根的影响Figure 7. Effects of different mass concentrations of IBA on the rooting of etiolated Quercus mongolica shoots![]() 图 8 不同质量浓度IBA对蒙古栎未黄化嫩枝扦插生根的影响Figure 8. Effects of different mass concentrations of IBA on the rooting of non-etiolated Quercus mongolica shoots
图 8 不同质量浓度IBA对蒙古栎未黄化嫩枝扦插生根的影响Figure 8. Effects of different mass concentrations of IBA on the rooting of non-etiolated Quercus mongolica shoots3. 讨论与结论
植物扦插生根诱导是一个复杂的生理过程,它受到生理、形态、生化等多方面因素的影响,其中植物生长调节剂和插穗处理方式是关键的制约因素。本研究以选育的栓皮栎、蒙古栎优良种源当年生实生苗半木质化的黄化嫩枝为扦插材料,未黄化嫩枝作对照,选取了NAA、IBA两种外源植物生长调节剂进行研究,发现植物生长调节剂质量浓度过高或过低都会较大地影响插穗生根效果。IBA分别为2 000、1 500、750、 600、400 mg/L时处理黄化或未黄化栓皮栎、蒙古栎嫩枝的生根效果均优于NAA相对应的5个不同质量浓度处理的生根效果。因此,栓皮栎与蒙古栎黄化与未黄化嫩枝扦插最适合的生根生长调节剂为IBA,最适合的质量浓度为1 500 mg/L。外源植物生长调节剂通过调节插穗细胞新陈代谢,增加组织再生能力,提高酶活性诱导根源基的发生,从而有利于植物扦插繁殖形成不定根[14-15],但不同种类外源植物调节剂对植物插穗生根效果的影响不同。郭素娟[16]认为用植物生长调节剂处理是促进难生根树种插穗生根的重要技术手段,不仅有利于根原始体的诱导,而且能够促进不定根的发育和生长。例如,IBA在蒙古栎扦插中对蒙古栎嫩枝插穗不定根的诱导效果优于ABT1号、NAA和IBA+NAA的处理[6];IBA在国槐(Sophora japonica)嫩枝扦插生根中,生根效果显著高于NAA、ABT1号生根粉[17]。申展[18]通过对闽楠(Phoebe bournei)无性繁殖的研究发现:多年生母树上的当年生半木质化的闽楠枝条和当年实生苗茎段作为插穗,均能生根成苗,在生根率和生根时间方面均有较好的效果;采用多年生母树上的当年生木质化枝条也能生根,但是效果远不如当年生实生苗好。高茜茜等[19]以晚松(Pinus rigida)当年生实生苗嫩枝为插穗,研究不同的扦插环境和扦插基质对晚松嫩枝扦插生根的影响情况,试验结果表明:不同扦插环境和基质均可显著影响晚松扦插的生根效果,其中扦插环境以180 cm的塑料拱棚最佳,生根率为26.67%,在带有自动间歇喷雾装置的扦插池中,扦插基质以珍珠岩为最佳,生根率为28.00%[19]。由于栎树属于难生根树种,绝大多数的栎树扦插生根还不理想,本研究充分考虑了材料的幼化效应与生根困难的问题,因此对插穗选择了黄化处理,结果表明:采用1 500 mg/L的IBA速蘸30 s扦插时,无论栓皮栎还是蒙古栎,其生根率均最高,且根系发达,主根粗壮,有侧根和须根;在相同质量浓度下,NAA、IBA处理后的黄化嫩枝扦插的生根率、平均生根数和平均根长,均高于未黄化嫩枝扦插。因此,我们推测黄化处理对于生根能力的影响,可能是由于黄化处理对枝条生长的方向性和极性的改变,引起枝条内源激素分布的变化。黄化处理不仅能够抑制生根阻碍物质的生成,增强植物生根激素的活性,使叶绿素消失,组织黄化,皮层增厚,薄壁细胞增多,淀粉和生长素有所积累,单宁减少,有利于根原始体分化,而且还可以使插穗木质化速度减慢,保持组织的幼嫩性。黄化处理结合生长调节剂处理可使难生根树种获得生根能力[2],经过黄化处理的树木通常具有良好的生根能力,保持树木幼态特性,不仅对于林木生产和良种无性繁殖具有重要的实践意义,而且也对体细胞再生和生物技术在林木上应用有借鉴价值[20]。
致谢 真诚感谢李云老师和孙宇涵老师的悉心指导,感谢河北省洪崖山国有林场管理局的工作人员及师弟李健康等人给予的帮助,正是集体的努力才使得本试验进展顺利!
-
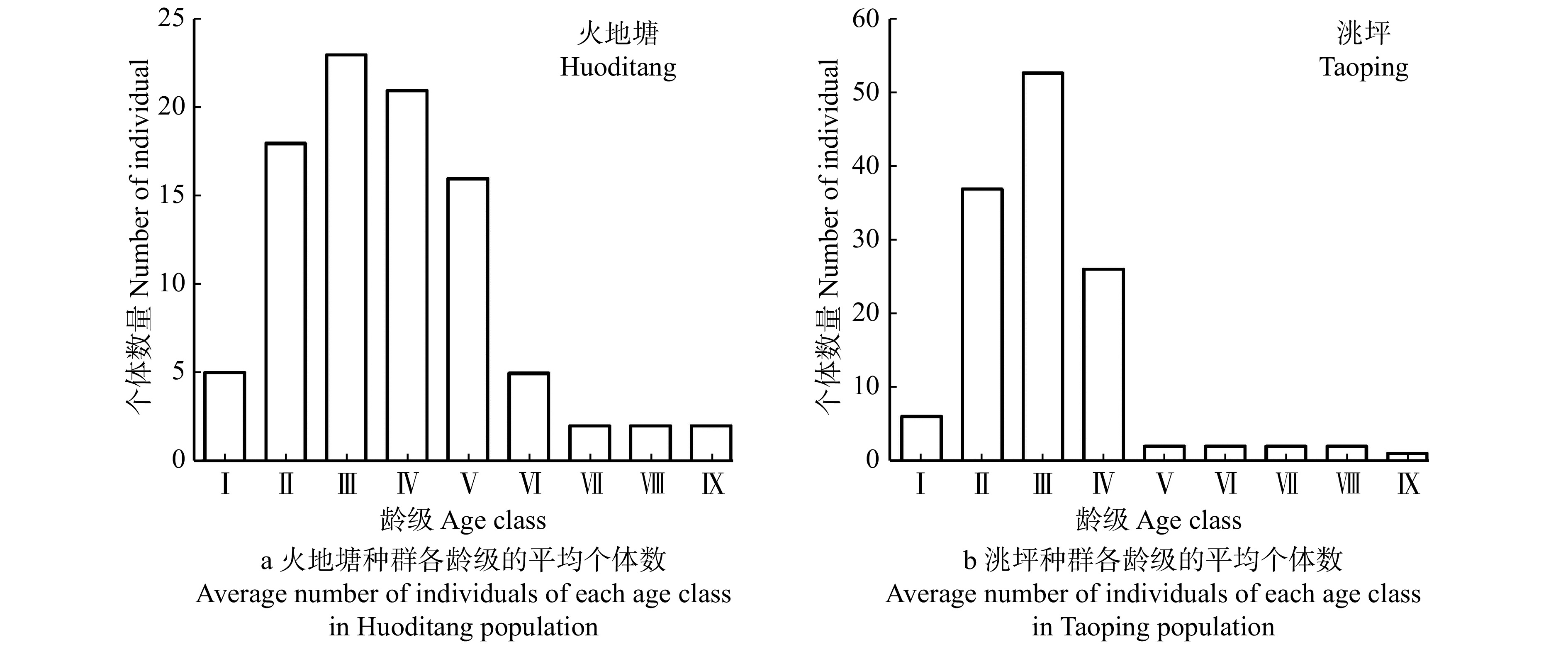
![]()
图 1 红桦种群年龄结构
Ⅰ. DBH < 2.5 cm; Ⅱ. 2.5 cm ≤ DBH < 7.5 cm; Ⅲ. 7.5 cm ≤ DBH < 12.5 cm; Ⅳ. 12.5 cm ≤ DBH < 17.5 cm; Ⅴ. 17.5 cm ≤ DBH < 22.5 cm; Ⅵ. 22.5 cm ≤ DBH < 27.5 cm; Ⅶ. 27.5 cm ≤ DBH < 32.5 cm; Ⅷ. 32.5 cm ≤ DBH < 37.5 cm; Ⅸ. DBH ≥ 37.5 cm. 下同。The same below.
Figure 1. Age structure of Betula albo-sinensis populations
![]()
图 3 红桦种群死亡率和消失率曲线
Figure 3. Mortality and vanish rate of Betula albo-sinensis populations
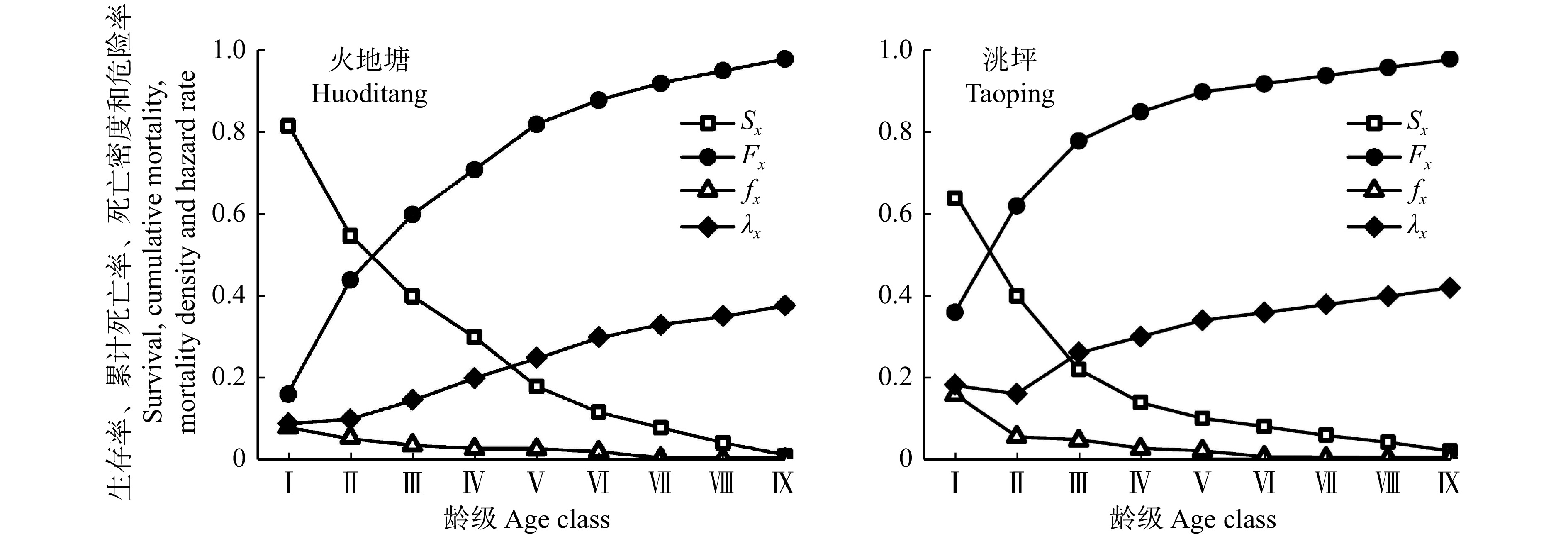
![]()
图 4 红桦种群生存率、累计死亡率曲线、死亡密度和危险率曲线
Sx. 红桦种群x龄级生存率;Fx. 红桦种群x龄级的累计死亡率;fx. 红桦种群x龄级的死亡密度;λx. 红桦种群x龄级的危险率。Sx, survival rate of Betula albo-sinensis population; Fx, fumulative death rate of Betula albo-sinensis population; fx, death density of Betula albo-sinensis population; λx, risk rate of Betula albo-sinensis population.
Figure 4. Survival rate, cumulative mortality rate, mortality density and hazard rate of Betula albo-sinensis populations
表 1 两地调查样地概况
Table 1 Survey of sample plots in the two regions
样地
Sample plot海拔
Elevation/m坡度
Slope/(°)郁闭度
Canopy closure红桦平均树高
Mean tree height of
B. albo-sinensis/m红桦平均胸径
Mean DBH of
B. albo-sinensis/cm伴生树种
Associated tree species火地塘
Huoditang2 000 ~ 2 200 8 ~ 15 0.80 9.90 ± 0.12 9.15 ± 0.03 灯台树、木姜子
Bothrocaryum controversum,
Litsea cubeba洮坪
Taoping2 100 ~ 2 600 12 ~ 23 0.85 11.12 ± 0.06 12.27 ± 0.16 荚蒾、蔷薇
Viburnum schensianum,
Rosa multifiora 下载: 导出CSV
下载: 导出CSV
表 2 红桦种群动态变化指数
Table 2 Dynamic indices of Betula albosinensis populations
种群动态指数级
Population dynamic index动态指数
Dynamic index/%火地塘
Huoditang洮坪
TaopingV1 −62.50 −98.41 V2 −46.67 −40.00 V3 6.67 65.71 V4 64.29 86.11 V5 40.00 80.00 V6 66.67 0.00 V7 0.00 0.00 V8 0.00 0.00 Vpi 16.78 36.36 {V'_{{\rm{pi}}} } 0.93 4.00 Pmax 0.06 0.11 注:Vn.种群从n到n + 1级的数量动态变化指数;Vpi.忽略外部干扰因素时整个种群结构数量动态变化指数;{V'_{{\rm{pi}}}} .考虑外部干扰因素时整个种群结构的数量动态变化指数;Pmax.随机干扰风险极大值。Notes: Vn, number dynamic change index of the population from n to n + 1; Vpi, number dynamic change index of the entire population structure when external interference factors are ignored; V′pi, quantitative dynamic change index of the entire population structure when external interference factors are considered; Pmax, the maximum value of random interference risk.
下载: 导出CSV
表 3 红桦种群静态生命表
Table 3 Static life table of Betula albosinensis populations
研究地区
Study area龄级
Age class
Ax
ax
lx
dx
lnlx
qx
Lx
Tx
ex
Kx
火地塘
HuoditangⅠ 5 27 1 000 185 6.908 0.185 907 2981 2.981 0.205 Ⅱ 18 22 815 259 6.703 0.318 685 2074 2.545 0.383 Ⅲ 23 15 556 148 6.320 0.267 481 1388 2.499 0.310 Ⅳ 21 11 407 111 6.010 0.273 352 907 2.226 0.318 Ⅴ 16 8 296 111 5.691 0.375 241 555 1.873 0.470 Ⅵ 5 5 185 74 5.221 0.400 148 314 1.697 0.511 Ⅶ 2 3 111 37 4.711 0.333 93 166 1.496 0.405 Ⅷ 2 2 74 37 4.305 0.500 56 74 0.994 0.693 Ⅸ 2 1 37 37 3.612 1.000 19 18 0.487 洮坪
TaopingⅠ 6 51 1 000 373 6.908 0.373 814 2069 2.069 0.466 Ⅱ 37 32 627 235 6.442 0.375 510 1255 2.001 0.470 Ⅲ 53 20 392 176 5.972 0.450 304 745 1.901 0.598 Ⅳ 26 11 216 78 5.374 0.364 176 442 2.047 0.452 Ⅴ 2 7 137 59 4.922 0.429 108 265 1.931 0.560 Ⅵ 2 4 78 20 4.362 0.250 69 157 2.005 0.288 Ⅶ 2 3 59 20 4.075 0.333 49 89 1.506 0.405 Ⅷ 2 2 39 20 3.669 0.500 29 40 1.010 0.693 Ⅸ 1 1 20 24 2.976 1.000 10 10 0.519 注:x.年龄级;Ax. x龄级内现有个体数;ax. 匀滑后x龄级内的存活个体数;lx.x龄级开始时标准化存活个体数;dx.从x到x + 1龄级间隔期内标准化死亡;qx.从x到x + 1龄级间隔期间死亡率;Lx.从x到x + 1龄级间隔期间还存活的个体数;Tx.从x龄级到超过x龄级的个体总数;ex.进入x龄级个体的生命期望或平均期望寿命;Kx.消失率。Notes: x, age class; Ax, number of existing individuals in the x age class; ax, number of surviving individuals in the x age class after smoothing; lx, standardized number of surviving individuals at the beginning of the x age class; dx, from x to x + 1 age standardized deaths during the interval; qx, mortality during the interval from x to x + 1 age; Lx, number of individuals still alive during the interval from x to x + 1; Tx, total number of individuals in the class from age x to over x age; ex, life expectancy or average life expectancy of individuals entering the x age class; Kx, disappearance rate.
下载: 导出CSV
表 4 红桦种群动态变化的时间序列分析
Table 4 Time sequence analysis of age structure of Betula albo-sinensis populations
龄级
Age class火地塘
Huoditang洮坪
Taoping个体数
Number of individualM2 M4 M6 M8 个体数
Number of individualM2 M4 M6 M8 Ⅰ 5 6 Ⅱ 18 8 37 17 Ⅲ 23 17 53 43 Ⅳ 21 22 15 26 36 26 Ⅴ 16 14 16 2 11 27 Ⅵ 5 6 14 12 2 2 19 18 Ⅶ 2 3 10 13 2 1 6 18 Ⅷ 2 2 4 10 10 2 1 2 13 14 Ⅸ 2 2 2 6 12 1 1 1 4 14 注:M2、M4、M6、M8分别表示经过2、4、6、8龄级时间后龄级的种群大小。Notes: M2, M4, M6, M8 indicate population sizes of age class after 2, 4, 6, 8 age class time, respectively.
下载: 导出CSV
-
[1] Wu X P, Zheng Y, Ma K P. Population distribution and dynamics of Quercus liaotungensis, Fraxinus rhynchophlla and Acer mono in Dongling Mountain[J]. Acta Botanica Sinica, 2002, 44(2): 212−222.
[2] Arista M. The structure and dynamics of an Abies pinsapo forest in southern Spain[J]. Forest Ecology and Management, 1995, 74(1−3): 81−89.
[3] 张文辉, 卢彦昌, 周建云. 巴山北坡不同干扰条件下栓皮栎种群结构与动态[J]. 林业科学, 2008, 44(7): 11−16. doi: 10.3321/j.issn:1001-7488.2008.07.003 Zhang W H, Lu Y C, Zhou J Y. Population structure and dynamics of Quercus variabilis in different habitats on northern slope of Bashan Mountain[J]. Scientia Silvae Sinicae, 2008, 44(7): 11−16. doi: 10.3321/j.issn:1001-7488.2008.07.003
[4] 李先琨. 濒危植物元宝山冷杉与南方红豆杉种群生态学研究[M]. 北京: 科学出版社, 2006. Li X K. Study on population ecology of endangered plant Abies yuanbao shanensis and Taxus chinensis var. mairei [M]. Beijing: Science Press, 2006.
[5] 李清河, 高婷婷, 刘建锋, 等. 荒漠珍稀灌木半日花种群的年龄结构与生命表分析[J]. 植物研究, 2009, 29(2): 176−181. doi: 10.7525/j.issn.1673-5102.2009.02.009 Li Q H, Gao T T, Liu J F, et al. The age structure and life table of rare eremophyte Helianthemum ordosicum population[J]. Plant Research, 2009, 29(2): 176−181. doi: 10.7525/j.issn.1673-5102.2009.02.009
[6] 张文辉, 祖元刚. 濒危种裂叶沙参种群生命表和存活曲线的研究及其与广布种泡沙参的对照[J]. 植物生态学报, 1999, 23(1): 76−86. doi: 10.3321/j.issn:1005-264X.1999.01.011 Zhang W H, Zu Y G. Study on population life and survivorship curves of Adenophora lobophylla, an endangered species, compared with A. potaninii, a widespread species[J]. Chinese Journal of Plant Ecology, 1999, 23(1): 76−86. doi: 10.3321/j.issn:1005-264X.1999.01.011
[7] 潘丽梅, 朱建华, 刘冰浩, 等. 广西龙虎山自然保护区龙荔种群生命表分析[J]. 园艺学报, 2011, 38(7): 1349−1355. Pan L M, Zhu J H, Liu B H, et al. Life table analysis of Dimocarpus confinis population in Guangxi Longhushan Nature Reserve[J]. Acta Horticulturae Sinica, 2011, 38(7): 1349−1355.
[8] 申仕康, 马海英, 王跃华, 等. 濒危植物猪血木自然种群结构及动态[J]. 生态学报, 2008, 28(5): 2404−2412. doi: 10.3321/j.issn:1000-0933.2008.05.058 Shen S K, Ma H Y, Wang Y H, et al. The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang[J]. Acta Ecologica Sinica, 2008, 28(5): 2404−2412. doi: 10.3321/j.issn:1000-0933.2008.05.058
[9] 解婷婷, 苏培玺, 周紫鹃, 等. 荒漠绿洲过渡带沙拐枣种群结构及动态特征[J]. 生态学报, 2014, 34(15): 4272−4279. Xie T T, Su P X, Zhou Z J, et al. Structure and dynamic characteristics of Calligonum mongolicum population in the desert-oasis ecotone[J]. Acta Ecologica Sinica, 2014, 34(15): 4272−4279.
[10] 姜景民. 中国桦木属植物地理分布的研究[J]. 林业科学研究, 1990, 3(1): 55−62. Jiang J M. The study of the geographical distribution of the Betula in China[J]. Forest Research, 1990, 3(1): 55−62.
[11] 任坚毅, 林玥, 岳明. 太白山红桦种子的萌发特性[J]. 植物生态学报, 2008, 32(4): 883−890. doi: 10.3773/j.issn.1005-264x.2008.04.017 Ren J Y, Lin Y, Yue M. Seed germination characteristics of Betula albo-sinensis at Mountain Taibai, China[J]. Chinese Journal of Plant Ecology, 2008, 32(4): 883−890. doi: 10.3773/j.issn.1005-264x.2008.04.017
[12] 刘志伟. 红桦汁保健饮料的研制[J]. 武汉工业学院学报, 2000(2): 4−6. Liu Z W. Preparation of red-brich juice health beverage[J]. Journal of Wuhan Polytechnic University, 2000(2): 4−6.
[13] 李家俊. 太白山自然保护区综合考察论文集[M]. 西安: 陕西师范大学出版社, 1989. Li J J. Collection of comprehensive investigation papers on Taibai Mountain Nature Reserve[M]. Xi’an: Shaanxi Normal University Press, 1989.
[14] 傅志军, 郭俊理. 太白山红桦林的初步研究[J]. 植物生态学报, 1994, 18(3): 261−270. doi: 10.3321/j.issn:1005-264X.1994.03.001 Fu Z J, Guo J L. Preliminary studies of the Betula albo-sinensis forest in Taibai Mountain[J]. Chinese Journal of Plant Ecology, 1994, 18(3): 261−270. doi: 10.3321/j.issn:1005-264X.1994.03.001
[15] 朱志诚. 秦岭太白山桦林的稳定性[J]. 武汉植物学研究, 1991, 9(2): 169−175. Zhu Z C. Stability of birch forest in Taibai Mountain of Qinling[J]. Plant Science Journal, 1991, 9(2): 169−175.
[16] 吴彦, 刘庆, 何海, 等. 光照与温度对云杉和红桦种子萌发的影响[J]. 应用生态学报, 2004, 15(12): 2229−2232. doi: 10.3321/j.issn:1001-9332.2004.12.007 Wu Y, Liu Q, He H, et al. Effects of light and temperature on seed germination of Picea asperata and Betula albo-sinensis[J]. Chinese Journal of Applied Ecology, 2004, 15(12): 2229−2232. doi: 10.3321/j.issn:1001-9332.2004.12.007
[17] 苏建文, 岳明, 王永军. 太白山红桦林林隙特征的研究[J]. 应用与环境生物学报, 2006, 12(2): 195−199. doi: 10.3321/j.issn:1006-687X.2006.02.011 Su J W, Yue M, Wang Y J. Gap characteristics of Betula albo-sinensis forest in the Taibai Mountains[J]. Chinese Journal of Applied and Environmental Biology, 2006, 12(2): 195−199. doi: 10.3321/j.issn:1006-687X.2006.02.011
[18] 谢晋阳. 森林植物立木种群静态生命表的一种编制方法[J]. 南昌大学学报, 1992, 16(4): 383−386. Xie J Y. A method of founding the life table of forest plants[J]. Journal of Nanchang University (Natural Science), 1992, 16(4): 383−386.
[19] 江洪. 云杉种群生态学[M]. 北京: 中国林业出版社, 1992. Jiang H. Spruce population ecology [M]. Beijing: China Forestry Publishing House, 1992.
[20] 姜在民, 和子森, 宿昊,等. 濒危植物羽叶丁香种群结构与动态特征[J]. 生态学报, 2018, 38(7): 2471−2480. Jiang Z M, He Z S, Su H, et al. Population structure and dynamic characteristics of the endangered plant Lilac pinnae[J]. Acta Ecologica Sinica, 2018, 38(7): 2471−2480.
[21] 陈晓德. 植物种群与群落结构动态量化分析方法研究[J]. 生态学报, 1998, 18(2): 214−217. Chen X D. A study on the method of quantitative analysis for plant population and community structural dynamics[J]. Acta Ecologica Sinica, 1998, 18(2): 214−217.
[22] 肖宜安, 何平, 李晓红, 等. 濒危植物长柄双花木自然种群数量动态[J]. 植物生态学报, 2004, 28(2): 252−257. doi: 10.3321/j.issn:1005-264X.2004.02.017 Xiao Y A, He P, Li X H, et al. Study on numeric dynamics of natural populations of the endangered species Disanthus cercidifolius var. longipes[J]. Chinese Journal of Plant Ecology, 2004, 28(2): 252−257. doi: 10.3321/j.issn:1005-264X.2004.02.017
[23] 何斌, 李青, 陈群利, 等. 贵州省西北部马尾松人工林种群数量特征与动态[J]. 中南林业科技大学学报, 2020, 40(44): 129−137, 155. He B, Li Q, Chen Q L, et al. Quantitative characteristics and population dynamics of Pinus massoniana plantation in northwest Guizhou Province[J]. Journal of Central South University of Forestry, 2020, 40(44): 129−137, 155.
[24] Taylor A H, Qin Z. Tree regeneration after bamboo die-back in Chinese Abies-Betula forests[J]. Journal of Vegetation Science, 1992, 3(2): 253−260. doi: 10.2307/3235687
[25] Taylor A H, Huang J Y, Zhou S Q. Canopy tree development and undergrowth bamboo dynamics in old-growth Abies-Betula forests in southwestern China[J]. Forest Ecology and Management, 2004, 25(10): 347−360.
[26] 吴语嫣, 李守中, 孙眭涛, 等. 长汀水土流失区侵蚀劣地马尾松种群动态[J]. 生态学报, 2019, 39(6): 2082−2089. Wu Y Y, Li S Z, Sun S T, et al. Population dynamics of Pinus massoniana on eroded land in the soil erosion area of Changting County[J]. Acta Ecologica Sinica, 2019, 39(6): 2082−2089.
-
期刊类型引用(9)
1. 包崇寅,孙永玉,李敏敏,邢洪铭,戚建华. 不同生境濒危植物龙棕种群结构及其动态特征. 西北植物学报. 2024(03): 479-490 .  百度学术
百度学术
2. 邱燕连,刘晨舒,梁晖,肖义锡,黄勤飞,傅书智. 黄楮林保护区福建青冈年龄结构及种群动态. 福建林业科技. 2024(02): 91-96 . 百度学术
3. 贾淇宇,刘雄盛,廖南燕,黄寒梅,黄荣林,肖玉菲,徐刚标,李娟. 极小种群野生植物狭叶坡垒种群结构与动态特征. 西南农业学报. 2024(05): 1078-1086 . 百度学术
4. 林碧华,陈绪辉,罗敏贤,陈泽平,张雨晨,刘宝,刘益鹏,郑世群. 珍稀濒危植物江南油杉群落区系与种群动态分析. 西北林学院学报. 2024(04): 71-78 . 百度学术
5. 陈思佚,唐燕,何腾,江永康,杜光源. 秦岭9个树种的木质部栓塞特性与水力安全风险. 植物生态学报. 2024(09): 1213-1222 . 百度学术
6. 吴万波,高文耀. 祁连山东段废弃渣台治理中乡土树种扦插培育及栽植试验分析. 农业与技术. 2023(10): 49-51 . 百度学术
7. 宋佳佳,李钢铁,郭靖捷,谷忠厚,刘坤,李治龙,康霞. 黄土丘陵区天然杜松种群结构及动态分析. 干旱区研究. 2023(08): 1304-1311 . 百度学术
8. 胡千惠,陈琳,赖媛,彭巧华,方平福,张盼盼,廖婷,高建新,潘永鑫,杨清培. 江西官山濒危植物穗花杉种群结构与动态. 江西农业大学学报. 2023(06): 1418-1430 . 百度学术
9. 张琴琴,胡雪平,雷曦,汪洋,刘汉才. 黄龙山金钱松种群结构与动态特征. 广西林业科学. 2022(06): 746-751 . 百度学术
其他类型引用(6)












计量
- 文章访问数: 757
- HTML全文浏览量: 276
- PDF下载量: 88
- 被引次数: 15
