
Identification and expression analysis during ovule development of Hsp90 gene family in Castanea mollissima
-
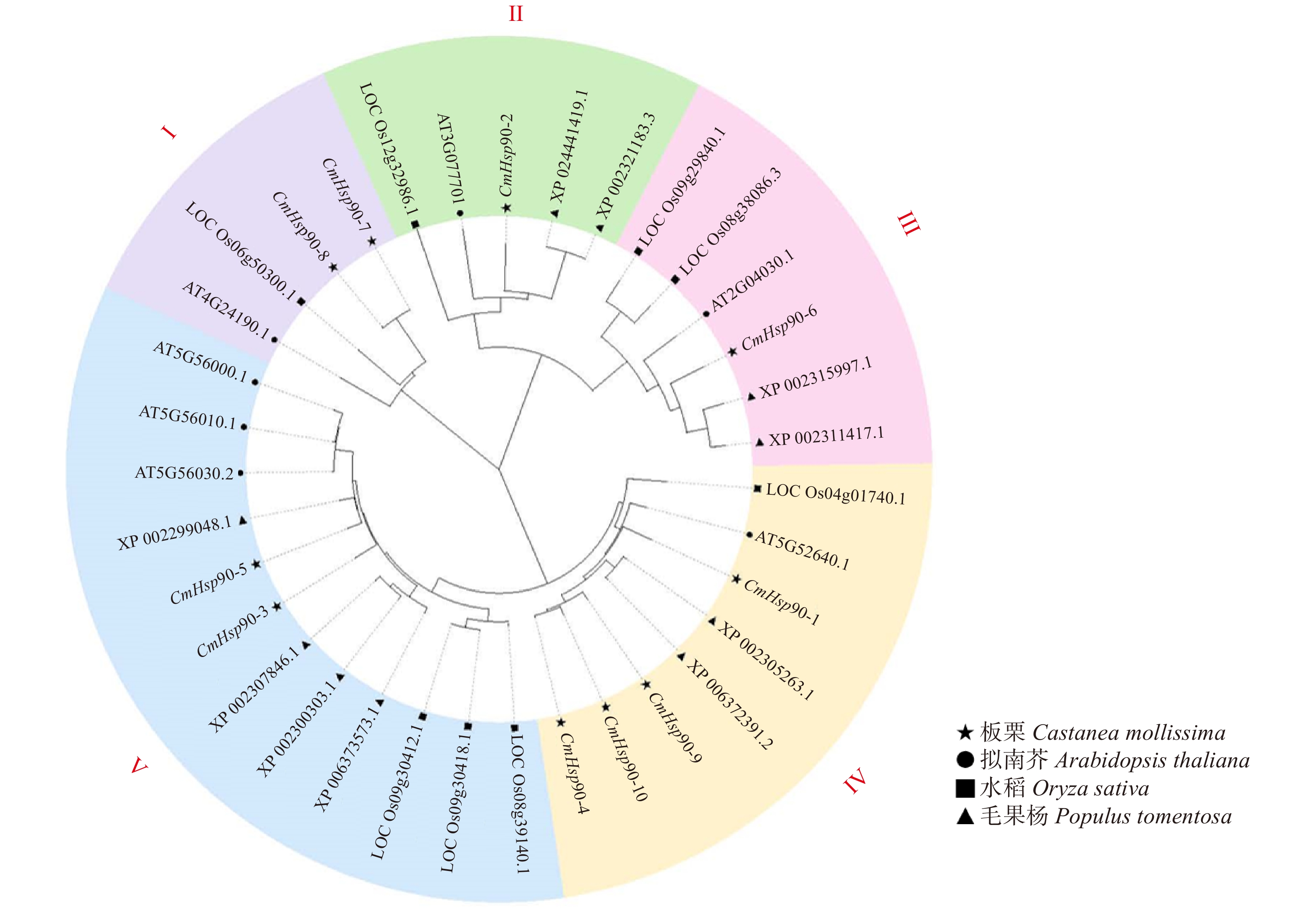
摘要:目的 热激蛋白(Hsp90)是一种在细菌、植物、动物中均广泛存在的高度保守的蛋白质分子,在植物应激反应和生长发育等过程中发挥着重要的作用。板栗作为我国重要的坚果树种,雌雄花比例不同、授粉受精困难及胚败育等现象致使板栗单产低,严重制约板栗的产量。近期有研究发现Hsp90基因家族参与植物的胚形成,本研究首次对板栗中Hsp90基因家族进行鉴定及表达分析,旨在揭示Hsp90基因家族的功能及其在授粉受精和胚珠发育过程中的作用,进而为提高板栗产量提供理论依据。方法 利用生物信息学方法对板栗Hsp90基因家族成员进行了鉴定,分析家族成员的理化性质、基因结构、进化关系、启动子元件。基于转录组数据分析CmHsp90基因在板栗胚珠不同发育时期的表达情况。结果 本研究在板栗中共鉴定到10个CmHsp90基因,该基因家族成员的编码区(CDS)长度范围是2 112 ~ 2 481 bp,含有4 ~ 22个外显子,编码703 ~ 826个氨基酸,相对分子量范围为80.67 ~ 94.21 kDa,等电点pI范围介于4.81 ~ 5.26;不稳定系数介于36.15 ~ 46.75之间,除CmHsp90-6、CmHsp90-4、CmHsp90-9外,其他基因编码的蛋白质相对比较稳定。对CmHsp90基因家族蛋白进行亚细胞定位预测发现:5个成员(CmHsp90-1、CmHsp90-4、CmHsp90-5、CmHsp90-9、CmHsp90-10)定位到了细胞质上,2个成员(CmHsp90-7、CmHsp90-8)定位到了内质网上,其余3个成员分别定位到细胞核(CmHsp90-3)、线粒体(CmHsp90-2)和叶绿体上(CmHsp90-6)上。对CmHsp90基因家族蛋白信号肽预测发现,仅CmHsp90-7和CmHsp90-8存在潜在的信号肽位点。CmHsp90基因家族蛋白的motif、保守结构域基本一致,均包含Hsp90结构域(PF00183)和HATPase_C结构域(PF02518)。基于系统进化树分析,10个CmHsp90家族蛋白被分为5个组。对CmHsp90基因家族启动子区域进行顺式作用元件预测,发现光响应元件是CmHsp90基因家族启动子上的主要调控元件,其次为ABA响应元件,此外,有4个基因(CmHsp90-1、CmHsp90-5、CmHsp90-7、CmHsp90-8)存在温度响应元件。通过对板栗胚珠不同发育阶段的RNA-seq数据进行分析发现:除CmHsp90-3基因外,其他基因在受精前胚珠中的表达量均高于受精后胚珠;CmHsp90-2、CmHsp90-4、CmHsp90-5、CmHsp90-7、CmHsp90-8、CmHsp90-9、CmHsp90-10基因在发育胚珠中的表达量高于败育胚珠。结论 CmHsp90基因在胚珠不同发育过程中的表达存在差异,在同一子房内发育胚珠和败育胚珠中的表达情况也不尽相同,研究结果为下一步研究CmHsp90基因在板栗授粉受精以及胚发育过程中的具体作用奠定理论基础。Abstract:Objective Recent studies have found that Hsp90 gene family is related to embryo development, but the details of Hsp90 gene family in chestnut are not clear. The identification and analysis of Hsp90 gene family in chestnut provide a theoretical basis for studying the function of Hsp90 gene family and its role in pollination, fertilization and ovule development, so as to improve the yield of chestnut.Method The members of chestnut Hsp90 gene family were identified by bioinformatics. The physical and chemical properties, gene structure, evolutionary relationship and promoter elements of the family members were analyzed. The expression of CmHsp90 genes at different developmental stages of ovule was analyzed based on transcriptome data.Result A total of 10 CmHsp90 genes were identified in Chinese chestnut. The length of CDS of 10 CmHsp90 gene family members ranged from 2 112 to 2 481 bp, the number of amino acids encoded ranged from 703 to 826. The relative molecular weight ranged from 80.67 to 94.21 kDa, and the pI ranged from 4.81 to 5.26 kDa. The coefficient of instability ranged from 36.15 to 46.75. Except CmHsp90-6, CmHsp90-4 and CmHsp90-9, the proteins encoded by other genes were relatively stable. According to the subcellular localization prediction of CmHsp90 family protein members, five members (CmHsp90-1, CmHsp90-4, CmHsp90-5, CmHsp90-9 and CmHsp90-10) were located in the cytoplasm, two members (CmHsp90-7 and CmHsp90-8) were located in the endoplasmic reticulum, and the other three members were located in the nucleus (CmHsp90-3), mitochondria (CmHsp90-2) and chloroplast (CmHsp90-6). By predicting the signal peptide of CmHsp90 gene family proteins, it was found that only CmHsp90-7 and CmHsp90-8 had potential signal peptide sites. The motif and protein domains of CmHsp90 gene family proteins were basically the same, including Hsp90 domain (PF00183) and HATPase_c domain (PF02518). Based on phylogenetic tree analysis, 10 CmHsp90 proteins were divided into 5 groups. The cis acting elements in the promoter region of CmHsp90 gene family were predicted. It was found that the light response element was the main regulatory element on the promoter of CmHsp90 gene family, followed by ABA response element. In addition, four genes (CmHsp90-1, CmHsp90-5, CmHsp90-7, CmHsp90-8) had temperature response elements. By analyzing the RNA-seq data of ovules at different developmental stages, it was found that the expression of other genes in ovules before fertilization was higher than that after fertilization, except CmHsp90-3 gene; the expression of CmHsp90-2, CmHsp90-4, CmHsp90-5, CmHsp90-7, CmHsp90-8, CmHsp90-9 and CmHsp90-10 genes in developing ovules was higher than that in abortive ovules.Conclusion The CmHsp90 genes in chestnut were identified and analyzed for the first time. The results show that the expression of Hsp90 genes is different in varied ovule development processes, and the expression in developing ovules and abortive ovules in the same ovary is also different. The research results lay a foundation for the further study of the specific role of CmHsp90 gene in chestnut pollination, fertilization and embryo abortion.
-
Keywords:
- Castanea mollissima /
- heat shock protein /
- gene family /
- embryo development /
- expression pattern
-
从古至今,木材作为一种天然可再生的环保型材料,以其独特的性能和优良的环境学特性深受人们喜爱,并广泛应用于生产生活的各个方面。随着现代科学技术的突飞猛进、人类学理论的不断发展,考古木材作为进行植物考古学和环境考古学研究的一种典型生物材料,正成为专家学者的研究热点,国内外考古作业挖掘出大量的木质文物[1-2],例如:“Riksapplet号”沉船[3]、“小白礁Ⅰ号”沉船[4]、“南海Ⅰ号”船木[5]等为出土考古木材的研究提供了丰富的材料。
Crestini等[6]运用X射线衍射和扫描电镜的方法对埃及古代棺木进行研究,研究发现:考古木材中纤维素、半纤维素、木质素均产生不同程度的降解,小分子化合物发生改变,使考古木材变得脆弱而柔软。Łucejko等[7]运用热裂解气相色谱–质谱联用法对取自比萨圣罗索雷古船的木材样品进行了主成分分析,样品木质素中愈创木基和紫丁香基的去甲基化表明考古木材中多糖产生了流失。卢芸[8]指出大多木质文物纤维素结晶区被严重破坏,葡聚糖侧链的乙酰基消失,多糖类物质遭到严重降解,木质素相对含量明显增加。
考古木材微观结构的变化,将在很大程度上影响其水分吸着行为,因而可利用等温吸附曲线和水分吸着理论对考古木材与吸着水分子的相互作用进行分析,如Guggenheim-Anderson-de Boer模型[9]、Hailwood-Horrobin模型[10]等。Guo等[11]利用Guggenheim-Anderson-de Boer模型计算得到每千克古代润楠木(Machilus pingii Cheng ex Yang)样品的最大单层吸水量为7.388 kg,而相同树种现代木材的最大单层吸水量仅为4.602 kg,考古木材的最大单层吸水量比现代木材增加了61%。
木材的水分吸着过程必然伴随着热量或能量的变化[12],因此基于吸附热力学考察水分吸着机理,有助于从本质上获得考古木材与水分相互作用方面的信息。曹金珍[13]发现西藏云杉(Picea spinulosa)在水分吸着过程中微分吸着热、微分吸着熵值随吸湿过程不断增大。Simón 等[14]研究了在吸湿和解吸平衡态下辐射松(Pinus radiata)的微分吸着熵值随含水率的变化曲线,结果显示在相同平衡含水率下辐射松解吸的微分吸着熵值大于吸湿值。
在以往的研究中,研究者多从成分、结构角度对考古木材水分吸着行为进行研究,且取得了一定进展,而对考古木材的吸附热力学研究较少,而且对各种热力学量变化的系统分析也很欠缺。本研究将聚焦考古木材在吸湿和解吸平衡状态下微分吸着热、自由能变化及微分吸着熵三大热力学量的研究,并分别利用电子显微镜(scanning electron microscope,SEM)、傅里叶红外光谱(Fourier transform infrared spectroscopy,FTIR)分析考古木材的细胞壁结构和化学成分变化,从而建立考古木材微观结构与其水分吸着行为间的构效关系,探究考古木材与现代木材在水分吸附热力学方面的差异及其原因,为从能量角度解析考古木材与水分之间的相互作用机制奠定理论基础,同时也可为我国木质文物的保护提供科学依据,以减少出土饱水木质文物与环境中水分的相互作用,提高木质文物的尺寸稳定性。
1. 材料与方法
1.1 材 料
本研究所用考古木材取自成都文物考古研究院发掘出土的南宋木棺残片,由研究院提供的材料和信息确定为考古柏木,并采集四川产柏木(Cupressus sp.)作为现代对照材。
1.2 细胞壁微观形貌表征
利用滑动切片机(REM-710,Yamato Kohki industrial Co.,Ltd,Japan)制得尺寸为1 mm(轴向) × 2 mm(弦向) × 2 mm(径向)的柏木考古木材与现代木材薄片试样(试样重复数为3),用于SEM观察。在80 ℃的鼓风干燥箱中将试样干燥至质量恒定后进行喷金处理,设置SEM(S-3400,Hitachi,日本)加速电压为3 kV,对柏木试样的细胞结构进行观察,初步了解考古木材的降解、腐朽情况。
1.3 化学成分表征
利用研磨机将考古木材与现代木材样品加工成100目以下的木粉,并在80 ℃的真空干燥箱中干燥至质量恒定,之后将木粉与溴化钾充分混合研磨压制成透明薄片,利用红外光谱仪(BRUKER vertex 70 V,德国)对试样进行检测分析,扫描范围设为400 ~ 4 000 cm−1,分辨率为4 cm−1,每个试样扫描次数为32次。重复测试3次,取平均值。
1.4 吸附热力学量测定
将柏木考古木材、现代木材样品加工为20 mg左右木片,并分别置于(103 ± 2)℃下干燥至质量恒定,用于动态水分吸附分析(dynamic vapor sorption,DVS)实验。将各组试样放入DVS中,分别预设恒定温度为25和50 ℃,相对湿度以10%为梯度从0%增加到90%,继续增加至95%,然后降回至90%,再以10%为梯度从90%降低到0%,试样在每个相对湿度下逐渐达到吸湿/解吸平衡后(当样品质量变化速率连续在10 min内小于0.000 1%/min,则认为样品达到吸湿/解吸平衡),仪器自动进入下一个相对湿度梯度。仪器每隔1 min记录样品质量、温度和相对湿度等数据,得到考古木材和现代木材的吸湿和解吸平衡的等温吸附曲线。
利用Hailwood-Horrobin水分吸着理论[10]分析所获得的等温吸附曲线,木材在不同相对湿度条件下达到的平衡含水率可表达为
M=Mh+Md=18m(KhKdH1+KhKdH+KdH1−KdH) (1) 式中:M为试样平衡含水率(%);Mh为水合水质量分数(%);Md为溶解水质量分数(%);m为单位摩尔数吸着位点的绝干木材质量(g/mol);Kh为水合水与溶解水之间的平衡常数;Kd为溶解水与环境温湿度之间的平衡常数;H为相对湿度(%)。
将式(1)整理成多项式,具体为
HM=A+BH−CH2 (2) 式中:A、B、C为多项式的拟合参数,A、B、C与Kh、Kd、m的关系为
A=m18×1Kd(Kh+1) (3) B=m18×Kh−1Kh+1 (4) C=m18×KhKdKh+1 (5) 通过不同相对湿度环境试样的等温吸湿曲线数据分析可得到拟合参数A、B、C的值,进而可以求解m、Kh和Kd的值,公式中
m18 的倒数18m 表示木材中有效吸着基团被水合水饱和时的含水率,间接表达了水分吸着位点的数量。利用各组试样的DVS数据拟合等温吸附曲线,进一步通过Clausius-Clapeyron公式[15]分别计算考古木材与现代木材的微分吸着热QS、自由能变化ΔG和微分吸着熵ΔS,分析考古木材与现代木材在吸附热力学量间的差异,各热力学量的计算公式如下。
QS=0.254d(ln1H)/d(ln1H)d(1T)d(1T) (6) ΔG=−RT18lnH (7) ΔS=QS−ΔG/T (8) 式中:T为温度(K);R为气体常数。
T和H的值均由DVS实验的拟合数据获取,然后分别通过公式(6)、(7)计算得到QS和ΔG的值,进一步计算得到ΔS。
2. 结果与讨论
2.1 细胞壁结构
图1为考古木材与现代木材横切面、纵切面的SEM图片。对比考古木材与现代木材图像,可以发现在经历长时间的饱水腐朽下,考古木材除复合胞间层、细胞角隅保存相对完整外[16-17],其细胞壁结构发生了明显的变化,与现代木材完整有序的细胞排列相比(图1b),考古木材由于长时间遭受微生物降解,细胞结构发生破坏,细胞壁产生腐朽。
具体而言,考古木材细胞壁上出现少量圆形孔洞(图1a),除此之外,从考古木材纵切面可见纹孔膜腐朽细节(图1c),并有菌丝体的存在(图1d),该菌丝体是导致考古木材在所处环境中受到降解的各类真菌的聚集体。Guo等[11]通过氮气吸附法和压汞法对山桐子(Idesia polycarpa)考古木材多孔结构的研究结果表明,考古木材的中孔隙的总孔体积比现代木材样品增加了471%,这也进一步证明了本研究中考古木材孔洞增多的事实。
2.2 化学成分变化
对等量考古木材与现代木材样品的红外光谱进行归一化处理后,可用吸收峰高度半定量表示吸收峰所代表基团的含量[18],通过对比考古木材与现代木材试样红外光谱(图2)的特征吸收峰位置变化及消失情况,结合各特征吸收峰所代表的化学基团,研究得到考古木材3种主要化学成分(纤维素、半纤维素、木质素)的含量变化情况及极性基团−OH、−COOH类型和含量的变化情况。
3 337 ~ 3 401 cm−1范围内的吸收峰代表极性基团−OH(O−H伸缩振动),考古木材在此处的吸收峰高度小于现代木材,证明考古木材中−OH含量减少。选取1 742 cm−1(半纤维素C=O伸缩振动)、1 231 cm−1(半纤维素酰氧键−COO伸缩振动)为半纤维素的代表特征峰,与现代木材相比,柏木考古木材的红外吸收光谱在1 742、1 231 cm−1处吸光度变小,特别是在1 742 cm−1处吸收峰几乎完全消失,证明考古木材细胞壁结构中的半纤维素降解严重;另一方面,1 231 cm−1吸收峰代表半纤维素酰氧键−COO,考古木材在1 231 cm−1处的吸收峰高度明显小于现代木材,证明考古木材半纤维素上的−COOH含量减少。
此外,由于考古木材纤维素结晶区中纤维素链的氢键网络的部分断裂,导致892 cm−1处的峰几乎消失,而该处波峰代表C−H弯曲振动,并且对氢键系统的性质敏感,因此该峰几乎消失表明,在考古木材试样中形成更稳定氢键的羟基数量减少,同时这也表明考古木材中无论是无定形区还是结晶区中的纤维素链都发生部分降解[19]。
指纹区吸收光谱峰数多、较为复杂,包含了更多能反映分子结构细微变化的信息[20]。为了提供更多的信息,将FTIR光谱图分析细化至3个指纹区域(1 550 ~ 1 800 cm−1、1 200 ~ 1 550 cm−1和850 ~ 1 200 cm−1),在这个范围内通过直接的红外光谱分析考古木材和现代木材仍存在一定局限性,因为指纹区往往存在高度重叠的峰,难以分辨和解析。为此,通过对考古木材和现代木材指纹区光谱进行分峰拟合处理,得到分峰拟合曲线,可以更直观地分析考古木材化学成分的变化。
通过对比考古木材和现代木材FTIR在1 550 ~ 1 800 cm−1波数范围内的分峰拟合曲线(图3a)可以发现,考古木材在1 654 cm−1(木质素中共轭C=O伸缩振动)处吸收峰吸光度值较高,这说明考古木材木质素相对含量升高。通过对比考古木材和现代木材FTIR在1 200 ~ 1 550 cm−1波数范围内的分峰拟合曲线(图3b)可以发现,考古木材在1 418 cm−1(木质素甲基中C−H弯曲振动)处吸收峰强度变大,代表考古木材木质素相对含量升高。通过对比分析考古木材和现代木材FTIR在850 ~ 1 200 cm–1波数范围内的分峰拟合曲线(图3c)可以得到:考古木材在892 cm–1(纤维素C−H弯曲振动)处峰几乎消失,这表明在考古木材中形成更稳定氢键的羟基数目减少;1 159 cm–1(综纤维素C−O−C变形振动)处,考古木材吸收峰吸光度值很低,这表明综纤维素也有一定程度的降解。
![]() 图 3 考古木材与现代木材在1 550 ~ 1 800 cm–1(a)、1 200 ~ 1 550 cm–1(b)、850 ~ 1 200 cm–1(c)波数范围内的分峰拟合曲线Figure 3. Deconvolution of the “fingerprint” region for archaeological wood and recent wood FTIR spectrum 1 550 − 1 800 cm–1 (a), 1 200 − 1 550 cm–1(b), 850 −1 200 cm–1(c)
图 3 考古木材与现代木材在1 550 ~ 1 800 cm–1(a)、1 200 ~ 1 550 cm–1(b)、850 ~ 1 200 cm–1(c)波数范围内的分峰拟合曲线Figure 3. Deconvolution of the “fingerprint” region for archaeological wood and recent wood FTIR spectrum 1 550 − 1 800 cm–1 (a), 1 200 − 1 550 cm–1(b), 850 −1 200 cm–1(c)综上可以发现,考古木材与现代木材相比,半纤维素、纤维素降解严重,纤维素骨架结构受到破坏,因此在SEM下可观察到细胞壁产生腐朽,这种化学成分和化学结构的变化进一步导致了细胞壁孔洞的出现,从化学成分变化角度解释了SEM中观察到考古木材出现孔洞等腐朽现象的原因。通过Xia等[21]的研究,考古木材木质素的化学结构也发生了变化,木质素仍会有降解,但考古木材中多糖类物质(纤维素、半纤维素)比芳环类物质(木质素)降解程度更高,因而木质素相对含量总体有升高趋势[22-23]。除纤维素、半纤维素含量降低及木质素相对含量升高的变化外,考古木材中极性基团−OH、−COOH含量均减少,导致考古木材对水分的吸着减少。
2.3 吸附热力学分析
2.3.1 Hailwood-Horrobin模型拟合
通过Hailwood-Horrobin模型拟合等温吸附曲线,不仅可以得到给定温度下木材吸湿和解吸任意平衡含水率状态下的相对湿度,更可以通过研究考古木材与现代木材的曲线差异,帮助理解考古木材水分吸附行为的差异本质。
根据Hailwood-Horrobin模型分别拟合出在25和50 ℃下考古木材、现代木材的等温吸附曲线(图4)。从中可以看出各等温吸附曲线的拟合度R2值均高于0.99,即Hailwood-Horrobin模型能够较好地描述考古木材及现代木材的等温吸附曲线。所有的等温吸附曲线均呈现“S”形,这说明无论考古木材还是现代木材都属于Ⅱ类等温吸附曲线,具备多孔材料的吸附特征[24-25]。在给定温度的每一个相对湿度条件下,考古木材的平衡含水率均高于现代木材,以温度为50 ℃,相对湿度为0.6时为例,考古木材平衡含水率为7.94%,现代木材为7.46%,考古木材是现代木材的1.1倍。这是因为图1a的 SEM图像显示考古木材物理环境遭到改变,细胞壁孔洞增多,即木材降解程度越大,木材细胞壁内孔隙越多,对水分的容纳空间增加,含水率越高,所以考古木材在经历长时间的降解后平衡含水率高于现代木材。
![]() 图 4 考古木材与现代木材的等温吸附曲线Figure 4. Adsorption isotherms of archaeological wood and recent wood
图 4 考古木材与现代木材的等温吸附曲线Figure 4. Adsorption isotherms of archaeological wood and recent wood图5为50 ℃下考古木材吸湿和解吸平衡状态的等温吸附曲线,图5显示考古木材的解吸曲线始终位于吸湿之上,即出现了水分吸湿滞后现象。表1总结了在不同相对湿度条件下,考古木材、现代木材分别在25、50 ℃下的滞后率。从表1可以看出:随着温度或相对湿度的升高,考古木材和现代木材的滞后率均增大,即水分吸湿滞后均减小,这与以往研究结果一致[13];另一方面,考古木材与现代木材的水分吸湿滞后大小差异不大。
![]() 图 5 50 ℃下考古木材的吸湿和解吸等温吸附曲线Figure 5. Adsorption and desorption isotherms of archaeological wood at 50 ℃
图 5 50 ℃下考古木材的吸湿和解吸等温吸附曲线Figure 5. Adsorption and desorption isotherms of archaeological wood at 50 ℃2.3.2 吸湿平衡状态下的热力学量分析
2.3.2.1 微分吸着热
图6是吸湿平衡状态下考古木材与现代木材中水分的微分吸着热(QS)随含水率(moisture content,MC)的变化曲线,QS值越大表示木材中水分子与周围分子之间的结合力越强[13]。与以往研究结果略有不同的是,在MC为5%处产生一明显拐点。根据曲线变化趋势,可以将曲线分为两个阶段,当MC小于 5%的低含水率状态下时,QS值逐渐增大直至达到最大值,这表明水分子和木材之间的氢键结合首先表现为单层分子之间的相互结合,且随MC增大,单个水分子和木材中的−OH形成了更多的氢键结合,相互作用更强,形成的键能更高[18]。当含水率大于5%时,QS值随MC的增大而减小,此时单分子层吸着水几乎达到饱和,吸着水分子与木材实质之间形成的氢键结合作用变弱,键能降低[13]。
表 1 考古木材、现代木材在25、50 ℃下不同相对湿度下的吸湿滞后Table 1. Moisture sorption hysteresis of archaeological wood and recent wood under different relative humidities at 25 and 50 ℃相对湿度
Relative humidity滞后率 Hysteresis rate 25 ℃考古木材
25 ℃-archaeological wood50 ℃考古木材
50 ℃-archaeological wood25 ℃现代木材
25 ℃-recent wood50 ℃现代木材
50 ℃-recent wood0.1 0.62 0.74 0.63 0.71 0.2 0.63 0.72 0.64 0.73 0.3 0.64 0.72 0.65 0.74 0.4 0.66 0.73 0.67 0.76 0.5 0.68 0.75 0.70 0.78 0.6 0.71 0.77 0.73 0.81 0.7 0.76 0.81 0.77 0.84 0.8 0.82 0.86 0.84 0.89 0.9 0.91 0.94 0.93 0.95 ![]() 图 6 考古木材与现代木材中水分的微分吸着热(QS)随含水率(MC)的变化曲线Figure 6. Variation curves of differential heat of sorption (QS) of moisture in archaeological wood and recent wood against moisture content (MC)
图 6 考古木材与现代木材中水分的微分吸着热(QS)随含水率(MC)的变化曲线Figure 6. Variation curves of differential heat of sorption (QS) of moisture in archaeological wood and recent wood against moisture content (MC)考古木材与现代木材相比,整体曲线形状基本相同,但考古木材的QS值随MC的变化曲线位于现代木材之下。根据上文FTIR分析,考古木材化学成分中纤维素、半纤维素大量降解,因此外界水分子可结合−OH、−COOH位点数量减少,这种化学环境的改变导致吸着水与木材分子之间的结合减少,QS值降低[26]。
2.3.2.2 自由能变化
水分的自由能变化的物理意义是水分子润胀木材并切断木材分子相互间氢键结合,裸露木材吸着点所需做的功[13]。基于图7所示吸湿平衡状态下考古木材与现代木材中水分的自由能变化ΔG值随MC的变化曲线,可以发现考古木材与现代木材的ΔG值均随MC的增大而减小。ΔG值与MC变化呈负相关可以解释为,MC升高使木材可塑性增大,吸着水分子与木材实质分子的协同运动加剧,因此裸露木材吸着点所需做的功减少。
![]() 图 7 考古木材(a)与现代木材(b)中水分的自由能变化(ΔG)随含水率的变化曲线Figure 7. Variation curves of free energy change (ΔG) of moisture in archaeological wood (a) and recent wood (b) against moisture content
图 7 考古木材(a)与现代木材(b)中水分的自由能变化(ΔG)随含水率的变化曲线Figure 7. Variation curves of free energy change (ΔG) of moisture in archaeological wood (a) and recent wood (b) against moisture content其次,无论是考古木材或是现代木材,50 ℃相较于25 ℃的ΔG值都有一定程度的减小,这是因为温度升高吸着水分子能量增加,活动加剧,所需做的功减小。
2.3.2.3 微分吸着熵
微分吸着熵(ΔS)体现了木材中吸着的水分子与液态水分子相比排列规则上的差异,ΔS > 0代表木材中水分子排列更有规律[13]。根据吸湿平衡状态下考古木材与现代木材中水分的ΔS值随MC的变化曲线(图8)可知:ΔS值与QS值随MC的变化曲线在MC小于5%呈现相似的变化趋势,都随MC的增大而增大。在MC小于5%的低含水率阶段,ΔS < 0表明木材中吸着的水分子比液态水分子更无序,这是因为此时木材中吸着的水分子主要为单分子层吸着水,它们比处于液体状态的水分子显示出更无序的状态;在经过5%的拐点之后,ΔS值随MC变化不明显。而随着MC的增大,ΔS值经历了由负到正、由无序到有序的变化,这说明吸着在木材中的水分子排列越来越规则。
![]() 图 8 考古木材与现代木材中水分的微分吸着熵(ΔS)随含水率的变化曲线Figure 8. Variation curves of differential entropy of sorption (ΔS) of moisture in archaeological wood and recent wood againstmoisture content
图 8 考古木材与现代木材中水分的微分吸着熵(ΔS)随含水率的变化曲线Figure 8. Variation curves of differential entropy of sorption (ΔS) of moisture in archaeological wood and recent wood againstmoisture content进一步分析考古木材与现代木材的曲线差异,考古木材ΔS值低于现代木材,这是因为考古木材在经历长期酸碱环境下软腐菌等真菌的降解后[27],细胞壁出现的孔洞分布是随机的,且与水结合的−OH分布也十分不均匀,造成考古木材中吸着的水分子排列更无序。
2.3.3 水分吸湿与解吸的比较
以50 ℃考古木材为例,分别绘制QS、ΔG、ΔS三大热力学量随MC的变化曲线,每张图包含吸湿和解吸平衡态下的两条曲线(图9)。发现吸湿和解吸变化曲线形状基本一致,但解吸的QS、ΔG和ΔS值均大于吸湿的值[14],对应于水分吸湿滞后(图5),这个现象即为“热力学吸湿滞后”,其表征在吸湿与解吸平衡态下,每一个吸着水分子与木材吸着点之间的氢键结合平均数的差异[13]。
![]() 图 9 50 ℃下考古木材吸湿与解吸过程中的QS(a)、ΔG(b)、ΔS(c)随含水率的变化曲线Figure 9. Variation curves of QS (a), Δ G (b), Δ S (c) in archaeological wood against moisture content during moisture adsorption and desorption at 50 ℃
图 9 50 ℃下考古木材吸湿与解吸过程中的QS(a)、ΔG(b)、ΔS(c)随含水率的变化曲线Figure 9. Variation curves of QS (a), Δ G (b), Δ S (c) in archaeological wood against moisture content during moisture adsorption and desorption at 50 ℃热力学吸湿滞后同样可以用水分吸湿滞后的“有效羟基说”[28]解释,即在干燥状态下,部分木材吸着点之间形成氢键结合,当木材从干燥状态开始吸湿时,由于这部分已经形成的氢键结合的影响,处于游离状态能够吸着水分子的吸着点的数量减少,每个吸着水分子与木材吸着点之间的氢键结合平均数减少,从而吸湿的热力学量也低于解吸过程。
3. 结 论
为探究考古木材的水分吸着行为与现代木材的差异机理,本研究从考古木材与现代木材微观结构与吸附热力学两个层面进行了研究。
考古木材三大主要成分(纤维素、半纤维素、木质素)均发生不同程度的降解,且考古木材中极性基团−OH、−COOH含量均减少,导致考古木材对水分的吸着减少。其中半纤维素、纤维素降解程度更大,因此在SEM中可观察到细胞壁产生腐朽,且可见孔洞分布;而考古木材中多糖类物质(纤维素、半纤维素)比芳环类物质(木质素)降解程度更高,因而木质素相对含量升高。
在吸附热力学研究中,通过DVS等温吸附实验并利用Hailwood-Horrobin模型拟合等温吸附曲线,在给定温度条件下考古木材的平衡含水率高于现代木材。结合Clausius-Clapeyron公式计算考古木材与现代木材的QS、ΔG和ΔS三大热力学量。考古木材因纤维素、半纤维素降解导致−OH、−COOH数量减小,QS值低于现代木材;考古木材与现代木材的ΔG值差异不大,且随温度升高而减小;考古木材的细胞壁孔洞、−OH分布不均匀,所以ΔS值低于现代木材;在MC小于5%,考古木材QS值和ΔS值随MC的变化曲线趋势相似,在此范围内木材中吸着的水分子主要为单分子层吸着水,随后在5%处产生一拐点,此时单分子层吸着水达到饱和,此后木材中吸着的水分子以多分子层吸着水占主导。最后,通过对比吸湿和解吸过程的热力学量(吸湿 < 解吸),发现考古木材存在热力学吸湿滞后现象。
基于以上对于考古木材水分吸着行为差异的研究,考古木材的吸湿性在经历长时间的泡水、腐朽下发生了较大变化,因此在出土饱水木质文物的保护工作中,建议采用各种物理、化学手段减少木质文物与环境中水分的相互作用,如涂覆防水涂层、浸渍脱水材料等,从而提高木质文物的尺寸稳定性[29],为我国出土饱水木质文物的保护提供理论支撑。
-
![]()
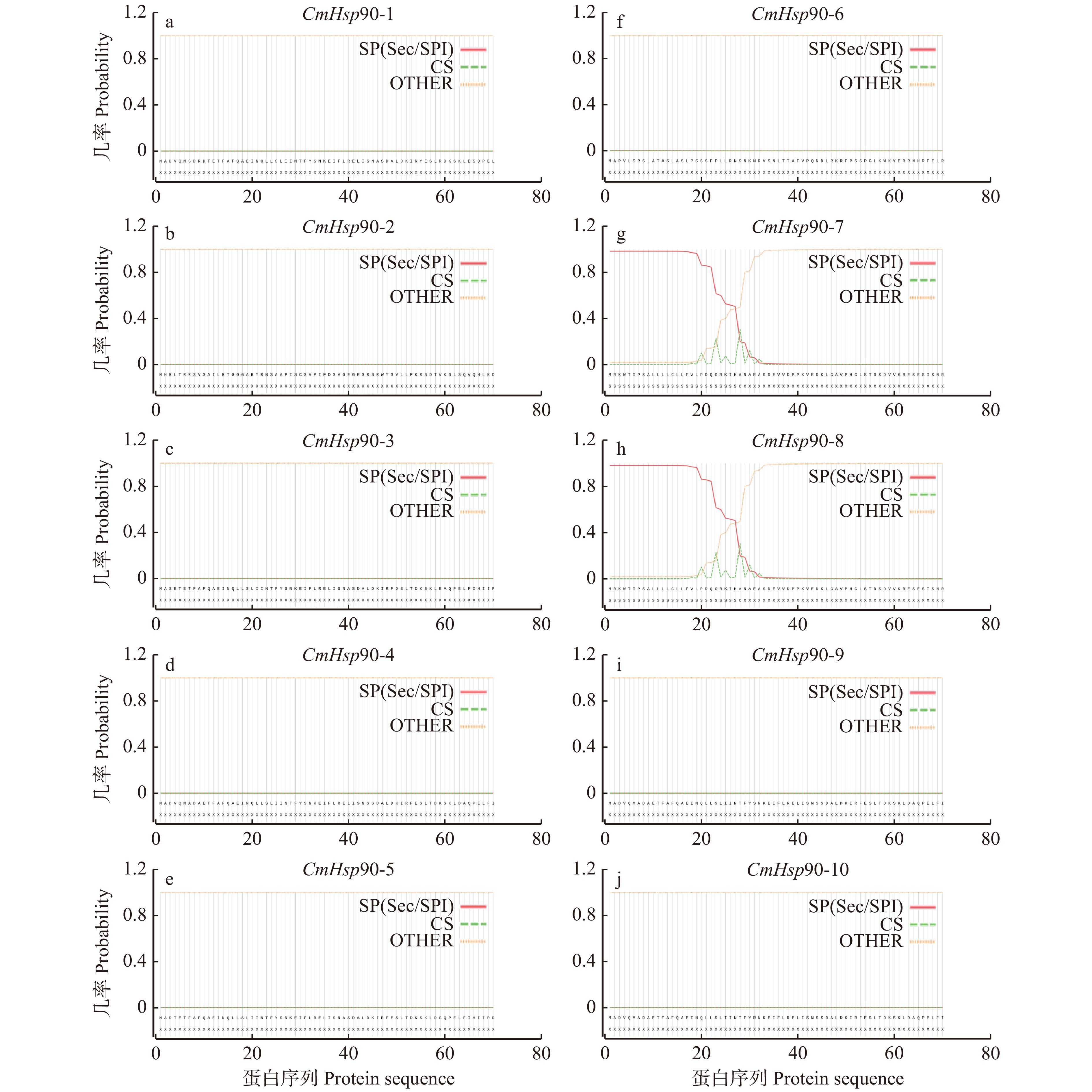
图 1 CmHsp90家族蛋白序列的信号肽位点预测
SP(Sec/SPI). 由Sec转座子转运并被信号肽酶I裂解的“标准”分泌信号肽;CS. 裂解位点;OTHER. 序列没有任何信号肽。SP(Sec/SPI), ‘standard’ secretory signal peptides transported by the Sec translocon and cleaved by signal peptidase I; CS, the cleavage site; OTHER, no signal peptide at all.
Figure 1. Prediction of signal peptide sites in CmHsp90 family protein sequence
![]()
图 2 CmHsp90家族氨基酸多序列比对
Figure 2. Amino acid multiple sequence alignment of CmHsp90 gene family
![]()
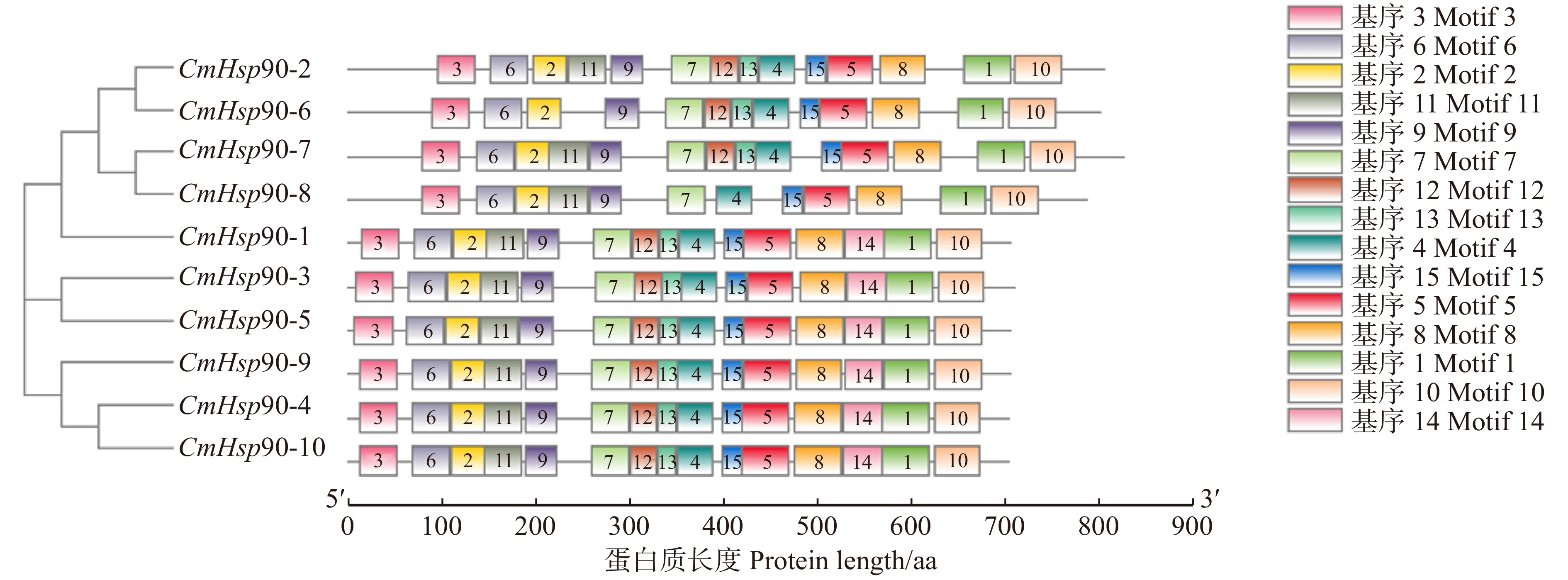
图 3 CmHsp90家族蛋白基序预测分析
Figure 3. Motif prediction and analysis of CmHsp90 family protein
![]()
图 4 板栗CmHsp90蛋白功能域预测
Figure 4. Functional domain prediction of CmHsp90 protein in C. mollissima
![]()
图 5 板栗CmHsp90家族基因启动子顺式作用元件预测分析
Figure 5. Functions of promoter cis-acting elements in CmHsp90 family genes in C. mollissima
![]()
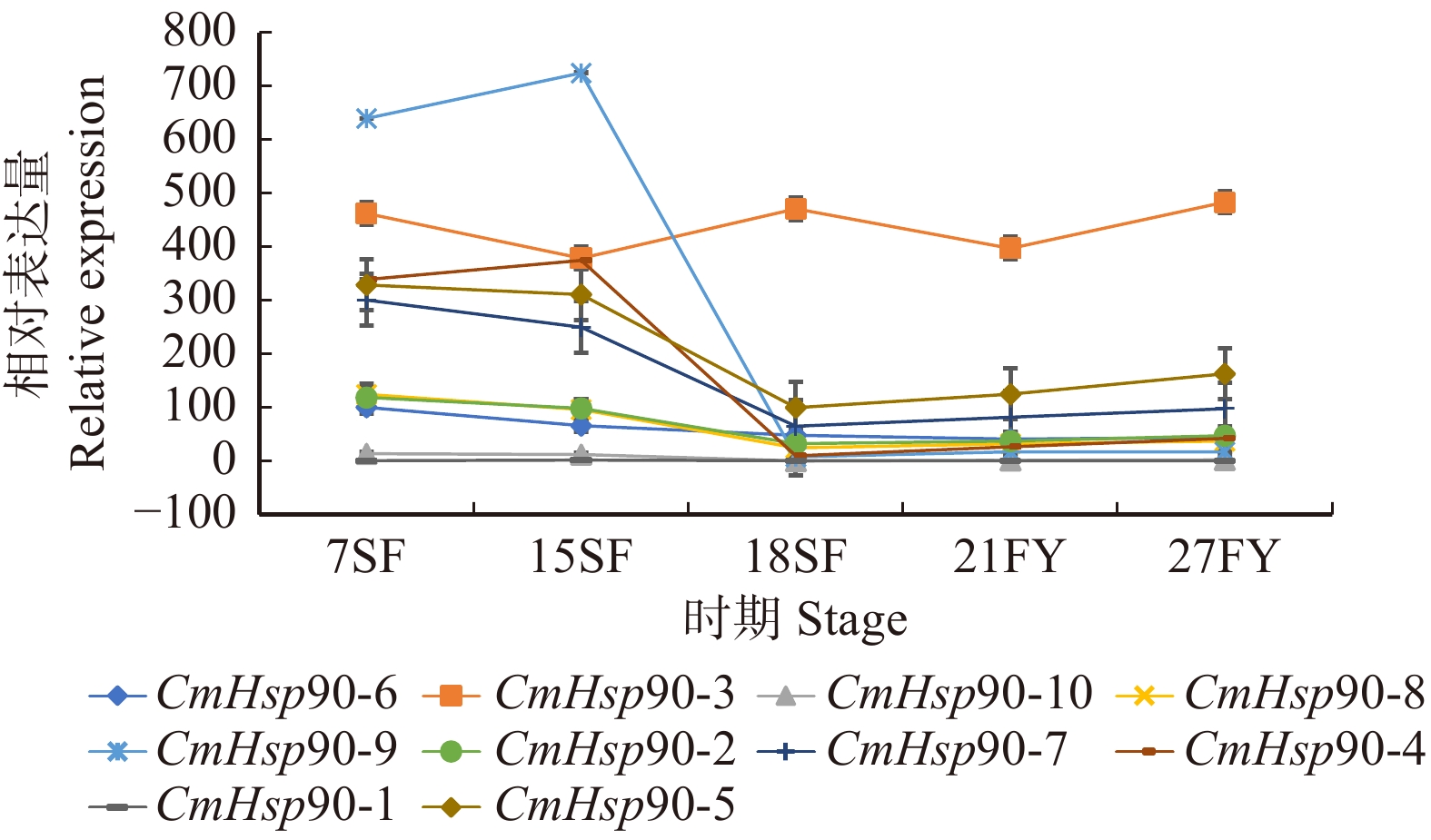
图 7 CmHsp90基因在胚珠不同发育时期的表达分析
7SF. 授粉后7d;15SF. 授粉后15 d;18SF. 授粉后18 d;21FY. 授粉后27 d发育胚珠;27FY. 授粉后27 d发育胚珠。下同。7SF, 7 days after pollination; 15SF, 7 days after pollination; 18SF, 18 days after pollination; 21FY, ovule developed 27 days after pollination; 27FY, ovule developed 27 days after pollination. The same below.
Figure 7. Expression analysis of CmHsp90 gene family in different development stages of ovule
![]()
图 8 CmHsp90基因在授粉后18 d和未授粉胚珠中的表达情况
18WF. 未授粉,与18 d授粉胚珠(18SF)同时采集。18WF, unpollinated, collected at the same time with 18 day pollinated ovules (18SF).
Figure 8. Expression of CmHsp90 gene in 18 days after pollination and unpollinated ovules
![]()
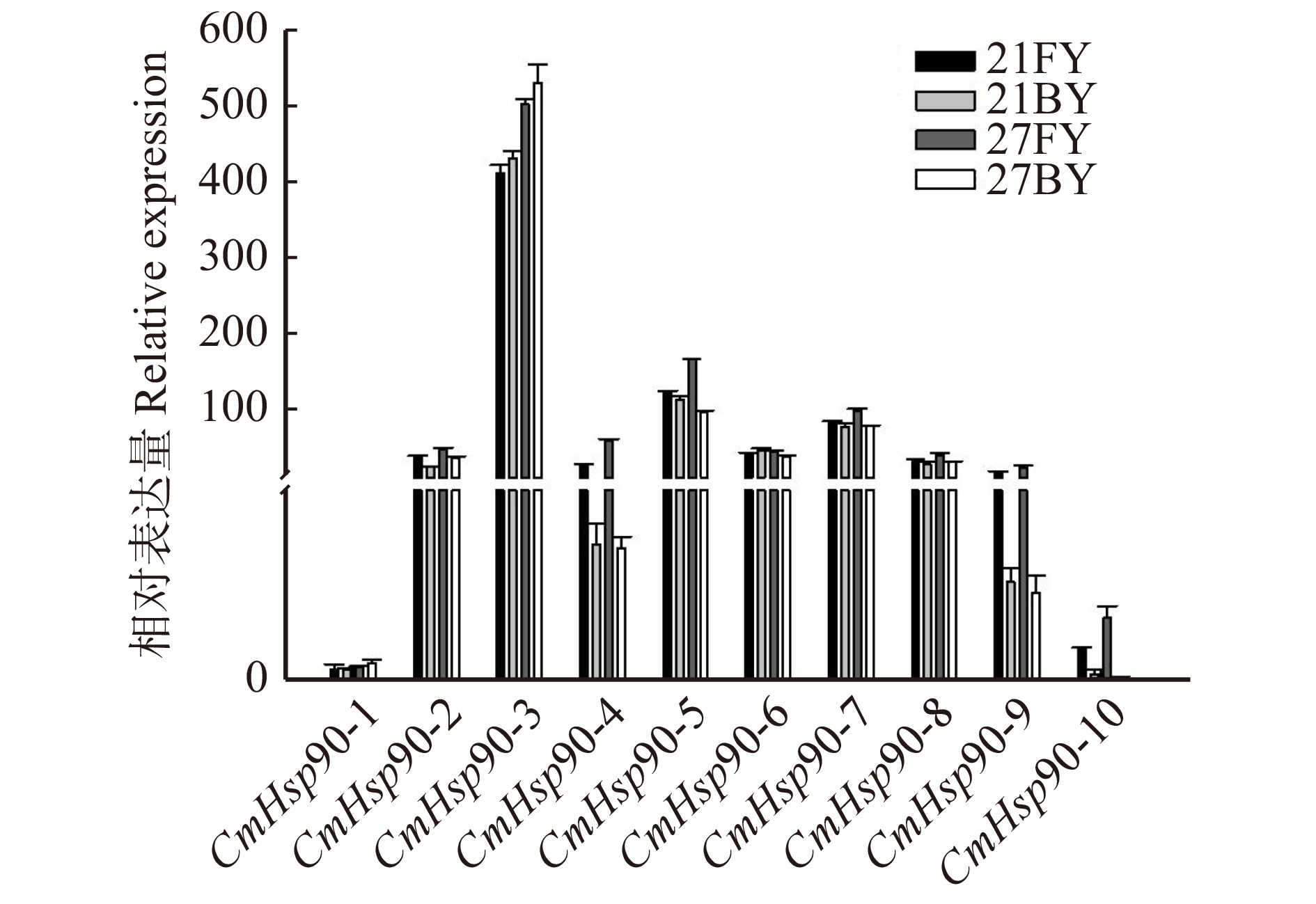
图 9 CmHsp90基因在发育胚珠与败育胚珠中的表达情况
21BY. 授粉后21 d败育胚珠;27BY. 授粉后27 d败育胚珠。21BY, ovule aborted 21 days after pollination; 27BY, ovule aborted 27 days after pollination.
Figure 9. Expression of CmHsp90 gene in ovule of pollination development and pollination abortion
表 1 板栗CmHsp90基因家族信息
Table 1 Information of CmHsp90 gene family in Castanea mollissima
基因名称
Gene name基因ID
Gene ID亚细胞定位
Subcellular localization编码区长度
CDS length/bp外显子数目
Exon numbert编码蛋白质特性
Characteristic of the coding protein氨基酸长度
Amino acid length/aa不稳定性系数
Instability coefficient分子质量
Molecular mass/kDa等电点
Isoelectric pointCmHsp90-1 BUA.CMHBY201654 细胞质 Cytosol 2 118 4 705 39.04 81.22 5.03 CmHsp90-2 BUA.CMHBY202913 线粒体 Mitochondrion 2 415 22 804 36.97 91.02 5.26 CmHsp90-3 BUA.CMHBY204630 细胞核 Nucleus 2 127 3 708 38.93 81.11 4.94 CmHsp90-4 BUA.CMHBY213842 细胞质 Cytosol 2 112 4 703 40.96 80.74 4.97 CmHsp90-5 BUA.CMHBY214124 细胞质 Cytosol 2 118 3 705 39.89 80.67 4.97 CmHsp90-6 BUA.CMHBY216742 叶绿体 Chloroplast 2 406 19 801 46.75 91.45 4.99 CmHsp90-7 BUA.CMHBY217539 内质网
Endoplasmic reticulum2 481 14 826 36.38 94.21 4.87 CmHsp90-8 BUA.CMHBY217715 内质网
Endoplasmic reticulum2 358 15 785 36.15 89.42 4.81 CmHsp90-9 BUA.CMHBY217794 细胞质 Cytosol 2 115 4 704 40.28 80.97 4.95 CmHsp90-10 BUA.CMHBY229404 细胞质 Cytosol 2 112 4 703 37.98 80.72 5.05  下载: 导出CSV
下载: 导出CSV
-
[1] Johnson J L, Brown C. Plasticity of the Hsp90 chaperone machine indivergent eukaryotic organisms[J]. Cell Stress Chaperones, 2009, 14: 83−94. doi: 10.1007/s12192-008-0058-9
[2] 汤佳乐, 徐海, 苑平, 等. 植物Hsp90s与耐热性关系的研究进展[J]. 生物技术通报, 2020, 36(10): 173−179. doi: 10.13560/j.cnki.biotech.bull.1985.2020-0264 Tang J L, Xu H, Yuan P, et al. Advance in relationship between heat shock protein 90s and thermo-tolerance in plants[J]. Biotechnology Bulletin, 2020, 36(10): 173−179. doi: 10.13560/j.cnki.biotech.bull.1985.2020-0264
[3] Chen J, Gao T, Wan S, et al. Genome-wide identification, classification and expression analysis of the HSP gene superfamily in tea plant (Camellia sinensis)[J]. International Journal of Molecular Sciences, 2018, 19: 2633. doi: 10.3390/ijms19092633
[4] Hu W, Hu G, Han B. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice[J]. Plant Science, 2009, 176(4): 583−590.
[5] Sangster T A, Queitsch C. The HSP90 chaperone complex, an emerging force in plant development and phenotypic plasticity[J]. Current Opinion in Plant Biology, 2005, 8: 86−92. doi: 10.1016/j.pbi.2004.11.012
[6] Johnson J L. Evolution and function of diverse Hsp90 homologs and cochaperone proteins[J]. Biochim Biophys Acta, 2012, 1823: 607−613. doi: 10.1016/j.bbamcr.2011.09.020
[7] Kravats A N, Hoskins J R, Reidy M, et al. Functional and physical interaction between yeast Hsp90 and Hsp70[J]. PNAS, 2018, 115(10): E2210.
[8] Mayer M P, Breton L L. Hsp90: breaking the symmetry[J]. Molecular Cell, 2015, 58: 8−20. doi: 10.1016/j.molcel.2015.02.022
[9] Jackson S E. Hsp90: structure and function[J]. Topics in Current Chemistry, 2013, 328: 155−240.
[10] Goldberg R B, de Paiva G, Yadegari R. Plant embryogenesis: zygote to seed[J]. Science, 1994, 266: 605−614. doi: 10.1126/science.266.5185.605
[11] Samakovli D, TicháT, VavrdováT, et al. HEAT SHOCK PROTEIN 90 proteins and YODA regulate main body axis formation during early embryogenesis[J]. Plant Physiology, 2021, 186(3): 1526−1544. doi: 10.1093/plphys/kiab171
[12] Luo A, Li X, Zhang X, et al. Identification of AtHsp90.6 involved in early embryogenesis and its structure prediction by molecular dynamics simulations[J/OL]. Royal Society Open Science, 2019, 6: 190219[2021−08−11]. http://dx.doi.org/10.1098/rsos.190219.
[13] Lee Y K, Kim S H, Hong C B, et al. Heat-shock protein 90 may be involved in differentiation of the female gametophytes in Griffithsia japonica (Ceramiales, Rhodophyta)[J]. Journal of Phycology, 1998, 34: 1017−1023. doi: 10.1046/j.1529-8817.1998.341017.x
[14] 智研咨询集团. 2019年全球及中国板栗行业产量、进出口贸易分析[Z]. 中国产业信息, 2020−10−11. Zhiyan Consulting Group. Analysis of global and Chinese chestnut industry output and import and export trade in 2019[Z]. Industry Information of China, 2020−10−11.
[15] 李玲. 板栗受精作用与胚发育分子机理的初步研究[D]. 北京: 北京林业大学, 2020. Li L. Basic study on the molecular mechanism of fertilization and embryonic development in Castanea mollissima BL.[D]. Beijing: Beijing Forestry University, 2020.
[16] 杜国华, 周良骝, 谢中稳, 等. 板栗空苞机理的研究[J]. 果树学报, 1995, 12(1): 5−9. doi: 10.13925/j.cnki.gsxb.1995.01.002 Du G H, Zhou L L, Xie Z W, et al. Study on empty bud mechanism of chestnut[J]. Journal of Fruit Science, 1995, 12(1): 5−9. doi: 10.13925/j.cnki.gsxb.1995.01.002
[17] 王倩. 品种配置对燕山板栗结实情况及果实品质影响研究[D]. 北京: 北京林业大学, 2012. Wang Q. Effect on fruiting conditions and nut quality from variety configuration[D]. Beijing: Beijing Forestry University, 2012.
[18] 李玲, 苏淑钗, 寇艳茹. 板栗座果及果实早期发育与内源激素质量分数的关系[J]. 东北林业大学学报, 2020, 48(5): 55−61. doi: 10.3969/j.issn.1000-5382.2020.05.011 Li L, Su S C, Kou Y R. Fruit-set and early fruit development in chestnut are associated with endogenous hormones contents[J]. Journal of Northeast Forestry University, 2020, 48(5): 55−61. doi: 10.3969/j.issn.1000-5382.2020.05.011
[19] Letunic I, Khedkar S, Bork P. SMART: recent updates, new developments and status in 2020[J]. Nucleic Acids Research, 2021, 49(D1): D458−D460. doi: 10.1093/nar/gkaa937
[20] Chen C, Chen H, Zhang Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant, 2020, 13(8): 1194−1202. doi: 10.1016/j.molp.2020.06.009
[21] 刘阳. 板栗花发育相关 MADS-box 基因的挖掘和表达分析[D]. 北京: 北京农学院, 2019. Liu Y. Mining and expression analysis of MADS-box genes related to flower development of Castanea mollissima [D]. Beijing: Beijing University of Agriculture, 2019.
[22] Kim D, Langmead B, Salzberg S L. HISAT: a fast spliced aligner with low memory requirements[J]. Nature Methods, 2015, 12(4): 357−360. doi: 10.1038/nmeth.3317
[23] Pertea M, Pertea G M, Antonescu C M, et al. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads[J]. Nature Biotechnology, 2015, 33(3): 290−295. doi: 10.1038/nbt.3122
[24] Mayer M P, Bukau B. Molecular chaperones: the busy life of Hsp90[J]. Current Biology, 1999, 9: R322−R325. doi: 10.1016/S0960-9822(99)80203-6
[25] Krishna P, Gloor G. The Hsp90 family of proteins in Arabidopsis thaliana[J]. Cell Stress Chaperones, 2001, 6: 238−246. doi: 10.1379/1466-1268(2001)006<0238:THFOPI>2.0.CO;2
[26] 张海. 水稻OsHSP90基因家族功能研究[D]. 雅安: 四川农业大学, 2016. Zhang H. Function analysis of the OsHSP90 family in rice[D]. Ya’an: Sichuan Agricultural University, 2016.
[27] Zhang J, Liu B B, Zhang L, et al. Genome-wide analysis of the Populus Hsp90 gene family reveals differential expression patterns, localization, and heat stress responses[J]. BMC Genomics, 2013, 13: 532.
[28] 杜志如. 水稻热激蛋白Hsp90基因的克隆及互作蛋白的筛选[D]. 雅安: 四川农业大学, 2008. Du Z R. Cloning of heat shock protein90 gene from rice and screening for interacted proteins[D]. Ya’an: Sichuan Agriculture University, 2008.
[29] Feng J, Fan P, Jiang P, et al. Chloroplast-targeted Hsp90 plays essential roles in plastid development and embryogenesis in Arabidopsis possibly linking with VIPP1[J]. Plant Physiology, 2014, 150(2): 292−307. doi: 10.1111/ppl.12083
[30] Panaretou B, Prodromou C, Roe S M, et al. ATP binding and hydrolysis areessential to the function of the Hsp90 molecular chaperone in vivo[J]. The EMBO Journal, 1998, 17(16): 4829−4836. doi: 10.1093/emboj/17.16.4829
[31] 袁凌云, 吴颖, 张利婷, 等. 白菜HSP90基因家族的鉴定及表达分析[J/OL]. 分子植物育种, 2012[2021−06−30]. https://kns.cnki.net/kcms/detail/46.1068.S.20210311.1553.004.html. Yuan L Y, Wu Y, Zhang L T, et al. Identification and expression analysis of HSP90 gene family in Chinese cabbage[J/OL]. Molecular Plant Breeding, 2012[2021−06−30]. https://kns.cnki.net/kcms/detail/46.1068.S.20210311.1553.004.html.
[32] 刘云飞, 万红建, 杨悦俭, 等. 番茄热激蛋白90的全基因组鉴定及分析[J]. 遗传, 2014, 36(10): 1043−1052. Liu Y F, Wan H J, Yang Y J, et al. Genome-wide identification and analysis of heat shock protein 90 in tomato[J]. Hereditas, 2014, 36(10): 1043−1052.
[33] Song Z P, Pan F L, Yang C, et al. Genome-wide identification andexpression analysis of HSP90 gene family in Nicotiana tabacum[J/OL]. BMC Genet, 2019, 20(1): 35[2021−06−30]. https://pubmed.ncbi.nlm.nih.gov/30890142/.
[34] Collins G G, Nie X L, Saltveit M E. Heat shock protein and chilling sensitivity of mung bean hypcotyls[J]. Journal Experimental Botany, 1995, 46(7): 795−802. doi: 10.1093/jxb/46.7.795
[35] Sabehat A, Weiss D, Lurie S, et al. The correlation between heat-shock protein accumulation and persistence and chilling tolerance in tomato fruit[J]. Plant Physiology, 1996, 110: 531−537. doi: 10.1104/pp.110.2.531
[36] Gilmour S J, Hajela R K, Thomashow M F. Cold acclimation in Arabidopsis thaliana[J]. Plant Physiology, 1988, 87: 745−750. doi: 10.1104/pp.87.3.745
[37] Prasinos C, Krampis K, Samakovli D, et al. Tight regulation of expression of two Arabidopsis cytosolic Hsp90 genes during embryo development[J]. Journal of Experimental Botany, 2005, 56(412): 633−644. doi: 10.1093/jxb/eri035
[38] Sangster T A, Bahrami A, Wilczek A, et al. Phenotypic diversity and altered environmental plasticity in Arabidopsis thaliana with reduced Hsp90 levels[J/OL]. PLoS One, 2007, 2(7): e648[2021−06−30]. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0000648.
-
期刊类型引用(6)
1. 冯林艳,周火艳,赵晓迪. 乌兰布和沙漠两种植物的分布格局及其变化. 南京林业大学学报(自然科学版). 2024(01): 155-160 .  百度学术
百度学术
2. 曾红,徐永艳,邵琳亚,闻永慧,夏小丽,汪琼. 4种植物叶片浸提液成分分析及其对珊瑚樱种子萌发的影响. 西南林业大学学报(自然科学). 2023(04): 39-46 . 百度学术
3. 马光宗,徐高峰,杨韶松,杨云海,张付斗,温丽娜,陶琼,申时才,叶敏. 甘薯提取物对3种杂草种子萌发和幼苗生长的化感作用. 西南农业学报. 2022(06): 1295-1302 . 百度学术
4. 路文杰,佛芒芒,肖毅,王永新,杜利霞,钟华,赵祥,董宽虎. 草地植物凋落物浸提液对根际微生物碳源利用的影响. 中国草地学报. 2021(06): 35-42 . 百度学术
5. 张林媚,刘姝玲,郭彩云. 立地条件对榆林沙区樟子松嫁接红松生长的影响. 林业科技通讯. 2021(11): 32-37 . 百度学术
6. 王方琳,尉秋实,柴成武,王理德,张德魁,王昱淇,王飞,胡小柯. 沙蒿(Artemisia desertorum)浸提液对自身种子萌发与幼苗生长的化感作用. 中国沙漠. 2021(06): 21-28 . 百度学术
其他类型引用(3)























计量
- 文章访问数: 718
- HTML全文浏览量: 159
- PDF下载量: 88
- 被引次数: 9
