
Stand biomass growth model of broadleaved forest with parameter classification in Guangdong Province of southern China
-
摘要:目的 建立区域尺度林分生物量生长模型,为预测未来某一时段广东省天然阔叶林生物量和碳储量提供方法学支持。方法 基于广东省1997—2017年5期森林清查数据,选择栎类、木荷和其他软阔类等6个阔叶树种为优势树种的203个天然林样地,以参数分级反映立地质量差异,以竞争指数表示密度影响,以分步建模(一元非线性回归法)和联合建模(非线性联立方程组法)区别建模方式,采用理论生长方程构建胸径生长模型估计林龄进而构建多种林分生物量生长模型,以决定系数和平均预估误差等4个指标评价模型拟合优度;对拟合优度较高的模型,以2002—2017年4期连清的183块样地为检验样本,用总相对误差来验证其应用效果。结果 对比模型拟合效果和区域尺度及样地水平上的估计精度,以探究林分密度、不同参数分级、分级方法和建模方法共4项影响因素对生物量生长模型的效果,结果表明:非线性联立方程组法优于分步建模法;与生长速度有关的参数b分级模型优于与生长潜力有关的参数a分级模型;考虑林木竞争和分级方程中加入竞争指数对优化模型性能影响不大。参数b分级、自变量和分级方程皆不含竞争指数的联合模型(模型10)为最优模型,其生物量生长模型确定系数R2为0.970 1;预测4期生物量时,估计效果较好,后期估计误差明显低于前期,如采用模型10预估栎类2002—2017年区域尺度生物量时,4期的估计误差分别为6.22%、15.27%、4.80%、−1.84%。结论 以Richards理论生长方程为基础构建林分生物量生长模型来估测区域尺度生物量是一种可行的方法,为评估未来某一时段区域尺度森林生态系统的固碳能力提供依据,也为其他区域的林分生物量生长模型研建提供参考。Abstract:Objective A regional-scale stand biomass growth model was established to provide methodological support for predicting the biomass and carbon storage of natural broadleaved forests in Guangdong Province of southern China in the future.Method Based on the five forest inventory data of Guangdong Province from 1997 to 2017, 203 natural forest sample plots with six broadleaved tree species such as Quercus spp., Schima superba and other soft broadleaved species as dominant tree species were selected. The site quality difference was reflected by parameter classification, the density effect was expressed by competition index, and the modeling method was distinguished by step-by-step modeling (univariate nonlinear regression method) and joint modeling (nonlinear simultaneous equations method). The DBH growth model, constructed by the theoretical growth equation, was used to estimate the stand age, and then various stand biomass growth models were constructed. The goodness of fit of the model was evaluated by four indexes such as determination coefficient and average prediction error. For the model with high goodness of fit, 183 sample plots by continuously inventory in four periods from 2002 to 2017 were taken as test samples, and the total relative error was used to verify its application effect.Result To compare the fitting effect and the estimation accuracy at regional scale and sample plot level for exploring the influence of four factors including stand density, different parameter classification, classification method and modeling method on the biomass growth model, it was found that nonlinear simultaneous equation was better than step-by-step modeling; the classification of model parameter b related to growth rate was better than that of model parameter a related to growth potential; considering the stand density and adding competition index to the hierarchical equation had little effect on optimizing model performance. Based on the classification of parameter b, the joint model without competition index in independent variable and the hierarchical equation was the optimal model, i.e. Model 10. The determination coefficient of the biomass growth model was 0.970 1. When Model 10 was used to predict the biomass of four periods, the prediction effect was good. But the estimation error in the later stage was significantly lower than that in the earlier stage. For example, when Model 10 was used to estimate the biomass of Quercus spp. at regional scale from 2002 to 2017, the estimation errors of four periods were 6.22%, 15.27%, 4.80% and −1.84%, respectively.Conclusion It is a feasible method to establish stand biomass growth model based on the Richards growth equation to estimate regional-scale biomass, which not only provides a basis for evaluating the carbon sink capacity of forest ecosystem at regional scale in a certain period in the future, but also provides a reference for the construction of stand biomass growth model in other regions.
-
在树木的生活史中,幼树阶段的生长和定居非常关键,决定着其种群的更新与发展[1]。分析幼树的生长状况、空间分布与环境之间的相互关系有助于揭示其在生活史早期对环境的适应机制,对制定科学的物种保育措施有着重要的意义[2]。幼树生活在森林下层,光照是其生长发育的主要限制因子[3],受到林冠层结构的影响显著[4]。此外,林下幼树生长还与物种的耐阴性有关[5−6]。
林窗是群落中因树木死亡而形成的不连续的林冠空隙[7],对森林更新具有重要意义。林窗会改变林下的光照强度[8]、温湿度[9]以及化感作用[10]等环境条件,增加了林分空间结构的复杂性和林下生境的异质性,并显著影响林下幼树的分布和生长发育[11]。精确的林窗描述可以更好地模拟林窗干扰对林下幼树的作用,推断生态学机制[12−14]。研究林窗对林下幼树的影响,如何准确得量化林窗空间结构是关键。传统方法常使用鱼眼镜头拍摄、椭圆模型拟合等方法得到冠层林窗结构,但是这些都很难测定林窗连续的空间变化和准确结构信息。激光雷达能够精确得呈现目标物的三维结构信息以获得这些极具价值但过去难以测量的林分参数,相较于传统方法,可以更加真实的提取到林窗的面积、形状和位置等特征。
黄檗(Phellodendron amurense)是芸香科(Rutaceae)黄檗属(Phellodendron)的落叶乔木[15],在2021年公布的《国家重点保护野生植物名录》中被列为国家二级重点保护野生植物。由于过去人为破坏严重,致使其种群数量急剧减少,因此开展黄檗的迁地保护研究对其种群的恢复非常重要。我们于2014年在北京百花山自然保护区实验区内的两块样地中随机种植了800株黄檗幼树,经过7年自然淘汰后,于2021年调查了存活黄檗的生长状况和空间分布情况,本文据此数据揭示了黄檗幼树对环境的适应,尤其是对林窗引起的环境变化的适应,以期为黄檗的迁地保护提供理论依据。
1. 研究区概况与研究方法
1.1 研究区概况
百花山国家级自然保护区位于北京市门头沟区清水镇境内( 115°25′ ~ 115°42′E,39°48′ ~ 40°05′N),总面积为21 743.1 hm2。该地区年平均降水量450 ~ 720 mm,年平均温度6 ~ 7 ℃,最热月8月份,平均温度22 ℃;最冷月1月份,平均温度−5.7 ℃,全年无霜期110 d左右。土壤为本地区地带性土类褐色土。百花山国家级自然保护区分布有野生黄檗种群,气候适宜黄檗的生长。
课题组于2014年在自然保护区实验区内建立2块40 m × 40 m的固定样地。两个样地均为1970年开始营造的人工林,未经择伐和抚育。样地A(115°34′56″E,39°50′11″N)是以胡桃楸(Juglans mandshurica)和华北落叶松(Larix gmelinii var. principis-rupprechtii)为建群种的针阔混交林,伴生树种有蒙古栎(Quercus mongolica),郁闭度0.7,林分密度为650株/hm2,平均胸径18.3 cm,平均树高11.1 m,平均冠幅4.8 m。样地B(115°34′49″E,39°49′52″N)是以胡桃楸和华北落叶松为建群种的针阔混交林,伴生树种有白桦(Betula platyphylla)、北京丁香(Syringa pekinensis)等,郁闭度0.7,林分密度为600株/hm2,平均胸径21.1 cm,树高12.1 m,平均冠幅5.3 m。研究区域位于自然保护区封闭区域,人为干扰较弱,群落中的乔木树种均为野生黄檗的常见伴生树种。在每个样地内随机种植黄檗幼树400株,共800株,平均密度0.25株/m2,栽种的黄檗幼树为同一批次苗圃苗,其平均基径为1 cm,平均高度为1 m。
1.2 数据收集与处理
(1)样地植物数据的收集。经过7年自然生长和淘汰,我们于2021年9月对样地内的黄檗进行了调查,调查样地内所有存活黄檗幼树的基径、冠幅、树高并记录在样地内的相对坐标。同时对样地中的枯立木和幼龄乔木也进行了调查,记录树种、胸径、树高和冠幅。枯立木通过树皮和枝干形态辨认种类,其冠幅通过样地中同种个体的胸径和冠幅比计算。
(2)样地环境数据的收集与处理。使用Li Backpack DGC50背包式激光雷达对样地进行采样,获取样地内所有植物的三维点云数据,采样路径如图1所示。
![]() 图 1 行走路径规划①为起点,沿Z字形走到② Start from ① and walk along the Z-shaped path to ②Figure 1. Walking path planning
图 1 行走路径规划①为起点,沿Z字形走到② Start from ① and walk along the Z-shaped path to ②Figure 1. Walking path planning使用LiDAR360软件对样地的点云数据进行去噪、滤波、根据地面点分类、根据地面点归一化等预处理。地面激光雷达点云是从冠层下面获取,可以清晰地识别树干,并以此分割出单木,使用点云分割工具将预处理完的点云数据进行单木分割,然后用单木点云编辑工具将林下高度低于6 m的低矮乔灌点云删除后,再按树ID提取点云[16],得到样地的冠层图(图2)。最后将点云数据转为栅格大小为0.1 m的TIFF文件,导入到ArcGIS 10.2中进行下一步分析。
由于研究目的和研究区域不同,对林窗的判定标准存在差异[17−19]。依据前人研究,我们将面积大于4 m2的林间空隙定义为林窗区域[20−21],并按照Omelko等[22]的方法,依据个体所处位置来划分其与林窗的关系类型,由于黄檗幼树树冠平均半径为1 m,在林窗边缘1 m范围内的黄檗幼树树冠不是完全处于林窗或林冠下的,所以我们将林窗边缘向林冠内1 m和向林窗中心1 m的范围定为林窗边缘,以此将样地的林冠覆盖情况分为林窗中心、林窗边缘和林冠区3类生境。
1.3 数据分析
1.3.1 空间点格局分析
点格局以每个树木个体的空间坐标为基本数据,所有个体构成了二维空间上种群空间分布的点图[23],Ripley’s K(r)函数可以从多尺度上研究种群分布格局,是空间格局分析最主要的方法[24]。Ripley’s K(r)函数是以任意点为圆心,r为半径的圆内期望点数与样方点密度的比值[25],公式为
K(r)=An2n∑i=1n∑j=1Ir(dij)Wij(i≠j) 式中:A为样地面积;n为样地内黄檗数量;Ir(dij)为指示函数,dij为圆心i和圆心j两点间的距离,当dij ≤ r时,Ir(dij) = 1,否则Ir(dij) = 0;Wij为边缘矫正的权重。
L(r)函数为K(r)函数的修正函数,可以更直观地解析种群空间格局,其公式为
L(r)=√K(r)/π−r 在自然群落中,很难区分幼树的空间分布格局是源于环境异质性,还是因由于种子扩散、母树分蘖等扩散限制因素所致[26],通常会选择Thomas等聚集型性零模型模拟扩散限制以消除对幼树空间分布的影响[27]。本研究中黄檗幼树为随机栽种,样本之间相互独立,所以采用完全随机零模型来分析黄檗幼树的分布格局,使用Monte Carlo方法检验观测点的L(r)值偏离随机分布的显著程度。通过完全随机模拟技术得到显著性为95%的置信区间,如果L(r)值分布在置信区间内,则分布格局为随机分布;在置信区间之上为聚集分布,在置信区间之下为均匀分布[28];研究尺度r的取值范围为0 ~ 15 m,在R 4.02中使用spatstat包完成。
1.3.2 生境关联性检验
生境关联分析时,首先要进行生境类型划分[29],有多种方法和标准来决定分类结果,比如Zhang等[2]使用多元回归树进行划分,Zuleta等[30]使用了Ward最小方差法,但目前还没有固定统一的方法进行划分。本文以乔木层树冠覆盖情况作为变量划分生境类型,将40 m × 40 m的样地划分为1 600个1 m × 1 m的小样格,根据样地内的树冠和林窗的分布情况,利用ArcGIS的空间统计分析功能,将每个小样格划分为林窗中心、林窗边缘和林冠区3种生境类型(以下称为生境1、生境2、生境3)中的一种。
生境关联分析同样采用物种完全随机模型(complete spatial randomness,CSR)作为零模型,检验幼树与生境之间的关联性。利用CSR模型在空间上随机生成相同数量的植株,关联真实的生境地图,计算每种生境类型中黄檗幼树的多度,重复上述过程1 000次后得到黄檗幼树在不同生境中的多度分布(视其为零分布)。在显著水平α = 0.05条件下,比较幼树在不同生境中真实分布和模拟分布的多度 [2]。
1.3.3 幼树个体生长与生境的关系
由于研究中黄檗幼树均为同一批次、苗圃1年生黄檗幼树,所以本次测量的幼树基径、树高和冠幅可以视为黄檗的生长量,以表征植株对环境因子的响应。利用S-W检验样本正态性,使用独立样本t检验比较不同生境的黄檗幼树生长指标,并在R中进行统计分析和绘图。
2. 结果与分析
2.1 黄檗幼树的空间分布格局
两块样地中共有154株黄檗幼树存活。其中样地A存活79株,存活率19.75%;样地B存活75株,存活率18.75%。基于CSR模型的点格局分析显示(图3),存活黄檗幼树的空间分布呈现明显的聚集格局,样地A的幼树在2 ~ 15 m尺度上表现为聚集分布,样地B幼树在3 ~ 9 m尺度上表现为聚集分布。
![]() 图 3 黄檗幼树空间点格局分析Figure 3. Spatial point pattern analyses of Phellodendron amurense saplings
图 3 黄檗幼树空间点格局分析Figure 3. Spatial point pattern analyses of Phellodendron amurense saplings2.2 黄檗幼树的生境关联性分析
黄檗样地的林窗空间格局如图4所示,2个样地共提取林窗15个,其中样地A 8个,样地B 7个,单个林窗面积4.1 ~ 155.4 m2。样地A中华北落叶松数量较多,树冠相对较小并且稀疏,林窗形状相对狭长;样地B中由于树木枯死,留下较大的冠层空隙。
按照上述生境划分的方法将样地划分为3种不同的生境类型(图5),样地A中林窗中心占总比例的7.31% (117个),林窗边缘占32.62% (522个),林冠区占60.06% (961个);样地B中林窗中心占10% (160个),林窗边缘占29.43% (471个),林冠区占60.5%(969个)。物种完全随机模型检验结果表明(表1),黄檗幼树在2个样地的空间分布均与林窗边缘呈显著正相关,与林冠区显著负相关,与林窗中心没有明显倾向性。林窗中心密度为0.076株/m2和0.043株/m2,林窗边缘为0.088株/m2和0.093株/m2,林冠区为0.024株/m2和0.029株/m2。黄檗幼树在各生境类型中的存活密度梯度为林窗边缘 > 林窗中心 > 林冠区,林冠结构与存活黄檗幼树的空间分布有显著关联性,林窗边缘的生境条件最适宜黄檗存活。
![]() 图 5 样地生境类型划分每个小样格为1 m × 1 m。小样格内的数字表示生境类型。不同的颜色代表不同的生境类型,蓝色为林窗中心,标记为生境类型1;黄色为林窗边缘,标记为生境类型2;绿色为林冠区,标记为生境类型3。The size of each plot is 1 m × 1 m. The number in the small square indicates the habitat type. Different colors represent varied habitat types, blue indicates the center of forest gaps, marked as habitat type 1; yellow indicates the edge of forest gaps, marked as habitat type 2; green indicates the area of canopy, marked as habitat type 3.Figure 5. Classification of habitat types in the sample plot表 1 幼树与每种生境类型的关联性Table 1. Association between saplings and each habitat type
图 5 样地生境类型划分每个小样格为1 m × 1 m。小样格内的数字表示生境类型。不同的颜色代表不同的生境类型,蓝色为林窗中心,标记为生境类型1;黄色为林窗边缘,标记为生境类型2;绿色为林冠区,标记为生境类型3。The size of each plot is 1 m × 1 m. The number in the small square indicates the habitat type. Different colors represent varied habitat types, blue indicates the center of forest gaps, marked as habitat type 1; yellow indicates the edge of forest gaps, marked as habitat type 2; green indicates the area of canopy, marked as habitat type 3.Figure 5. Classification of habitat types in the sample plot表 1 幼树与每种生境类型的关联性Table 1. Association between saplings and each habitat type样地
Sample plot生境1 Habitat 1 生境2 Habitat 2 生境3 Habitat 3 幼树数量
Sapling number显著性
Significance幼树数量
Sapling number显著性
Significance幼树数量
Sapling number显著性
SignificanceA 9 N 46 + 24 − B 7 N 44 + 24 − 注:+.正关联;−.负关联;N.中性。Notes: +, positively associated; −, negatively associated; N, neutral. 2.3 幼树个体发育与其所处位置的关系
物种对不同生境的响应和利用存在差异,其基径、树高和冠幅大小等性状是植株对其所处生境适应性的表现。黄檗幼树长势的t检验结果表明(图6),冠层结构对林下黄檗幼树的生长产生了显著的影响,黄檗幼树基径的生长状况在林窗边缘显著高于林窗中心和林冠区(P < 0.01),但在林冠区与林窗中心之间没有显著差异。黄檗的冠幅和树高则呈现显著的梯度变化(P < 0.05),即林窗边缘 > 林冠区 > 林窗中心。
![]() 图 6 黄檗幼树在各生境类型中的生长状况Figure 6. Growth status of Phellodendron amurense saplings in various habitat types
图 6 黄檗幼树在各生境类型中的生长状况Figure 6. Growth status of Phellodendron amurense saplings in various habitat types3. 讨 论
分析树木的空间分布可以推断该种群对环境的适应性[12],幼树空间分布主要受生境异质性和扩散限制因素的影响[31−32],在已经排除扩散限制的前提下,样地中存活黄檗的空间格局在0 ~ 4 m的小尺度上没有表现出明显的倾向性,但随着尺度增大,逐渐表现为聚集分布格局(图3),说明黄檗的分布格局主要受到生境异质性的影响。林下的微生境条件通常分布不均匀,有限资源的斑块状分布会影响植株的空间格局[33],导致黄檗个体在样地中的某些区域中存活率更高,经过7年的自然生长和死亡,我们发现黄檗对不同生境的适应性不同。
林窗对林下生境最直接的影响是改变生境中的光照条件,但林下的光环境并不是同质的,大林窗比小林窗可获得更多的光照,林窗中心的光照强度高于林窗边缘[34−35]。黄檗幼树基径的生长状况在林窗边缘的显著高于其他区域,而在林冠区和林窗中心之间没有显著差异;黄檗幼树的冠幅和树高则呈现显著的梯度变化(P < 0.05),即林窗边缘 > 林冠区 > 林窗中心(图6)。耐阴的树种在弱光环境中会在基径、树高和树冠中进行碳的重分配以适应低光环境,通常会通过降低基径来增加树高和冠幅生长的方式以最大程度获取阳光[3],植株表现为“细长”而“冠大”的特征。林冠区的黄檗幼树可能采取了类似的生长策略,将更多碳分配于植株高度和树冠的生长,以提高其对光的截获能力,所以其树高和树冠相较于林窗中心长势更好;在林窗中心,由于光照相对更强烈,树木暴露在无遮蔽的强光下,得到的有效光辐射可能超出了它们所能利用的最高水平,抑制了其树冠和树高的生长;在林窗边缘的中等遮蔽条件下,黄檗的生长状况好于林窗中心和林冠下,这与前人研究结论一致,李霞等[36]和张玲等[37]对不同遮荫条件下黄檗形态特征、生理指标的研究中发现轻度和中度遮蔽处理有利于黄檗植株地上形态生长,并增加叶绿素含量以适应遮荫环境,但重度遮蔽会降低其可溶性糖的积累。
不适宜的生境会抑制植物的生长速度,甚至导致个体死亡,存活个体的空间分布可以表示其对特定生境的倾向性[38]。生境关联分析的结果表明,存活黄檗幼树的空间分布与林窗边缘呈显著正关联,与林冠区显著负关联,而林窗中心对其存活没有显著影响(表1),黄檗幼树在各类生境的存活密度梯度为林窗边缘 > 林窗中心 > 林冠区,这与黄檗生长状况的结果存在差异,虽然黄檗树高和树冠的长势在林冠下高于林窗中心,但可能是其在弱光环境胁迫下引起的徒长现象[39],长期的逆境胁迫最终会导致其死亡。
即使在相同的林冠环境下,不同植物的幼苗存活率也可能存在显著差异,这与该物种对林下遮蔽环境的耐受性[40]有关,顶级种的幼苗通常生活在郁闭度高的森林下层,其幼年期可以忍耐较荫蔽的环境,而先锋种幼苗则需要较强的光照[41]。在野生黄檗分布及生长的研究中,通常认为黄檗为喜光植物[15,42−43],而试验条件下的黄檗幼苗在全光照下受到抑制,更适宜中等遮蔽的光照条件[36−37]。这可能是因为研究中黄檗的生活史阶段不同,对野生黄檗的研究多为成年木,而试验条件下的黄檗为幼年期。结合前人研究,我们认为黄檗在幼树期具有一定的耐阴性,但如其他演替后期物种一样[44],黄檗需要借助林窗的帮助以到达主林层,所以林窗边缘的中等光照条件最适宜其定植。
此外,枯立木和幼龄乔木可以表示样地的林分动态和冠层结构变化情况。森林中的枯木分解时间较长,枯立木可以证明其所处位置在过去几年曾有过树冠遮蔽。在样地调查过程中,我们对研究区域的枯立木及幼龄乔木的数量和分布进行了记录(图5),但研究区域的枯立木数量很少,这可能有两个原因:(1)关于冠层扰动的研究表明,在10年尺度上的冠层扰动不会很多[22],我们的样地布置时间相对而言仍较短;(2)人类活动干扰造成的树木死亡是林窗形成的重要原因,我们的研究区域位于自然保护区中且没有进行人工择伐和抚育,人类干扰较弱,从而没有较多的冠层扰动情况。幼龄乔木可以代表其所处位置在过去几年可能没有树冠遮蔽,我们在点云数据处理时,已将低矮的幼龄乔木去掉。所以本文认为冠层动态在研究区域较弱,没有进行冠层动态对林下黄檗幼树影响的研究。
4. 结 论
本文分析了林窗空间格局对林下黄檗幼树生长和分布的影响,研究结果表明,林窗空间结构导致的林下异质性生境是影响黄檗幼树生长发育和空间分布的重要原因,林窗中心和林冠区都会对黄檗幼树的生长发育产生抑制,林窗边缘的生境条件更适宜黄檗存活。如果对黄檗进行迁地保护,建议在微生境选择上,以林窗边缘为宜,或者在其偏上方开辟人工林窗,可以为黄檗幼树的生长和存活提供适宜的生境条件。
-
![]()
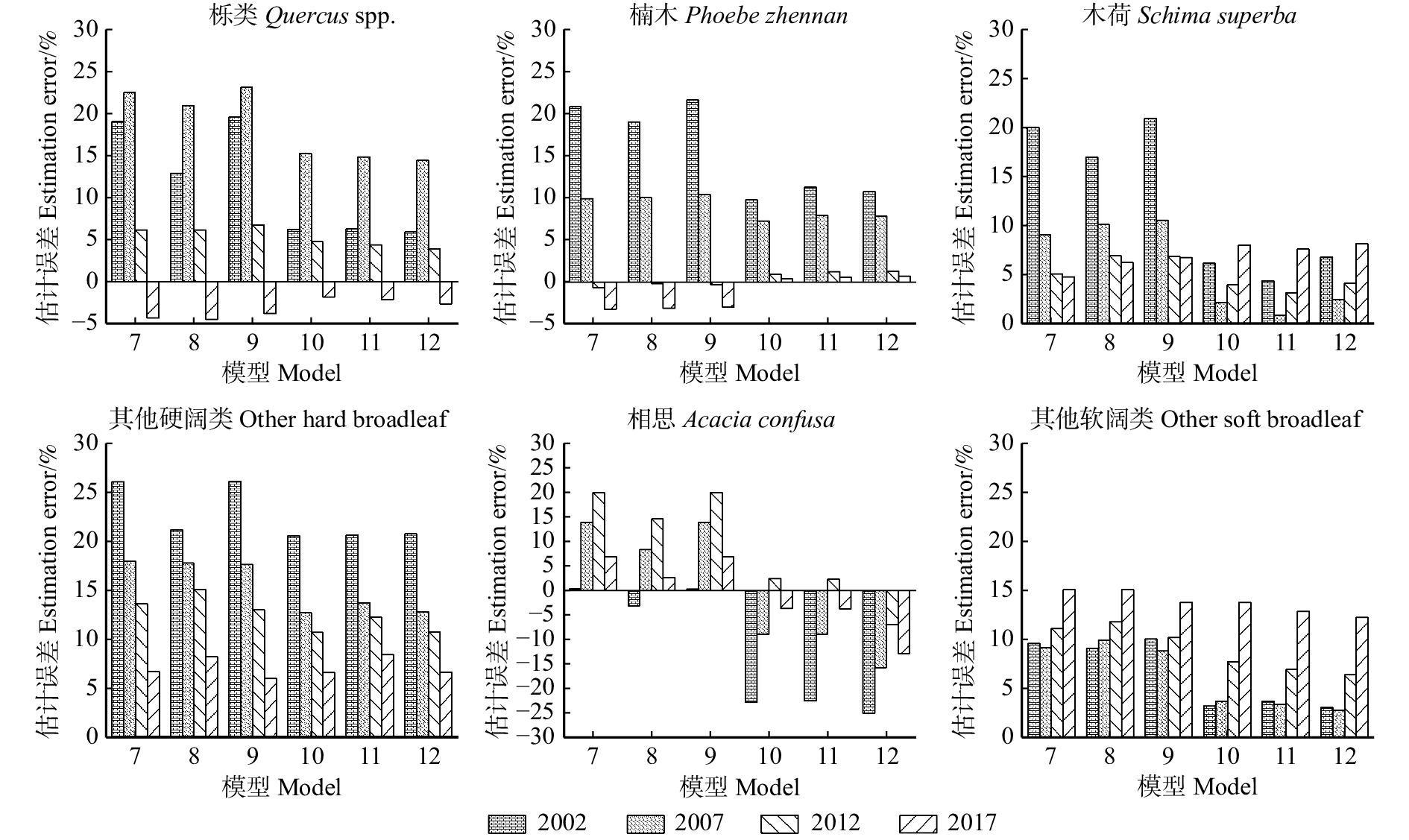
图 2 6种模型预测区域水平生物量的误差比较
Figure 2. Error comparison of six models in predicting regional biomass
![]()
图 3 预测栎类、楠木和木荷样地生物量的估计误差
Figure 3. Estimation error of biomass for Quercus spp., Phoebe zhennan and Schima superba sample plots
![]()
图 4 预测其他硬阔类、相思和其他软阔类样地生物量的估计误差
Figure 4. Estimation error of biomass for predicting other hard broadleaf, Acacia confusa and other soft broadleaf sample plots
表 1 建模数据统计特征
Table 1 Statistical characteristics of modeling data
树种 Tree species 变量 Variable 年份 Year 1997 2002 2007 2012 2017 栎类
Quercus spp.胸径 DBH/cm 10.7 ± 4.0 13.7 ± 4.2 15.8 ± 4.4 17.5 ± 4.7 19.5 ± 5.1 生物量/(t·hm−2) Biomass/(t·ha−1) 17.4 ± 23.8 22.3 ± 21.7 29.2 ± 25.7 42.4 ± 39.0 53.6 ± 44.2 楠木
Phoebe zhennan胸径 DBH/cm 10.8 ± 3.3 13.6 ± 3.9 15.5 ± 4.1 17.3 ± 4.3 19.4 ± 5.0 生物量/(t·hm−2) Biomass/(t·ha−1) 10.1 ± 10.8 15.3 ± 14.4 19.7 ± 17.0 24.7 ± 20.9 30.0 ± 23.1 木荷
Schima superba胸径 DBH/cm 10.0 ± 3.6 13.0 ± 3.7 15.1 ± 3.9 16.9 ± 4.3 18.7 ± 4.6 生物量/(t·hm−2) Biomass/(t·ha−1) 8.6 ± 8.6 15.4 ± 14.5 20.4 ± 18.3 24.1 ± 19.7 29.6 ± 23.1 其他硬阔类
Other hard broadleaf胸径 DBH/cm 10.9 ± 3.3 13.5 ± 3.9 15.2 ± 4.3 16.8 ± 4.8 18.4 ± 5.3 生物量/(t·hm−2) Biomass/(t·ha−1) 13.5 ± 16.8 19.3 ± 22.3 24.7 ± 26.4 29.7 ± 30.5 37.2 ± 35.8 相思
Acacia confusa胸径 DBH/cm 9.1 ± 1.1 14.1 ± 2.9 16.6 ± 4.0 18.9 ± 4.6 21.3 ± 4.9 生物量/(t·hm−2) Biomass/(t·ha−1) 10.5 ± 5.9 33.5 ± 20.8 51.3 ± 32.3 71.2 ± 44.1 92.8 ± 54.0 其他软阔类
Other soft broadleaf胸径DBH/cm 9.8 ± 2.4 12.7 ± 3.3 14.8 ± 4.0 16.3 ± 4.5 18.3 ± 4.8 生物量/(t·hm−2) Biomass/(t·ha−1) 5.5 ± 4.1 12.3 ± 8.6 16.7 ± 11.0 20.8 ± 13.0 23.8 ± 15.3 注:数据为平均值 ± 标准差。下同。Notes: data is average ± standard deviation.The same below.  下载: 导出CSV
下载: 导出CSV
表 2 验证数据统计特征
Table 2 Statistical characteristics of validation data
树种 Tree species 变量 Variable 年份 Year 2002 2007 2012 2017 栎类
Quercus spp.胸径 DBH/cm 9.0 ± 2.3 11.8 ± 2.7 14.3 ± 3.3 17.2 ± 4.0 生物量/(t·hm−2) Biomass/(t·ha−1) 6.3 ± 5.4 11.7 ± 9.4 21.2 ± 17.0 32.4 ± 24.4 楠木
Phoebe zhennan胸径 DBH/cm 8.6 ± 2.0 11.5 ± 2.3 14.3 ± 3.2 16.7 ± 3.8 生物量/(t·hm−2) Biomass/(t·ha−1) 4.4 ± 3.3 8.4 ± 5.6 13.8 ± 7.6 18.8 ± 9.1 木荷
Schima superba胸径 DBH/cm 8.3 ± 1.3 11.5 ± 1.9 14.1 ± 2.7 16.5 ± 3.2 生物量/(t·hm−2) Biomass/(t·ha−1) 5.5 ± 4.6 11.3 ± 8.5 18.3 ± 14.1 25.4 ± 19.4 其他硬阔类
Other hard broadleaf胸径 DBH/cm 9.5 ± 2.7 12.0 ± 3.1 14.1 ± 3.6 16.1 ± 4.1 生物量/(t·hm−2) Biomass/(t·ha−1) 4.6 ± 2.7 8.3 ± 5.2 12.4 ± 8.1 17.7 ± 11.9 相思
Acacia confusa胸径 DBH/cm 8.5 ± 1.9 11.8 ± 3.5 14.9 ± 4.6 18.4 ± 4.9 生物量/(t·hm−2) Biomass/(t·ha−1) 4.4 ± 1.7 8.7 ± 3.1 15.9 ± 10.2 27.5 ± 20.5 其他软阔类
Other soft broadleaf胸径 DBH/cm 9.5 ± 2.1 12.0 ± 2.7 14.3 ± 3.3 16.6 ± 3.7 生物量/(t·hm−2) Biomass/(t·ha−1) 5.1 ± 5.4 8.6 ± 8.1 12.5 ± 10.8 16.5 ± 13.8
下载: 导出CSV
表 3 不同分级模式下各立地质量等级所对应的样地数量
Table 3 Number of sample plots corresponding to each site quality grade under different classification modes
分级模式
Classification mode立地质量等级
Site quality grade1 2 3 4 5 6 7 AC7 22 48 36 44 23 24 6 BC7 53 67 45 28 5 4 1 AC7-1 17 48 43 34 33 21 7 BC7-1 49 68 48 28 5 4 1 注:AC7表示对参数a分级,BC7表示对参数b分级,AC7-1表示对参数a分级且分级方程中含有竞争指数,BC7-1表示对参数b分级且分级方程中含有竞争指数。Notes: AC7 indicates that the parameter a is graded. BC7 indicates that the parameter b is graded. AC7-1 indicates that the parameter a is graded and the competition index is included in the hierarchical equation. BC7-1 indicates that the parameter b is graded and the competition index is included in the hierarchical equation.
下载: 导出CSV
表 4 12种模型
Table 4 Twelve models
模型 Model 方程 Equation 模型 Model 方程 Equation 1 Dgij=am1(1−e−bk1⋅(AGEi+ADj)) BIOij=am2(1−e−bk2⋅PAGEij)ck2 7 {Dgij=am1(1−e−bk1⋅(AGEi+ADj))BIOij=am2(1−e−bk2⋅(AGEi+ADj))ck2 2 Dgij=a′m1(1−e−bk1⋅(eJUTij)b0⋅(AGEi+ADj)) BIOij=a′m2(1−e−bk2⋅PAGEij)ck2 8 {Dgij=a′m1(1−e−bk1⋅(eJUTij)b0⋅(AGEi+ADj))BIOij=a′m2(1−e−bk2⋅(AGEi+ADj))ck2 3 Dgij=am1(1−e−bk1⋅(eJUTij)b0⋅(AGEi+ADj)) BIOij=am2(1−e−bk2⋅PAGEij)ck2 9 {Dgij=am1(1−e−bk1⋅(eJUTij)b0⋅(AGEi+ADj))BIOij=am2(1−e−bk2⋅(AGEi+ADj))ck2 4 Dgij=ak1(1−e−bm1⋅(AGEi+ADj)) BIOij=ak2(1−e−bm2⋅PAGEij)ck2 10 {Dgij=ak1(1−e−bm1⋅(AGEi+ADj))BIOij=ak2(1−e−bm2⋅(AGEi+ADj))ck2 5 Dgij=ak1(1−e−b′m1⋅(eJUTij)b0⋅(AGEi+ADj)) BIOij=ak2(1−e−b′m2⋅PAGEij)ck2 11 {Dgij=ak1(1−e−b′m1⋅(eJUTij)b0⋅(AGEi+ADj))BIOij=ak2(1−e−b′m2⋅(AGEi+ADj))ck2 6 Dgij=ak1(1−e−bm1⋅(eJUTij)b0⋅(AGEi+ADj)) BIOij=ak2(1−e−bm2⋅PAGEij)ck2 12 {Dgij=ak1(1−e−bm1⋅(eJUTij)b0⋅(AGEi+ADj))BIOij=ak2(1−e−bm2⋅(AGEi+ADj))ck2
下载: 导出CSV
表 5 12种生物量生长模型的拟合效果评价指标
Table 5 Evaluation index of fitting effect of twelve biomass growth models
模型 Model R2 SEE/kg MPE/% TRE/% 1 0.960 8 30.073 1 1.103 2 −0.350 3 2 0.954 6 32.366 4 1.187 3 −0.532 2 3 0.960 0 30.391 2 1.114 8 −0.404 5 4 0.962 4 29.463 8 1.080 8 −0.237 8 5 0.960 3 30.241 9 1.109 4 −0.260 5 6 0.949 7 34.049 6 1.249 0 −0.001 0 7 0.967 8 27.013 2 0.990 9 −0.441 6 8 0.966 0 27.756 6 1.018 2 −0.306 3 9 0.967 8 27.011 1 0.990 8 −0.446 4 10 0.970 1 26.049 0 0.955 5 −0.222 3 11 0.969 3 26.402 5 0.968 5 −0.222 4 12 0.970 1 26.046 3 0.955 4 −0.228 2
下载: 导出CSV
表 6 基于参数b分级的联合模型立地质量等级哑变量参数
Table 6 Site quality grade dummy variable parameters of joint model based on parameter b classification
模型 Model 变量 Variable 立地质量等级Site quality grade 1 2 3 4 5 6 7 10 胸径 DBH 0.007 4 0.012 4 0.017 5 0.024 8 0.035 6 0.045 7 0.031 8 生物量 Biomass 0.007 4 0.012 4 0.017 4 0.024 2 0.034 3 0.044 8 0.027 8 11 胸径 DBH 0.007 3 0.012 1 0.017 3 0.024 5 0.035 6 0.044 9 0.030 6 生物量 Biomass 0.007 2 0.012 0 0.017 2 0.023 7 0.034 1 0.043 8 0.026 6 12 胸径 DBH 0.007 5 0.012 4 0.017 6 0.024 9 0.035 6 0.045 8 0.031 8 生物量 Biomass 0.007 4 0.012 4 0.017 4 0.024 3 0.034 3 0.044 8 0.027 9
下载: 导出CSV
表 7 基于参数b分级的联合模型的树种哑变量参数和竞争指数参数
Table 7 Dummy variable parameters of tree species and competition index parameters of joint model based on parameter b classification
树种
Tree species变量
Variable模型 Model 10 11 12 a b0/c a b0/c a b0/c 栎类
Quercus spp.胸径 DBH 46.11 46.47 −0.080 4 46.20 −0.084 6 生物量 Biomass 1 940.67 2.392 4 1 997.97 2.412 2 1 940.58 2.400 6 楠木
Phoebe zhennan胸径 DBH 44.41 44.77 −0.080 4 44.49 −0.084 6 生物量 Biomass 1 061.14 2.031 9 1 073.96 2.032 5 1 061.88 2.039 5 木荷
Schima superba胸径 DBH 46.53 46.65 −0.080 4 46.54 −0.084 6 生物量 Biomass 1 427.33 2.235 8 1 458.30 2.260 0 1 428.33 2.245 2 其他硬阔类
Other hard broadleaf胸径 DBH 48.71 48.19 −0.080 4 48.80 −0.084 6 生物量 Biomass 1 826.08 2.277 7 1 781.28 2.281 0 1 826.00 2.286 6 相思
Acacia confusa胸径 DBH 41.40 41.94 −0.080 4 41.69 −0.084 6 生物量 Biomass 1 115.52 2.461 7 1 139.54 2.466 1 1 117.12 2.468 9 其他软阔类
Other soft broadleaf胸径 DBH 42.74 43.70 −0.080 4 42.82 −0.084 6 生物量 Biomass 1 152.07 2.296 8 1 196.78 2.291 0 1 153.38 2.306 4
下载: 导出CSV
-
[1] Somogyi Z, Cienciala E, Mkip R, et al. Indirect methods of large-scale forest biomass estimation[J]. European Journal of Forest Research, 2007, 126(2): 197−207. doi: 10.1007/s10342-006-0125-7
[2] 罗云建, 张小全, 王效科, 等. 森林生物量的估算方法及其研究进展[J]. 林业科学, 2009, 45(8): 129−134. doi: 10.3321/j.issn:1001-7488.2009.08.023 Luo Y J, Zhang X Q, Wang X K, et al. Forest biomass estimation methods and their prospects[J]. Scientia Silvae Sinicae, 2009, 45(8): 129−134. doi: 10.3321/j.issn:1001-7488.2009.08.023
[3] 林开淼. 亚热带常绿阔叶林生物量模型及其分析[J]. 中南林业科技大学学报, 2017, 37(11): 115−120, 126. Lin K M. Research and analysis on biomass allometric equations of subtropical broad-leaved forest[J]. Journal of Central South University of Forestry & Technology, 2017, 37(11): 115−120, 126.
[4] Zianis D, Muukkonen P, Makipaa R, et al. Biomass and stem volume equations for tree species in Europe[J]. Silva Fennica Monographs, 2005, 4: 63.
[5] Andrzej M J, Dyderski M K, Gsikiewicz K, et al. Tree- and stand-level biomass estimation in a Larix decidua Mill. chronosequence[J]. Forests, 2018, 9(10): 587. doi: 10.3390/f9100587
[6] 曾伟生, 孙乡楠, 王六如, 等. 东北林区10种主要森林类型的蓄积量、生物量和碳储量模型研建[J]. 北京林业大学学报, 2021, 43(3): 1−8. doi: 10.12171/j.1000-1522.20200058 Zeng W S, Sun X N, Wang L R, et al. Developing stand volume, biomass and carbon stock models for ten major forest types in forest region of northeastern China[J]. Journal of Beijing Forestry University, 2021, 43(3): 1−8. doi: 10.12171/j.1000-1522.20200058
[7] Wang C K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology and Management, 2006, 222(1−3): 9−16. doi: 10.1016/j.foreco.2005.10.074
[8] Brown S, Lugo A E. Biomass of tropical forests: a new estimate based on forest volumes[J]. Science, 1984, 223: 1290−1293. doi: 10.1126/science.223.4642.1290
[9] Fang J Y, Chen A P, Peng C H, et al. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 2001, 292: 2320−2322. doi: 10.1126/science.1058629
[10] Fang J Y, Wang G G, Liu G H, et al. Forest biomass of China: an estimate based on the biomass-volume relationship[J]. Ecological Applications, 1998, 8(4): 1084−1091.
[11] Sánchez-González M, Tomé M, Montero G. Modelling height and diameter growth of dominant cork oak trees in Spain[J]. Annals of Forest Science, 2005, 62(7): 633−643. doi: 10.1051/forest:2005065
[12] 刘帅, 李建军, 卿东升, 等. 气候敏感的青冈栎单木胸径生长模型[J]. 林业科学, 2021, 57(1): 95−104. doi: 10.11707/j.1001-7488.20210110 Liu S, Li J J, Qing D S, et al. A climate-sensitive individual-tree DBH growth model for Cyclobalanopsis glauca[J]. Scientia Silvae Sinicae, 2021, 57(1): 95−104. doi: 10.11707/j.1001-7488.20210110
[13] 龙时胜, 曾思齐, 甘世书, 等. 基于林木多期直径测定数据的异龄林年龄估计方法[J]. 中南林业科技大学学报, 2018, 38(9): 1−8. Long S S, Zeng S Q, Gan S S, et al. Age estimation method of uneven-aged forest based on data of multistage diameter measurement[J]. Journal of Central South University of Forestry & Technology, 2018, 38(9): 1−8.
[14] 雷相东, 李希菲. 混交林生长模型研究进展[J]. 北京林业大学学报, 2003, 25(3): 105−110. doi: 10.3321/j.issn:1000-1522.2003.03.022 Lei X D, Li X F. A review on growth models of mixed forest[J]. Journal of Beijing Forestry University, 2003, 25(3): 105−110. doi: 10.3321/j.issn:1000-1522.2003.03.022
[15] 国红, 雷渊才, 郎璞玫. 年龄无关的生长模型研究: 以落叶松平均高为例[J]. 林业科学研究, 2020, 33(5): 129−136. Guo H, Lei Y C, Lang P M. Study on age-independent tree model: taking the average height of Larix gmelinii as an example[J]. Forest Research, 2020, 33(5): 129−136.
[16] 葛宏立, 项小强, 何时珍, 等. 年龄隐含的生长模型在森林资源连续清查中的应用[J]. 林业科学研究, 1997, 10(4): 81−85. Ge H L, Xiang X Q, He S Z, et al. Application of the age implicit growth model to continuous forest inventory[J]. Forest Research, 1997, 10(4): 81−85.
[17] 曹磊. 基于多期保留木实测胸径估计吉林省蒙古栎天然林年龄[D]. 北京: 中国林业科学研究院, 2020. Cao L. Estimating stand age of Quercus mongolica natural forest in Jilin based on diameter data of periodical measurements[D]. Beijing: Chinese Academy of Forestry, 2020.
[18] 龙时胜, 曾思齐, 甘世书, 等. 基于林木多期直径测定数据的异龄林年龄估计方法Ⅱ[J]. 中南林业科技大学学报, 2019, 39(6): 23−29, 59. Long S S, Zeng S Q, Gan S S, et al. Age estimation method Ⅱ of uneven-aged forest based on the data of multistage diameter measurement[J]. Journal of Central South University of Forestry & Technology, 2019, 39(6): 23−29, 59.
[19] Gargaglione V, Peri P L, Rubio G. Allometric relations for biomass partitioning of Nothofagus antarctica trees of different crown classes over a site quality gradient[J]. Forest Ecology & Management, 2010, 259(6): 1118−1126.
[20] Peri P L, Gargaglione V, Pastur G M, et al. Carbon accumulation along a stand development sequence of Nothofagus antarctica forests across a gradient in site quality in southern Patagonia[J]. Forest Ecology & Management, 2010, 260(2): 229−237.
[21] 孟宪宇. 测树学[M]. 3版. 北京: 中国林业出版社, 2006. Meng X Y. Forest measurement[M]. 3rd ed. Beijing: China Forestry Publishing House, 2006.
[22] Li H K, Zhao P X. Improving the accuracy of tree-level aboveground biomass equations with height classification at a large regional scale[J]. Forest Ecology & Management, 2013, 289: 153−163.
[23] 赵菡, 雷渊才, 符利勇. 江西省不同立地等级的马尾松林生物量估计和不确定性度量[J]. 林业科学, 2017, 53(8): 81−93. doi: 10.11707/j.1001-7488.20170810 Zhao H, Lei Y C, Fu L Y. Biomass and uncertainty estimates of Pinus massoniana forest for different site classes in Jiangxi Province[J]. Scientia Silvae Sinicae, 2017, 53(8): 81−93. doi: 10.11707/j.1001-7488.20170810
[24] 雷相东, 符利勇, 李海奎, 等. 基于林分潜在生长量的立地质量评价方法与应用[J]. 林业科学, 2018, 54(12): 116−126. doi: 10.11707/j.1001-7488.20181213 Lei X D, Fu L Y, Li H K, et al. Methodology and applications of site quality assessment based on potential mean annual increment[J]. Scientia Silvae Sinicae, 2018, 54(12): 116−126. doi: 10.11707/j.1001-7488.20181213
[25] 薛春泉, 徐期瑚, 林丽平, 等. 基于异速生长和理论生长方程的广东省木荷生物量动态预测[J]. 林业科学, 2019, 55(7): 86−94. doi: 10.11707/j.1001-7488.20190709 Xue C Q, Xu Q H, Lin L P, et al. Biomass dynamic predicting for Schima superba in Guangdong based on allometric and theoretical growth equation[J]. Scientia Silvae Sinicae, 2019, 55(7): 86−94. doi: 10.11707/j.1001-7488.20190709
[26] 曹磊, 刘晓彤, 李海奎, 等. 广东省常绿阔叶林生物量生长模型[J]. 林业科学研究, 2020, 33(5): 61−67. Cao L, Liu X T, Li H K, et al. Biomass growth models for evergreen broad-leaved forests in Guangdong[J]. Forest Research, 2020, 33(5): 61−67.
[27] 李巍, 王传宽, 张全智. 林木分化对兴安落叶松异速生长方程和生物量分配的影响[J]. 生态学报, 2015, 35(6): 1679−1687. Li W, Wang C K, Zhang Q Z. Differentiation of stand individuals impacts allometry and biomass allocation of Larix gmelinii trees[J]. Acta Ecologica Sinica, 2015, 35(6): 1679−1687.
[28] 臧颢, 刘洪生, 黄锦程, 等. 竞争和气候及其交互作用对杉木人工林胸径生长的影响[J]. 林业科学, 2021, 57(3): 39−50. Zang H, Liu H S, Huang J C, et al. Effects of competition, climate factors and their interactions on diameter growth for Chinese fir plantations[J]. Scientia Silvae Sinicae, 2021, 57(3): 39−50.
[29] 董利虎, 李凤日, 贾炜玮. 林木竞争对红松人工林立木生物量影响及模型研究[J]. 北京林业大学学报, 2013, 35(6): 15−22. Dong L H, Li F R, Jia W W. Effects of tree competition on biomass and biomass models of Pinus koraiensis plantation[J]. Journal of Beijing Forestry University, 2013, 35(6): 15−22.
[30] 国家林业局. 立木生物量模型及碳计量参数−栎树 (LY/T 2658—2016)[S]. 北京: 中国标准出版社, 2016. State Forestry Administration. Tree biomass models and related parameters to carbon accounting for Quercus (LY/T 2658−2016)[S]. Beijing: China Standard Press, 2016.
[31] 国家林业局. 立木生物量模型及碳计量参数−木荷 (LY/T 2660—2016)[S]. 北京: 中国标准出版社, 2016. State Forestry Administration. Tree biomass models and related parameters to carbon accounting for Schima superba (LY/T 2660−2016)[S]. Beijing: China Standard Press, 2016.
[32] 李海奎, 雷渊才. 中国森林植被生物量和碳储量评估[M]. 北京: 中国林业出版社, 2010. Li H K, Lei Y C. Estimation and evaluation of forest biomass carbon storage in China[M]. Beijing: China Forestry Publishing House, 2010.
[33] 张少昂, 王冬梅. Richards方程的分析和一种新的树木理论生长方程[J]. 北京林业大学学报, 1992, 14(3): 99−105. Zhang S A, Wang D M. New theoretical growth model based on analysis of Richards’s equation[J]. Journal of Beijing Forestry University, 1992, 14(3): 99−105.
[34] 魏晓慧, 孙玉军, 马炜. 基于Richards方程的杉木树高生长模型[J]. 浙江农林大学学报, 2012, 29(5): 661−666. doi: 10.11833/j.issn.2095-0756.2012.05.004 Wei X H, Sun Y J, Ma W. A height growth model for Cunninghamia lanceolata based on Richards’s equation[J]. Journal of Zhejiang A&F University, 2012, 29(5): 661−666. doi: 10.11833/j.issn.2095-0756.2012.05.004
[35] 惠刚盈, 胡艳波, 赵中华, 等. 基于交角的林木竞争指数[J]. 林业科学, 2013, 49(6): 68−73. doi: 10.11707/j.1001-7488.20130610 Hui G Y, Hu Y B, Zhao Z H, et al. A forest competition index based on intersection angle[J]. Scientia Silvae Sinicae, 2013, 49(6): 68−73. doi: 10.11707/j.1001-7488.20130610
[36] Wensel L C, Meerschaert W J, Biging G S. Tree height and diameter growth models for northern California conifers[J]. Hilgardia A Journal of Agricultural Science, 1987, 55(8): 1−20.
[37] Pretzsch H, Biber P. Size-symmetric versus size-asymmetric competition and growth partitioning among trees in forest stands along an ecological gradient in central Europe[J]. Canadian Journal of Forest Research, 2010, 40(2): 370−384. doi: 10.1139/X09-195
[38] Daniels R F, Burkhart H E, Clason T R. A comparison of competition measures for predicting growth of loblolly pine trees[J]. Canadian Journal of Forest Research, 1986, 16(6): 1230−1237. doi: 10.1139/x86-218
[39] Kuehne C, Weiskittel A R, Waskiewicz J. Comparing performance of contrasting distance-independent and distance-dependent competition metrics in predicting individual tree diameter increment and survival within structurally-heterogeneous, mixed-species forests of northeastern United States[J]. Forest Ecology and Management, 2019, 433: 205−216. doi: 10.1016/j.foreco.2018.11.002
[40] 符利勇, 雷渊才, 曾伟生. 几种相容性生物量模型及估计方法的比较[J]. 林业科学, 2014, 50(6): 42−54. Fu L Y, Lei Y C, Zeng W S. Comparison of several compatible biomass models and estimation approaches[J]. Scientia Silvae Sinicae, 2014, 50(6): 42−54.
[41] 何静, 朱光玉, 张学余, 等. 基于立地与密度效应的湖南栎类天然林平均木胸径生长模型[J]. 中南林业科技大学学报, 2021, 41(10): 75−82. He J, Zhu G Y, Zhang X Y, et al. A growth model of average tree diameter at breast height of Quercus natural forests in Hunan based on site and density effects[J]. Journal of Central South University of Forestry & Technology, 2021, 41(10): 75−82.
-
期刊类型引用(11)
1. 秦孝天,郭梦鸽,秦少华,陈瑞丹. 梅花新品种‘治章骨红重翠’跨品种群特性机制探究. 生物工程学报. 2024(01): 239-251 .  百度学术
百度学术
2. 周成宇,武怀燕,圣倩倩,曹福亮,祝遵凌. 33个观赏文冠果品系花瓣色彩的动态变化特征分析. 西部林业科学. 2023(05): 84-94 . 百度学术
3. 苏江硕,贾棣文,王思悦,张飞,蒋甲福,陈素梅,房伟民,陈发棣. 中国菊花遗传育种60年回顾与展望. 园艺学报. 2022(10): 2143-2162 . 百度学术
4. 付瀚森,张亚雯,赵阳阳,罗婷婷,邓慧杰,孟晨伟,王彩云. 菊花‘绿叮当’与毛华菊杂交后代花部性状杂种优势与混合遗传分析. 园艺学报. 2021(01): 96-106 . 百度学术
5. 周琦,赵峰,张慧会,祝遵凌. 香水莲花色素成分及含量的初步研究. 黑龙江农业科学. 2021(04): 72-78 . 百度学术
6. 侯瑞丽,武倩,闫星蓉,张芸香,郭晋平. 观赏型文冠果新品种花期颜色特征及其表型稳定性研究. 西北农业学报. 2021(01): 143-151 . 百度学术
7. 吴芳芳,原鑫,苏少文,贺丹,刘艺平,孔德政. 荷花品种的花器官表型性状及花色多样性分析. 河南农业大学学报. 2020(01): 24-29+37 . 百度学术
8. 赵晋陵,金玉,叶回春,黄文江,董莹莹,范玲玲,马慧琴,江静. 基于无人机多光谱影像的槟榔黄化病遥感监测. 农业工程学报. 2020(08): 54-61 . 百度学术
9. 丁苏芹,孙忆,李玺,唐东芹,史益敏. 小苍兰品种花色表型数量分类研究. 北方园艺. 2019(04): 85-91 . 百度学术
10. 袁培森,任守纲,翟肇裕,徐焕良. 基于半监督主动学习的菊花表型分类研究. 农业机械学报. 2018(09): 27-34 . 百度学术
11. 刘海英,高远,邢晨涛,甄俊琦,陆顺丽,王玉芝. 花青素苷提取专用菊种质及适宜采收期的筛选. 河南农业科学. 2018(09): 120-125 . 百度学术
其他类型引用(14)








计量
- 文章访问数: 876
- HTML全文浏览量: 258
- PDF下载量: 109
- 被引次数: 25
