
Properties and mechanism of poplar wood modified by melamine-urea-glucose (MUG) biomass resin and sodium silicate compound
-
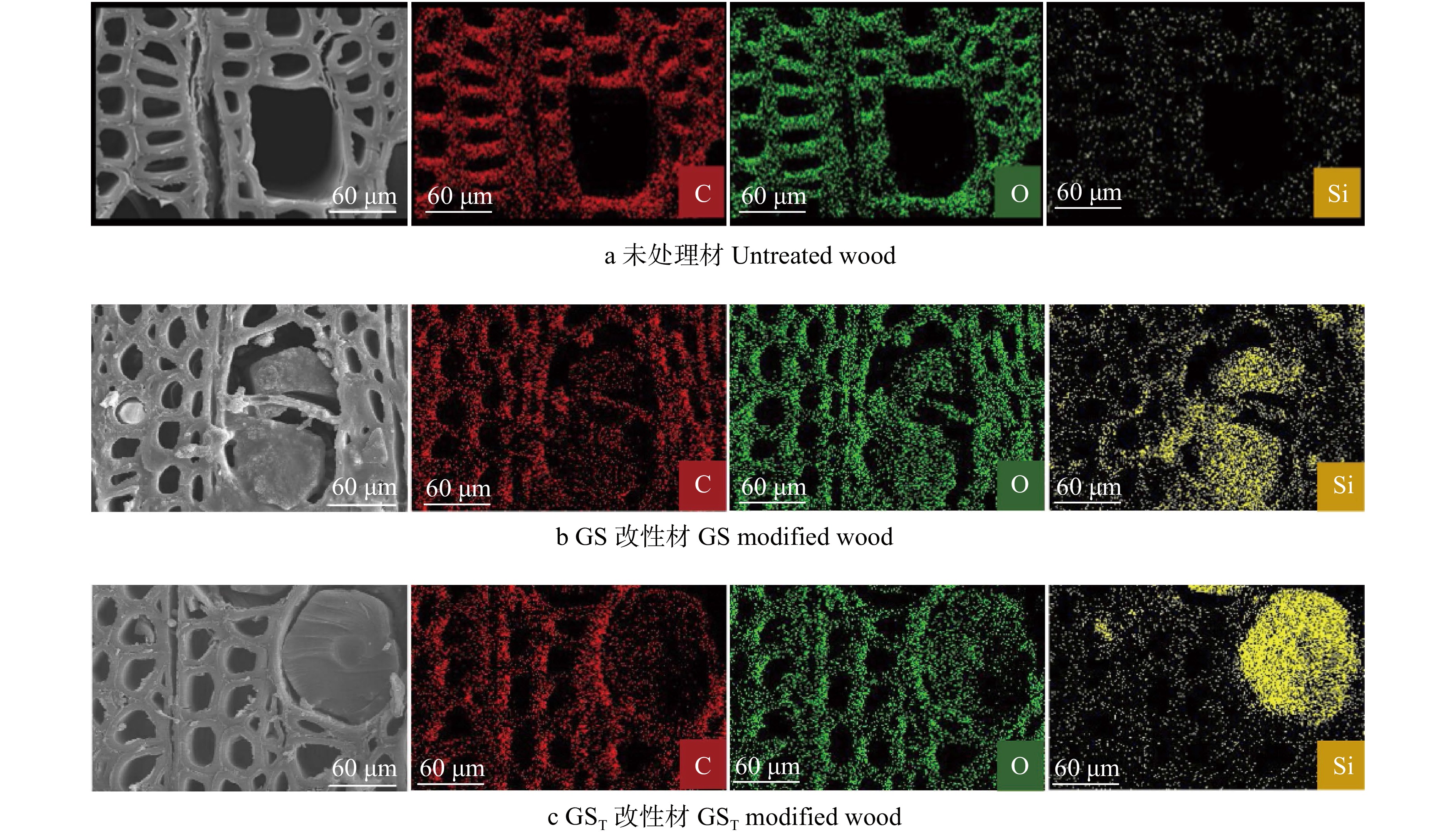
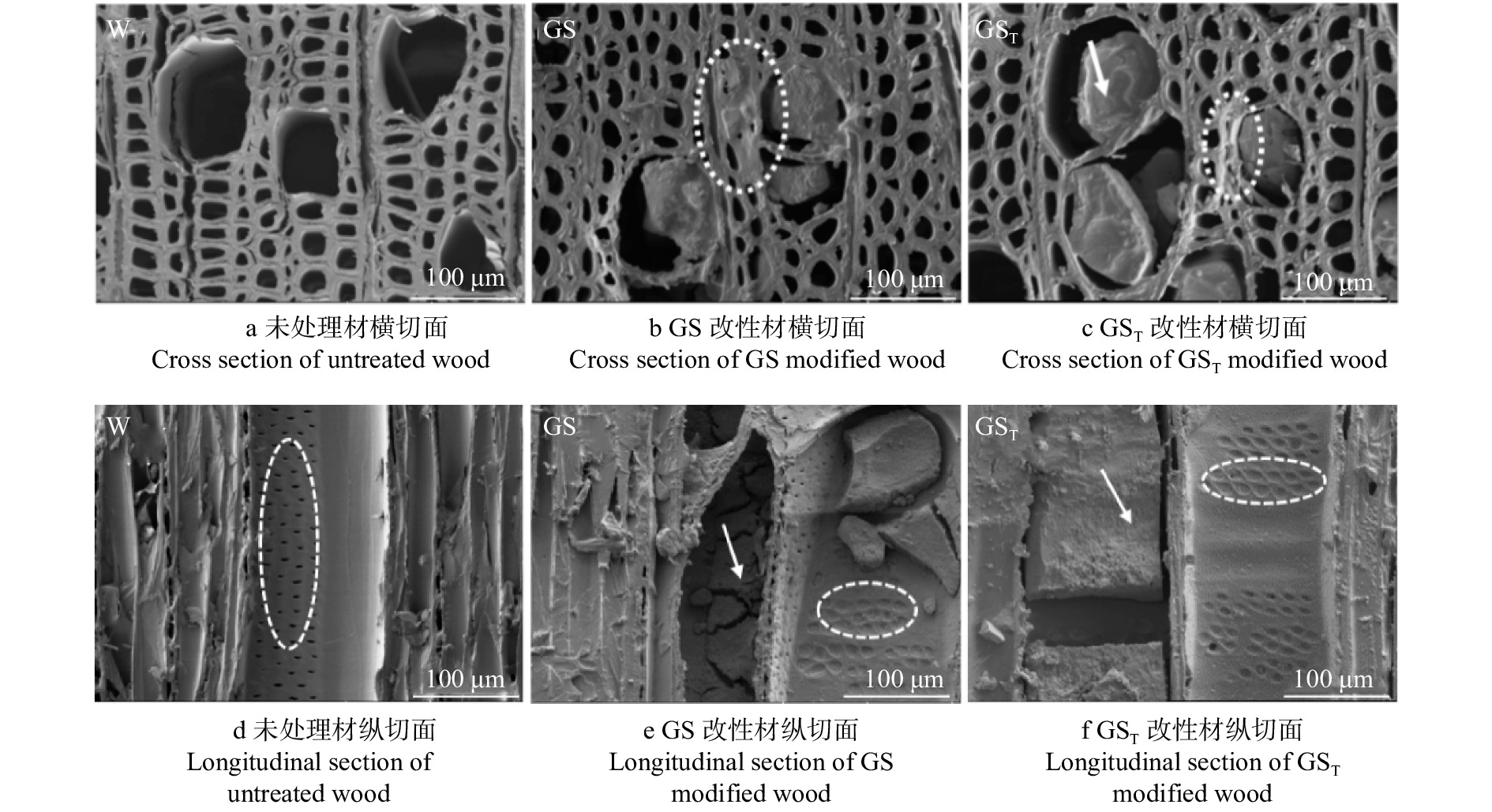
摘要:目的 研究改性材的微观形貌、化学结构和元素成分等变化,探讨三聚氰胺–尿素–葡萄糖(MUG)生物质树脂复合改性剂对杨木的改性作用机理,旨在为MUG复合改性剂的应用提供依据,促进木材的绿色改性。方法 通过将有机硅烷等疏水基团,引入MUG生物质树脂与硅酸钠的复配溶液中,制备硅烷杂化MUG树脂/硅酸钠复合改性剂(GST),对杨木进行真空加压浸渍处理,测试改性杨木的物理力学性能,采用扫描电镜–X射线能谱仪(SEM-EDX)、傅里叶变换红外光谱仪(FTIR)、X射线光电子能谱仪(XPS)和X射线衍射仪(XRD)等,表征改性材的微观形貌、化学结构、元素成分和结晶度,利用微型量热仪(MCC)测试其燃烧性能和热解特性。结果 SEM-EDX分析表明:GST改性剂渗透性好,能有效渗入木材细胞腔和细胞壁中;改性材的C、O、Si元素无规分布于木材细胞腔、细胞壁、细胞间隙等处,导管沉积最为明显;改性剂对木材孔隙的填充以及对纤维素非结晶区的充胀,有效提高了木材的尺寸稳定性和力学性能。FTIR分析表明:GST改性材中半纤维素等多糖与改性剂发生了交联反应,减少了C=O、—OH等吸水性基团。XPS分析表明:GST改性材的C1最多,C3最少,木材的多糖类物质、木质素醇羟基、酚羟基以及羰基等活性基团与改性剂发生反应,减少了羰基等活性基团,增加了C—H、C—C结构含量。XRD分析表明:GST改性材衍射峰无明显变化,相对结晶度增大,说明改性剂进入纤维素非结晶区使其分子排列更加有序。MCC分析表明:GST改性材的热释放能力、热释放速率峰值和总热释放量分别下降了65.7%、66.2%和6.2%,800 ℃残炭率提高了122.6%,热释放强度大大降低,火灾危险性减小。结论 GST复合改性剂可有效渗入杨木内部,与木材中半纤维素等多糖发生交联反应,减少糖基等活性基团,使非结晶区排列更为有序,从而提升改性杨木的物理力学性能。
-
关键词:
- 杨木 /
- 三聚氰胺–尿素–葡萄糖树脂 /
- 硅酸钠 /
- 有机杂化 /
- 改性机理
Abstract:Objective In order to provide basis for the application of melamine-urea-glucose (MUG) composite modifier and promote wood green modification, the changes of micro morphology, chemical structure and element composition of modified wood were studied, and the modification mechanism of biomass resin compound modifier on wood was discussed.Method By introducing hydrophobic groups such as organosilane into the compound solution of MUG biomass resin and sodium silicate, the silane hybrid modifier (GST) was prepared. Then the poplar wood was modified with it by vacuum pressure impregnating treatment. The physical and mechanical properties of modified wood were tested. Its micro-morphology, chemical structure, element composition and crystallinity were characterized by scanning electron microscopy-energy dispersive X-ray spectrometer (SEM-EDX), Fourier transform infrared spectrometer (FTIR), X-ray photoelectron spectroscopy (XPS) and X-ray diffractometer (XRD). The combustion performance and pyrolysis characteristics were tested by micro calorimeter (MCC).Result SEM-EDX analysis showed that GST modifier had good permeability and can effectively penetrate into wood cell cavity and cell wall; the C, O and Si element of modified wood were irregularly distributed in wood cell cavity, cell wall and cell gap, and the modifier was most deposited in wood vessels. Due to the effective filling of wood pores and the swelling of cellulose amorphous zone, wood dimensional stability and mechanical properties were improved. FTIR analysis showed that hemicellulose and other polysaccharides in GST modified wood had a cross-linking reaction with the modifier, reducing C=O, —OH and other hydroscopic groups. XPS analysis found that the C1 of GST modified wood was the most and its C3 was the least. During the modification process, the active groups such as polysaccharides, lignin alcohol hydroxyl, phenol hydroxyl and carbonyl reacted with the modifier, reducing the active groups and increasing the contents of C—H and C—C structure. XRD analysis showed that the diffraction peaks of the GST modified wood had no obvious change, its relative crystallinity increased, indicating that the modifier entered the amorphous region of cellulose to make its molecular arrangement more orderly. MCC analysis showed that the heat release capacity, peak heat release rate and total heat release of GST modified wood decreased by 65.7%, 66.2% and 6.2%, respectively, the residual carbon rate at 800 ℃ increased by 122.6%, its heat release intensity was greatly reduced and its fire risk was reduced.Conclusion GST compound modifier can effectively penetrate into poplar wood, cross-linked with its hemicellulose and other polysaccharides, reduce active groups such as sugar groups, and make the arrangement of amorphous regions more orderly, so as to improve its physical and mechanical properties.-
Keywords:
- poplar wood /
- melamine-urea-glucose /
- sodium silicate /
- organic hybrid /
- modification mechanism
-
毛白杨(Populus tomentosa)是中国独有的乡土树种,具有速生、干直、抗性好、材质优良和适应性强等优点,是我国重要的工业用材和纸浆用材[1-2]。然而,毛白杨属于轻度耐盐树种,它在NaCl含量超过0.3%的土壤中不能正常生长,这也是影响毛白杨在中国大面积推广的主要原因之一[3],培育耐盐毛白杨新品种对于充分发挥其生态效益与经济价值具有重要的意义。与传统育种相比较,基因工程育种具有高效性和专一性,能够加快育种进程,提高育种效率[4]。目前,进入田间试验阶段的转基因杨树多达10例,进入商品化应用阶段的转基因杨树有2例[5]。杨树基因工程育种发展迅猛的同时,其潜在生态风险性也应该予以相应的重视。
转基因林木生物安全性主要包括外源基因的稳定性、基因流,转基因林木对非靶标物种、生物多样性和生态系统结构的影响等[5-6]。基因流又称基因转移、基因逃逸,是转基因生物安全性评价的重要内容之一,由于其可能导致严重的生物安全性问题引起了全世界的广泛关注[7]。基因流具有方向性,包括垂直转移和水平转移。花粉传播和种子扩散是转基因植物外源基因发生垂直转移的主要途径[8],特别是虫媒传粉的植物,其花粉传播距离可达数公里以上[9],而培育雄性不育的转基因植物是阻止该途径发生的有效方式。基因水平转移是指基因不通过有性杂交在生物体之间转移的过程,细菌间遗传物质的转化、转导、结合,以及土壤农杆菌利用其自身的Ti 质粒将遗传物质整合到植物细胞的染色体上都是证明基因水平转移的最有力的证据[10]。转基因植物的外源DNA通过根际分泌物、残体和花粉等方式不断进入到土壤环境中,土壤中丰富的DNA酶能够将土壤中残留的各种生物体的DNA降解[11]。然而,在一定条件下,土壤对DNA具有一定的吸附能力,被吸附的DNA能够不被酶降解而且仍然具有转化能力,这些受保护的DNA可能在土壤中不被检测出来[12]。Zhu等[13]应用实时定量PCR对连续种植3年转苏云金芽孢杆菌(Bt)基因玉米的大田土壤进行检测,结果表明:在连续种植转基因玉米的土壤中,其外源基因一年四季都能够被检测到,并且转基因玉米中所携带的选择标记基因(NPTII)存在向具有转化能力的假单胞菌细胞水平转移的风险。目前,对于转基因植物外源基因在土壤中残留的周期和水平转移等研究还存在诸多不一致的结论,且研究多集中在转基因作物方面,而对转基因林木的生态安全性研究相对较少,尤其是大田种植多年的转基因树木研究鲜有报道。本研究对大田种植13年的转基因杨树的生物安全性进行监测,探究其基因流发生的可能性,为转AhDREB1基因三倍体毛白杨环境释放和生产性试验提供依据,也为促进转基因杨树商品化提供理论依据。
1. 材料与方法
1.1 材 料
AhDREB1 基因是从山菠菜(Prunella asiatical)的cDNA中克隆得到的一个编码DREB类转录因子基因,异源转化三倍体毛白杨,以提高受体植株耐盐能力。试验地位于山东省东营市垦利县(36°56′N,118°17′E)。年均气温12.1 ℃,年均降水量556.1 mm,县域尚未开发的盐碱地4.47万 hm2。2005 年春将转AhDREB1 基因三倍体毛白杨株系T-46种植于大田中,试验林位于黄河入海口处,土壤为海滨盐碱土,转基因毛白杨与非转基因毛白杨混交,间距5 m × 5 m,并与刺槐(Robinia pseudoacacia)、白蜡(Fraxinus excelsior)等混交。
分别于2016年2月、2018年3月从山东省东营市垦利县试验地采集转基因三倍体毛白杨雄花枝和非转基因三倍体毛白杨(受体)雄花枝。从山东省聊城市冠县杨树基地采集对照毛新杨(Populus tomentosa × P. bolleana)、银腺杨(P. alba × P. glandulosa)的雌花枝和雄花枝,用于转基因杨树花粉活力检测和室内控制杂交试验。
1.2 研究方法
1.2.1 转基因杨树花粉活力检测、杂交试验方法
在温室中对采集到的杨树花枝进行切枝水培,待雄花枝上的花粉成熟后,收集花粉,一部分花粉用于杨树杂交试验,另一部分干燥后于− 20 ℃保存用于花粉活力检测试验。2016年杂交组合:毛新杨♀ × 转基因三倍体毛白杨♂,毛新杨♀ × 非转基因三倍体毛白杨♂,毛新杨♀ × 毛新杨♂;2018年杂交组合:银腺杨♀ × 转基因三倍体毛白杨♂,银腺杨♀ × 非转基因三倍体毛白杨♂,银腺杨♀ × 银腺杨♂。采用花粉管体外萌发法、TTC染色法和MTT染色法3种方法进行花粉活力检测,试验所需试剂均为国产分析纯试剂。花粉管体外萌发法培养基的配方为:0.7%的琼脂 + 300 mg/L的CaCl2 + 20%的蔗糖 + 100 mg/L的硼酸,pH值5.8[14-15]。显微镜观察时随机选择7个视野,每个视野不少于50粒花粉,记录每个视野内花粉的萌发数量。计算花粉萌发率时,7个数值去掉一个最大值和一个最小值,其余5个视野平均值作为花粉萌发率(%),萌发率(%) = (萌发的花粉数/花粉总数)× 100%[16]。TTC染色法检测花粉活力:吸取少量0.2%TTC染色液于载玻片上,用解剖针蘸取少量新鲜花粉于染色液上混匀,盖上盖玻片,在25 ~ 28 ℃的温度下放置15 ~ 20 min,显微镜下观察并统计花粉活力百分数[17]。MTT染色法检测花粉活力:采用1%的MTT染色液对新鲜花粉进行染色[18],操作方法与TTC染色法一致。
1.2.2 转基因三倍体毛白杨枯落物降解及外源基因检测
2017年在试验林开展转基因三倍体毛白杨枯落物中外源基因的检测试验,模拟其外植体在自然环境中的凋落情况,凋落于林下土壤表面、杂草表面和土壤中(图1)。分别于5月、7月、9月将转基因三倍体毛白杨外植体的叶、枝(直径0.5 cm,长5 cm)和茎(直径 0.2 cm,长3 cm)分别装入一种透气网兜中,然后再固定于转基因杨树林下杂草表面、土壤表面和土壤中,每隔一个月分别将上述3种材料带回实验室,从中提取DNA并检测是否存在外源基因。
![]() 图 1 转基因杨树枯落物外源基因检测试验Figure 1. Detection experiment of exogenous gene in transgenic poplar litter
图 1 转基因杨树枯落物外源基因检测试验Figure 1. Detection experiment of exogenous gene in transgenic poplar litter1.2.3 转基因三倍体毛白杨外源基因向林下根际土壤可培养细菌水平转移的研究
2018年3月在该试验地挑选13年生转基因三倍体毛白杨15棵,采用等边三角形取样法采集距离转基因三倍体毛白杨基部10 cm、深度20 cm的根际土壤,装入自封袋中密封保存。使用保温箱低温运回实验室,过2 mm 筛后装入自封袋,贴好标签,置于4 ℃ 保存。使用添加卡那霉素的细菌培养基对转基因三倍体毛白杨根际土壤中的微生物进行筛选培养,使用两对外源基因和选择标记基因(NPTII)的PCR特异性引物对筛选出的具卡那霉素抗性细菌的DNA进行PCR特异性检测。
细菌培养使用传统的牛肉膏蛋白胨培养基5.0 g/L牛肉膏 + 10.0 g/L蛋白胨 + 5.0 g/L NaCl + 20.0 g/L琼脂,pH = 7.4 ~ 7.6。转基因三倍体毛白杨枯落物、根际土壤和细菌基因组DNA的提取使用天根公司生产的植物基因组DNA提取试剂盒(DP305)、土壤基因组DNA提取试剂盒(DP336)和细菌基因组提取试剂盒(DP302)。外源基因的PCR扩增特异性引物:AhDREB1-1号引物(Forward:5′-TGCGTTTACGGAGGAATG-3′,Reverse:5′- CGTTTACCCAAGAACAGGAC-3′),预期片段长度198 bp;AhDREB1-2号引物(Forward:5′-AATTCTCCGATCCCATTTC-3′,Reverse:5′-AGCTGCTTTGGCTTACGA-3′),预期片段长度443 bp。NPTII选择标记基因引物(Forward:5′-GAACAAGATGGATTGCACGC-3′,Reverse:5′- GAAGAACTCGTCAAGAAGGC-3′),预期片段长度730 bp。细菌16S rDNA 通用引物(Forward:5′-ACGGGCGGTGTGTAC-3′,Reverse:5′-CCTACGGGAGGCAGCAG-3′)。
1.2.4 数据分析
采用Excel 2010 和SPSS 17. 0软件进行数据统计与分析。
2. 结果与分析
2.1 转基因三倍体毛白杨成年树木花粉可育性分析
2016年2月我们对转AhDREB1基因三倍体毛白杨的花粉进行了检测,大部分对照毛新杨花粉被染成红色,而无论是转基因三倍体毛白杨的花粉还是非转基因毛白杨(受体)都几乎没有被染上色。2018年3月,使用TTC检测和MTT检测两种方法进行花粉活力检测试验(图2、图3),试验结果表明:大部分对照银腺杨花粉被染成红色,而无论是转基因三倍体毛白杨的花粉还是非转基因三倍体毛白杨(受体)都几乎没有被染上色。试验结果与2016年的检测结果一致,这表明转基因三倍体毛白杨的花粉活力很差,其原因可能是转基因三倍体毛白杨的受体本身花粉活力较差。
![]() 图 2 2018年TTC法检测杨树花粉活力Figure 2. Poplar pollen viability tested by TTC staining in 2018
图 2 2018年TTC法检测杨树花粉活力Figure 2. Poplar pollen viability tested by TTC staining in 2018![]() 图 3 2018年MTT法检测杨树花粉活力Figure 3. Poplar pollen viability tested by MTT staining in 2018
图 3 2018年MTT法检测杨树花粉活力Figure 3. Poplar pollen viability tested by MTT staining in 2018经统计,2016年两种方法(花粉活性TTC 检测和花粉体外培养检测)鉴定下,转基因三倍体毛白杨和非转基因三倍体毛白杨花粉的活力均为0,而对照植株毛新杨的花粉活力分别为56.7%( TTC 检测)和67.4%(花粉体外培养检测)(表1)。2018年两种方法(花粉活性TTC 检测和花粉活性MTT 检测)鉴定下,转基因三倍体毛白杨和非转基因三倍体毛白杨(受体)的活力均为0,而对照植株银腺杨的花粉活力分别为48.7%(TTC 染色法)、86.4%(MTT染色法)(表2)。
表 1 2016年花粉活力测试结果Table 1. Pollen viability test results in 2016品种 Variety TTC检测 TTC test 花粉体外培养检测 Pollen culture in vitro 转基因三倍体毛白杨 Transgenic triploid P. tomentosa 0 0 非转基因三倍体毛白杨(受体) Non-transgenic triploid P. tomentosa (receptor) 0 0 毛新杨 P. tomentosa × P. bolleana 56.7% ± 6.5% 67.4% ± 8.7% 表 2 2018年花粉活力测试结果Table 2. Pollen viability test results in 2018品种 Variety TTC检测 TTC test MTT 检测 MTT test 转基因三倍体毛白杨 Transgenic triploid P. tomentosa 0 0 非转基因三倍体毛白杨(受体) Non-transgenic triploid P. tomentosa (receptor) 0 0 银腺杨 P. alba × P. glandulosa 48.7% ± 7.2% 86.4% ± 5.4% 2016年以毛新杨雌株为母本,以转基因三倍体毛白杨、非转基因三倍体毛白杨(受体)以及对照毛新杨雄株为父本,进行杨树杂交试验,收集种子。没能得到毛新杨与转基因三倍体毛白杨的杂交种子,也没有得到毛新杨与非转基因三倍体毛白杨(受体)的杂交种子,只得到少量对照毛新杨与毛新杨的杂交种子。2018年3月,以银腺杨为母本再次进行杨树杂交试验,仍未能得到转基因三倍体毛白杨或非转基因三倍体毛白杨(受体)做父本与银腺杨母本的杂交种子,仅能得到对照父本银腺杨与母本银腺杨的杂交种子(图4)。两年的杂交试验结果(表3、表4)与花粉活力检测结果相符合,表明转AhDREB1基因三倍体毛白杨花粉确实没有活力,花粉不具备可授性和育性。
表 3 2016年转基因三倍体毛白杨杂交试验结果Table 3. Hybridization assay results of transgenic triploid P. tomentosa in 2016亲本 Parent 授粉小花数量
Number of flowers
pollinated种子数量
Number of seeds结籽率
Seed rate/%母本 Female 父本 Male 毛新杨
P. tomentosa × P. bolleana转基因三倍体毛白杨
Transgenic triploid P. tomentosa1 585 0 0 毛新杨
P. tomentosa × P. bolleana非转基因三倍体毛白杨(受体)
Non-transgenic triploid P. tomentosa (receptor)1 376 0 0 毛新杨
P. tomentosa × P. bolleana毛新杨
P. tomentosa × P. bolleana1 104 36 3.26 表 4 2018年转基因三倍体毛白杨杂交试验结果Table 4. Hybridization assay results of transgenic triploid P. tomentosa in 2018亲本Parent 授粉小花数量
Number of flowers
pollinated种子数量
Number of seeds结籽率
Seed rate/%母本 Female 父本 Male 银腺杨
P. alba × P. glandulosa转基因三倍体毛白杨
Transgenic triploid P. tomentosa1 210 0 0 银腺杨
P. alba × P. glandulosa非转基因三倍体毛白杨(受体)
Non-transgenic triploid P. tomentosa (receptor)1 085 0 0 银腺杨
P. alba × P. glandulosa银腺杨
P. alba × P. glandulosa875 128 14.63 ![]() 图 4 毛白杨飞絮的果实、败育的种子和银腺杨的种子Figure 4. Fruit of P. tomentosa, abortive seeds and the seeds of P. alba × P. glandulosa
图 4 毛白杨飞絮的果实、败育的种子和银腺杨的种子Figure 4. Fruit of P. tomentosa, abortive seeds and the seeds of P. alba × P. glandulosa2.2 转基因三倍体毛白杨枯落物降解及外源基因检测分析
2017年5月、7月、9月对转基因三倍体毛白杨枯落物中外源基因进行检测,5月15日在试验林铺设试验,3个月后(8月30日)已经无法从这些枯落物基因组中检测到外源基因的残留;又于7月11日在试验林铺设试验,3个月后(10月15日)也未能检测到外源基因的残留;再于8月30日在试验林铺设试验,3个月后(11月29日)腐烂的材料中已无法检测到外源基因残留。结果表明:转基因杨树的枯落物(枝、茎和叶)无论是埋在土里,飘落在土壤表面,飘落在杂草表面上,在3个月后均未能检测到其外源基因(图5)。
![]() 图 5 转基因三倍体毛白杨枯落物中外源基因的检测1 ~ 3:埋在土里的转基因三倍体毛白杨枝、茎和叶;4 ~ 6:飘落在地表的转基因三倍体毛白杨枝、茎和叶;7 ~ 9:飘落在杂草上的转基因三倍体毛白杨枝、茎和叶;CK−:阴性对照,CK+:阳性对照,M: 2 000 bp DNA标记。1−3:transgenic hybrid Populus tomentosa branches, stems and leaves buried in the earth;4−6:transgenic hybrid Populus tomentosa branches, stems and leaves falt on the earth’s surface;7−9 transgenic hybrid Populus tomentosa branches, stems and leaves falt on the lawn. CK−:negative control,CK+:positive control, M:2 000 bp DNA marker.Figure 5. Detection of exogenous genes in transgenic triploid Populus tomentosa litter
图 5 转基因三倍体毛白杨枯落物中外源基因的检测1 ~ 3:埋在土里的转基因三倍体毛白杨枝、茎和叶;4 ~ 6:飘落在地表的转基因三倍体毛白杨枝、茎和叶;7 ~ 9:飘落在杂草上的转基因三倍体毛白杨枝、茎和叶;CK−:阴性对照,CK+:阳性对照,M: 2 000 bp DNA标记。1−3:transgenic hybrid Populus tomentosa branches, stems and leaves buried in the earth;4−6:transgenic hybrid Populus tomentosa branches, stems and leaves falt on the earth’s surface;7−9 transgenic hybrid Populus tomentosa branches, stems and leaves falt on the lawn. CK−:negative control,CK+:positive control, M:2 000 bp DNA marker.Figure 5. Detection of exogenous genes in transgenic triploid Populus tomentosa litter2.3 转基因三倍体毛白杨外源基因向林下根际土壤可培养细菌水平转移的研究
从转基因杨树林下根际土壤中筛选得到具卡那霉素抗性细菌单菌落44个,使用目的基因的两对特异性引物AhDREB1-1号、AhDREB1-2号和选择标记基因NPTII特异性引物对上述DNA样本进行PCR扩增检测,没有扩增出相符合的目的片段(图6)。研究结果表明:种植13年的转基因三倍体毛白杨枯落物中的外源基因暂未水平转移到根际土壤可培养细菌基因组中,该试验结果与课题组2017年的研究结果[19]相一致。
![]() 图 6 转基因杨树林下根际土壤中卡那霉素抗性细菌外源基因和选择标记基因的PCR检测1 ~ 44:转基因杨树林下根际土壤中卡那霉素抗性细菌,CK−:阴性对照,CK+:阳性对照,M:2 000 bp DNA 标记。1−44:Kanamycin resistant bacteria in the rhizosphere soil of transgenic poplar forest, CK−:negative control, CK+:positive control, M: 2 000 bp DNA marker.Figure 6. PCR detection of exogenous and selectable marker genes in kanamycin resistant bacteria in rhizospheric soils of transgenic poplar
图 6 转基因杨树林下根际土壤中卡那霉素抗性细菌外源基因和选择标记基因的PCR检测1 ~ 44:转基因杨树林下根际土壤中卡那霉素抗性细菌,CK−:阴性对照,CK+:阳性对照,M:2 000 bp DNA 标记。1−44:Kanamycin resistant bacteria in the rhizosphere soil of transgenic poplar forest, CK−:negative control, CK+:positive control, M: 2 000 bp DNA marker.Figure 6. PCR detection of exogenous and selectable marker genes in kanamycin resistant bacteria in rhizospheric soils of transgenic poplar3. 结论与讨论
国内外对于转基因植物安全性的研究主要集中在作物上,关于转基因林木的研究也大多集中在林木苗期或者幼树,对成龄大树研究不是特别多。然而,林木用材林多是在成龄期取材,生态林则会长期存在于自然环境中,因此,在转基因林木大面积种植和商品化种植前应对其进行长期系统性的风险评估[20]。此外,林木与作物几个重要特征的差异必须在风险评估中加以考虑,林木生命周期长,栽培环境复杂,管理粗放,其花粉和种子可以传播到很远的地方,转基因林木对环境的潜在风险性更大[21-23]。转基因植物的花粉漂移一直是转基因植物生态安全性评价的重要研究内容,外源基因通过花粉向非转基因品种或野生近缘物种漂移将会对生态环境造成巨大危害[24]。路兴波等[25]对转抗除草剂基因玉米外源基因向周边环境遗传漂移的距离和频率进行研究,发现转基因玉米(Zea mays L.)的外源基因可向周边玉米品种进行漂移,其最大漂移频率为45.10%,150 m处仍能检测到外源基因的漂移。此外,转基因油菜(Brassica campestris L.)、水稻(Oryza sativa L.)、棉花(Gossypium spp.)等的外源基因通过花粉漂移的研究均有报道[26-28]。本试验中我们连续两年对转基因三倍体毛白杨花粉活力和杂交结实率进行检测,两年的数据结果表明我们课题组培育的转AhDREB1基因三倍体毛白杨花粉可育性极低,转基因受体植株自身花粉活力低可能是造成转基因三倍体毛白杨杂交可育性低的原因,因此,其通过花粉向近缘物种扩散的可能性也很低。基因的水平转移也一直被认为是转基因植物潜在的安全隐患之一。有研究表明土壤微生物能够利用自由存在的DNA并将其整合到自身基因组中,尤其是一些抗生素标记基因[29-30]。Zuo 等[31]对转多个抗虫基因的741杨树进行了连续5年的生态安全性研究,5年的实地研究均未能从非转基因741杨树、转基因741杨树林下植被和土壤中检测到外源基因。Saxena等[32]对转Bt基因玉米根系分泌物中释放的抗鳞翅类毒素(Cry1Ab蛋白)进行研究,发现这些通过根系分泌物所释放的毒素会在土壤中积累,并迅速附着在土壤表面活性颗粒上(如黏土和腐殖质),并保留至少180天的杀虫活性。Gebhard等[33]研究表明:大田种植的转基因甜菜(Beta vulgaris L.)其外源DNA可以在不同类型土壤中残留数月,甚至2年。邓欣等[34]采用菌落PCR方法对分离的叶围卡那霉素抗性细菌进行NPTII基因漂移监测,在部分转基因抗虫棉的叶围细菌中检测到阳性片段,研究结果表明转基因抗虫棉中的NPTII选择标记基因可能向叶围细菌漂移。更有学者认为:基因水平转移是抗生素抗性基因传播的重要方式,是造成抗性基因环境污染日益严重的原因之一[35]。因此,在转基因林木商品化应用之前,我们有必要对其枯落物中外源基因的残留时间进行检测,以确保这些具有外源基因的枯落物在凋落后能自然分解,不会对环境造成影响。本试验应用PCR技术对转基因三倍体毛白杨枯落物中的外源基因进行检测,并从转基因三倍体毛白杨林下根际土壤中筛选出44株卡那霉素抗性细菌,对其基因组DNA进行外源基因PCR检测,结果表明:转基因三倍体毛白杨枯落物中的外源基因在自然环境中持留3个月后均未能被检测到,同时,也没有从44株卡那霉素抗性细菌基因组中扩增出相符合的目的片段。转基因枯落物中外源基因存在2 ~ 3个月的滞留周期,那么在落叶季,源源不断的枯落物从树体脱落进入到自然环境中,将会造成枯落物中的外源基因暴露时间增长。现阶段没有足够的证据证明这些“潜伏”的外源基因是否能水平转移到土壤微生物基因组中,但这可能是因为现阶段我们的检测方法以及检测技术的局限性。此外,我们通过抗性筛选培养的方式所培养的细菌在整个土壤微生物中的占比微乎其微,未来需要我们借助更为先进的方法与技术对这片转基因杨树成年林进行更为严格的检测,以评估长期存在于自然环境中的转基因林木对周围环境可能带来的潜在风险。
林木转基因安全性问题一直是从事林木遗传改良工作者关切的热点问题,存在较多争议,而国内外学者开展的林木转基因安全性研究大多为短期研究,长期系统性的研究鲜有报道。未来,需要科研工作者对该领域投入更多的研究,以科学的风险评估为依据,加快推进转基因林木商品化进程。
-
![]()
图 2 杨木及其改性材横切面SEM-EDX图
Figure 2. Cross section’s SEM-EDX images of poplar wood and its modified wood
![]()
图 5 杨木及其改性材的热释放曲线和测试数据
Figure 5. Heat release curves and MCC data of poplar wood and its modified wood
表 1 杨木及其改性材的物理力学性能
Table 1 Physical and mechanical properties of poplar wood and its modified wood
组别
Group密度 Density/
(g·cm−3)抗胀缩率 Anti-swelling efficiency (ASE)/% 抗弯弹性模量Modulus of elasticity
(MOE)/GPa抗弯强度Modulus of rapture
(MOR)/MPa顺纹抗压强度Compressive strength
parallel to grain (CS)/MPa径向 Radial 弦向 Tangential 体积 Volume W 0.325(0.01) 8.84(1.7) 76.10(7.8) 53.75(8.2) GS 0.523(0.01) 60.81(6.1) 34.20(9.1) 51.87(6.3) 13.73(2.0) 112.47(9.4) 109.11(7.6) GST 0.525(0.01) 63.71(9.3) 45.90(3.8) 57.54(4.3) 14.25(2.1) 116.02(7.4) 111.30(6.1) 注:W为未处理材,GS为MUG树脂/硅酸钠改性材,GST为硅烷改性MUG树脂/硅酸钠改性材。下同。括号内为变异系数。Notes: W is untreated wood, GS is MUG resin/sodium silicate modified wood, and GST is silane modified MUG resin/sodium silicate modified wood. The same below. The coefficient of variation is showed in brackets.  下载: 导出CSV
下载: 导出CSV
表 2 杨木及其改性材表面的元素成分和相对含量
Table 2 Surface elements and their relative contents of poplar wood and its modified wood
组别
GroupC/% O/% N/% Si/% O/C Si/C C1/% C2/% C3/% W 64.21 32.80 0.70 2.29 0.51 0.04 20.06 51.24 28.70 GS 58.78 33.73 3.30 4.19 0.57 0.07 32.84 55.89 11.27 GST 57.70 33.85 2.01 6.44 0.58 0.11 44.22 48.11 7.67
下载: 导出CSV
-
[1] 赵鹏炜, 徐国祺, 杨鸿. 纳米CuO/硅溶胶制剂处理杨木性能的研究[J]. 北京林业大学学报, 2021, 43(11): 109−117. doi: 10.12171/j.1000-1522.20210299 Zhao P W, Xu G Q, Yang H. Research on the performance of poplar wood treated by nano-CuO/silica sol formulations[J]. Journal of Beijing Forestry University, 2021, 43(11): 109−117. doi: 10.12171/j.1000-1522.20210299
[2] 刘强强, 吕文华, 石媛, 等. 复合硅改性热处理杨木的制备及性能[J]. 北京林业大学学报, 2021, 43(1): 136−143. Liu Q Q, Lü W H, Shi Y, et al. Preparation and properties of heat-treated poplar wood modified with composite silicon modifier[J]. Journal of Beijing Forestry University, 2021, 43(1): 136−143.
[3] Viswanathan T, Richardson T. Thermosetting adhesives resins from whey and whey by-products[J]. Industrial and Engineering Chemistry Product Research and Development, 1984, 23(4): 644−647. doi: 10.1021/i300016a027
[4] Liu Q, Du H, Lyu W. Physical and mechanical properties of poplar wood modified by glucose-urea-melamine resin/sodium silicate compound[J]. Forests, 2021, 12(2): 127.
[5] Wan Y Z, Luo H, He F, et al. Mechanical, moisture absorption, and biodegradation behaviours of bacterial cellulose fibre-reinforced starch biocomposites[J]. Composites Science and Technology, 2009, 69(7-8): 1212−1217. doi: 10.1016/j.compscitech.2009.02.024
[6] Lang Q, Bi Z, Pu J W. Poplar wood-methylol urea composites prepared by in situ polymerization. II. characterization of the mechanism of wood modification by methylol urea[J]. Journal of Applied Polymer Science, 2015, 132(41): 280.
[7] 付菁菁, 何春霞, 王思群. 浸渍过程对纳米纤维素/二氧化硅复合气凝胶结构与性能研究[J]. 光谱学与光谱分析, 2017, 37(7): 2019−2023. Fu J J, He C X, Wang S Q. Effect of immersion process the properties and structure of cellulose nanofibril/silica composite aerogels[J]. Spectroscope and Spectral Analysis, 2017, 37(7): 2019−2023.
[8] Tjeerdsma B, Militz H. Chemical changes in hydrothermal treated wood: FTIR analysis of combined hydrothermal and dry heat-treated wood[J]. Holz Roh-Werkst, 2005, 63: 102−111. doi: 10.1007/s00107-004-0532-8
[9] 张涛, 于建芳, 王喜明, 等. 山苍子油对人工林北京杨的改性效果[J]. 应用化工, 2020, 49(7): 1661−1665. Zhang T, Yu J F, Wang X M, et al. Effect of Litsea cubeba oil on the modification of Beijing poplar plantation[J]. Applied Chemical Industry, 2020, 49(7): 1661−1665.
[10] 李萍, 吴义强, 左迎峰, 等. XPS和FTIR分析仿生呼吸法对硅酸盐改性杉木浸渍效果的影响[J]. 光谱学与光谱分析, 2021, 41(5): 1430−1435. Li P, Wu Y Q, Zuo Y F, et al. Effect of biometic respiration method on the impact of silicone modified Chinese fir by XPS and FTIR analysis[J]. Spectroscope and Spectral Analysis, 2021, 41(5): 1430−1435.
[11] 钱曹厉. CaCl2-NaCO3内部反应沉积改性杨木的制备及性能研究[D]. 南京: 南京林业大学, 2020. Qian C L. Preparation and performance of CaCl2-NaCO3 internal reaction deposition modified poplar[D]. Nanjing: Nanjing Forestry University, 2020.
[12] 刘雪纯, 王凯伦, 甘卫星, 等. 葡萄糖三聚氰胺甲醛树脂的热性能[J]. 桂林理工大学学报, 2018, 38(3): 513−518. doi: 10.3969/j.issn.1674-9057.2018.03.021 Liu X C, Wang K L, Gan W X, et al. Thermal properties of glucose-melamine-formaldehyde resin[J]. Journal of Guilin University of Technology, 2018, 38(3): 513−518. doi: 10.3969/j.issn.1674-9057.2018.03.021
[13] Liu C, Wang S, Shi J, et al. Fabrication of superhydrophobic wood surfaces via a solution-immersion process[J]. Applied Surface Science, 2011, 258(2): 761−765. doi: 10.1016/j.apsusc.2011.08.077
[14] Jiang J, Cao J Z, Wang W. Characteristics of wood-silica composites influenced by the pH value of silica sols[J]. Holzforschung, 2018, 72(4): 311−319. doi: 10.1515/hf-2017-0126
[15] 彭尧, 王雯, 曹金珍. 蒙脱土对木粉/聚丙烯复合材料光降解及老化抑制作用[J]. 北京林业大学学报, 2018, 40(8): 116−122. Peng Y, Wang W, Cao J Z. Photodegradation and anti-weathering effects of montmorillonite on WF/PP composites[J]. Journal of Beijing Forestry University, 2018, 40(8): 116−122.
[16] 潘明珠, 梅长彤. 纳米SiO2-APP对木塑复合材料界面特性及力学性能的影响[J]. 北京林业大学学报, 2013, 35(5): 117−122. Pan M Z, Mei C T. Effects of nano SiO2-ammonium polyphosphate on the interfacial and mechanical properties of wood fiber-polyethylene composites[J]. Journal of Beijing Forestry University, 2013, 35(5): 117−122.
[17] Meng F, Yu Y, Zhang Y, et al. Surface chemical composition analysis of heat-treated bamboo[J]. Applied Surface Science, 2016, 371: 383−390. doi: 10.1016/j.apsusc.2016.03.015
[18] Miao X, Chen H, Lang Q, et al. Characterization of Ailanthus altissima veneer modified by urea-formaldehyde pre-polymer with compression drying[J]. Bioresources, 2014, 9(4): 5928−5939.
[19] 王喆, 刘君良, 孙柏玲, 等. 真空热处理人工林落叶松木材吸湿性变化机理研究[J]. 光谱学与光谱分析, 2017, 37(10): 3160−3164. Wang Z, Liu J L, Sun B L, et al. Study on mechanism of moisture absorption change of larch plantation under vacuum heat treatment[J]. Spectroscope and Spectral Analysis, 2017, 37(10): 3160−3164.
[20] Wang J, Zhang M, Chen M, et al. Catalytic effects of six inorganic compounds on pyrolysis of three kinds of biomass[J]. Thermochimica Acta, 2006, 444(1): 110−114. doi: 10.1016/j.tca.2006.02.007
[21] Nguyen T T, Nguyen T V K, Xiao Z, et al. Combustion behavior of poplar (Populus adenopoda Maxim.) and radiata pine (Pinus radiata Don.) treated with a combination of styrene-acrylic copolymer and sodium silicate[J]. Holz Als Roh Und Werkstoff, 2019, 77(3): 439−452. doi: 10.1007/s00107-019-01401-2
-
期刊类型引用(10)
1. 王蕾,郭秋菊,艾训儒,姚兰,朱江,刘西尧. 林分空间结构对天然林木本植物多样性的影响. 森林与环境学报. 2024(01): 20-27 .  百度学术
百度学术
2. 朱临渊,曹受金,颜惠芳,彭翠英,廖德志,梁军生,杨鹏华,龚雄夫,王旭军. 杉木凋落物对魔芋的生长及其生理生化影响研究. 湖南林业科技. 2024(01): 1-9 . 百度学术
3. 于佳乐,刘志明,王海英. 木醋液对4种蔬菜种子萌发的影响. 中国野生植物资源. 2024(02): 64-72 . 百度学术
4. 荆蓉,彭祚登,李云,王少明. 刺槐林下凋落物浸提液对刺槐种子萌发和胚生长的化感作用. 浙江农林大学学报. 2023(01): 97-106 . 百度学术
5. 李梦琪,赵冲,罗航,陈杭,刘博,王正宁. 不同凋落物水浸提液对杉木种子萌发和幼苗早期生长的化感作用. 江苏农业科学. 2023(07): 138-146 . 百度学术
6. 荆蓉,彭祚登,李云,王少明. 刺槐林下枯落物浸提液对自身幼苗生长的化感效应. 西北林学院学报. 2023(04): 27-33 . 百度学术
7. 罗合一,李美玄,贠民强,马印玺,赵萧汀,赵蕾,曲同宝. 四种凋落物对入侵植物火炬树种子萌发和幼苗生长的影响. 山东农业科学. 2023(10): 59-65 . 百度学术
8. 马永林,武利玉,张砡嫣,杨玉凤. 兰州主要阔叶造林树种凋落物对火炬树种子萌发的影响. 草原与草坪. 2022(03): 91-99 . 百度学术
9. 晋梦然,贾梅花,肖倩茹,刘金福,沈彩霞,施友文,何中声. 林窗凋落物化感作用对格氏栲幼苗生长的影响. 生态学报. 2022(20): 8288-8299 . 百度学术
10. 徐来仙,姚兰,周大寨,郭秋菊,朱江,邓楚,艾鑫,夏煜轩. 水杉凋落物水浸提液对其种子萌发和生长的化感作用. 广西植物. 2022(11): 1949-1958 . 百度学术
其他类型引用(4)









计量
- 文章访问数: 870
- HTML全文浏览量: 269
- PDF下载量: 139
- 被引次数: 14
