
Relationship between tree growth and tree size, competition as well as species diversity in spruce-fir natural forests
-
摘要:目的
林分内林木大小、竞争和树种多样性等多方面因素影响着林木的生长,而胸高断面积生长量通常被用来描述树木生长状态。本文利用长白山云冷杉针阔混交林主要树种的单木胸高断面积生长量建立随机森林模型,研究和量化影响树木生长的环境因素,旨在为该地区的云冷杉针阔混交林生长预估提供理论依据。
方法连续24年(1987—2010年)对总样木数为6 903株的固定样地进行数据调查,应用随机森林算法,选取单木、竞争因子、多样性和气候方面共11个调查因子,对混交林中6个主要树种建立胸高断面积生长量模型,并使用10折交叉验证法来优化超参数mtry和评估模型结果。
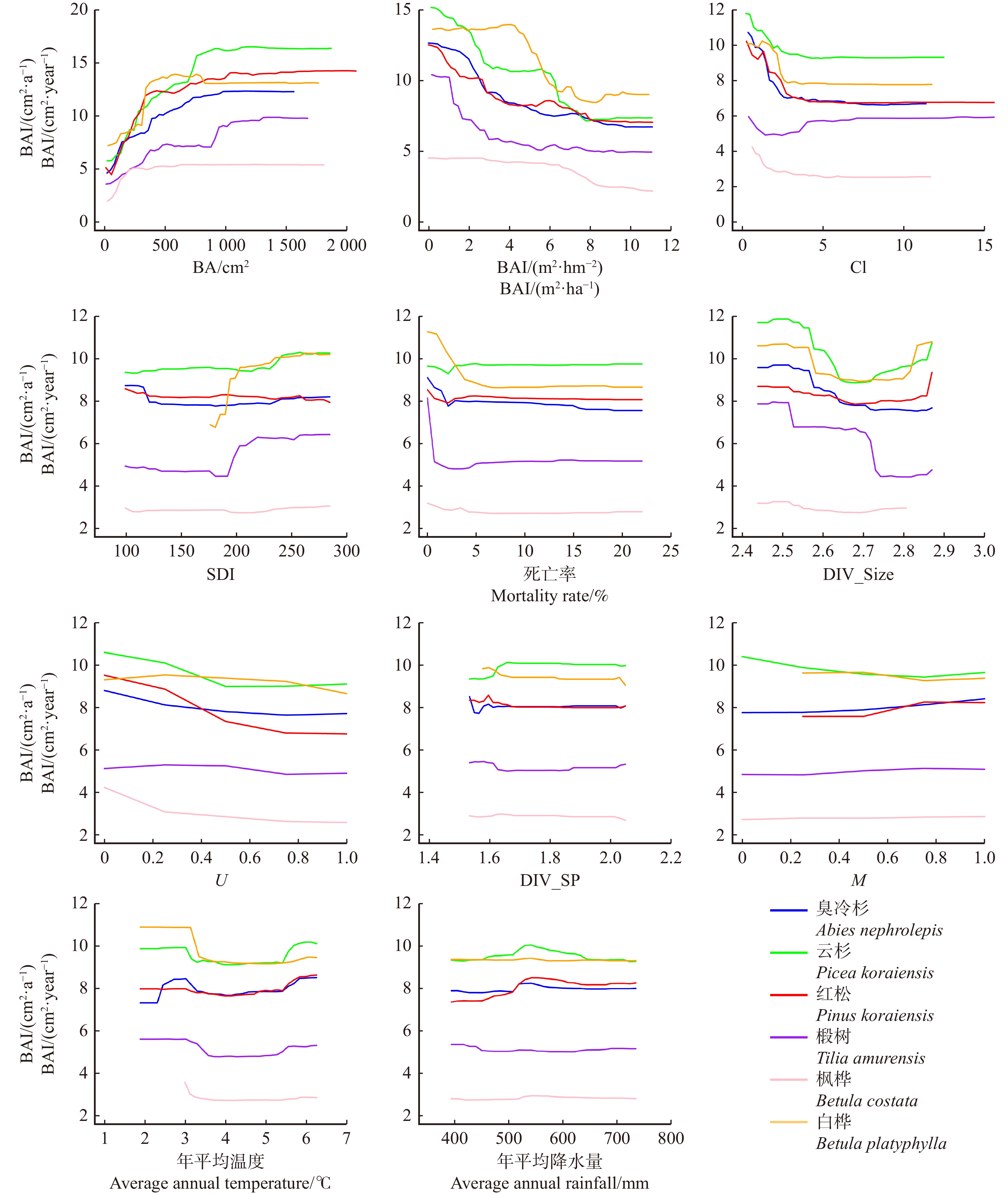
结果(1)臭冷杉、云杉、红松、椴树、枫桦、白桦6个主要树种胸高断面积生长量模型的决定系数分别为0.663、0.683、0.695、0.459、0.384和0.568。(2)单木胸高断面积是最重要的因子,对树木生长有着很强的正向影响;竞争因子和树木大小多样性是影响树木生长的主要因素,单木胸高断面积生长量随着竞争因子、树木大小多样性增加而下降。(3)树种多样性对树木生长的影响比较有限,树种多样性指数和混交度的增加会一定程度加快云杉、臭冷杉和红松的生长速度;而气候因子对树木生长的影响则相对较小。
结论树木生长在很大程度上依赖于其自身的生长潜力,在外界环境中主要受到来自林木间竞争和树木大小多样性的抑制,而树种多样性的增加也能在一定程度促进林内优势树种的生长;随机森林模型能够很好地量化和显出各变量与单木胸高断面积生长量之间复杂的关系,可以作为森林管理评价工具,为森林生长收获预估提供新的方法。
Abstract:ObjectiveTree size, competition, tree species diversity and other factors in the forest stand affect the growth of forest trees, while the individual basal area increment (BAI) is often used to describe tree growth. A random forest model was established to study the individual tree BAI of main tree species in the mixed forest of Changbai Mountain of northeastern China, study and quantify the environmental mechanisms affecting tree growth, aiming to provide a theoretical basis for growth projections.
MethodData were investigated for 24 consecutive years (1987−2010) in a fixed sample plot with a total sample size of 6 903 trees. Random forest algorithm was used to build individual tree BAI model with 11 independent variables as individual tree, competition factor, diversity, and climate for 6 main tree species in mixed forest. And 10-fold cross-validation was used to optimize hyper parameter mtry and evaluate these models.
Result(1) The coefficients of determination for the model of BAI for the six major species of Abies nephrolepis, Picea koraiensis, Pinus koraiensis, Tilia amurensis, Betula costata, and Betula platyphylla were 0.663, 0.683, 0.695, 0.459, 0.384, and 0.568, respectively. (2) The basal area (BA) of individual tree was the most important independent variable, and had a strong positive effect on the growth of trees. Competitive factors and tree size diversity were the main factors affecting tree growth, and individual BAI decreased with increasing competitive factors and tree size diversity. (3) The effects of species diversity on tree growth were relatively limited, with increases in species diversity index and mingling degrees accelerating the growth of Abies nephrolepis, Picea koraiensis and Pinus koraiensis to some extent; whereas the effects of climatic factors on tree growth were relatively small.
ConclusionTree growth is largely depends on its own growth potential, which is mainly inhibited by competition and tree size heterogeneity in the external environment, while increased species diversity also promotes the growth of dominant species within the forest to some extent. The random forest model can well quantify and express the complex relationship between the variables and BAI of individual tree. It can be used as a tool for forest management practice and providing a new method for forest growth and yield prediction.
-
Keywords:
- tree growth /
- tree size /
- competition /
- tree species diversity /
- random forest model
-
绿色空间是各类自然、人工绿地及水域覆盖的地表空间,良好的绿色空间体系是协调城市发展和自然保护的重要保障[1-2]。在我国城镇化到绿色城镇化的发展历程中,学者不断梳理绿色空间与生态环境的关系,将绿色空间特征与调节城市热环境[3]、评估生态系统服务[4]、促进公众健康等目标有机融合[5],在城市群[6]、省市县[7]、村镇[8]等多尺度研究中广泛运用。其中,土地利用的快速转变影响了绿色空间的景观结构和生态功能,在城市区域尺度的响应尤为突出[9]。
自19世纪初期景观生态学萌芽,到分别以自然景观和人类活动为主要研究对象的路径分野,再到基于地理信息系统的空间定量计算 [10] ,区域绿色空间研究经历了静态向过程、客体向主体、现象向规律等扩展。明确绿色空间时空演变的规律和机制是区域绿色空间体系优化的依据,近年在学界形成广泛共识。其中,演变的研究主要涉及结构指标的计算,基于多时段遥感影像解译,通过 Fragstats软件计算绿色空间景观格局指数(landscape pattern indices,LPI)或景观形态空间格局分析(morphological spatial pattern analysis,MSPA)分析绿色空间格局演化[11],也有学者关注了各类绿色空间面积比例的城乡梯度特征 [12]。既有研究多直接采用土地利用一级分类划分区域绿色空间类型,一般分为林地、草地、耕地、水域和未利用地,忽略了植被覆盖密度及其生态价值的差异[12];而土地利用二级分类数量繁多,不便于以区域绿地规划为目标的研究采用。归一化差分植被指数(normalized difference vegetation index,NDVI)的遥感估算能够弥合这一缺陷,以植被覆盖密度为依据合并土地利用二级分类,进而划分绿色空间类型[13]。演变机制的研究则重点涉及空间效应的测度[14]。在空间效应特征识别方面,主要途径是通过莫兰指数(Moran’s I)、热点分析(Getis-Ord Gi*)、地理加权回归(geographically weighted regression,GWR)等空间统计方法测度绿色空间类型在研究区内的相关或异质性[15];在空间效应的驱动判断方面,多基于地理探测器等工具测度潜在影响因子的显著性和影响力 [16]。不局限于理论研究,国土空间、公园城市规划等实际操作中逐渐关注可选择的规划模式以确保生态系统服务价值等目标的最大实现[17],识别绿色空间时空演变特征是模拟预测的前置环节。
黄河下游是我国工农业生产核心区之一[18],沿岸城市发展是中国城镇化的典型缩影[19]。历史以来,河流与滩区为下游城市创造了丰厚独特的农业条件和农耕文化基础,也参与构建区域绿色空间,影响城市生态安全[20]。《黄河流域生态保护和高质量发展规划纲要》提出,有效推进下游绿色生态走廊建设和滩区生态综合整治。目前,黄河下游城市区域绿色空间时空演变研究数量有限,既有研究中尚缺对于滩区空间的考虑。基于上述认识,本文以10年为间隔,将1990、2000、2010、2020年作为时间节点,基于土地利用二级分类和植被覆盖密度划分黄河下游4个典型城市区域绿色空间类型,识别绿色空间类型转移、景观格局与城乡梯度等时空变化规律,并以地理探测方法揭示绿色空间演变的驱动机制,旨在丰富和深化黄河下游地区绿色空间发展的研究成果,为优化国土空间格局、协调生态保护与经济发展提供参考。
1. 研究区概况与数据
1.1 研究区概况
根据黄河下游自然环境的异质性和城市发展的代表性,选取郑州、新乡、济南、菏泽4城市市域范围为研究区(图1)。从自然环境看,黄河主河道流经郑州、新乡、菏泽边缘,贯穿济南市域空间;郑州市、新乡市、菏泽市境内为黄河宽滩区,其中新乡市封丘县境内还有一定面积的倒灌区,济南市境内则均为窄滩区[21]。此外,济南、郑州、新乡境内有山地分布,菏泽全境地形均为平原。从城市发展看,济南和郑州为区域性中心城市,40多年间经历了剧烈的城市化进程,新乡、菏泽则在近10年间经济增速较快。各城市基本情况见表1。
表 1 研究城市基本情况Table 1. Summary of selected cities城市 City 常住人口数量
Permanent resident population2020年国民生产总值/亿元
GDP in 2020/108 CNY行政区面积
Administrative area/km2济南 Jinan 920.2 × 104 10 140.9 10 220.0 菏泽 Heze 879.6 × 104 3 483.1 12 142.8 郑州 Zhengzhou 1 260.1 × 104 12 691.0 7 515.5 新乡 Xinxiang 617.1 × 104 3 232.5 8 271.0 1.2 数据来源
研究主要数据来源如下:
(1)土地利用/覆盖数据(https://www.resdc.cn/)。来源于由中科院地理所构建的中国多时期土地利用/土地覆盖遥感监测数据库(CNLUCC)。1990—2010年、2020年数据分别以Landsat-TM和Landsat-8遥感图像为信息源,采用5 月上旬至10 月中旬无云图,经波段提取、色彩合成、图像纠正与解译处理等过程生成,数据库Kappa系数为0.889,空间分辨率为30 m × 30 m。
(2)水系数据(https://www.webmap.cn/)。来源于国家基础地理信息中心发布的全国1∶100万公众版地理信息数据库。
(3)滩区类型数据。来源于既有文献中的判断和划定,将研究区划分宽滩区、倒灌区、窄滩区、涉滩区和滩外区[21-23]。
(4)土壤类型数据(https://www.resdc.cn/)。来源于中科院地理所发布的中国土壤类型空间分布数据库。
(5)高程数据(https://search.asf.alaska.edu/#/)。来源于NASA官方网站发布的ALOS卫星12.5 m DEM数据库。
(6)重点城镇中心点空间数据。包括4个城市主城区与周边的一、二级城镇[24],结合中国县级行政边界数据与百度地图识别获得政府所在地的空间点位。
(7)各区县经济、人口和产业数据。整理自各市统计局发布的历年统计年鉴及当年国民经济和社会发展统计公报。
2. 研究框架与方法
2.1 研究框架
研究以类型转移、景观格局和城乡梯度变化作为基本表征,识别绿色空间演变规律;进而进行自然环境和社会经济因素的地理探测,分析绿色空间演变机制(图2)。
2.2 研究方法
2.2.1 空间分类
根据既有研究对绿色空间的界定[4,25],结合CNLUCC 数据分类体系说明书,将研究区域划分为裸露地表和3类绿色空间分别为低覆盖密度绿色空间、中高覆盖密度绿色空间和高覆盖密度绿色空间(以下称为低、中、高密度绿色空间)。裸露地表包括建设用地及植被覆盖度在5%以下的空间。低密度绿色空间包括耕地及覆盖度为5% ~ 50%的草地。中密度绿色空间包括郁闭度 > 40%的矮林地与灌丛林地、郁闭度为10% ~ 30%的林地和苗圃、覆盖度 > 50%的草地及坑塘等小面积水体。高密度绿色空间包括郁闭度 > 30%的林地及河渠、湖泊、水库等大面积水体[25]。
2.2.2 空间转移矩阵
通过计算研究期初各类型面积转出和研究期末各类型空间面积转入的数值构建各类空间转移矩阵,以反映4个城市30年间各类空间相互转化的动态过程信息。转移矩阵的通用形式如下:
{{\boldsymbol{S}}_{ ij}} = \left[ {\begin{array}{*{20}{c}} {{s_{11}}}&{{s_{12}}}& \cdots &{{s_{1n}}} \\ {{s_{21}}}&{{s_{22}}}& \cdots &{{s_{2n}}} \\ \cdots & \cdots & \cdots &{{s_{3n}}} \\ {{s_{n1}}}&{{s_{n2}}}& \cdots &{{s_{nn}}} \end{array}} \right] (1) 式中:Sij为研究初期到末期时间段内i类向j类空间转移的面积矩阵;s为空间转移面积;n为空间总类型,本研究中n = 4。空间转移方向与转移量在ArcMap 10.5软件中计算,后在Origin 2021平台上绘制桑基图。
2.2.3 景观格局指数计算
选取表征空间斑块面积、密度、形状和聚散性等维度特征的6个景观格局指数,分别为斑块面积比(percentage of landscape,PLAND)、斑块数量(number of patches,NP)、最大斑块面积比(largest patch index,LPI)、斑块平均面积(mean patch size,MPS)、面积加权平均形状指数(area-weighted mean shape index,AWMSI)、功能连通度(functional connectivity,FC)[10]。所有景观格局指数均在Fragstats 4.2软件中计算。
2.2.4 距离指数与城乡梯度计算
参照中外学者对于“城市中心”的普遍定义,以各空间斑块与研究区4城市CBD的欧氏距离表征所处的城乡梯度[26]。考虑城市之间的横向比较,引入归一化距离指数(normalized distance index,NDI),其公式如下:
{\rm{NDI}} = \displaystyle \sum\nolimits_{i=1}^n {\frac{{\sqrt {{{\left( {{x_i} - X} \right)}^2} + {{\left( {{y_i} - Y} \right)}^2}} }}{{{d_{\max }}}}} (2) 式中:X、Y表示城市中心的横纵坐标,xi和yi表示绿色空间斑块中心的横纵坐标,分子即为斑块i中心到城市中心的欧氏距离,dmax为所有斑块中心到城市中心欧式距离中的最大值。欧氏距离在ArcMap 10.5软件中计算,以0.01个距离为单位计算各类空间面积占比,后在RStudio 3.3.1 平台上调用ggplot2 软件包,插入“Loess”统计曲线模型,基于局部加权回归原理形成城乡梯度轨迹曲线[27]。
2.2.5 影响因素的地理探测
使用地理探测器工具中的因子探测计算影响因子对绿色空间地理分布的影响,使用交互探测计算两两因子的共同作用。地理探测器是一种探测自变量对因变量空间异质性解释力的统计学方法,基本假设为:如果某种自变量能够影响因变量的地理空间分布,那么自变量与因变量应具有相似的地理空间特征[28]。
{q} = 1 - \frac{1}{{n{\sigma ^2}}}\sum\nolimits_{h = 1}^L {{n_h}} \sigma _h^2 (3) 式中:q为影响因子对绿色空间地理分布的解释力;n、
{\sigma ^2} 分别为整个研究区域中绿色空间地理分布的样本量和方差,{n_h}、\sigma _h^2 分别为次级区域h中绿色空间地理分布的样本量和方差。q取值范围为[0,1],数值越接近1表示解释力越强。在地理探测步骤,对所有数值型数据进行离散化处理得到类型栅格数据集,建立渔网并作采样。3. 结果与分析
3.1 绿色空间转移面积
由表2计算,1990—2020年研究区各类空间转移面积为5 139 km2,占总面积的13.47%。绿色空间总计向裸露地表转出3 223 km2;裸露地表向各类绿色空间转出1 181 km2;绿色空间内部互相转换735 km2,其中较高密度向较低密度绿色空间转出466 km2,较低密度向较高密度绿色空间转出269 km2。4个城市转移量按比例大小排顺序为济南、菏泽、郑州、新乡。山东省两城市绿色空间呈现一增三减的变化,裸露地表面积增加,高、中、低密度绿色空间面积减少。河南省两城市为二增二减,裸露地表与高密度绿色空间面积增加,中低密度绿色空间面积减少。各级城镇边缘、宽滩区沿线绿色空间转移明显(图3)。
表 2 1990—2020年绿色空间转移矩阵Table 2. Matrix of greenspace transfer from 1990 to 2020km2 年份 Year 城市 City 指标 Index 2020 裸露地表
Bare surface低密度
Low density中密度
Medium density高密度
High density1990 济南 Jinan 裸露地表 Bare surface 904 296 1 36 低密度 Low density 1 137 5 319 6 85 中密度 Medium density 16 19 349 3 高密度 High density 115 139 1 1 792 菏泽 Heze 裸露地表 Bare surface 1 462 532 1 25 低密度 Low density 1 005 8 727 0 66 中密度 Medium density 9 35 6 0 高密度 High density 26 106 0 139 郑州 Zhengzhou 裸露地表 Bare surface 1 453 54 0 54 低密度 Low density 583 4 098 1 65 中密度 Medium density 24 56 141 21 高密度 High density 37 57 6 864 新乡 Xinxiang 裸露地表 Bare surface 1 166 127 0 55 低密度 Low density 253 5 473 0 20 中密度 Medium density 0 4 43 2 高密度 High density 18 42 1 1 062 ![]() 图 3 1990—2020年绿色空间转移的空间分布Figure 3. Distribution of greenspace transfer from 1990 to 2020
图 3 1990—2020年绿色空间转移的空间分布Figure 3. Distribution of greenspace transfer from 1990 to 202030年间,低密度绿色空间一直是裸露地表面积的主要转入来源(图4)。2000—2010年间,济南、菏泽两市低密度绿色空间与裸露地表大量相互转化,主要是受2006年山东省出台的《城镇建设用地增加与农村建设用地减少相挂钩管理办法》等相关政策影响[29],大量零星分散的农村居民点发生搬迁撤并或土地流转。新乡、郑州则主要是沿黄滩涂向耕地的转移。
![]() 图 4 1990—2020年绿色空间转移桑基图Figure 4. Sankey diagram of greenspace transfer from 1990 to 2020
图 4 1990—2020年绿色空间转移桑基图Figure 4. Sankey diagram of greenspace transfer from 1990 to 20203.2 绿色空间景观指数变化
3.2.1 景观指数总体变化
由1990—2020年景观指数变化图(图5)可知:在斑块面积比(PLAND)方面,郑州和济南低密度绿色空间数值减少较多,分别为6.41%和6.50%,裸露地表分别增加7.13%、9.14%,均远高于平均值。中、高密度绿色空间变化较小。
![]() 图 5 1990—2020年绿色空间景观指数总体变化PLAND.斑块面积比;NP.斑块数量;LPI.最大斑块面积比;MPS.斑块平均面积;AWMSI.面积加权平均形状指数;FC.功能连通度。下同。PLAND, percentage of landscape; NP, number of patch; LPI, largest patch index; MPS, mean patch size; AWMSI, area-weighted mean shape index; FC, functional connectivity. The same below.Figure 5. Overall changes of landscape pattern indices from 1990 to 2020
图 5 1990—2020年绿色空间景观指数总体变化PLAND.斑块面积比;NP.斑块数量;LPI.最大斑块面积比;MPS.斑块平均面积;AWMSI.面积加权平均形状指数;FC.功能连通度。下同。PLAND, percentage of landscape; NP, number of patch; LPI, largest patch index; MPS, mean patch size; AWMSI, area-weighted mean shape index; FC, functional connectivity. The same below.Figure 5. Overall changes of landscape pattern indices from 1990 to 2020斑块数量(NP)方面,除济南低密度绿色空间与裸露地表数值增加外,其余各城市均有所减少,低、中、高密度绿色空间减少量均以郑州为最大,新乡次之,可见4个城市大多数绿色空间斑块均发生整合。
最大斑块面积比(LPI)方面,4个城市裸露地表数值均有所增加,低密度绿色空间数值均有减少,以郑州与济南的变化量最大,均达到平均水平的2倍以上。中、高密度绿色空间数值变化量相对较小,个体斑块面积增减量未影响整体优势性,表明大规模自然林地格局总体平稳。
面积加权平均形状指数(AWMSI)方面,济南裸露地表数值增加明显,低密度绿色空间数值减少。在NP增加的情况下,表明无植被覆盖的城乡建设用地向外延伸而降低了整体形态的规则程度,同时耕地、草地被人工方整切割。此外,郑州和新乡裸露地表的数值也有较大程度的减少,在PLAND增加、NP减少的情况下,能够反映出郑州城乡建设用地以集聚扩张的形式增长,具有向心性特征。
平均斑块面积(MPS)指数方面,变化主要发生于低密度绿色空间。除济南数值减少外,郑州、新乡、菏泽均有增加。结合NP值变化,可知济南的大面积的低密度绿色空间被切分独立;菏泽低密度绿色空间数值增加最大,表明大量小碎斑块转出。新乡的高密度绿色空间数值增量最大,河南城市增量明显大于山东城市。
功能连接度(FC)方面,郑州、济南的低密度绿色空间,郑州、济南、菏泽的中密度绿色空间,以及济南的高密度绿色空间数值有所增加,且区域性中心城市的改善更为全面。同时,4个城市的裸露地表的数值变化较小,在NP减少、PLAND增加的前提下,较小的FC变化表明斑块之间的最大可能连接数变小,即斑块面积的增加与数量的减少在空间中是相对均匀发生的,反映了城市与村镇建设用地共同发生扩张。
3.2.2 景观格局指数变化轨迹
由1990—2020 年景观格局指数变化折线图(图6)可知, 4个城市区域绿色空间景观格局指数在过去 30 年间的变化轨迹具有一定共性。低密度绿色空间的PLAND、AWMSI、FC,中密度绿色空间的PLAND、NP、LIP、AWMSI,高密度绿色空间的各项指数及裸露地表的PLAND、NP、LIP、MPS、FC变化轨迹均较为相似。比较特殊的轨迹表现于以下方面:郑州中密度绿色空间的MPS、AWMSI和FC的基数在30年内均为最大,其中MPS在1990—2010年间的上升趋势强于其他3市,2010年以来与其他城市呈现一致的平稳趋势。济南裸露地表的AWMSI在2000—2010年间发展迅速并超越郑州至首位,空间限制使建设用地形态不规则度上升。同一时期低密度绿色空间的LIP明显单调下降,耕地和草地面积有所收缩;NP单调上升,变化趋势与其他3市相反,并超越郑州发展至首位。上述变化在2010年后恢复平稳,严格的耕地保护和土地集约利用政策效果显现。菏泽低密度绿色空间的MPS、FC基数远大于其他城市,受黄淮海平原农业开发战略影响,相较于其他城市的平稳轨迹经历了先上升后下降的明显变化。与其他城市的平稳趋势相比,新乡高密度绿色空间的MPS在30年间保持速率均匀的单调上升趋势; FC以2000年前后为转折,呈现“∩”型发展趋势,但始终处于首位。
![]() 图 6 1990—2020年景观格局指数变化折线图Figure 6. Point-fold line chart of landscape pattern indices from 1990 to 2020
图 6 1990—2020年景观格局指数变化折线图Figure 6. Point-fold line chart of landscape pattern indices from 1990 to 2020此外,4个城市高密度绿色空间的LIP、AWMSI数值在30年间均无较大波动,由高到低顺序始终为新乡、郑州、济南、菏泽,河南两城市总体高于山东两城市。中密度绿色空间的LIP、MPS、AWMSI值也未发生排位顺序的变化,由高到低顺序为郑州、济南、新乡、菏泽,区域性中心城市总体高于一般城市。
3.3 绿色空间面积比例的城乡梯度变化
![]() 图 7 1990—2020年绿色空间面积占比的城乡梯度变化Figure 7. Urban-to-rural trajectory of the area percentage of greenspace from 1990 to 2020
图 7 1990—2020年绿色空间面积占比的城乡梯度变化Figure 7. Urban-to-rural trajectory of the area percentage of greenspace from 1990 to 2020低密度绿色空间的城乡梯度轨迹在时间序列中的变化较为复杂。1990和2000年,济南的轨迹由中心向外围先缓慢降低再升高,降低区间为山区地带;2010和2020年,由于城市建设用地扩张导致近郊低密度绿色空间比例锐减,而在NDI > 0.750的区间上保持平稳。2000年,郑州的轨迹在0.125 < NDI < 0.375的区间较前期上升,但2010年后又下降,到2020年,在近远郊地带均出现一定程度下降。新乡较早时期的城乡梯度变化轨迹是“W”形,波峰出现在NDI = 0.500的中间地带。2000和2020年,这一波峰逐渐削弱,轨迹逐步转变为“U”形。菏泽市的轨迹则在 30年间均可见波谷,其形态在30年间由“U”向“V”发展,波谷稳定存在于0.375 < NDI < 0.625的区间。
各城市中密度绿色空间的城乡梯度轨迹在时间序列上体现出一致性。济南中心到外围趋于水平,整体缓慢降低。菏泽的轨迹更加平直,且始终处于低位。郑州、新乡两城市的城乡梯度变化轨迹呈现较为相似的“U”形,分别在NDI = 0.50和NDI = 0.375的距离地带出现了小的峰值。新乡在1990—2010年间的轨迹为“U”形,中部地带为较宽波谷。
高密度绿色空间的梯度相对固定,并反映出城市自然本底、城镇布局方式等空间特征。如济南的峰值出现在NDI = 0.500的中段地带,郑州则在NDI > 0.750的区间上升,均对应于山区自然林地。平原城市菏泽城乡梯度轨迹在30年间没有明显差异,从中心向外保持平稳波动,整体比例处于与中密度绿色空间轨迹类似的低位,最大值出现在远郊水域地带。早期新乡的轨迹在市域中间地带存在小的峰值,后随着城市扩张而被削弱,但可见近郊高密度空间在近年出现增量,主要是依托引黄干渠等水系而发展的城市绿地。
裸露地表的城乡梯度轨迹反映了市域城、镇、村建设用地的发展格局。济南30年间的轨迹表现为单调递增的指数形态,反映城镇布局向外延伸,以及泰山山脉南侧莱芜、钢城两区的发育态势。郑州在1990前年有类似的情况,2000和2010年,在中心城区边缘地带内先后有中牟、荥阳和新郑等城镇发育扩张,使整体变化轨迹呈现出“U”形。新乡的轨迹则在1990和2000年从中心向外围缓慢升高,2010和2020年在NDI = 0.500的距离上形成波峰,对应于第一圈层城镇的扩张成熟。菏泽轨迹的波动幅度不大(图7)。
3.4 绿色空间演变的驱动机制
以绿色空间分布为地理探测中的因变量,以4个表征自然环境的因子(X1 ~ X4)与4个表征社会经济情况的因子(X5 ~ X8)为自变量,计算单一因子、因子间交互解释力。由于1990年的社会经济数据难以全面获取,以下仅探讨2000—2020年间各项因子对绿色空间演变的驱动机制。
根据因子探测的结果(表3),高程、与水系距离、与城镇距离、一产比例、GDP和城乡人口比例6项因子在20年间均对济南区域绿色空间分布有显著贡献,综合解释力由高至低,其中社会经济因素解释力的q累计值上升5.2%。与滩区关系在20年间始终对菏泽区域绿色空间分布显著贡献,除此之外有显著解释力的均为社会经济因素,q累计值上升3.1%。土壤类型外的其他7项因子均在20年间对郑州区域绿色空间分布有显著贡献,高程的解释力最大,社会经济因素解释力的q累积值上升3.9%。自然环境因素在新乡占据明显主导,社会经济因素解释力的q累计值上升8.4%。
表 3 因子探测结果Table 3. Factor detection results城市
City年份
Year与水系距离
Distance from waters (X1)土壤类型
Soil type (X2)与滩区关系
Relationship with floodplain (X3)高程
Altitude (X4)与城镇距离
Distance from town (X5)生产总值
GDP (X6)城乡人口比例
Proportion of
urban and rural
population (X7)一产比例
Proportion of
primary industry (X8)济南 Jinan 2000 0.304*** 0.001 0.003** 0.328*** 0.094*** 0.029*** 0.003*** 0.024*** 2010 0.244 0.001 0.001 0.283*** 0.092*** 0.018*** 0.013*** 0.028*** 2020 0.243*** 0.001 0.001 0.283*** 0.073*** 0.048*** 0.014*** 0.067*** 菏泽 Heze 2000 0.004 0.002 0.005* 0.002 0.004 0 0 0.005** 2010 1.000 0.872 0.011*** 0.002 0.032*** 0.003 0.004* 0.003* 2020 0 0 0.011*** 0.002 0.036*** 0.000 0.001 0.001 郑州 Zhengzhou 2000 0.039*** 0 0.012*** 0.005 0.027*** 0.014*** 0.031*** 0.022*** 2010 0.037*** 0.001 0.012*** 0.194 *** 0.034*** 0.026*** 0.077*** 0.073*** 2020 0.045*** 0.001 0.017*** 0.192*** 0.037*** 0.056*** 0.077*** 0.063*** 新乡 Xinxiang 2000 0.412*** 0.004 0.047*** 0.531*** 0.096*** 0.109*** 0.145*** 0.067*** 2010 0.401*** 0.004 0.043*** 0.526*** 0.103*** 0.166*** 0.086*** 0.082*** 2020 0.393*** 0.005 0.042*** 0.518*** 0.111*** 0.164*** 0.045*** 0.181*** 注:表中数字为解释力q值;*表示在P < 0.1水平上差异显著,**表示在P < 0.05水平上差异显著,***表示在P < 0.01水平上差异显著。Notes: the number in the table is the explanatory power q-value; * means significant differences at P < 0.1 level, ** means significant differences at P < 0.05 level, and *** means significant differences at P < 0.01 level. 根据交互探测结果,菏泽未见因子间的显著交互作用,而郑州、新乡、济南3城市均可见因子共同作用的效果大于独立效果之和的情况,即两两交互的类型为非线性增强。以郑州为例(表4),在2000年仅有土壤类型∩高程产生显著非线性增强效果;2010年增加两对,分别是与滩区关系∩高程、土壤类型∩一产比例;2020年,土壤类型分别与高程、GDP、城乡人口比例、一产比例产生非线性增强效果。表明社会经济因素与自然环境因素体现愈加明显的交互协同。
表 4 交互探测结果(以郑州为例)Table 4. Interactive detection results (taking Zhengzhou as an example)年份
YearX1 X2 X3 X4 X5 X6 X7 X8 2000 X1 0.039 X2 0.003 X3 0.012 X4 0.221 0.205 X5 0.027 X6 0.014 X7 0.031 X8 0.022 2010 X1 0.037 X2 0.001 X3 0.012 X4 0.209 0.209 0.194 X5 0.034 X6 0.026 X7 0.077 X8 0.088 0.073 2020 X1 0.045 X2 0.001 X3 0.017 X4 0.240 0.211 0.192 X5 0.037 X6 0.067 0.056 X7 0.065 0.057 X8 0.073 0.063 注:表中数字为交互作用解释力q值,此表仅显示具有显著性的q值。Notes: the number in the table is the explanatory power q-values of interaction. This table only shows the q values with significant interaction relationship. 4. 讨 论
4.1 区域绿色空间演变特征识别
(1)各类空间的面积增减。研究显示,30年间黄河下游4个城市裸露地表规模持续扩张,低密度绿色空间面积减量最大,中、高密度绿色空间面积增减相对平稳。研究区城乡建设用地共同经历扩张,但区域性中心城市与一般城市之间存在增量和形态上的差别,即新增城市建设用地更多直接接壤城市边缘,集中连片的增长现象更为显著。济南中心城区以山前平原为核心,北向黄河、南向浅山地带逐步延展,在空间的制约中侵占了一定面积中、高密度绿色空间。郑州中心城区的发展以近郊城镇集聚和自身边缘扩张为主,北部边缘向滩区与水域靠近的同时,也建设了有规模的高密度绿色空间,将滩地滩涂转为坑塘湿地和林地为主的生态公园。4个城市内,小而碎的低密度绿色空间均有转出,但城乡建设行为也挤压或分割了大量规模性的耕地、草地。近年来,在严格耕地保护政策出台和城市存量发展导向下,低密度绿色空间减量得到控制。
(2)同类空间的整合连通。30年间,破碎分散向整合连通是各城市区域绿色空间斑块的主要发展趋势。中、高密度绿色空间表现更为明显,其破碎度降低、连通度和集聚水平增加,能够对物种保护、能量交换等生态系统服务价值改善起到积极作用[8],同时可见区域性中心城市的改善尤为全面、明显。此外,济南中心城区低密度绿色空间在2010年以前的形态破碎而不规则,但这一现象在近10年有所改观,反映出山东省土地综合整治等政策的效果。
(3)面积占比的时空分异。30年间,城乡轨迹变化最明显的是区域性中心城市的低密度绿色空间,而各城市中、高密度绿色空间30年间的轨迹具有相对一致性。研究范围内的黄淮海冲击平原区和鲁西平原区是典型的农业景观,耕地为主的低密度绿色空间是最主要的景观类型,外围太行、嵩山、泰山山区内则以中、高密度绿色空间为主,构成了市域重要的生态屏障。在此自然本底上,绿色空间面积比例的城乡梯度本身具有一定特殊性,剧烈的城市化进程中使之发生了不同幅度的变化,与“从城区到郊区,人造绿地数量不断减少,自然绿色空间逐渐增加”的西方经验有所区别[30]。
(4)“自然−社会”的互构作用。研究呈现了“水”“山”“滩”对黄河下游城市区域绿色空间长期性、大尺度的影响,探测了“自然−社会”交互作用的增长,与黄河下游城市“经济系统和谐发展指数最高,环境系统次之”的研究结果相互印证[31]。从同一时期的城乡梯度上看,菏泽、新乡的低密度绿色空间在城市近郊和远郊占比最大,这一现象与两个城市圈层式的布局有关。高密度植被面积比例的城乡梯度轨迹则主要体现了市域范围内山地、滩区等地理单元的空间特征,同时反映出区域政策的引导下城市边缘区生态质量的改变。
4.2 区域绿色空间优化建议
区域绿色空间规划需充分考虑其演变的规律和机制,“因城制宜”。如与其他城市相比,菏泽市域范围自然基底更均质,绿色空间演变多以社会经济发展为动力,需加强规划决策对区域绿色空间建设的助推作用。现状低密度绿色空间在市域范围内均占据主导,应科学规划林地−树团−点状树木等防护绿地结构,丰富农田为主的大尺度国土景观,保障林地、水网为非农生境提供稳定的生物多样性及生态效应[32];中心城区近郊地带中高密度绿色空间面积比例偏低但有较好的连通性基础,水系与林网构成的生态廊道应是优化重点[33]。自然环境因素对新乡区域绿地格局影响力的主导地位最为突出,未来仍需围绕自然要素优化绿色空间格局,将高密度绿色空间面积平稳上升的趋势由山区延伸至近郊与中心城区,进一步发挥滩区与水系的诱导效应,提升滩区沿线、引黄干渠沿线的生境质量。在区域性中心城市郑州、济南,区域绿色空间与社会经济因素关联性日益增强,中心城区周边生态改善趋势显现。未来建设中,一方面需重点考虑城−滩关系,通过资源的保护利用引导现状滩区沿线中密度绿色空间向高密度过度,提升城市边缘区高密度绿色空间的连通性和植被多样性,以公园、绿道建设为引力增进绿色空间建设与经济发展之间的良性互动;另一方面要规划生态分区,防止中、高密度绿色空间规模损失,防范城市建设行为对滩区和山区的重要生境造成扰动。值得注意的是,土壤类型因子在4个城市中往往不具备独立影响力,但与各项表征社会经济的因子交互协同,表明人工绿地建设受到土壤条件的制约。黄河下游尤其是滩区周边城市在进行区域绿色空间规划时,需根据立地条件合理配置不同生长习性的植被,保障区域绿地的可持续发展。
5. 结 论
研究定量测度了1990—2020年间黄河下游4城市区域绿色空间的类型转移、景观格局、城乡梯度的时空变化规律,并揭示了绿色空间演变的驱动机制,主要结论如下:(1)30年间,区域绿色空间总量损失,绿色空间内部转移以中、高密度向低密度的转出为主,各级城镇建成区边缘、宽滩区沿线转移较多;(2)景观格局的总体变化趋势为由明显波动到趋近平稳、由破碎分散向整合连通,中、高密度绿色空间指标的改善态势在区域性中心城市更为明显;(3)城乡梯度特征在城市间、绿色空间类型间存在较大差异;(4)影响因子间对绿色空间地理分布的协同作用渐增,绿色空间逐渐成为自然−社会互构的结果。伴随黄河下游簇群城镇形态的成熟与黄河流域高质量发展战略的深入实施,未来的绿色空间应由“屏障”转为“枢纽”,在区域生态保护和经济社会发展中发挥更多助益。
受限于数据的可获取性,本研究仅对近20年的绿色空间演变机制进行研究,且基于时间节点数据的分析结果相对局限。另一方面,不同遥感数据的差异可能会对比较研究造成干扰。未来将加强遥感、经济、人口等面板数据的追踪分析,构建黄河下游城市多精度LUCC、社会经济、政策发展综合数据库,并深入生态系统服务、市域生态网络构建等多目标研究。
-
![]()
图 1 各因子对不同树种BAI生长的偏依赖关系
Figure 1. Partial dependence of various factors on BAI growth of different tree species
表 1 样地信息统计表(1987)
Table 1 Statistical table of sample plot information (1987)
样地号
Sample plot No.面积/hm2
Area/ha平均胸径
Mean DBH/cm海拔
Altitude/m坡向
Slope aspect坡度
Slope degree/(°)树种组成
Tree species composition1 0.160 18.8 780 东北 Northeast 13 5云3冷1红1枫
5P3A1PK1B2 0.250 22.0 650 西南 Southwest 28 4冷2红2椴1枫1白
4A2PK2T1B1BP3 0.360 15.9 750 东北 Northeast 7 4冷3云1红1椴1枫
4A3P1PK1T1B4 0.312 16.3 780 东北 Northeast 10 5冷3云1红1枫
5A3P1PK1B5 0.200 20.4 634 东北 Northeast 26 3云3红2椴1冷1白
3P3PK2T1A1BP注:P. 红皮云杉 Picea koraiensis; A. 臭冷杉 Abies nephrolepis; PK. 红松 Pinus koraiensis; B. 枫桦 Betula costata; T. 椴树 Tilia amurensis; BP. 白桦 Betula platyphylla.  下载: 导出CSV
下载: 导出CSV
表 2 模型因子属性统计表
Table 2 Statistics table of model factor attributes
类别
Category因子
Factor最小值
Min.最大值
Max.平均值
Mean单木因子 Individual tree factor BA/(10−4 m2) 9.62 2341.40 295.64 竞争因子 Competitive factor BAL/(m2·ha−1) 0 11.10 6.08 CI 0.23 26.28 2.73 SDI 99.03 284.49 214.33 死亡率 Mortality rate/% 0 22 5.1 多样性 Diversity 树木大小多样性
Tree size diversityDIV_Size 2.44 2.87 2.67 U 0 1 0.50 树种多样性
Tree species diversityDIV_SP 1.54 2.05 1.74 M 0 1 0.67 气候 Climate 年平均温度
Average annual temperature/℃1.89 6.25 5.02 年平均降雨量
Average annual rainfall/mm393.90 735.38 588.15 注:BA. 单木胸高断面积;BAL. 大于对象木的林木断面积之和;CI. 对象木的竞争指数;SDI. Reineke的林分密度指数;DIV_Size. 树木大小多样性;U. 大小比数;DIV_SP. 林分树种多样性;M. 混交度。Notes: BA, basal area of individual tree; BAL, sum of basal area of forest tree that is greater than the object trees; CI, competition index of object tree; SDI, stand density index of Reineke; DIV_Size, tree size diversity; U, dominance degree; DIV_SP, tree species diversity of stand; M, mingling degree.
下载: 导出CSV
表 3 主要树种信息统计表
Table 3 Statistical table of main tree species information
树种
Tree species株数
Number of plant胸高断面积
BA/cm2胸高断面积生长量/(cm2·a−1)
BAI/(cm2·year−1)最小值
Min.q25 q50 q75 最大值
Max.平均值
Mean最小值
Min.q25 q50 q75 最大值
Max.平均值
Mean臭冷杉 Abies nephrolepis 2 375 19.6 91.6 243.3 447.7 1 562.3 311.2 0 2.7 6.5 11.7 40.6 8.0 云杉 Picea koraiensis 1 650 20.4 95.0 240.5 526.9 1 870.4 346.1 0 2.0 6.5 14.8 48.7 9.6 红松 Pinus koraiensis 898 9.6 59.4 124.3 411.9 2 341.4 293.8 0 1.5 4.2 11.6 47.6 8.1 椴树 Tilia amurensis 661 12.8 124.7 264.5 481.1 1 676.4 337.3 0 1.1 3.2 7.4 36.5 5.0 枫桦 Betula costata 899 23.8 50.3 78.5 118.3 1 809.6 109.8 0 0.3 1.8 3.9 21.6 2.8 白桦 Betula platyphylla 420 28.3 172.0 268.8 401.2 1 764.6 346.1 0 4.2 7.8 13.4 37.7 9.2 注:q25、q50、q75分别为下四分位数、中位数和上四分位数。Notes: q25, q50, and q75 are the lower quartiles, median, and the upper quartiles, respectively.
下载: 导出CSV
表 4 10折交叉验证结果
Table 4 10-fold blocked cross-validation results
树种 Tree species mtry R2 ± SD RMSE ± SD 臭冷杉 Abies nephrolepis 4 0.663 ± 0.052 3.897 ± 0.263 云杉 Picea koraiensis 4 0.683 ± 0.058 5.272 ± 0.445 红松 Pinus koraiensis 4 0.695 ± 0.064 5.067 ± 0.737 椴树 Tilia amurensis 4 0.459 ± 0.069 3.969 ± 0.294 枫桦 Betula costata 2 0.384 ± 0.075 2.523 ± 0.262 白桦 Betula platyphylla 5 0.568 ± 0.101 4.540 ± 0.719
下载: 导出CSV
表 5 模型因子相对重要性
Table 5 Relative importance of model factors
% 类别
Category因子
Factor臭冷杉
Abies
nephrolepis云杉
Picea
koraiensis红松
Pinus
koraiensis椴树
Tilia
amurensis枫桦
Betula
costata白桦
Betula
platyphylla单木因子
Individual tree factorBA 13.3 13.1 19.2 19.8 22.6 17.5 竞争因子
Competitive factorBAL 10.6 9.6 10.0 14.6 13.8 11.6 CI 12.6 10.1 11.4 5.8 8.0 10.2 SDI 8.3 9.3 8.5 10.6 7.0 14.0 死亡率 Mortality rate 8.9 8.2 4.8 10.6 6.7 9.7 多样性 Diversity 树木大小多样性
Tree size diversityDIV_Size 10.1 12.8 8.0 14.0 6.6 9.7 U 6.8 9.0 11.4 7.4 10.8 5.1 树种多样性
Tree species diversityDIV_SP 8.5 7.8 6.0 5.5 7.1 7.4 M 6.6 5.8 7.3 2.7 4.9 7.1 气候 Climate 年平均降雨量
Average annual rainfall5.1 6.0 5.8 2.0 5.0 2.3 年平均温度
Average annual temperature9.2 8.2 7.6 6.9 7.4 5.4
下载: 导出CSV
-
[1] Schlamadinger B, Bird N, Johns T, et al. A synopsis of land use, land-use change and forestry (lulucf) under the kyoto protocol and marrakech accords[J]. Environmental Science & Policy, 2007, 10(4): 271−282.
[2] 胡松. 湖南栎类天然林断面积生长模型研究[D]. 长沙: 中南林业科技大学, 2019. Hu S. Research on basal area growth model for oak natural forest in Hunan Province [D]. Changsha: Central South University of Forestry & Technology, 2019.
[3] Jevsenak J, Skudnik M. A random forest model for basal area increment predictions from national forest inventory data[J]. Forest Ecology and Management, 2021, 479: 118601.
[4] Jiang X, Huang J, Cheng J, et al. Interspecific variation in growth responses to tree size, competition and climate of western canadian boreal mixed forests[J]. Science of the Total Environment, 2018, 631−632: 1070−1078. doi: 10.1016/j.scitotenv.2018.03.099
[5] 江小雷, 岳静, 张卫国, 等. 生物多样性, 生态系统功能与时空尺度[J]. 草业学报, 2010, 19(1): 219−225. Jiang X L, Yue J, Zhang W G, et al. Biodiversity, ecosystem functioning and spatio-temporal scales[J]. Acta Prataculturae Sinica, 2010, 19(1): 219−225.
[6] Lei X, Wang W, Peng C. Relationships between stand growth and structural diversity in fpruce-dominated forests in new Brunswick, Canada[J]. Canadian Journal of Forest Research, 2009, 39(10): 1835−1847. doi: 10.1139/X09-089
[7] Subedi N, Sharma M. Climate-diameter growth relationships of black spruce and jack pine trees in Boreal Ontario, Canada[J]. Global Change Biology, 2013, 19(2): 505−516. doi: 10.1111/gcb.12033
[8] Chen L, Huang J, Stadt K, et al. Drought explains variation in the radial growth of white spruce in western Canada[J]. Agricultural and Forest Meteorology, 2017, 233: 133−142. doi: 10.1016/j.agrformet.2016.11.012
[9] Cortini F, Filipescu C, Groot A, et al. Regional models of diameter as a function of individual tree attributes, climate and site characteristics for six major tree species in Alberta, Canada[J]. Forests, 2011, 2(4): 814−831. doi: 10.3390/f2040814
[10] Ashraf M, Zhao Z, Bourque C, et al. Integrating biophysical controls in forest growth and yield predictions with artificial intelligence technology[J]. Canadian Journal of Forest Research, 2013, 43(12): 1162−1171. doi: 10.1139/cjfr-2013-0090
[11] 方匡南, 吴见彬, 朱建平, 等. 随机森林方法研究综述[J]. 统计与信息论坛, 2011, 26(3): 32−38. Fang K N, Wu J B, Zhu J P, et al. A review of technologies on random forests[J]. Journal of Statistics and Information, 2011, 26(3): 32−38.
[12] 高若楠, 苏喜友, 谢阳生, 等. 基于随机森林的杉木适生性预测研究[J]. 北京林业大学学报, 2017, 39(12): 36−43. Gao R N, Su X Y, Xie Y S, et al. Prediction of adaptability of Cunninghamia lanceolata based on random forest[J]. Journal of Beijing Forestry University, 2017, 39(12): 36−43.
[13] 董灵波, 梁凯富, 张一帆, 等. 基于Landsat 8时间序列数据的翠岗林场森林类型划分[J]. 应用生态学报, 2022, 33(9): 2339−2346. Dong L B, Liang K F, Zhang Y F, et al. Classification of forest types in Cuigang Forest Farm based on time series data of Landsat 8[J]. Chinese Journal of Applied Ecology, 2022, 33(9): 2339−2346.
[14] 徐丽, 欧阳勋志, 潘萍, 等. 基于GF-1 WFV与MODIS时空融合的南方森林植被类型识别[J]. 应用生态学报, 2022, 33(7): 1948−1956. Xu L, Ouyang X Z, Pan P, et al. Identification of forest vegetation types in southern China based on spatio- temporal fusion of GF-1 WFV and MODlS data[J]. Chinese Journal of Applied Ecology, 2022, 33(7): 1948−1956.
[15] 孙铭辰, 姜立春. 基于机器学习算法的樟子松立木材积预测[J]. 南京林业大学学报(自然科学版), 2023, 47(1): 31−37. Sun M C, Jiang L C. Standing volume prediction of Pinus sylvestris var. mongolica based on machine learning algorithm[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2023, 47(1): 31−37.
[16] Lh R. Perfecting a stand-density tndex for even-aged forests [J]. Journal of Agricultural Research, 1933, 46: 627−638.
[17] Hegyi F. A simulation model for managing Jack-pine stands simulation[J]. Royalcoll. For, Res. Notes, 1974, 30: 74−90.
[18] 惠刚盈, 胡艳波. 混交林树种空间隔离程度表达方式的研究[J]. 林业科学研究, 2001, 14(1): 23−27. doi: 10.3321/j.issn:1001-1498.2001.01.004 Hui G Y, Hu Y B. Measuring species spatial lsolation in mixed forests[J]. Forest Research, 2001, 14(1): 23−27. doi: 10.3321/j.issn:1001-1498.2001.01.004
[19] 惠刚盈. 一个新的林分空间结构参数: 大小比数[J]. 林业科学研究, 1999, 12(1): 4−9. Hui G Y. A new parameter for stand spatial structure: neighbourhood comparison[J]. Forest Research, 1999, 12(1): 4−9.
[20] Breiman L. Random forests[J]. Machine Learning, 2001, 45(1): 5−32. doi: 10.1023/A:1010933404324
[21] Arlot S, Celisse A. A survey of cross-validation procedures for model selection[J]. Statistics Surveys, 2010, 4: 40−79.
[22] Willmott C J, Matsuura K. Advantages of the mean absolute error (mae) over the root mean square error (rmse) in assessing average model performance[J]. Climate Research, 2005, 30(1): 79−82.
[23] 董雪婷, 张静, 张志东, 等. 树种相互作用、林分密度和树木大小对华北落叶松生产力的影响[J]. 应用生态学报, 2021, 32(8): 2722−2728. Dong X T, Zhang J, Zhang Z D, et al. Effects of tree species interaction, stand density, and tree size on the productivity of Larix principis-rupprechtii[J]. Chinese Journal of Applied Ecology, 2021, 32(8): 2722−2728.
[24] 吕沅杭, 伊利启, 王儒林, 等. 基于空间结构参数的大兴安岭天然落叶松单木直径生长模型[J]. 林业科学研究, 2021, 34(2): 81−91. Lü Y H, Yi L Q, Wang R L, et al. Diameter growth model using spatial structure parameters of natural Larix gmelinii stand in Daxing’anling Mountains, northeast China[J]. Forest Research, 2021, 34(2): 81−91.
[25] Lin S, Li Y, Li Y H, et al. Influence of tree size, local forest structure, topography, and soil resource availability on plantation growth in Qinghai Province, China[J]. Ecological Indicators, 2021, 120: 106957.
[26] Adame P, Hynynen J, Canellas I, et al. Individual-tree diameter growth model for rebollo oak (Quercus pyrenaica Willd.) coppices[J]. Forest Ecology and Management, 2008, 255(3): 1011−1022.
[27] 任玫玫, 杨华. 长白山云冷杉林优势树种的竞争[J]. 应用生态学报, 2016, 27(10): 3089−3097. Ren M M, Yang H. Competition among dominant tree species in a natural spruce-fir forest in Changbai Mountain[J]. Chinese Journal of Applied Ecology, 2016, 27(10): 3089−3097.
[28] Liu D, Zhou C, He X, et al. The effect of stand density, biodiversity, and spatial structure on stand basal area increment in natural spruce-fir-broadleaf mixed forests[J]. Forests, 2022, 13(2): 162.
[29] 陈梦飞. 青海东部黄土区青海云杉人工林空间结构对径向生长的影响[D]. 北京: 北京林业大学, 2019. Chen M F. Effects of spatial structure of Picea crassifolia plantationon radial growth in the loess area of eastern Qinghai[D]. Beijing: Beijing Forestry University, 2019.
[30] 吕延杰, 杨华, 张青, 等. 云冷杉天然林林分空间结构对胸径生长量的影响[J]. 北京林业大学学报, 2017, 39(9): 41−47. Lü Y J, Yang H, Zhang Q, et al. Effects of spatial structure on DBH increment of natural spruce-fir forest[J]. Journal of Beijing Forestry University, 2017, 39(9): 41−47.
[31] 余黎, 雷相东, 王雅志, 等. 基于广义可加模型的气候对单木胸径生长的影响研究[J]. 北京林业大学学报, 2014, 36(5): 22−32. Yu L, Lei X D, Wang Y Z, et al. Impact of climate on individual tree radial growth based on generalized additive model[J]. Journal of Beijing Forestry University, 2014, 36(5): 22−32.
[32] 欧强新, 雷相东, 沈琛琛, 等. 基于随机森林算法的落叶松–云冷杉混交林单木胸径生长预测[J]. 北京林业大学学报, 2019, 41(9): 9−19. Ou Q X, Lei X D, Shen C C, et al. Individual tree DBH growth prediction of larch-spruce-fir mixed forests based on random forest algorithm[J]. Journal of Beijing Forestry University, 2019, 41(9): 9−19.
[33] 覃鑫浩. 基于混合效应的兴安落叶松单木断面积生长模型研究[J]. 西北林学院学报, 2022, 37(4): 223−230. Qin X H. Growth model of individual-tree basal area for Larix gmelinii based on mixed effect approach[J]. Journal of Northwest Forestry University, 2022, 37(4): 223−230.
[34] 杜志, 陈振雄, 孟京辉, 等. 基于混合效应的马尾松单木断面积预估模型[J]. 中南林业科技大学学报, 2020, 40(9): 33−40. Du Z, Chen Z X, Meng J H, et al. Prediction model of individual-tree basal area for Pinus massoniana based on mixed effect[J]. Journal of Central South University of Forestry & Technology, 2020, 40(9): 33−40.
[35] Kindermann G E. The development of a simple basal area increment, model[J]. Nature Precedings, 2011, 6: 1−25.
[36] Forrester D. Linking forest growth with stand structure: tree size inequality, tree growth or resource partitioning and the asymmetry of competition[J]. Forest Ecology and Management, 2019, 447: 139−157. doi: 10.1016/j.foreco.2019.05.053
[37] Ni R, Baiketuerhan Y, Zhang C, et al. Analysing structural diversity in two temperate forests in northeastern China[J]. Forest Ecology and Management, 2014, 316: 139−147. doi: 10.1016/j.foreco.2013.10.012
[38] Mcroberts R E, Hahn J T, Hefty G J, et al. Variation in forest inventory field measurements[J]. Canadian Journal of Forest Research, 1994, 24(9): 1766−1770. doi: 10.1139/x94-228
[39] Andreassen K, Tomter S. Basal area growth models for individual trees of Norway spruce, scots pine, birch and other broadleaves in Norway[J]. Forest Ecology and Management, 2003, 180(1): 11−24.
-
期刊类型引用(4)
1. 牟韵洁,高德民,龙腾腾,郭在军,牛海峰. 张家口崇礼区主要植物叶片理化性质及燃烧性分析. 南京林业大学学报(自然科学版). 2024(06): 239-244 .  百度学术
百度学术
2. 王博,韩树文,顾泽,陈锋,白夜,刘晓东. 不同烈度林火对油松林潜在地表火行为的影响. 生态学报. 2023(05): 1812-1821 . 百度学术
3. 韩懂懂,杨光,邸雪颖,李兆国. 兴安落叶松林火烧迹地土壤理化性质驱动因子. 生态学报. 2023(21): 8727-8738 . 百度学术
4. 王博,韩树文,武英达,牛树奎,刘晓东. 辽河源自然保护区油松林火烧迹地林木更新研究. 北京林业大学学报. 2020(04): 41-50 . 本站查看
其他类型引用(6)












计量
- 文章访问数: 340
- HTML全文浏览量: 45
- PDF下载量: 45
- 被引次数: 10
