
Using allometric model and game theory to analyze the genetic regulation mechanism of dynamic growth of Populus tibetica trunk
-
摘要:目的
高度与直径是树木生命历程中的2个重要生长性状,本文通过量化藏川杨主干高度和直径之间的相互作用,探究这2个性状的生长过程和生长模式,揭示藏川杨主干动态生长的遗传调控机制。
方法基于异速生长模型和博弈论构建藏川杨高度与直径性状的动态生长互作微分方程,利用系统作图构建藏川杨主干生长遗传解析的统计模型。进一步以藏川杨自然群体作为研究材料,调查其在温室内的生长动态数据,结合本群体的高通量分子标记数据,开展藏川杨的全基因组基因定位工作。
结果藏川杨主干高度和直径的整体生长曲线符合Logistic生长曲线,拆分整体生长曲线发现,藏川杨主干的直径生长对高度生长具有抑制作用,而高度生长对直径生长具有促进作用。所有藏川杨高度和直径的个体拟合优度 > 0.90,且2个性状拟合的残差均服从随机分布,说明使用广义Lotka-Volterra微分方程来拟合藏川杨高度和直径效果很好。基于功能作图方法共定位到78个显著位点,可注释到52个候选基因。以2号染色体上最显著的SNP为例,对藏川杨高度和直径进行遗传解析,发现该SNP的3种基因型(AA、AC、CC)具有相似的高度–直径相互作用模式。该SNP对整体生长、独立生长和依赖生长发挥了不同方式的遗传效应。对定位到的显著位点进行功能注释,可将这些位点所在的基因分为:与木质素/细胞壁合成相关基因、与生长发育相关基因、与抗病抗逆性相关基因、与光合作用相关基因这4类。
结论结合异速生长模型和博弈理论的基因定位模型,可检测基因如何通过合作或竞争策略来调控藏川杨主干的大小,识别树干动态生长的过程和模式,相关模型也可为其他物种重要性状之间的深层次遗传解析提供借鉴。
Abstract:ObjectiveHeight and diameter are two important growth traits in the life cycle of trees. This paper quantifies the interaction between the stem height and diameter of Populus szechuanica var. tibetica, explores the growth process and patterns of these two traits, and reveals the genetic regulatory mechanism of the dynamic growth of P. szechuanica var. tibetica stems.
MethodWe constructed dynamic growth interaction differential equations for the height and diameter traits of P. szechuanica var. tibetica based on the allometric growth model and game theory, and used system mapping to build a statistical model for genetic analysis of the stem growth of P. szechuanica var. tibetica. Furtherly, a natural population of P. szechuanica var. tibetica was used to investigate the dynamic stem growth in greenhouse. Combined with the high-throughput molecular marker data of this population, the genome-wide gene mapping of Populus tibetica was carried out.
ResultThe overall growth curves of main stem height and diameter of P. szechuanica var. tibetica conformed to the logistic growth curve. By splitting the overall growth curve, it was indicated that the diameter growth of P. szechuanica var. tibetica stems had an inhibitory effect on height growth, while height growth had a promoting effect on diameter growth. The individual fitting goodness of height and diameter of P. szechuanica var. tibetica was R2 > 0.90, and the residuals of the fitting of the two traits followed a random distribution, indicating that using the generalized Lotka-Volterra differential equations to fit the height and diameter of P. szechuanica var. tibetica was effective. Based on the functional plotting method, 78 significant loci were co-located, annotating 52 candidate genes. Taking the most significant SNP on chromosome 2 as an example, genetic analysis of height and diameter of P. szechuanica var. tibetica showed that the three genotypes of this SNP, AA, AC, and CC had similar height-diameter interaction patterns. This SNP exhibited different genetic effects on overall growth, independent growth, and dependent growth. Functional annotation of the significant loci identified genes related to lignin/cell wall synthesis, growth and development, disease resistance, and photosynthesis.
ConclusionA gene localization model combining allometric growth model and game theory can detect how genes regulate the size of the trunk of Populus szechuanica var. tibetica through cooperative or competitive strategies, identify the process and pattern of dynamic trunk growth, and provide reference for deep genetic analysis of important traits in other species.
-
碳气凝胶(CA)是一种轻质纳米多孔非晶固态材料,凭借其优良的物理化学性能和导电性能,例如:比表面积大、孔隙率高、热稳定性好和机械性能优异等,被广泛应用在锂离子电池、超级电容器等电化学器件的理想电极材料、吸附介质、催化剂载体、水处理材料、色谱分离系统等领域[1-5],在与传统无机多孔材料的竞争中显示出极大的优势和应用前景[6-8]。传统碳气凝胶由一些有机化合物(如间苯二酚)先经过溶胶−凝胶法缩聚得到有机水凝胶,再经过干燥过程制备得到有机气凝胶,随后再碳化形成碳气凝胶[9]。在干燥过程中,由于这些碳气凝胶前驱体不能承受液体表面张力引起的毛细作用力,从而导致微孔塌陷[10],因此,必须采用超临界CO2干燥等昂贵、耗时的干燥工艺和严格的干燥条件。为了简化制作工艺,学者们从反应条件、原料等方面进行了相关研究。Zhang等[11]以苯酚−甲醛的酚醛树脂作为碳源,经过蒸发、聚合、碳化等步骤制得碳气凝胶并应用于超级电容器领域,直接碳化避免使用复杂的超临界CO2干燥,但过程耗时较久,且制备所得样品电化学性能一般。
与传统制备方法相比,有研究表明:高盐条件可以很大程度简化碳气凝胶制备过程,且对碳气凝胶性质无不利影响。张璇等[12]利用苯酚等原料在高浓度ZnCl2条件下经溶剂热反应,再在氮气中高温热解得到碳气凝胶,无需特殊干燥方法,简化了干燥过程。近年来,Fechler等[13]提出了一种利用葡萄糖与二元盐混合物在水热碳化过程中制备多孔碳质材料的可行方法。盐作为制孔剂和稳定剂,很容易被水冲掉,从而打开微孔。Huang等[14]在高盐条件下用苯酚单体聚合制备多孔碳气凝胶,真空干燥后直接碳化形成低密度(25 mg/cm3)高比表面积(1 340 m2/g)的酚醛碳气凝胶,盐作为制孔剂和稳定剂,在后续步骤中可用盐酸清洗除去,形成微孔。这种在高盐条件下制备碳气凝胶的方法操作简便、价格低,应用于碳气凝胶制备领域具有显著优势。
传统碳气凝胶制备原料间苯二酚、苯酚等价格昂贵且有一定毒性,而木质素具备与其相似的结构,可代替一部分苯酚、间苯二酚等物质。木质素是一种广泛存在于植物体中的重要有机高分子化合物,为自然界含量第二丰富的聚合物,含碳量超过60%,被认为是最大的芳香烃源,是最具有吸引力的炭材料前体[15-16],由紫丁香基丙烷、愈创木基丙烷、对羟苯基丙烷3种单体构成,由于其具有特殊的酚类结构,可以代替部分苯酚、间苯二酚与甲醛等交联剂发生共聚合反应,形成木质素基酚醛树脂碳气凝胶[17-20]。
本研究利用工业碱木质素与甲醛在高盐条件下直接聚合,制备多孔碳气凝胶,其中盐起制孔和稳定作用,在碳化过程升华或分解除去。用工业碱木质素替代苯酚、间苯二酚等昂贵化学品,降低成本,减小毒性,高盐条件下可以直接碳化,也省去了特殊的干燥过程,操作简便,提供了一种木质素高值化利用的新途径。
1. 材料与方法
1.1 材 料
工业碱木质素(山东龙力生物科技股份有限公司),37% ~ 40%的甲醛溶液、氯化锌、氯化钠、碳酸钠、聚四氟乙烯(PTFE)、乙炔黑均为分析纯(北京化工厂)。
1.2 木质素高盐模板碳气凝胶的制备
将2 g工业碱木质素和x g盐混合均匀,加入1.5 mL甲醛,搅拌成黏稠浆状,转移到反应釜中,160 ℃反应2 h,得到一系列LCA/ZnCl2-x、LCA/Na2CO3-x、LCA/NaCl-x的木质素碳气凝胶前驱体。将得到的一系列碳气凝胶前驱体置于样品舟,在通氮气保护的管式炉中,以3 ℃/min的升温速率升温至900 ℃,保温3 h进行碳化,自然冷却后取出,磨粉,浸泡于盐酸中,搅拌后静置24 h,重复该洗涤步骤3次后用水洗涤2次,干燥后得到木质素高盐模板碳气凝胶,进行后续检测。
1.3 样品的结构和理化性质表征
将木质素高盐模板碳气凝胶样品用导电胶固定在样品台上,经喷金处理后采用扫描电镜(Hitachi SU-8010)观察样品表面形貌;用X射线多晶衍射仪(Bruker D8 ADVANCE x-ray)对样品的结构和种类进行表征,入射激光波长0.541 78 nm,扫描范围为10° ~ 60°;将100 mg左右样品在脱气站于300 ℃下脱气3 h后,采用比表面积孔径分布仪(Autosorb-iQ2-MP)对样品的比表面积进行分析。
1.4 电化学性能表征
将炭材料、PTFE和乙炔黑按质量比8∶1∶1混合,加入适量乙醇使其混合均匀,将其涂覆于尺寸为1 cm × 1 cm的泡沫镍上,压片制成工作电极,在60 ℃下进行干燥(30 min),然后使其浸没在6 mol/L KOH电解液,并在真空环境下浸泡24 h[21]。
电化学性能测试采用三电极系统,分别以铂片为对电极,以Hg/HgO为参比电极,以制备的碳气凝胶电极材料为工作电极,以6 mol/L KOH为电解液,采用瑞士万通电化学工作站进行循环伏安(CV)、恒流充放电(GCD)和开位电路阻抗(EIS)测试。循环伏安测试的电压区间为−1 ~ 0 V,扫描速度为2 ~ 50 mV/s;恒流充放电在−1 ~ 0 V的电压区间进行,电流密度0.2 ~ 10 A/g,开位电路阻抗在开路电压下进行,频率区间为10− 2 ~ 104 Hz。
2. 结果与讨论
2.1 炭材料的形貌分析和结构表征
2.1.1 比表面积分析
表1是不同盐模板木质素碳气凝胶的比表面积数据。从表1可以看出:不同盐模板样品的比表面积数值大约有一个数量级的差距,LCA/NaCl-4比表面积为6 m2/g,LCA/Na2CO3-4比表面积为75 m2/g,LCA/ZnCl2-4比表面积为711 m2/g,以ZnCl2为模板的碳气凝胶比表面积最大。以ZnCl2为模板的碳气凝胶,当ZnCl2加入量从LCA/ZnCl2-1增加到LCA/ZnCl2-4时,比表面积从490 m2/g增大到711 m2/g;当增加到LCA/ZnCl2-6时,比表面积减小到616 m2/g。得到和Huang等[14]类似的结论,随着盐加入量的增加,比表面积不会一直增大。可能的原因为ZnCl2加入量过多,导致ZnCl2对碳气凝胶过度蚀刻,或者是有剩余盐未除去而残留在碳气凝胶表面,导致比表面积略减小。因此使比表面积达到最大值的ZnCl2最佳加入量可继续通过实验找出。
表 1 不同盐模板制备的碳气凝胶比表面积Table 1. Specific surface area of carbon aerogels prepared by different salt templates前驱体
Precursor盐模板类型 Type of salt template 工业碱木质素∶盐
(质量比)
Industrial lignin∶salt (mass ratio)比表面积 Specific surface area/(m2·g−1) LCA/ZnCl2-1 ZnCl2 2∶1 490 LCA/ZnCl2-2 ZnCl2 2∶2 719 LCA/ZnCl2-4 ZnCl2 2∶4 711 LCA/ZnCl2-6 ZnCl2 2∶6 616 LCA/Na2CO3-4 Na2CO3 2∶4 75 LCA/NaCl-4 NaCl 2∶4 6 2.1.2 SEM分析
图1为LCA/NaCl-4、LCA/Na2CO3-4、LCA/ZnCl2-4的SEM图。从图1a可以观察到LCA/NaCl-4表面致密无明显孔洞,并能观察到NaCl结晶。NaCl使反应物发生盐析现象,苯酚类物质悬浮在表面,NaCl沉淀在底部,最终上层形成一种非常坚硬的有机碳,NaCl未对碳气凝胶起到制孔作用。图1b中可以观察到明显孔洞,但Na2CO3的蚀刻作用仍不明显,制孔能力较弱。LCA/ZnCl2-4表面十分粗糙,孔隙多(图1c),另一区域放大后可以观察到碳气凝胶状(图1d)。因此,推测3种盐的制孔能力ZnCl2 > Na2CO3 > NaCl,且只有以ZnCl2为模板的样品可以形成碳气凝胶,ZnCl2为发泡剂、制孔剂、稳定剂。通常非常精细的有机气凝胶从一开始就很坚固,致密前驱体的同时碳化和发泡,使其结构在整个成碳过程中十分稳定,所获得的坚固LCA/ZnCl2-x可以承受极性液体的毛细管压力,因此不再需要使用冷冻干燥或超临界干燥来避免毛细管力引起的微孔塌陷[13]。
![]() 图 1 不同盐模板制备的碳气凝胶扫描电镜图Figure 1. SEM diagrams of carbon aerogels prepared by different salt templates
图 1 不同盐模板制备的碳气凝胶扫描电镜图Figure 1. SEM diagrams of carbon aerogels prepared by different salt templates2.1.3 XRD分析
图2为不同盐模板制备的碳气凝胶和工业碱木质素的XRD图谱。从图2可以观察到:LCA/ZnCl2-4、LCA/Na2CO3-4、LCA/NaCl-4这3个样品的谱图在24°和44°附近存在两个明显且较宽的衍射峰,对应于石墨的(002)面和(101)面,表明得到的产物以无定型碳为主。其中LCA/ZnCl2-4的衍射峰强度较大,表明该碳气凝胶具备更好的石墨化程度和优良的电化学储能潜力。32°和46°处两个尖锐的衍射峰在工业碱木质素和碳气凝胶产物中均存在,推测为工业碱木质素原料中难除去的固有杂质[22]。
![]() 图 2 不同盐模板制备的碳气凝胶和碱木质素XRD图Figure 2. XRD diagrams of carbon aerogels prepared by different salt templates and alkali lignin
图 2 不同盐模板制备的碳气凝胶和碱木质素XRD图Figure 2. XRD diagrams of carbon aerogels prepared by different salt templates and alkali lignin2.2 电化学性能分析
2.2.1 循环伏安法测试
图3为不同盐模板的碳气凝胶电极在2 mV/s的扫描速度下的循环伏安曲线,3条样品电极的循环伏安曲线都近似矩形,说明该材料的电容具有双电层电容性质。以ZnCl2为模板的碳气凝胶电极循环伏安曲线的封闭面积最大,可以推测以ZnCl2为模板的碳气凝胶电极的比电容远大于以Na2CO3或NaCl为模板的碳气凝胶电极。
![]() 图 3 不同盐模板的碳气凝胶样品在2 mV/s时循环伏安测试曲线Figure 3. CV curves of carbon aerogel samples with different salt templates at the scan rate of 2 mV/s
图 3 不同盐模板的碳气凝胶样品在2 mV/s时循环伏安测试曲线Figure 3. CV curves of carbon aerogel samples with different salt templates at the scan rate of 2 mV/s图4为ZnCl2加入量不同的碳气凝胶电极在2 mV/s扫描速度下的循环伏安曲线,均为近似矩形的双电层电容。LCA/ZnCl2-2和LCA/ZnCl2-4曲线封闭面积相近,比电容大小也近似相同,符合同种原料比表面积增大比电容也增大的规律。LCA/ZnCl2-6曲线封闭面积略增大,可能是LCA/ZnCl2-6中ZnCl2过量,残留的ZnCl2使得比表面积减小,但是KOH电解液中,经过循环伏安预循环和活化过程使得残留的ZnCl2被冲掉,LCA/ZnCl2-6在电解液中电解质离子所能触及的实际比表面积大于前面几个样品。
![]() 图 4 ZnCl2盐模板不同盐加入量的碳气凝胶样品在2 mV/s时循环伏安测试曲线Figure 4. CV curves of samples with ZnCl2 template at the scan rate of 2 mV/s
图 4 ZnCl2盐模板不同盐加入量的碳气凝胶样品在2 mV/s时循环伏安测试曲线Figure 4. CV curves of samples with ZnCl2 template at the scan rate of 2 mV/s2.2.2 恒流充放电测试
图5为不同盐模板的碳气凝胶样品在0.2 A/g电流密度下的恒流充放电曲线。从图5可以看出:3个样品的恒流充放电曲线都为近似对称的等腰三角形形状,表现出双电层电容的充放电性质,表明3种样品电极的充放电都具备良好的化学可逆性。以ZnCl2为模板的碳气凝胶电极放电时间最长,表明该碳气凝胶具有更大的比电容。3条充放电曲线均出现轻微的圆弧,是由于充放电过程中发生少量氧化还原反应,产生了赝电容效应[15, 22]。图6为LCA/ZnCl2在不同电流密度下的恒流充放电曲线,随电流密度的增加,曲线仍保持近似等腰三角形,表明以ZnCl2为模板的碳气凝胶电极具备良好的化学可逆性。
![]() 图 5 不同盐模板的碳气凝胶样品在0.2 A/g下恒流充放电曲线Figure 5. GCD curves of carbon aerogel samples with different salt templates at 0.2 A/g
图 5 不同盐模板的碳气凝胶样品在0.2 A/g下恒流充放电曲线Figure 5. GCD curves of carbon aerogel samples with different salt templates at 0.2 A/g![]() 图 6 LCA/ZnCl2-4在不同电流密度下恒流充放电曲线Figure 6. GCD curves of LCA/ZnCl2-4 under different current densities
图 6 LCA/ZnCl2-4在不同电流密度下恒流充放电曲线Figure 6. GCD curves of LCA/ZnCl2-4 under different current densities为了进一步探索不同盐模板对倍率性能的影响,测试了3个样品在不同电流密度下的恒流充放电情况,将不同样品的电流密度与充放电时间换算成比电容(c)作图。计算方法为
c=I⋅TU ,I为电流密度(A/g),T为放电时间(s),U为放电电压(V)。图7为不同样品的倍率性能曲线。从图7可以看出:随电流密度的增加,所有样品的比电容都有小幅度的减小,但LCA/ZnCl2-4电极仍表现出比较好的倍率性能。LCA/ZnCl2-4在0.2 A/g的电流密度下,比电容达到124 F/g,当电流密度增加到10 A/g时,比电容保持在60 F/g,电容保持率为48%。LCA/Na2CO3-4在0.2 A/g的电流密度下,比电容为14 F/g,当电流密度增加到10 A/g时,比电容为10 F/g。LCA/NaCl-4在0.2 A/g的电流密度下,比电容为20 F/g,当电流密度增加到10 A/g时,比电容为8 F/g。以ZnCl2为模板的碳气凝胶电极具有更大的比电容和更优秀的倍率性能。2.2.3 开位电路阻抗测试
图8为不同盐模板碳气凝胶电极的开位电路阻抗曲线。3条阻抗曲线均由高频段的半圆弧和低频段的线性部分两部分组成,高频区圆弧与实轴阻抗上的截距表示内阻或等效串联电阻,表示离子在电极和电解液界面之间的迁移。以ZnCl2、NaCl、Na2CO3为盐模板的碳气凝胶的内阻分别约为0.1、11、5 Ω;中高频段的半圆弧直径对应电荷转移电阻。3种材料电荷转移电阻都很小,以ZnCl2为盐模板的碳气凝胶约为0.5 Ω;低频区近似垂直的直线表示离子在电极或电解液界面上的扩散阻力[15, 23-24]。3种盐模板中,以ZnCl2为模板的LCA/ZnCl2-4电极拥有较短的45°斜线,而且低频段直线近似垂直,所以阻抗性能更为优异,能保证电解液离子的快速传输和低电阻的电荷扩散[25-26]。
![]() 图 8 不同碳气凝胶样品的开位电路阻抗测试曲线Figure 8. Open circuit impedance test curves of different carbon aerogel samples
图 8 不同碳气凝胶样品的开位电路阻抗测试曲线Figure 8. Open circuit impedance test curves of different carbon aerogel samples3. 结 论
(1)工业碱木质素和甲醛混合后可分别与ZnCl2、Na2CO3、NaCl在水热条件下可直接聚合,再将碳气凝胶前驱体高温碳化得到碳气凝胶,其中盐作为制孔剂和稳定剂,致密前驱体的同时碳化和发泡过程使其结构在整个碳化过程中保持稳定,得到的坚固LCA可以承受极性液体的毛细管压力,允许在环境或真空条件下干燥,不需使用特殊干燥手段。
(2)3种盐模板中,ZnCl2的制孔能力最佳,可以制备出比表面积大,电化学性能优异的多孔碳气凝胶。其中ZnCl2在聚合过程中,就已经在多孔微观结构的形成中起着关键作用[14],ZnCl2参与制孔,并且可以在碳化过程中转变为ZnO后蒸发,完成模板作用,碳气凝胶骨架中碳原子主要为sp3杂化,得到以无定型碳为主的碳气凝胶。以LCA/ZnCl2-4为例,比表面积711 m2/g,在6 mol/L的 KOH电解液中,0.2 A/g的电流密度下,比电容达到124 F/g;在10 A/g的高电流密度下,比电容维持在60 F/g,电容保持率约为48%。
(3)高盐条件中的“高”是一个相对含义,是相对其他实验中盐的添加量和本实验中其他反应物的加入量而言。Huang等[14]的研究和本实验结果证明:随着盐加入量的增加,比表面积不会一直增大。在本实验中,LCA/ZnCl2-x(x最大为6)随着盐的增加,材料的比电容等电化学性能却更加优异。所以后续的研究可以继续探究使碳气凝胶综合性能更加优异的盐加入量,或者根据碳气凝胶的用途来决定盐的加入量。
综上,以工业碱木质素为原料制备碳气凝胶有以下优点:工业碱木质素代替有毒的苯酚、间苯二酚等酚类化学品,绿色环保;工业碱木质素价格远低于昂贵的苯酚等原料,可降低成本;工业碱木质素可直接参与反应,且制备过程无需冷冻干燥,操作简单;产物以无定形碳为主,比表面积大,电化学性能优异等。因此,木质素高盐模板碳气凝胶制备的电极材料具有优良的性能,在工业化利用方面具有广阔前景。
-
![]()
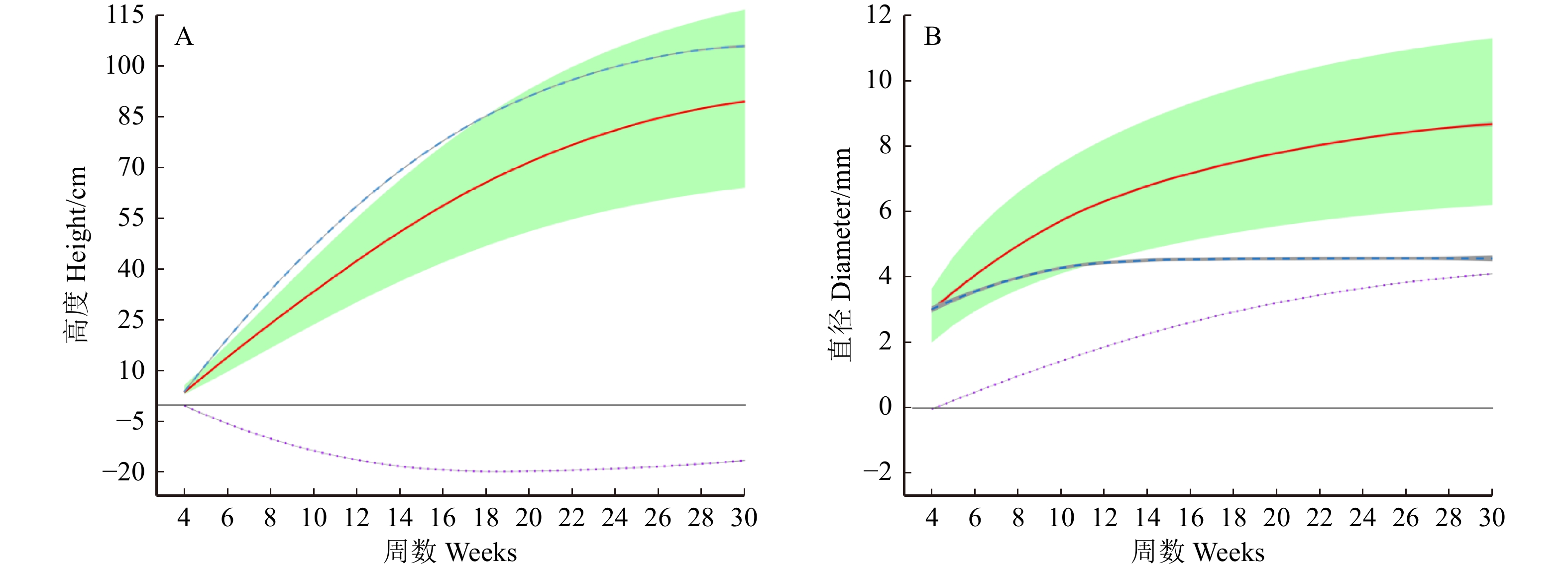
图 1 藏川杨主干高度(A)和直径(B)随时间变化的生长曲线
基因型变异的分布和程度以阴影显示,2个生长性状的整体生长曲线用实线表示。每条整体生长曲线是独立生长曲线(虚线)和交互生长曲线(点线)的总和。The distribution and degree of genotype variation are shown in shadows, and the overall growth curves of the two growth traits are represented by solid lines. Each overall growth curve is the sum of independent growth curves (dashed lines) and interactive growth curves (dotted lines).
Figure 1. Growth curves of stem height (A) and diameter (B) of P. szechuanica var. tibetica with time
![]()
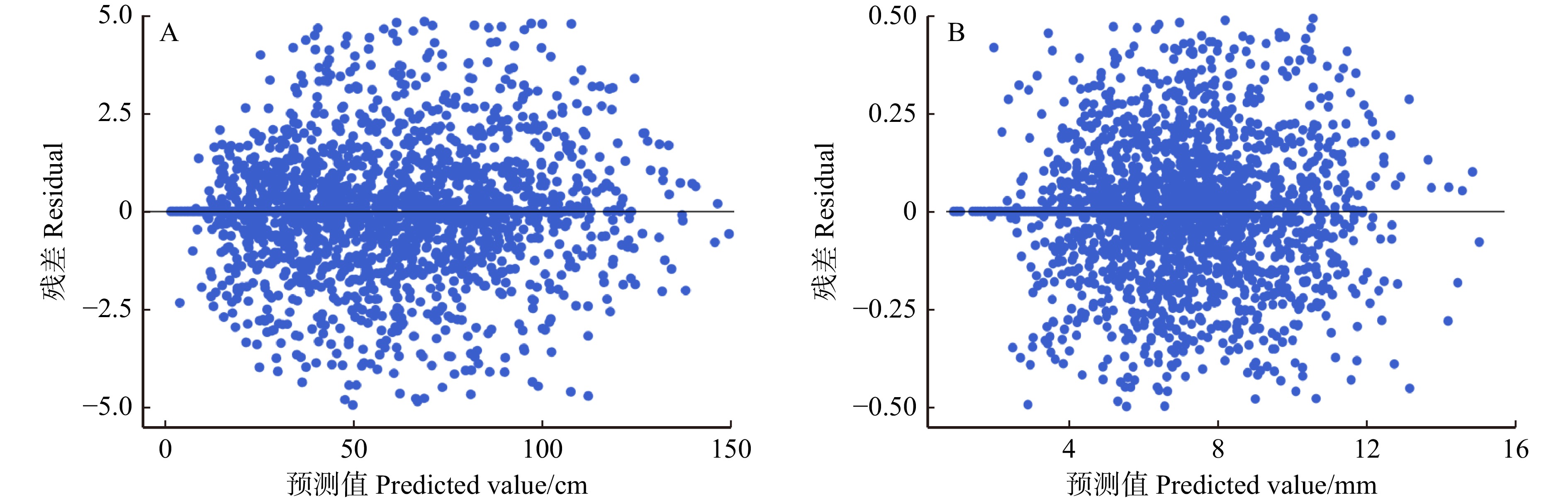
图 2 广义Lotka-Volterra微分方程组估计藏川杨主干高度(A)和直径(B)的残差分布
Figure 2. Residual distribution of stem height (A) and diameter (B) of Populus szechuanica var. tibetica estimated by generalized Lotka-Volterra differential equations
![]()
图 3 藏川杨全基因组SNP的曼哈顿图
水平虚线为临界阈值。Horizontal dashed line represents critical threshold.
Figure 3. Manhattan map of SNP in the whole genome of P. szechuanica var. tibetica
![]()
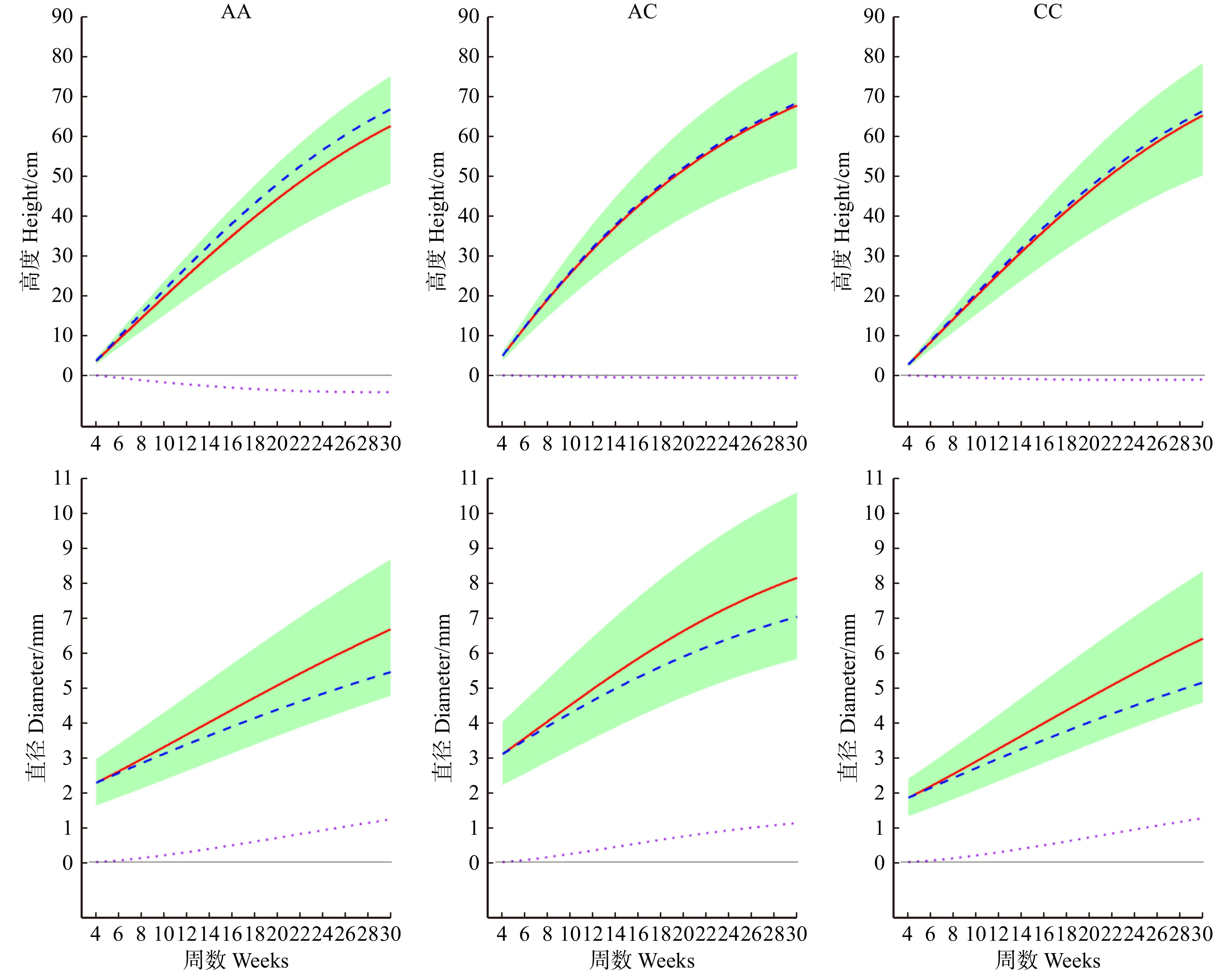
图 4 关于藏川杨GameQ.2位点的3种基因型的高度和直径的生长曲线
每个性状的整体生长(实线)由独立生长(虚线)和交互生长(点线)贡献。AA,AC和CC为3种基因型。The overall growth (solid line) of each trait is contributed by independent growth (dashed line) and interactive growth (dotted line). AA, AC, and CC are three genotypes.
Figure 4. Growth curves of height and diameter for three genotypes of GameQ.2 locus in P. szechuanica var. tibetica
![]()
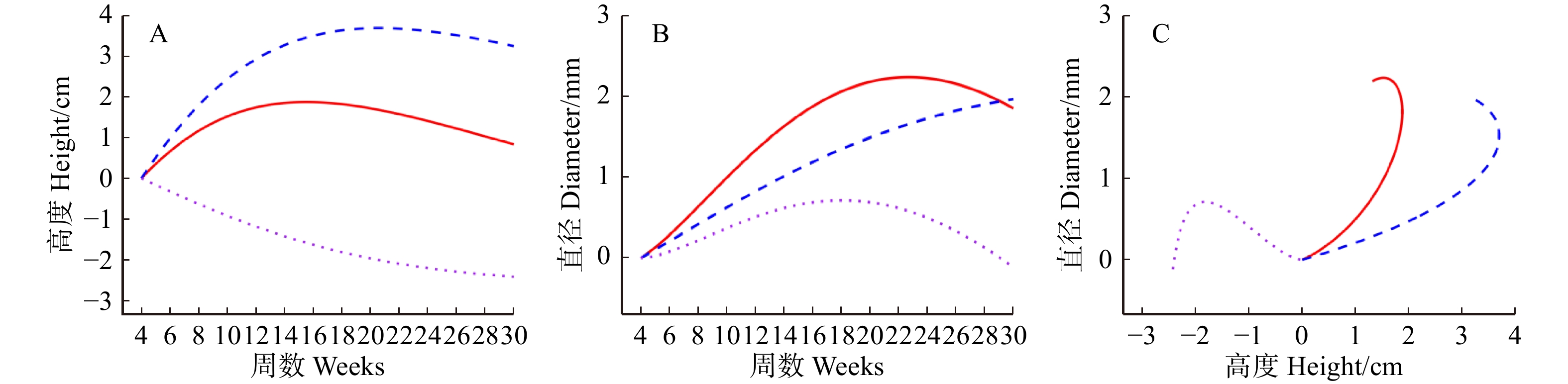
图 5 藏川杨GameQ.2位点上高度(A)、直径(B)随时间变化的遗传效应曲线以及二者之间的遗传效应曲线(C)
实线、虚线、点线分别表示整体生长、独立生长和交互生长的遗传效应。The solid line, dashed line, and dotted line represent the genetic effects of overall growth, independent growth, and interactive growth, respectively.
Figure 5. Genetic effect curves of height (A) and diameter (B) with time at the GameQ.2 of P. szechuanica var. tibetica, as well as the genetic effect curve between the two traits (C)
![]()
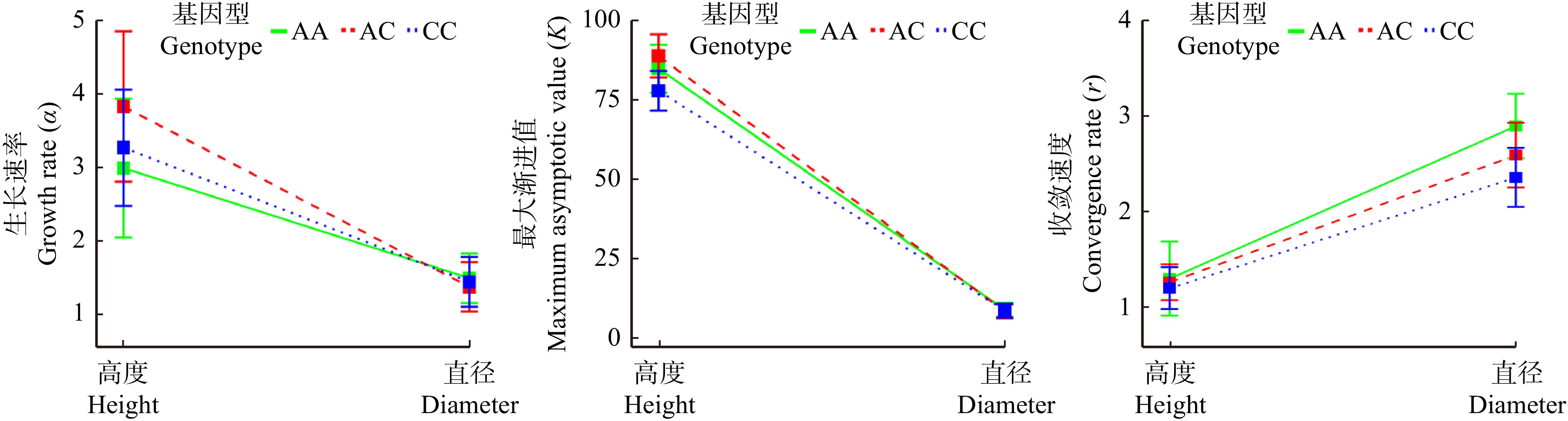
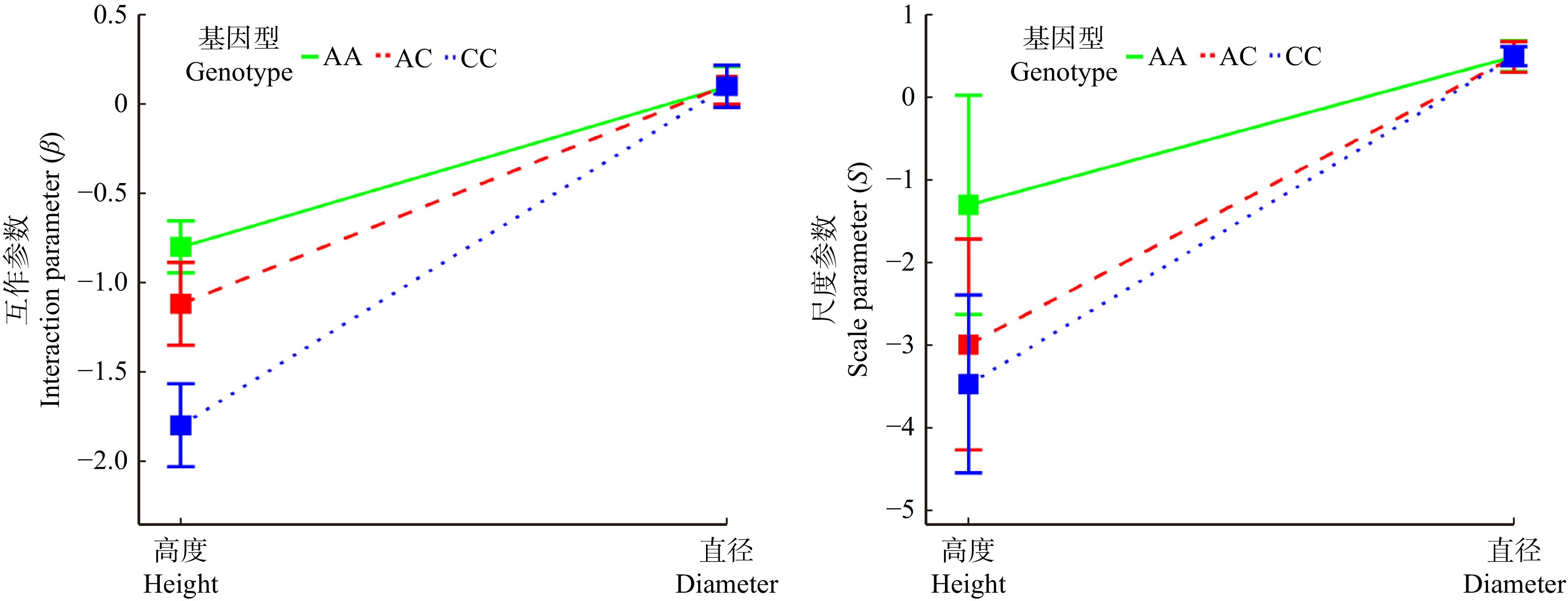
图 6 GameQ.2位点上不同基因型高度和直径的独立生长模式参数
Figure 6. Independent growth pattern parameters of different genotypes with different heights and diameters at GameQ.2 locus
![]()
图 7 GameQ.2位点上不同基因型高度和直径的交互生长模式参数
Figure 7. Interactive growth pattern parameters of different genotype heights and diameters at GameQ.2
表 1 200株藏川杨主干高度和直径的拟合优度(R2)统计结果
Table 1 Goodness-of-fit (R2) statistical results of stem height and diameter of 200 P. szechuanica var. tibetica plants
表型
PhenotypeR2 > 0.98的株数(百分比)
Plant number with
R2 > 0.98 (percentage)R2 > 0.95的株数(百分比)
Plant number with
R2 > 0.95 (percentage)R2 > 0.90的株数(百分比)
Plant number with
R2 > 0.90 (percentage)高度 Height 192(96.0%) 198(99.0%) 200(100.0%) 直径 Diameter 158(79.0%) 191(95.5%) 200(100.0%)  下载: 导出CSV
下载: 导出CSV
表 2 藏川杨显著位点的基因注释
Table 2 Gene annotation of significant sites in P. szechuanica var. tibetica
类别
Category染色体号
Chromosome No.基因编号
Gene ID基因名称
Gene name木质素/细胞壁合成相关基因
Lignin/cell wall synthesis related gene3 LOC18097081 内切葡聚糖酶25 Endoglucanase 25(EG25) 5 LOC18098789 UDP-葡糖醛酸:木聚糖α-葡糖醛酸基转移酶5
UDP-glucuronate: xylan alpha-glucuronosyltransferase 5(GUX5)10 LOC18102703 UDP-糖基转移酶76B1 UDP-glycosyltransferase 76B1(UGT76B1) 13 LOC18104206 LOB结构域蛋白15 LOB domain-containing protein 15(LBD15) 14 LOC7466295 可能的半乳糖醛酸基转移酶15
Probable galacturonosyltransferase 15(GAUT15)19 LOC18108359 蛋白质 SOMBRERO Protein SOMBRERO(SMB) 生长发育相关基因
Growth and development
related gene1 LOC18109358 嘌呤霉素敏感的氨肽酶 Puromycin-sensitive aminopeptidase(PSA) 4 LOC7470186 花同源异型蛋白基因 AGAMOUS Floral homeotic protein AGAMOUS(AG) 8 LOC7457804 E3泛素蛋白连接酶 RIE1 E3 ubiquitin protein ligase RIE1(RIE1) 15 LOC7457575 双特异性蛋白磷酸酶 PHS1 Dual specificity protein phosphatase PHS1(PHS1) 抗病抗逆性相关基因
Disease resistance and stress
resistance related gene2 LOC7479726 转录因子 bHLH110 Transcription factor bHLH110(BHLH110) 4 LOC18097671 钙/钙调蛋白依赖性蛋白激酶激酶 2
Calcium/calmodulin-regulated receptor-like kinase 2(CRLK2)13 LOC7489694 植物半胱氨酸氧化酶2 Plant cysteine oxidase 2(PCO2) 15 LOC7457571 转录因子 bHLH137 Transcription factor bHLH137(BHLH137) 18 LOC7479118 类增强抗病蛋白2
Protein ENHANCED DISEASE RESISTANCE 2-like(EDR2L)光合作用相关基因
Photosynthesis related gene1 LOC18095847 N-α-乙酰基转移酶 MAK3 N-alpha-acetyltransferase MAK3(MAK3) 4 LOC18097764 叶绿体亚铁螯合酶2 Ferrochelatase-2, chloroplastic(FC2) 其他 Qthers 2 LOC7471975 类 DMR6氧合酶1蛋 Protein DMR6-LIKE OXYGENASE 1(DLO1) 3 LOC7479194 染色体结构维持蛋白2-1
Structural maintenance of chromosomes protein 2-1(SMC2-1)5 LOC18099665 细胞色素 P450 94A1 Cytochrome P450 94A1(CYP94A1) 6 LOC7492290 叶绿体铜转运 ATP酶 PAA2
Copper-transporting ATPase PAA2, chloroplastic(PAA2)6 LOC7471545 富含脯氨酸的类受体蛋白激酶 PERK1
Proline-rich receptor-like protein kinase PERK1(PERK1)
下载: 导出CSV
-
[1] Wang D, Chen Y, Li W, et al. Vascular cambium: the source of wood formation[J/OL]. Frontiers in Plant Science, 2021, 12: 700928[2022−04−21]. https://doi.org/10.3389/fpls.2021.700928.
[2] Bartholome J, Salmon F, Vigneron P, et al. Plasticity of primary and secondary growth dynamics in Eucalyptus hybrids: a quantitative genetics and QTL mapping perspective[J]. BMC Plant Biology, 2013, 13: 120−133. doi: 10.1186/1471-2229-13-120
[3] Gorgens E B, Nunes M H, Jackson T, et al. Resource availability and disturbance shape maximum tree height across the Amazon[J]. Global Change Biology, 2021, 27(1): 177−189. doi: 10.1111/gcb.15423
[4] Kocillari L, Olson M E, Suweis S, et al. The widened pipe model of plant hydraulic evolution[J/OL]. Proceedings of the National Academy of Sciences of the United States of America, 2021, 118(22): e2100314118[2022−04−16]. https://doi.org/10.1073/pnas.2100314118.
[5] 李昕, 李姗, 邓丽萍, 等. 楸树木质部水分输导组织构造特征的轴向变化[J]. 北京林业大学学报, 2020, 42(1): 27−34. doi: 10.12171/j.1000-1522.20190238 Li X, Li S, Deng L P, et al. Axial variation of characteristics of water conductive tissue in xylem of Catalpa bungei[J]. Journal of Beijing Forestry University, 2020, 42(1): 27−34. doi: 10.12171/j.1000-1522.20190238
[6] Franceschini T, Martin-Ducup O, Schneider R. Allometric exponents as a tool to study the influence of climate on the trade-off between primary and secondary growth in major north-eastern American tree species[J]. Annals of Botany, 2016, 117(4): 551−563. doi: 10.1093/aob/mcw003
[7] Dharmawardhana P, Brunner A, Strauss S. Genome-wide transcriptome analysis of the transition from primary to secondary stem development in Populus trichocarpa[J]. BMC Genomics, 2010, 11(1): 150−168. doi: 10.1186/1471-2164-11-150
[8] Xu C, Shen Y, He F, et al. Auxin-mediated Aux/IAA-ARF-HB signaling cascade regulates secondary xylem development in Populus[J]. New Phytologist, 2019, 222(2): 752−767. doi: 10.1111/nph.15658
[9] Zhao B G, Li G, Wang Y F, et al. PdeHCA2 affects biomass in Populus by regulating plant architecture, the transition from primary to secondary growth, and photosynthesis[J]. Planta, 2022, 255(5): 101−118. doi: 10.1007/s00425-022-03883-6
[10] Huang S H, Titus S J, Wiens D P. Comparison of nonlinear height-diameter functions for major Alberta tree species[J]. Canadian Journal of Forest Research, 1992, 22(9): 1297−1304. doi: 10.1139/x92-172
[11] Avalos G, Gei M, Rios L D, et al. Scaling of stem diameter and height allometry in 14 neotropical palm species of different forest strata[J]. Oecologia, 2019, 190(4): 757−767. doi: 10.1007/s00442-019-04452-7
[12] Sullivan M, Lewis S L, Hubau W, et al. Field methods for sampling tree height for tropical forest biomass estimation[J]. Methods in Ecology and Evolution, 2018, 9(5): 1179−1189. doi: 10.1111/2041-210X.12962
[13] 曾伟生, 杨学云, 陈新云. 单木和林分水平一元与二元材积模型的预估精度对比[J]. 中南林业调查规划, 2017, 36(4): 1−6. Zeng W S, Yang X Y, Chen X Y. Comparison on prediction precision of one-variable and two-variable volume models on tree-level and stand-level[J]. Central South Forest Inventory and Planning, 2017, 36(4): 1−6.
[14] Osada N, Tateno R, Mori A, et al. Changes in crown development patterns and current-year shoot structure with light environment and tree height in Fagus crenata (Fagaceae)[J]. American Journal of Botany, 2004, 91(12): 1981−1989. doi: 10.3732/ajb.91.12.1981
[15] Vizcaino-Palomar N, Ibanez I, Gonzalez-Martinez S C, et al. Adaptation and plasticity in aboveground allometry variation of four pine species along environmental gradients[J]. Ecology and Evolution, 2016, 6(21): 7561−7573. doi: 10.1002/ece3.2153
[16] Daniel S F, Mark W. Plant height and evolutionary games[J]. Trends in Ecology and Evolution, 2003, 18(7): 337−343. doi: 10.1016/S0169-5347(03)00061-2
[17] McNickle G G, Dybzinski R. Game theory and plant ecology[J]. Ecology Letters, 2013, 16(4): 545−555. doi: 10.1111/ele.12071
[18] Wu R, Cao J, Huang Z, et al. Systems mapping: how to improve the genetic mapping of complex traits through design principles of biological systems[J]. BMC Systems Biology, 2011, 5: 84−94. doi: 10.1186/1752-0509-5-84
[19] Chen X, Ou C H. The complete chloroplast genome sequence of Populus szechuanica, a medicinal plant for child anesthesia application[J]. Mitochondrial DNA B Resources, 2021, 6(2): 696−698. doi: 10.1080/23802359.2021.1882907
[20] Raj A, Stephens M, Pritchard J K. fastSTRUCTURE: variational inference of population structure in large SNP data sets[J]. Genetics, 2014, 197(2): 573−589. doi: 10.1534/genetics.114.164350
[21] Li H, Durbin R. Fast and accurate short read alignment with Burrows-Wheeler transform[J]. Bioinformatics, 2009, 25(14): 1754−1760. doi: 10.1093/bioinformatics/btp324
[22] Li H, Handsaker B, Wysoker A, et al. The sequence alignment/map format and SAMtools[J]. Bioinformatics, 2009, 25(16): 2078−2079. doi: 10.1093/bioinformatics/btp352
[23] Danecek P, Auton A, Abecasis G, et al. The variant call format and VCFtools[J]. Bioinformatics, 2011, 27(15): 2156−2158. doi: 10.1093/bioinformatics/btr330
[24] Lotka A J. Analytical note on certain rhythmic relations in organic systems[J]. Proceedings of the National Academy of Sciences of the United States of America, 1920, 6(7): 410−415.
[25] West G B, Brown J H, Enquist B J. A general model for ontogenetic growth[J]. Nature, 2001, 413(6856): 628−631. doi: 10.1038/35098076
[26] Ma C X, Casella G, Wu R. Functional mapping of quantitative trait loci underlying the character process: a theoretical framework[J]. Genetics, 2002, 161(4): 1751−1762. doi: 10.1093/genetics/161.4.1751
[27] Hairer E, Iserles A, Sanz-Serna J M. Equilibria of Runge-Kutta methods[J]. Numerische Mathematik, 1900, 58: 243−254.
[28] Zhao W, Hou W, Littell R C, et al. Structured antedependence models for functional mapping of multiple longitudinal traits[J]. Statistical Applications in Genetics and Molecular Biology, 2005, 4(1): 1−28.
[29] Bland J M, Altman D G. Multiple significance tests: the Bonferroni method[J]. British Medical Journal, 1995, 310(6973): 170. doi: 10.1136/bmj.310.6973.170
[30] Piermattei A, von Arx G, Avanzi C, et al. Functional relationships of wood anatomical traits in Norway spruce [J/OL]. Frontiers in Plant Science, 2020, 11: 683[2022−04−18]. https://doi.org/10.3389/fpls.2020.00683.
[31] Akiyoshi N, Ihara A, Matsumoto T, et al. Functional analysis of poplar sombrero-type NAC transcription factors yields a strategy to modify woody cell wall properties[J]. Plant and Cell Physiology, 2021, 62(12): 1963−1974. doi: 10.1093/pcp/pcab102
[32] Rennie E A, Hansen S F, Baidoo E E, et al. Three members of the Arabidopsis glycosyltransferase family 8 are xylan glucuronosyltransferases[J]. Plant Physiology, 2012, 159(4): 1408−1417. doi: 10.1104/pp.112.200964
[33] Ohashi-Ito K, Iwamoto K, Fukuda H. LOB domain-containing protein 15 positively regulates expression of VND7, a master regulator of tracheary elements[J]. Plant and Cell Physiology, 2018, 59(5): 989−996. doi: 10.1093/pcp/pcy036
[34] Sun X, Feng Z, Meng L, et al. Arabidopsis ASL11/LBD15 is involved in shoot apical meristem development and regulates WUS expression[J]. Planta, 2013, 237(5): 1367−1378. doi: 10.1007/s00425-013-1844-x
-
期刊类型引用(3)
1. 张耀,李树锋,付鹏飞. 纤维素碳气凝胶的制备、性质及应用. 化工新型材料. 2023(07): 8-11+20 .  百度学术
百度学术
2. 刘沛静,辛福恩. 石墨烯/δ-MnO_2复合材料及其超级电容器性能研究. 电子制作. 2021(08): 87-88 . 百度学术
3. 陈浩伟,余先纯,张传艳,李銮玉,孙德林,郝晓峰. 木质素基模板炭的制备及电化学性能. 材料导报. 2021(24): 24164-24171 . 百度学术
其他类型引用(7)











计量
- 文章访问数: 408
- HTML全文浏览量: 86
- PDF下载量: 66
- 被引次数: 10
