
Characteristics and regeneration potential of soil seed banks in green space of urban parks in Beijing
-
摘要:目的
作为潜在的植物群落,土壤种子库在一定程度上代表地上植被的更新演替潜力,研究城市公园绿地土壤种子库特征及其对地上植被和土壤因子的响应,为近自然城市公园的营造提供理论依据。
方法以北京市六环内公园绿地为对象,将其划分为综合公园、郊野公园、文化遗址公园、社区公园4种类型。在随机选取的75个公园内,采用样方法进行地上植被调查,随机法进行土壤采样,土壤种子库采用萌发试验确定,常规方法获取土壤理化指标。
结果(1)北京城市公园土壤种子库共有77种植物,隶属34科74属,以草本植物为主(97.5%),其中菊科物种数最多(16种)。土壤种子库密度在352 ~ 899粒/m2之间,其中菊科种子储量最高,其次为禾本科和豆科,密度最大的物种为马唐(147.11粒/m2),其次为苜蓿(128.40粒/m2)、狗尾草(122.67粒/m2)等。4类城市公园均以一、二年生草本为主,郊野公园的多年生草本相对丰度要高于其他类型公园,其种子库密度显著大于其他类型公园(P < 0.05)。(2)城市公园土壤种子库的Shannon-Wiener多样性指数范围在1.04 ~ 1.46,Simpson优势度指数范围在0.53 ~ 0.62,Patrick丰富度指数范围在4.79 ~ 8.41,Pielou均匀度指数范围在0.74 ~ 0.84,其中郊野公园种子库物种丰富度显著大于其他类型公园(P < 0.05)。(3)相似系数(Sørensen指数)和非度量多维尺度排序分析表明,不同类型公园间土壤种子库、地上植被相似性系数较高,且群落结构差异不显著。4类城市公园土壤种子库与地上植被相似性偏低,相似系数为0.10 ~ 0.18,且土壤种子库与地上植被间群落构成存在明显差异。(4)土壤种子库密度与草本多样性指数、草本物种丰富度指数、土壤铵态氮和非毛管孔隙度呈显著正相关(P < 0.05),与土壤全氮和硝态氮呈显著负相关;土壤种子库物种丰富度指数与土壤全氮和硝态氮呈显著正相关,与草本多样性指数和土壤铵态氮呈显著负相关。土壤种子库均匀度、优势度和多样性指数与灌木物种丰富度指数呈显著正相关,与草本物种丰富度指数呈显著负相关。其中,草本多样性指数和灌木物种丰富度指数对城市公园土壤种子库的密度和物种多样性影响较大。
结论城市公园土壤种子库密度较小,与其对应地上植被间相似性也较低,且以草本植物种子为主;种子库密度和物种多样性主要受地上草本多样性指数和灌木物种丰富度指数影响较大。因此城市公园绿地土壤种子库更新潜力较低,采取人工促进木本植被更新,是营建近自然城市公园的关键。
Abstract:ObjectiveAs a potential plant community, soil seed bank represents the regeneration and succession potential of aboveground vegetation to some extent. To study the characteristics of soil seed banks and their response to aboveground vegetation and soil factors in urban park green spaces can provide theoretical basis for the construction of urban parks near nature.
MethodTaking the green spaces of parks within the Sixth Ring Road of Beijing as the object, they were divided into four types: comprehensive park, country park, cultural heritage park, and community park. In 75 randomly selected parks, aboveground vegetation surveys were carried out by sampling methods and soil sampling was carried out by randomization methods, the soil seed bank was determined by greenhouse germination method, soil physical and chemical indexes were obtained by conventional method.
Result(1) There were 77 species in the soil seed banks of Beijing City Parks, belonging to 34 families and 74 genera. Most of them were herbs (97.5%), with the highest number of species in the Asteraceae (16 species). Density of soil seed bank ranged from 352 to 899 grain/m2, with Asteraceae having the highest seed reserves, followed by Gramineae and Leguminosae. The species with the highest density was Digitaria sanguinalis (147.11 grain/m2), followed by Medicago sativa (128.40 grain/m2), Setaria viridis (122.67 grain/m2) and others. Annal and biennial herbs were dominant in the four types of urban parks. The relative abundance of perennial herbs in country parks was higher than that in other types of parks, and the density of their seed banks was significantly greater than that in other types of parks (P < 0.05). (2) The Shannon-Wiener diversity index of the soil seed banks in urban parks ranged from 1.04 to 1.46, the Simpson dominance index ranged from 0.53 to 0.62, the Patrick richness index ranged from 4.79 to 8.41, and the Pielou evenness index ranged from 0.74 to 0.84, with the species richness of seed banks of country parks significantly greater than that of other types of parks (P < 0.05). (3) Similarity coefficients (Sørensen index) and non-metric multidimensional scaling analysis indicated that the similarity coefficients of soil seed bank and aboveground vegetation among different types of parks were high, and the differences in community structure were not significant. The similarity between soil seed bank and aboveground vegetation in the four types of urban parks was low, with similarity coefficients ranging from 0.10 to 0.18, and there were significant differences in community composition between them. (4) The density of soil seed banks was significantly positively correlated with herb diversity index, herb species richness index, soil ammonium nitrogen, and non-capillary porosity (P < 0.05), and significantly negatively correlated with soil total nitrogen and nitrate nitrogen. The species richness index of soil seed banks was significantly positively correlated with soil total nitrogen and nitrate nitrogen, and significantly negatively correlated with herb diversity index and soil ammonium nitrogen. The soil seed bank evenness, dominance, and diversity indices were significantly positively correlated with shrub species richness index and negatively correlated with herb species richness index. Among them, the herb diversity index and shrub species richness index had a greater impact on the density and species diversity of soil seed bank in urban parks.
ConclusionThe density of soil seed banks in urban parks and the similarity between the soil seed banks and its corresponding aboveground vegetation is low, and the seeds of herb plants are dominated. The density and species diversity of the seed banks are mainly influenced by the diversity index of aboveground herb plants and the richness index of aboveground shrub species. Therefore, the soil seed bank renewal potential of urban parks is low, and artificial promotion of woody vegetation renewal is the key to build a near-natural urban park.
-
城市公园作为城市绿地的重要组成部分[1],园内植被所发挥的净化空气、降温增湿、保护物种多样性等多重生态作用,对调节城市微气候,改善城市生态环境有着重要影响。公园根据自身主导功能(休闲游憩、生态景观、文化科普、防灾避灾等)、服务范畴(服务半径、主要服务对象等)和分布特点可以分为不同类型(综合公园、郊野公园、社区公园、森林公园等)[2−3]。为了进一步探讨公园功能的发挥与公园类型之间的关联,国内外学者针对不同城市公园进行了大量研究,包括园内植被群落结构的差异及其对生物多样性的影响[4−5]、利用景观格局指数研究不同类型公园的服务质量[6]、不同公园在生态、社会效益等方面的差异[7−8]等,但对不同城市公园植被自然更新潜力的研究尚少。植被的自然更新指通过林下植被的自然生长来实现生态环境的维持、恢复和优化[9],对植物种群的繁殖发展、生物多样性的稳定提升以及生态系统功能的充分发挥具有重要影响[10]。而研究城市公园植被自然更新潜力,是城市公园近自然植被建设与低成本管理中最根本和最基础的科学问题。
土壤种子库指存在于土壤上层凋落物和土壤中全部存活种子的总和,是潜在的植物种群,也是地上植被种群基因多样性的储存库[11−12]。因此,土壤种子库是衡量植被自然更新潜力的重要指标[13]。土壤种子库特征与地上植被的组成关系密切[14],地上植被群落是土壤种子库的直接来源,其生长和繁殖情况直接影响着种子库的物种组成以及动态变化[15],研究两者物种组成和群落构成的关系,对植被群落更新演替以及生态系统的恢复重建十分重要。土壤作为种子直接生存的微环境,会影响种子的萌发和分布,进而改变种子库的物种组成[14]。陈颖颖等[16]发现种子库主要受土壤非毛管孔隙度和含水量的影响。李国旗等[17]研究表明土壤pH值和电导率会对种子库物种多样性产生较大影响。卢正宽等[14]指出土壤全氮会增加种子库的密度和物种丰富度,但 Basto等[18]认为过量氮会引起种子库的密度和物种丰富度降低。可见土壤因子对土壤种子库的影响会随研究内容有所改变。研究土壤理化性质和地上植被与土壤种子库之间的关系,有助于了解植被更新过程中影响土壤种子库的主要因子,为公园植被自然更新潜力研究提供理论基础。
以往研究表明,城市化提高了木本植物多样性,但降低了草本和土壤种子库多样性,限制了植物群落未来的更新潜力[19−20],在不同绿地类型中,土壤种子库多样性在森林公园中较高,这与适宜的人为管理模式相关[19]。然而,不同城市公园类型的植被群落更新潜力仍不清楚,探究不同类型城市公园土壤种子库物种构成和多样性有助于进一步理解植物群落的更新潜力,以提供针对性植被可持续发展管理模式和保护措施。此外,扩大城市公园绿地规模,提高绿量,降低维护成本,不仅被认为是应对全球气候变化与快速城市化导致的城市生态环境问题最为有效的基于自然的解决途径,也是实现城市持续管理的重要举措[21]。在城市用地日趋紧张和城市绿地高成本维护的背景下,如何有效利用植被自然更新能力营建与管理城市公园绿地,充分发挥城市公园生态、环境、健康等多种生态服务功能,是当前亟待解决的问题。北京作为世界一线城市,公园数量众多、植被规模较大,部分公园建设历史悠久,能够在一定程度上反映公园植被的更新状况与潜力。因此,本研究以北京市为例,按照综合公园、郊野公园、文化遗址公园、社区公园4种主要类型,通过系统采样、实验室分析测试,系统分析不同类型公园土壤种子库特征及其与地上植被间相似性的差异,进而推断城市公园植被更新潜力,分析制约植被更新的主要因素,以期为城市公园植被多样性保护、近自然营建和低成本维护提供科学依据。
1. 研究区概况与研究方法
1.1 研究区概况
北京位于中国北部、华北平原西北部(115°42′~117°24′E,39°24′~41°36′N),为典型的暖温带半湿润大陆性季风气候,年平均气温为13.8 ℃,多年平均降雨量600 mm,降水年际和年内分配不均匀,全年降水的75%集中在夏季。由于开发历时较长,北京市主城区的天然植被较少,绝大多数是人工植物群落,主要包括温带落叶阔叶林(栎属Quercus、槭属Acer、杨属Populus等)和针叶林(油松Pinus tabuliformis、侧柏Platycladus orientalis、刺柏Juniperus formosana等占优势)[22]。城市公园绿地植被也多为人工栽培,其群落结构主要为乔灌草、乔草、草地等。园内乔木植被主要有毛白杨(P. tomentosa)、油松(Pinus tabuliformis)、栾(Koelreuteria paniculata)等,灌木植被主要有北京丁香(Syringa reticulata)、金银忍冬(Lonicera maackii)、迎春花(Jasminum nudiflorum)等,草本植被主要有沿阶草(Ophiopogon bodinieri)、牛筋草(Eleusine indica)等。
1.2 试验方法
1.2.1 样地设置、植被调查和土壤取样
以城市绿地分类标准[23]为基础,结合北京市公园分类分级管理办法[24]以及管理实际,将北京城区六环以内的城市公园划分为综合公园、郊野公园、文化遗址公园、社区公园4类[2,25](图1和表1)。由于文化遗址公园数量较少,为保证研究结果具有统计学意义,平衡了每类公园的样品数量,在初步拟定每一类公园类型随机选取15个公园的基础上,根据公园实际数量,最终确定综合公园、郊野公园、文化遗址公园、社区公园分别为19个、17个、12个、27个,总共75个公园,进行野外调查与取样。在每个公园内随机设置1 ~ 3个20 m × 20 m的样方,进行乔木调查,当样地不足20 m × 20 m则调整形状使其面积为400 m2,共对201个样地进行了调查。采用手持便携式GPS(4G LTE Real-Time Portable GPS Tracker)记录样地位置、海拔等信息。对乔木层进行每木检尺,记录树种、数量、胸径、树高、生长情况等。在每个乔木样方内按照五点取样法设置5个5 m × 5 m的灌木样方和5个1 m × 1 m的草本样方进行调查。灌木层调查植物物种、数量等;草本层调查物种、盖度等[20]。
![]() 图 1 北京市六环内公园研究样地分布Figure 1. Distribution of study sample plots of parks in the Sixth Ring Road of Beijing表 1 公园分类Table 1. Classification of parks
图 1 北京市六环内公园研究样地分布Figure 1. Distribution of study sample plots of parks in the Sixth Ring Road of Beijing表 1 公园分类Table 1. Classification of parks公园类型
Type of park位置
Location功能特点
Functional characteristics人为管理
Artificial management综合公园
Comprehensive
park中心区
Central zone综合公园一般规模较大;以人工景观为主;具有休闲游憩、生态景观、文化科普、防灾避灾等功能并全面均衡发展;服务半径大
The scale of comprehensive park is generally large, mainly artificial landscape, and has the functions of recreation, ecological landscape, cultural science, disaster prevention and avoidance, and comprehensive and balanced development; the service radius of such park is large管理机制稳定健全,人为干扰强度较大;资源保护措施完善且效果好;绿地养护水平高
The management mechanism is stable and sound, and the intensity of human interference is large; the resource protection measures are perfect and effective; the level of green space maintenance is high文化遗址公园
Cultural
heritage park中心区
Central zone文化遗址公园规模适中;人文与自然景观相结合;具有突出的文化科普和休闲游憩的功能,对非本地游客有重要影响;服务半径适中
The scale of cultural heritage park is moderate, combining humanistic and natural landscapes, with outstanding functions of popularization of culture and recreation, and have an important impact on non-local visitors; the service radius of such park is moderate管理机制稳定健全,人为干扰强度较大;资源保护措施完善且效果好;绿地养护水平高
The management mechanism is stable and sound, and the intensity of human interference is large; the resource protection measures are perfect and effective; the level of green space maintenance is high社区公园
Community park中心区 (居民区附近)
Central zone (near a residential area)社区公园一般规模较小;以人工景观为主;周围居民为主要服务对象,具有休闲放松、生态景观和防灾避灾等功能;服务半径小
The scale of community parks is generally small, mainly artificial landscape, and the surrounding residents as the main service object, with the functions of leisure and relaxation, ecological landscape and disaster prevention and avoidance; the service radius of such park is small管理机制较稳定,人为干扰强度适中;资源保护措施比较完善且效果比较好;绿地养护水平中等
The management mechanism is relatively sound, and the intensity of human interference is moderate; the resource protection measures are sound and the effect is good; the level of green space maintenance is medium郊野公园
Country park近郊
Suburb郊野公园规模较大;以自然景观为主;生态服务功能突出,兼有休憩娱乐、文化科普和防灾减灾等功能;服务半径较大
The scale of country park is large, mainly natural landscape, with outstanding ecological service functions, and functions of leisure and recreation, culture and science popularization, and disaster prevention and mitigation; the service radius of such park is large管理机制有待提升,人为干扰强度较小;资源保护措施有待完善,效果较好;绿地养护水平一般
The management mechanism needs to be improved, and the intensity of human interference is small; the resource protection measures need to be improved and the effect is good; the level of green space maintenance is average注:引自文献[2, 23−25]。Note: cited from reference [2, 23−25]. 土壤取样与地上植被调查同时进行,均在2021年5—7月份完成[14,17,19,26]。为提高种子库采样精确性,采用大数量小样方法[27]进行土壤持久种子库取样。种子多数集中在土壤表层,加之城市环境采样过深会受建筑垃圾影响,因此,每个20 m × 20 m样方内采用直径3.6 cm、深10 cm的土钻随机采取10个0 ~ 10 cm表层土样,并将同一个样地的3个样方所采的30份表层土壤混合,作为该样地的代表土壤种子库土样,最终共获取75份土壤种子库试验样品。在每个样方的土壤种子库取样点周围用环刀随机取3份表层土壤样品,共603份,用于土壤物理性质的测定,同时用土钻(直径5 cm、深20 cm)在每个样方内随机取3份土样,每个样地的样方土样混为一份,共201份,去除植物根系和石块,用于土壤化学性质的测定。
1.2.2 土壤理化性质测定
土壤物理性质测定指标包括土壤含水量、土壤密度、土壤总孔隙度、毛管孔隙度、非毛管孔隙度;土壤化学性质测定指标包括pH值、有机质、全氮、全磷、铵态氮、硝态氮和有效磷。其中,土壤含水量、土壤密度和孔隙度采用烘干法和环刀法测定;土壤pH值采用SX751型便携式测量仪测定;有机质采用重铬酸钾氧化−外加热法测定[28];全氮采用凯氏定氮法测定[29];全磷用氢氧化钠熔融−钼锑抗比色法测定;有效磷、铵态氮和硝态氮用SmartChem450全自动间断化学分析仪(AMS Alliance, KPM Analytics, Italy)测定。
1.2.3 土壤种子库测定
采用萌发试验[14,17,19,28,30]测定土壤种子库的物种组成和数量。试验在北京林业大学苗圃温室大棚(116°20′E,40°00′N)中进行。将采集到的土样带回实验室,去除土壤种子库中的根系、石块等杂物后,将土样放置在底部铺有2 cm厚蛭石土层(蛭石经120 ℃高温处理24 h,以消灭其中有活力的种子)的塑料萌发盆(30 cm × 24 cm × 6 cm)中。为排除外来种子的干扰,试验设置了4个仅装有蛭石的萌发盆作为空白对照。从第一天开始每天09:00和17:00时浇水2次,保证土壤湿润,并控制室温在22 ℃左右[19],保证种子有良好的萌发环境。幼苗萌发出来后,及时进行数量记录和物种鉴定,并拔出鉴定过的幼苗。萌发两个月后,适当翻动土壤,促使休眠种子萌发[12,17],当连续4周萌发盆中没有新的植物萌发时,试验结束[19,31]。试验于2021年5月初开始,2022年1月中旬结束[12,14,17,19]。
1.3 数据处理
1.3.1 土壤种子库物种和生活型相对丰度
物种相对丰度用某物种个数在全部物种总数中所占比例来表示;生活型相对丰度用某生活型数量在全部生活型总数中所占比例来表示。
1.3.2 土壤种子库密度
土壤种子库密度采用每平方米土壤内所含的种子数量来表示[31]。
1.3.3 土壤种子库物种多样性
不同类型公园土壤种子库物种多样性采用Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)、Pielou均匀度指数(E)、Patrick丰富度指数(R)进行计算[17,32]。
H=−S∑i=1PilnPi (1) D=1−S∑i=1Pi2 (2) E=HlnS (3) R=S (4) 式中:S为种子库物种总数;Pi表示第i个物种的种子数占种子库种子总数的比值。
1.3.4 土壤种子库、地上植被及两者间的关系
采用相似系数(Sørensen指数)和距离测度为Bray-Curtis的非度量多维尺度分析(non-metric multidimensional scaling,NMDS)推断不同类型公园地上植被间、不同类型公园土壤种子库间以及不同类型公园土壤种子库与其对应地上植被间的相似性和群落构成关系[21]。Sørensen指数(Sc)[32]为
Sc=2Wa+b (5) 式中:W表示地上植被与其对应的土壤种子库共有植物种数;a和b分别表示地上植被与土壤种子库各自拥有的物种数量。
1.3.5 土壤种子库与环境要素的关系
土壤种子库密度和物种多样性指数作为响应变量,对其进行除趋势对应分析(decentered correspondence analysis,DCA),获取排序轴梯度长度(length of the gradient axis,LGA)。根据LGA确定合适的分析方法:当LGA < 3时,使用冗余分析(redundancy analysis,RDA);当LGA > 4时,使用典范对应分析;当3 < LGA < 4时,两种分析方法均可使用[33]。城市公园土壤种子库的LGA最大值为0.6(LGA < 3),因此选用RDA分析土壤种子库与地上植被和土壤性质的关系。
1.3.6 统计分析
用SPSS 28.0软件中单因子方差分析和多重比较方法对不同类型公园间土壤种子库的密度、物种多样性以及与地上植被间的相似性系数进行差异显著性分析(α = 0.05)。使用Origin 9.0对数据进行作图。使用R 4.1.2软件“vegan”包中的rda函数进行RDA分析,用metaMDS函数进行NMDS排序,为了满足正态分布要求,在分析前对部分数据进行转换。
2. 结果与分析
2.1 土壤种子库特征
2.1.1 不同类型公园土壤种子库的物种组成和密度
由表2可知:4类城市公园75个土壤种子库样品中共萌发77种植物,隶属于34科74属,平均密度变化范围在352 ~ 899粒/m2。菊科物种数最多(16种),其次为禾本科(6种)、豆科(4种)。车前(Plantago asiatica)、活血丹(Glechoma longituba)、夏至草(Lagopsis supina)、铁苋菜(Acalypha australis)、狗尾草(Setaria viridis)、马唐(Digitaria sanguinalis)、牛筋草、早熟禾(Poa annua)等19种植物在4类城市公园土壤种子库中都存在,表明这些物种在城市公园中分布广泛。物种密度较大的植物为马唐(147.11粒/m2)、苜蓿(Medicago sativa)(128.40粒/m2)、狗尾草(122.67粒/m2)、早熟禾(121.13粒/m2)、牛筋草(100.36粒/m2)、中华苦荬菜(Ixeris chinensis)(96.30粒/m2)等。
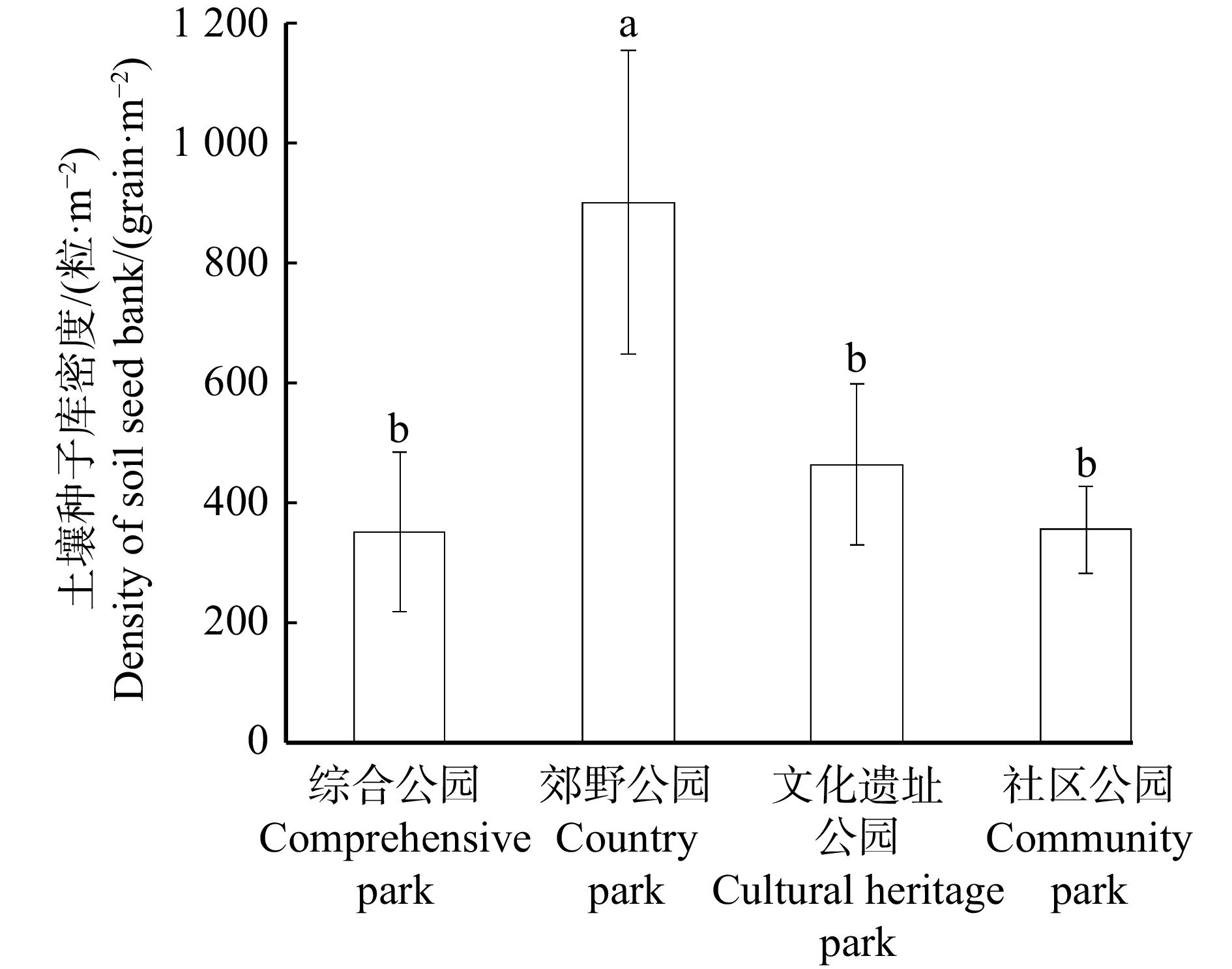
综合公园有植物种子41种,隶属21科40属,以马唐(34.4%)为主的禾本科物种相对丰度较大(46.3%),其次为蔷薇科(12.4%)、菊科(10.2%);种子库密度较大的物种为马唐(287.04粒/m2)和朝天委陵菜(Potentilla supina)(207.41粒/m2) 。郊野公园有植物种子40种,隶属22科39属,其中禾本科与莎草科物种相对丰度较大,分别为35.1%和26.1%,其中狗尾草(216.67粒/m2)的密度最大,其次为牛筋草(129.29粒/m2)、马唐(119.75粒/m2)等。文化遗址公园有植物种子36种,隶属21科36属,其中以早熟禾(21.2%)为主的禾本科物种相对丰度较大(42.6%),其次为马齿苋科(12.8%)、紫草科(9.8%)、酢浆草科(9.1%);密度较大的物种为马齿苋(Portulaca oleracea) (276.54粒/m2)、附地菜(Trigonotis peduncularis) (212.35粒/m2)、早熟禾(196.83粒/m2)和牛筋草(154.07粒/m2)。社区公园有植物种子47种,隶属31科46属,其中以马唐(23.5%)为主的禾本科物种相对丰度较大(39.3%),其次为紫草科(8.3%)、菊科(7.0%)、马齿苋科(6.0%);密度较大的物种为繁缕(Stellaria media)(488.89粒/m2)、苜蓿(148.15粒/m2)、马唐(140.74粒/m2)等。不同类型公园土壤种子库密度排序:郊野公园(899 ± 252)粒/m2 > 文化遗址公园(463 ± 134)粒/m2 > 社区公园(355 ± 72)粒/m2 > 综合公园(352 ± 133)粒/m2。从图2可以看出,郊野公园与文化遗址公园、综合公园、社区公园的种子库密度存在显著差异(P < 0.05)。
![]() 图 2 不同类型城市公园土壤种子库密度不同小写字母表示不同公园类型间差异显著(P < 0.05)。误差线代表标准误差。下同。Different small letters indicate significant differences among different park types at 0.05 level. The error line represents the standard error. Same as below.Figure 2. Density of soil seed banks in different types of urban parks表 2 不同类型城市公园土壤种子库物种组成和密度Table 2. Species composition and density of soil seed banks in different types of urban parks
图 2 不同类型城市公园土壤种子库密度不同小写字母表示不同公园类型间差异显著(P < 0.05)。误差线代表标准误差。下同。Different small letters indicate significant differences among different park types at 0.05 level. The error line represents the standard error. Same as below.Figure 2. Density of soil seed banks in different types of urban parks表 2 不同类型城市公园土壤种子库物种组成和密度Table 2. Species composition and density of soil seed banks in different types of urban parks科
Family属
Genus种
Species生活型
Life
form物种密度/(粒·m−2) Species density/(grain·m−2) 综合公园
Comprehensive
park郊野公园
Country
park文化遗址
公园
Cultural
heritage park社区公园
Community
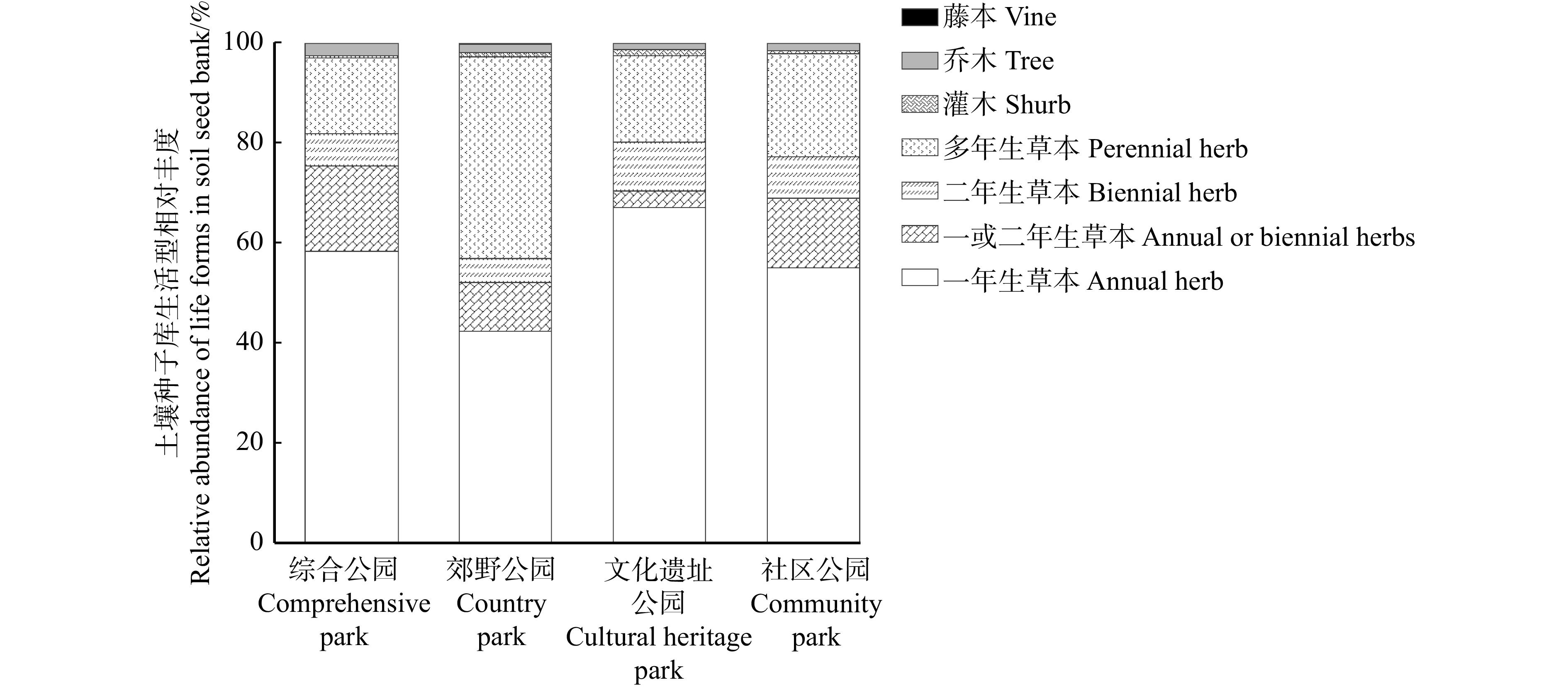
park车前科 Plantaginaceae 车前属 Plantago 车前 Plantago asiatica P 14.81 62.96 14.81 22.22 婆婆纳属 Veronica 蚊母草 Veronica peregrina A 14.81 唇形科 Lamiaceae 薄荷属 Mentha 薄荷 Mentha canadensis P 14.81 111.11 活血丹属 Glechoma 活血丹 Glechoma longituba P 14.81 71.96 14.81 37.04 夏至草属 Lagopsis 夏至草 Lagopsis supina P 29.63 22.22 14.81 14.81 大戟科 Euphorbiaceae 大戟属 Euphorbia 地锦草 Euphorbia humifusa A 29.63 29.63 19.75 铁苋菜属 Acalypha 铁苋菜 Acalypha australis A 22.22 29.63 143.21 22.22 豆科 Fabaceae 胡枝子属 Lespedeza 胡枝子 Lespedeza bicolor S 74.07 决明属 Senna 决明 Senna tora A 103.70 米口袋属 Gueldenstaedtia 少花米口袋 Gueldenstaedtia verna P 59.26 苜蓿属 Medicago 苜蓿 Medicago sativa P 88.89 148.15 禾本科 Poaceae 地毯草属 Axonopus 地毯草 Axonopus compressus P 29.63 狗尾草属 Setaria 狗尾草 Setaria viridis A 18.52 216.67 56.30 122.22 黑麦草属 Lolium 黑麦草 Lolium perenne P 14.81 马唐属 Digitaria 马唐 Digitaria sanguinalis A 287.04 119.75 48.68 140.74 穇属 Eleusine 牛筋草 Eleusine indica A 55.56 129.29 154.07 67.72 早熟禾属 Poa 早熟禾 Poa annua A 79.01 77.04 196.83 29.63 虎耳草科 Saxifragaceae 虎耳草属 Saxifraga 虎耳草 Saxifraga stolonifera P 44.44 29.63 夹竹桃科 Apocynaceae 鹅绒藤属 Cynanchum 萝藦 Cynanchum rostellatum V 14.81 堇菜科 Violaceae 堇菜属 Viola 堇菜 Viola arcuata P 19.75 紫花地丁 Viola philippica P 14.81 14.81 88.89 锦葵科 Grewia 扁担杆属 Grewia 扁担杆 Grewia biloba S 14.81 苘麻属 Abutilon 苘麻 Abutilon theophrasti A 14.81 14.81 田麻属 Corchoropsis 田麻 Corchoropsis crenata A 14.81 菊科 Asteraceae 飞蓬属 Erigeron 小蓬草 Erigeron canadensis A 22.22 一年蓬 Erigeron annuus AB 14.81 鬼针草属 Bidens 鬼针草 Bidens pilosa A 14.81 蒿属 Artemisia 青蒿 Artemisia caruifolia A 59.26 29.63 44.44 黄鹌菜属 Youngia 黄鹌菜 Youngia japonica P 74.07 29.63 蓟属 Cirsium 蓟 Cirsium japonicum P 74.07 29.63 103.70 91.85 假还阳参属 Crepidiastrum 假还阳参 Crepidiastrum lanceolatum P 103.70 59.26 金光菊属 Rudbeckia 黑心金光菊 Rudbeckia hirta AB 14.81 菊三七属 Gynura 白子菜 Gynura divaricata P 59.26 苦苣菜属 Sonchus 长裂苣荬菜 Sonchus brachyotus A 14.81 22.22 苦苣菜 Sonchus oleraceus AB 31.75 47.41 14.81 24.69 苦荬菜属 Ixeris 中华苦荬菜 Ixeris chinensis P 113.58 44.44 鳢肠属 Eclipta 鳢肠 Eclipta prostrata A 14.81 牛膝菊属 Galinsoga 牛膝菊 Galinsoga parviflora A 22.22 29.63 14.81 蒲公英属 Taraxacum 蒲公英 Taraxacum mongolicum P 29.63 29.63 旋覆花属 Inula 旋覆花 Inula japonica P 14.81 苦木科 Simaroubaceae 臭椿属 Ailanthus 臭椿 Ailanthus altissima T 14.81 蓼科 Polygonaceae 千叶兰属 Muehlenbeckia 竹节蓼 Muehlenbeckia platyclada P 44.44 列当科 Orobanchaceae 地黄属 Rehmannia 地黄 Rehmannia glutinosa P 14.81 马齿苋科 Portulacaceae 马齿苋属 Portulaca 马齿苋 Portulaca oleracea A 74.07 48.48 276.54 72.22 木樨科 Oleaceae 梣属 Fraxinus 白蜡树 Fraxinus chinensis T 22.22 29.63 茜草科 Rubiaceae 耳草属 Hedyotis 耳草 Hedyotis auricularia P 14.81 29.63 拉拉藤属 Galium 车轴草 Galium odoratum P 14.81 蔷薇科 Rosaceae 委陵菜属 Potentilla 朝天委陵菜 Potentilla supina AB 207.41 51.03 44.44 37.04 茄科 Solanaceae 茄属 Solanum 龙葵 Solanum nigrum A 14.81 忍冬科 Caprifoliaceae 忍冬属 Lonicera 金银忍冬 Lonicera maackii S 14.81 桑科 Moraceae 构属 Broussonetia 构 Broussonetia papyrifera T 14.81 桑属 Morus 桑 Morus alba T 29.63 14.81 14.81 莎草科 Cyperaceae 莎草属 Cyperus 莎草 Cyperus rotundus P 14.81 水蜈蚣属 Kyllinga 短叶水蜈蚣 Kyllinga brevifolia P 29.63 44.44 14.81 薹草属 Carex 薹草 Carex spp. P 29.63 88.89 十字花科 Brassicaceae 独行菜属 Lepidium 独行菜 Lepidium apetalum AB 29.63 88.89 14.81 37.86 荠属 Capsella 荠 Capsella bursa-pastoris AB 14.81 诸葛菜属 Orychophragmus 诸葛菜 Orychophragmus violaceus AB 29.63 64.20 石竹科 Caryophyllaceae 繁缕属 Stellaria 繁缕 Stellaria media AB 14.81 29.63 488.89 漆姑草属 Sagina 漆姑草 Sagina japonica AB 14.81 天门冬科 Asparagaceae 沿阶草属 Ophiopogon 沿阶草 Ophiopogon bodinieri P 138.27 14.81 55.56 通泉草科 Mazaceae 通泉草属 Mazus 通泉草 Mazus pumilus A 14.81 44.44 14.81 125.93 无患子科 Sapindaceae 栾属 Koelreuteria 栾 Koelreuteria paniculata T 29.63 44.44 22.22 14.81 槭属 Acer 元宝槭 Acer truncatum T 59.26 苋科 Amaranthaceae 沙冰藜属 Bassia 地肤 Bassia scoparia A 74.07 藜属 Chenopodium 藜 Chenopodium album A 29.63 32.59 39.51 48.15 苋属 Amaranthus 凹头苋 Amaranthus blitum A 29.63 44.44 旋花科 Convolvulaceae 打碗花属 Calystegia 打碗花 Calystegia hederacea A 14.81 虎掌藤属 Ipomoea 牵牛 Ipomoea nil A 14.81 荨麻科 Urticaceae 雾水葛属 Pouzolzia 雾水葛 Pouzolzia zeylanica P 14.81 苎麻属 Boehmeria 苎麻 Boehmeria nivea S 29.63 29.63 74.07 44.44 鸭跖草科 Commelinaceae 鸭跖草属 Commelina 鸭跖草 Commelina communis A 14.81 银杏科 Ginkgoaceae 银杏属 Ginkgo 银杏 Ginkgo biloba T 14.81 罂粟科 Papaveraceae 紫堇属 Corydalis 紫堇 Corydalis edulis A 14.81 鸢尾科 Iridaceae 鸢尾属 Iris 鸢尾 Iris tectorum P 29.63 紫草科 Boraginaceae 附地菜属 Trigonotis 附地菜 Trigonotis peduncularis B 107.41 69.63 212.35 80.00 酢浆草科 Oxalidaceae 酢浆草属 Oxalis 酢浆草 Oxalis corniculata P 19.75 37.04 98.77 59.26 注:A.一年生草本;B.二年生草本;AB.一或二年生草本;P.多年生草本;V.藤本;S.灌木;T.乔木。Notes: A, annual herb; B, biennial herb; AB, annual or biennial herb; P, perennial herb; V, vine; S, shrub; T, tree. 城市公园土壤种子库生活型相对丰度排序:一年生草本(55.73%) > 多年生草本(23.43%) > 一或二年生草本(10.97%) > 二年生草本(7.36%) > 乔木(1.69%) > 灌木(0.79%) > 藤本(0.03%)。其中草本相对丰度最大(平均占比达97.5%),乔、灌木和藤本植物种子数最少(占比均 < 2.0%)。由图3可看出,4类城市公园均以一、二年生草本为主(综合公园81.8%、郊野公园57.0%、文化遗址公园80.2%、社区公园77.2%),其中郊野公园多年生草本植物的相对丰度(40.4%)要大于其他类型公园(综合公园15.3%、文化遗址公园17.3%、社区公园20.7%)。整体来看,城市公园的土壤种子库主要以一、二年生草本为主,多年生草本为辅,草本层对城市公园植物群落结构以及物种多样性维持具有重要意义。
![]() 图 3 不同类型城市公园土壤种子库生活型组成Figure 3. Life type composition of soil seed banks in different types of urban parks
图 3 不同类型城市公园土壤种子库生活型组成Figure 3. Life type composition of soil seed banks in different types of urban parks2.1.2 不同类型公园土壤种子库物种多样性
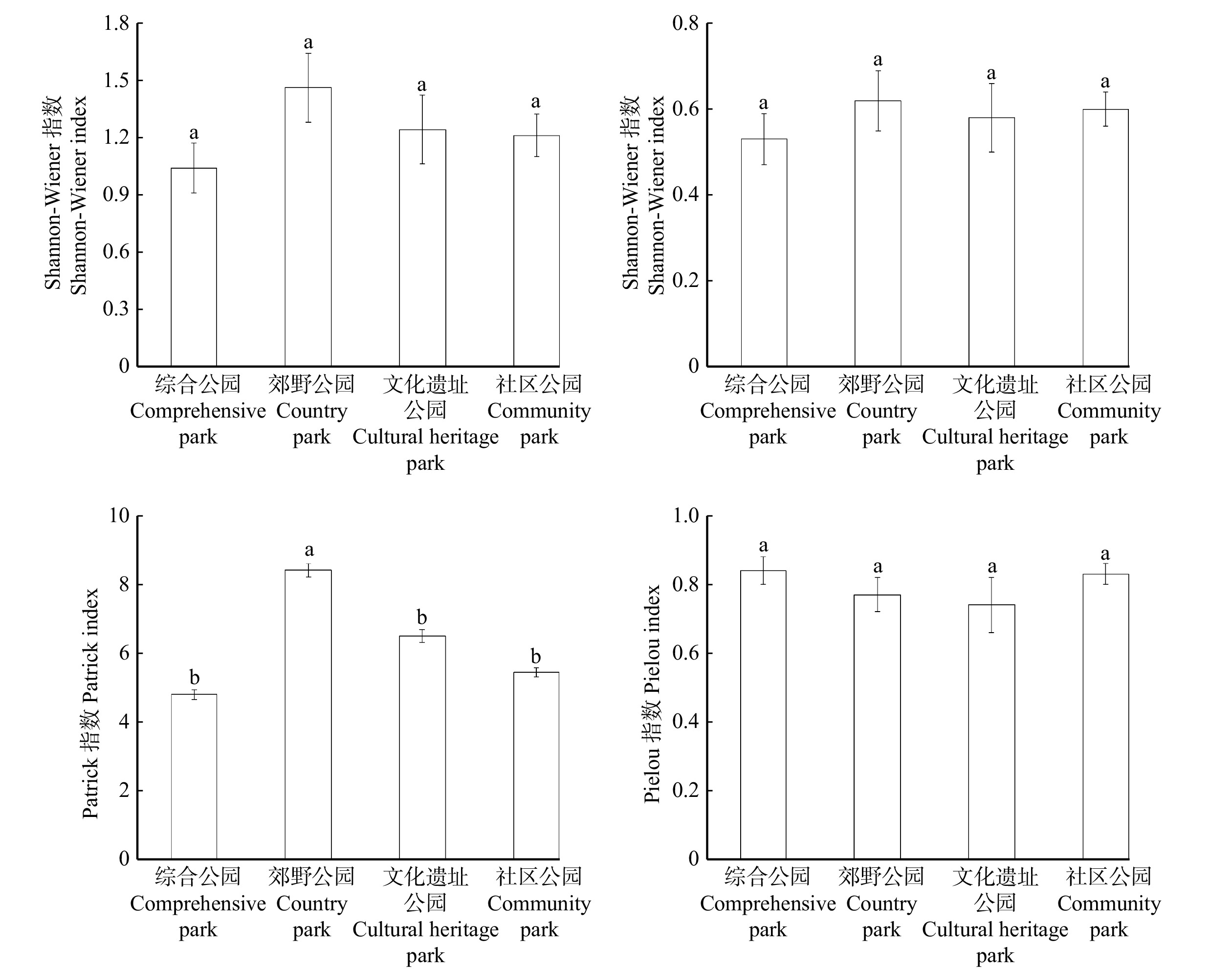
城市公园土壤种子库的Shannon-Wiener多样性指数范围在1.04 ~ 1.46,其中郊野公园(1.46 ± 0.18) > 文化遗址公园(1.24 ± 0.18) > 社区公园(1.21 ± 0.11) > 综合公园(1.04 ± 0.13);Simpson优势度指数范围在0.53 ~ 0.62,其中郊野公园(0.62 ± 0.07) > 社区公园(0.60 ± 0.04) > 文化遗址公园(0.58 ± 0.08) > 综合公园(0.53 ± 0.06);Patrick丰富度指数范围在4.79 ~ 8.41,其中郊野公园(8.41 ± 0.19) > 文化遗址公园(6.50 ± 0.19) > 社区公园(5.44 ± 0.14) > 综合公园(4.79 ± 0.15);Pielou均匀度指数范围在0.74 ~ 0.84,其中综合公园(0.84 ± 0.04) > 社区公园(0.83 ± 0.03) > 郊野公园(0.77 ± 0.05) > 文化遗址公园(0.74 ± 0.08)。城市公园土壤种子库的多样性较高,物种也较为丰富且分布较均匀。从图4可以看出,郊野公园种子库的丰富度Patrick指数最高且与其他3类公园间存在显著差异(P < 0.05)。
![]() 图 4 不同类型城市公园土壤种子库物种多样性指标Figure 4. Species diversity of soil seed banks in different types of urban parks
图 4 不同类型城市公园土壤种子库物种多样性指标Figure 4. Species diversity of soil seed banks in different types of urban parks2.2 土壤种子库与地上植被的关系
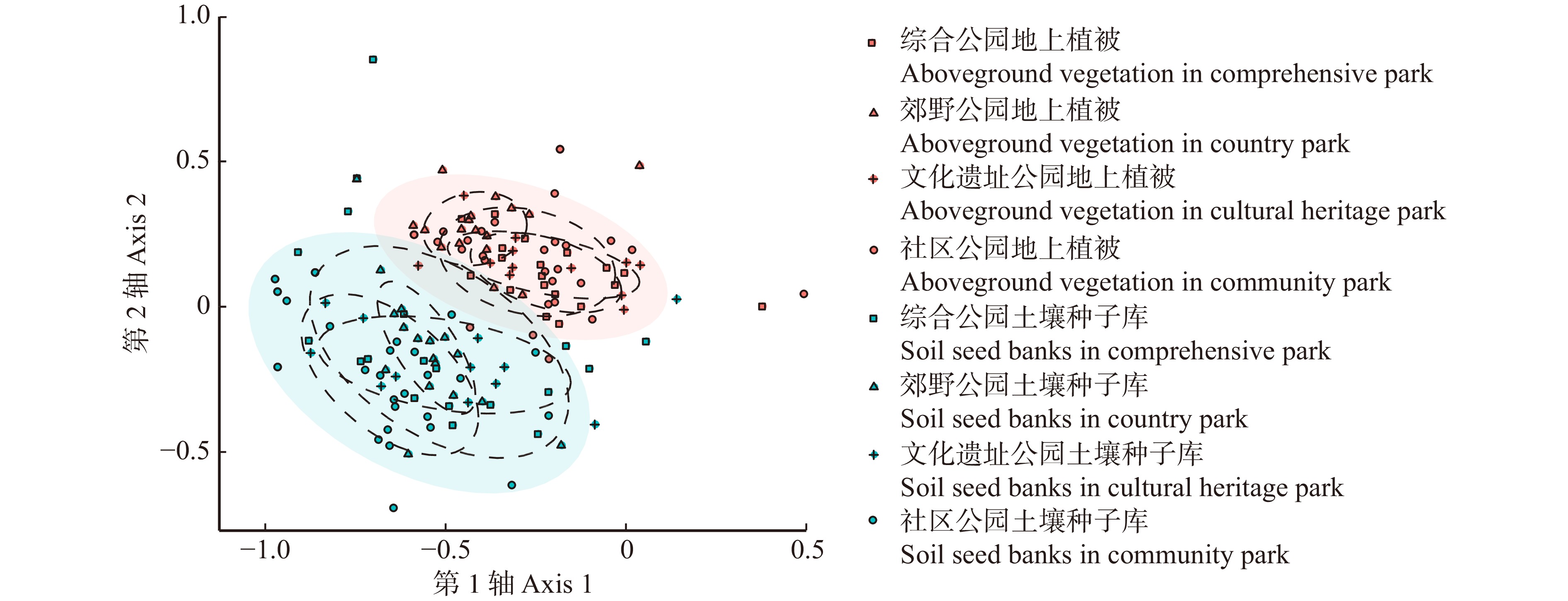
不同类型公园间土壤种子库相似性系数和不同类型公园间地上植被相似性系数分别在0.56 ~ 0.68和0.59 ~ 0.65之间(表3),相似性均较高,且群落构成差异不明显(图5)。其中综合公园与社区公园相似性最高(土壤种子库间相似性系数为0.68,地上植被间相似性系数为0.65)。
表 3 不同类型城市公园之间土壤种子库、地上植被Sørensen相似性系数Table 3. Sørensen similarity coefficients of soil seed banks and aboveground vegetation species among different types of urban parks公园类型 土壤种子库间物种相似性
Species similarity among soil seed banks地上植被间物种相似性
Species similarity among aboveground vegetations综合公园
Comprehensive park郊野公园
Country park文化遗址公园
Cultural heritage park综合公园
Comprehensive park郊野公园
Country park文化遗址公园
Cultural heritage park郊野公园
Country park0.64 0.62 文化遗址公园
Cultural heritage park0.64 0.57 0.60 0.59 社区公园
Community park0.68 0.56 0.57 0.65 0.64 0.60 ![]() 图 5 不同类型城市公园土壤种子库与地上植被NMDS排序Figure 5. NMDS ranking of soil seed banks and aboveground vegetation in different types of urban parks
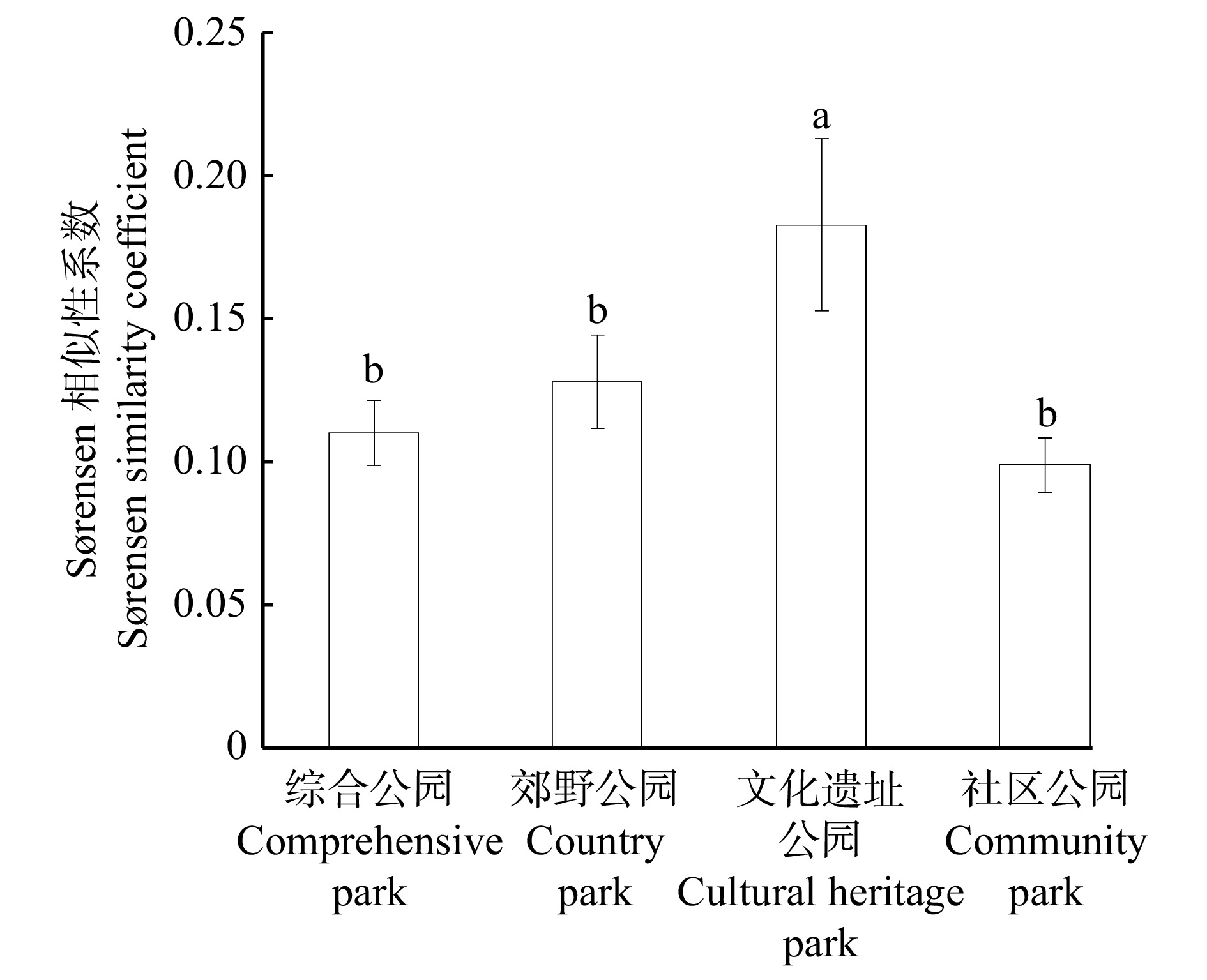
图 5 不同类型城市公园土壤种子库与地上植被NMDS排序Figure 5. NMDS ranking of soil seed banks and aboveground vegetation in different types of urban parks地上植被和土壤种子库间的NMDS分析结果表明(图5),两者间群落构成有一定的差异。不同类型公园间土壤种子库与其对应地上植被间的相似性系数范围为0.10 ~ 0.18,属于较低相似水平,其中文化遗址公园土壤种子库与其对应地上植被间的相似性系数最高(0.18),且与其他类型公园之间存在显著差异(P < 0.05)(图6)。
![]() 图 6 不同类型城市公园土壤种子库与地上植被之间Sørensen相似性系数Figure 6. Sørensen similarity coefficients between soil seed banks and aboveground vegetation in different types of urban parks
图 6 不同类型城市公园土壤种子库与地上植被之间Sørensen相似性系数Figure 6. Sørensen similarity coefficients between soil seed banks and aboveground vegetation in different types of urban parks2.3 地上植被和土壤性质对土壤种子库的影响
将城市公园土壤种子库特征(密度和物种多样性指标)作为响应变量,将除去了强共线性因子的地上植被以及土壤性质(草本多样性指数、草本物种丰富度指数、灌木物种丰富度指数、灌木均匀度指数、土壤铵态氮、硝态氮、全氮、总孔隙度、非毛管孔隙度和pH值)作为解释变量,进行RDA分析,筛选出影响土壤种子库特征的主要环境因子。
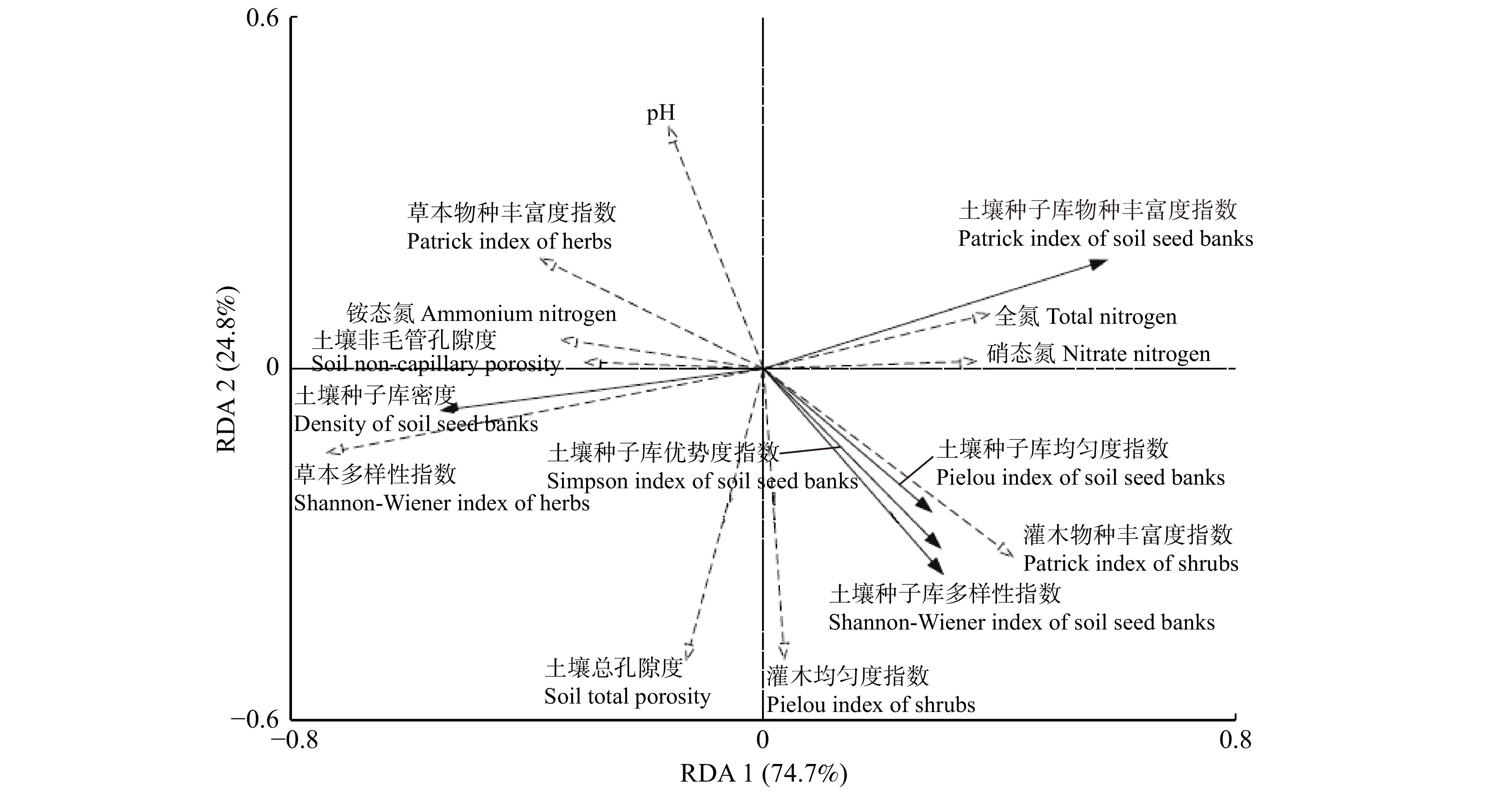
土壤种子库特征与地上植被和土壤性质的RDA排序结果表明(图7),地上植被草本多样性指数(r = 0.335)和灌木物种丰富度指数(r = 0.326)与RDA1轴的相关性较大,累计方差贡献率为74.7%,两轴的累计贡献率达99.5%,表明地上植被物种多样性以及土壤理化性质会显著影响土壤种子库特征。土壤种子库密度与草本多样性指数呈极显著正相关(P < 0.01),与草本物种丰富度指数、土壤铵态氮和非毛管孔隙度呈显著正相关(P < 0.05),与土壤全氮和硝态氮呈显著负相关;土壤种子库物种丰富度指数与草本多样性指数呈极显著负相关,与土壤铵态氮呈显著负相关,与土壤全氮和硝态氮呈显著正相关;土壤种子库均匀度、优势度和多样性指数与灌木物种丰富度指数呈显著正相关,与草本物种丰富度指数呈显著负相关。其中,草本多样性指数和灌木物种丰富度指数对城市公园土壤种子库的密度和物种多样性有显著影响(P < 0.05)。
![]() 图 7 土壤种子库特征与环境因子间RDA排序图Figure 7. RDA ranking diagram between soil seed banks and environmental factors
图 7 土壤种子库特征与环境因子间RDA排序图Figure 7. RDA ranking diagram between soil seed banks and environmental factors3. 讨 论
3.1 不同类型公园土壤种子库特征
土壤种子库是潜伏在土壤中的植物群落,是森林、农田、湿地和草地等生态系统植被更新和恢复的基础[34],其物种组成决定着生态系统的稳定性、生产力及营养动态等功能[35]。城市公园土壤种子库共萌发植物77种,隶属于34科74属,以菊科、禾本科物种为主。这两科植物多是自生植物[36],普遍克隆生长、有强竞争力且能在土壤中长时间存活,是种子库的主要物种,这与其他研究结果[28,37−38]相符。各个类型公园土壤种子库的组成相对简单,草本植物占绝对优势(平均占比97.5%),而乔、灌木占比均较少。草本植物的种子体型小、质量小、产量高,能在风力、动物携带等外力作用下散布,进入土壤更加容易[39],且部分草本植物,如中华苦荬菜、紫花地丁(Viola philippica)等,在5—7月份采样前已经产种,这都会增加草本植物在种子库中的比例。而大部分乔、灌木(如栾、元宝槭Acer truncatum、金银忍冬、北京丁香等)则在5—7月采样后才会散布种子,且种子落下后会被动物捕食[11]或随着凋落物的清理而被清除,部分落入土壤的种子会进入休眠期或者腐烂死亡[31,38]。此外,由于繁殖策略不同[17],部分木本植物的营养繁殖优于种子繁殖,如毛白杨、垂柳(Salix babylonica)、火炬树(Rhus typhina)等。相比其他类型公园,郊野公园多年生草本的相对丰度较大,这可能是因为该类公园多选取观赏效果好、管理方便且节约成本的多年生草本(如薹草(Carex spp.)、沿阶草等)作为公园绿化的草本植物,所以其土壤种子库的多年生草本种子相较更多。
土壤种子库作为植被更新演替的种源储备库,其数量和物种多样性特征是判断植被恢复能力的重要依据[13]。北京城市公园种子库密度在352 ~ 899粒/m2之间,与森林类型种子库密度(100 ~ 1 000粒/m2)相近[40]。不同城市不同研究区土壤种子库密度有所不同:上海城市废弃地土壤种子库密度为837 ~ 2675粒/m2[41];沈阳和抚顺交接的城市近郊林种子库密度为154 ~ 455粒/m2[42];北京山区自然边坡种子库密度为1.4 × 104 ~ 2.3 × 104粒/m2[43]。本研究种子库密度结果相较其他较小,一方面由于萌发试验周期为8个月,可能存在小部分土壤种子由于种皮较厚、休眠期过长等原因未萌发,另一方面可能是本研究样点的土壤pH值偏大(pH > 8)造成的。相关研究[44]表明:当土壤pH值偏高时,攻击种子的细菌、真菌数量增加,不利于种子的萌发和存储。另外,受现有采样技术[45](采样位置、大小等)和萌发条件(温度、光照、湿度等)等因素影响,很难获取土壤中所有的种子,也很难保证每个种子都拥有理想的萌发条件。不同植被群落土壤种子库的物种组成以及生态功能有一定差异,相似群落的种子库特征也会有不同之处[46]。郊野公园的土壤种子库密度显著高于其他3类公园(P < 0.05),这可能是由公园人为管理不同所致。地上植被在成熟期产种,大多数种子会先落在地表枯枝落叶层,再通过动物搬运、雨水输送等方式进入土壤[42],而郊野公园以自然景观为主,人为干扰较其他类型公园小,地表枯枝落叶多,因此落在该层的种子会被保护起来,避免被鸟类、脊椎动物等捕食[31,47],从而增加种子进入土壤的几率。与不同类型公园地上植被物种多样性的变化规律相似,郊野公园土壤种子库的多样性、优势度、丰富度最高,可见此类公园的潜在稳定性更高[17]。其中该类公园物种丰富度显著大于其他类型公园,可能是由于人为干扰和地上植被类型的差异,郊野公园能够为适应性强的自生植物提供更充足的生长空间[36],从而提高土壤种子库的物种丰富度,同时部分春季开花的植物会在夏季来临前撒种,进一步丰富了种子库的物种类型。
3.2 土壤种子库与地上植被的关系
地上植被产种是种子库形成的基础,种子库的种子萌发也会影响地上植被的结构和组成,研究两者之间的相似性,能够用来衡量该植被群落的稳定性并预测未来的发展趋势[37]。本研究对比了不同类型公园土壤种子库、地上植被以及两者之间的关系。结果表明:不同类型公园间土壤种子库、地上植被相似性系数均较高,且群落构成差异不大。这意味着各类公园有着相同的物种基础[43],如:油松、毛白杨、连翘(Forsythia suspensa)等北京公园常栽植物[48],以及狗尾草、早熟禾、藜(Chenopodium album)等土壤种子库常见的先锋物种[17,43]。其中综合公园与社区公园的土壤种子库相似性最高,这与两者地上植被构成相似有关,也表明两者潜在发展趋势较为一致[17]。Tedoradze等[49]发现高山坡地地上植被与种子库之间相似性普遍较低,两者间仅有18%的共有物种;Wang等[31,50]研究了不同地区人工林植被与土壤种子库之间的关系,均得出种子库与地上植被的相似性较低的结论。本研究也有着类似的结果,城市公园土壤种子库与其对应的地上植被间仅有16%的共有物种,包括栾、胡枝子(Lespedeza bicolor)、狗尾草、假还阳参(Crepidiastrum lanceolatum)等。这是由多种原因导致的[51]。一方面城市公园受人为干扰较大:公园土壤大多为客土回填,其种子库物种组成除了后期地上植被落入土壤的物种外,客土本身还存在一部分原有的植物种子;城市公园人为管护(如清扫枯枝落叶)较多,地上植被下落至土壤中的种子量较少[37],使种子库与地上植被间物种组成不稳定。另一方面城市内斑块密集,会有外来物种进入土壤,如未在地上植被调查到的虎耳草(Saxifraga stolonifera)、紫堇(Corydalis edulis)等11种草本植物,从而影响种子库的物种组成。在以往研究结果中,生态保育绿地土壤种子库与地上植被相似性显著高于其他绿地类型,生态保育绿地主要以生态防护功能为主,因此弱化了人为管理程度[19]。而在本研究4类城市公园中,由于文化遗址公园依地而建,存在年限长,绿地大多被管护起来,园内客土较少,有更多的地上植被种子能用来补充土壤种子库,因此其种子库与地上植被间的相似性系数(0.18)显著高于其他类型公园。因此,在城市绿地管理中,减少人为干扰有助于提高植被群落的更新潜力。
3.3 环境要素对土壤种子库的影响
土壤种子库的组成和发展不仅会受地上植被的影响,还会受到地形地理、土壤理化性质、土壤微生物等因素的影响[52]。除了外界干扰因素[44,53]通过影响地上植被间接影响种子库外,地上植被本身的群落结构[28,54]、物种多样性等因子也会对种子库产生影响。研究结果显示,地上植被草本多样性和灌木物种丰富度在较大程度上影响着土壤种子库特征。地上草本植物种子产量多、易进入土壤[39],因此种子库密度会随地上草本多样性的增加而增加;但地上草本植被多由土壤种子库萌发而来,且部分草本植物会优先、大量萌发(如藜、紫花地丁、中华苦荬菜等),因此种子库物种丰富度会随地上草本多样性的增加而减小。地上灌木植被物种丰富度越高,向种子库输送的物种也越丰富[55],其产生的枯落物在降低种子被捕食几率的同时[31],还能为土壤提供营养物质,从而促进种子的生长发育,因此土壤种子库多样性会随着地上灌木植被物种丰富度的增加而增加。先前研究表明,在不同研究区对土壤种子库影响较大的土壤因子有所不同[17,44,55−56]。本研究中土壤种子库密度和物种丰富度主要受土壤全氮、铵态氮和硝态氮的影响,这与胡安[34]的研究结果一致。氮作为植物生长发育的必需营养元素,能够直接或间接影响植物体内某些元素的组成[14],对种子萌发影响较大。土壤中的铵态氮会抑制种子的萌发,从而增加土壤中种子库的密度,种子未萌发则地上植被物种丰富度会降低,进而影响土壤种子库的物种丰富度;全氮和硝态氮会促进禾本科、豆科等多科的植物种子萌发,从而降低土壤种子库密度,但萌发后的植被又会产生新种子,进而增加土壤种子库的丰富度。此外,土壤种子库密度还受土壤非毛管孔隙度的影响。土壤非毛管孔隙度是土壤通气性的指标之一,影响着土壤透气、渗水性能。当土壤较为密实、非毛管孔隙度较小时,其高保水性会促进攻击种子的细菌、真菌的增殖以及降低土壤交换气体的能力,增加种子腐烂的几率[57]。本研究结果显示,土壤非毛管孔隙度与种子库密度呈显著正相关,表明非毛管孔隙度较高的土壤对种子萌发、存活有正向影响,有利于种子的积累。
4. 结论与建议
城市公园土壤种子库密度较小,物种组成以草本为主,且与其对应的地上植被间相似性低。地上植被草本多样性和灌木物种丰富度对土壤种子库密度和物种多样性影响较大,表现为:地上草本多样性越高,土壤种子库密度越大,物种丰富度越小;地上灌木物种丰富度越高,土壤种子库多样性、优势度和均匀度越高。其中,郊野公园种子库密度和物种丰富度显著高于其他类型公园;由于建存时间长,客土较少,文化遗址公园土壤种子库与地上植被间的相似性显著高于其他类型公园。可见,城市公园中郊野公园和文化遗址公园土壤种子库更新潜力较大,其他类型公园应在保留自身功能特点定位的前提下,将郊野公园和文化遗址公园中的人为管理作为参考,增加土壤种子库规模、多样性及与相应地上植被间的相似性,提高植被更新潜力。总体而言,城市公园土壤种子库维持植被物种多样性,促进植被更新的能力有限,在城市公园绿地管理中,可减少草本植物的拔除,适当增加灌木树种的栽植比例,增大土壤种子库规模和物种多样性,促进公园木本植被的更新演替,营建近自然城市公园。由于种子萌发试验持续8个月,可能存在小部分持久种子库由于种皮较厚、休眠期过长等原因未萌发,以后研究中可以适当延长萌发时间,并注意公园中自生植物及其种子库的特点,进一步研究城市公园土壤种子库的特征及植被的自然更新潜力。
-
![]()
图 1 北京市六环内公园研究样地分布
Figure 1. Distribution of study sample plots of parks in the Sixth Ring Road of Beijing
![]()
图 2 不同类型城市公园土壤种子库密度
不同小写字母表示不同公园类型间差异显著(P < 0.05)。误差线代表标准误差。下同。Different small letters indicate significant differences among different park types at 0.05 level. The error line represents the standard error. Same as below.
Figure 2. Density of soil seed banks in different types of urban parks
![]()
图 3 不同类型城市公园土壤种子库生活型组成
Figure 3. Life type composition of soil seed banks in different types of urban parks
![]()
图 4 不同类型城市公园土壤种子库物种多样性指标
Figure 4. Species diversity of soil seed banks in different types of urban parks
![]()
图 5 不同类型城市公园土壤种子库与地上植被NMDS排序
Figure 5. NMDS ranking of soil seed banks and aboveground vegetation in different types of urban parks
![]()
图 6 不同类型城市公园土壤种子库与地上植被之间Sørensen相似性系数
Figure 6. Sørensen similarity coefficients between soil seed banks and aboveground vegetation in different types of urban parks
![]()
图 7 土壤种子库特征与环境因子间RDA排序图
Figure 7. RDA ranking diagram between soil seed banks and environmental factors
表 1 公园分类
Table 1 Classification of parks
公园类型
Type of park位置
Location功能特点
Functional characteristics人为管理
Artificial management综合公园
Comprehensive
park中心区
Central zone综合公园一般规模较大;以人工景观为主;具有休闲游憩、生态景观、文化科普、防灾避灾等功能并全面均衡发展;服务半径大
The scale of comprehensive park is generally large, mainly artificial landscape, and has the functions of recreation, ecological landscape, cultural science, disaster prevention and avoidance, and comprehensive and balanced development; the service radius of such park is large管理机制稳定健全,人为干扰强度较大;资源保护措施完善且效果好;绿地养护水平高
The management mechanism is stable and sound, and the intensity of human interference is large; the resource protection measures are perfect and effective; the level of green space maintenance is high文化遗址公园
Cultural
heritage park中心区
Central zone文化遗址公园规模适中;人文与自然景观相结合;具有突出的文化科普和休闲游憩的功能,对非本地游客有重要影响;服务半径适中
The scale of cultural heritage park is moderate, combining humanistic and natural landscapes, with outstanding functions of popularization of culture and recreation, and have an important impact on non-local visitors; the service radius of such park is moderate管理机制稳定健全,人为干扰强度较大;资源保护措施完善且效果好;绿地养护水平高
The management mechanism is stable and sound, and the intensity of human interference is large; the resource protection measures are perfect and effective; the level of green space maintenance is high社区公园
Community park中心区 (居民区附近)
Central zone (near a residential area)社区公园一般规模较小;以人工景观为主;周围居民为主要服务对象,具有休闲放松、生态景观和防灾避灾等功能;服务半径小
The scale of community parks is generally small, mainly artificial landscape, and the surrounding residents as the main service object, with the functions of leisure and relaxation, ecological landscape and disaster prevention and avoidance; the service radius of such park is small管理机制较稳定,人为干扰强度适中;资源保护措施比较完善且效果比较好;绿地养护水平中等
The management mechanism is relatively sound, and the intensity of human interference is moderate; the resource protection measures are sound and the effect is good; the level of green space maintenance is medium郊野公园
Country park近郊
Suburb郊野公园规模较大;以自然景观为主;生态服务功能突出,兼有休憩娱乐、文化科普和防灾减灾等功能;服务半径较大
The scale of country park is large, mainly natural landscape, with outstanding ecological service functions, and functions of leisure and recreation, culture and science popularization, and disaster prevention and mitigation; the service radius of such park is large管理机制有待提升,人为干扰强度较小;资源保护措施有待完善,效果较好;绿地养护水平一般
The management mechanism needs to be improved, and the intensity of human interference is small; the resource protection measures need to be improved and the effect is good; the level of green space maintenance is average注:引自文献[2, 23−25]。Note: cited from reference [2, 23−25].  下载: 导出CSV
下载: 导出CSV
表 2 不同类型城市公园土壤种子库物种组成和密度
Table 2 Species composition and density of soil seed banks in different types of urban parks
科
Family属
Genus种
Species生活型
Life
form物种密度/(粒·m−2) Species density/(grain·m−2) 综合公园
Comprehensive
park郊野公园
Country
park文化遗址
公园
Cultural
heritage park社区公园
Community
park车前科 Plantaginaceae 车前属 Plantago 车前 Plantago asiatica P 14.81 62.96 14.81 22.22 婆婆纳属 Veronica 蚊母草 Veronica peregrina A 14.81 唇形科 Lamiaceae 薄荷属 Mentha 薄荷 Mentha canadensis P 14.81 111.11 活血丹属 Glechoma 活血丹 Glechoma longituba P 14.81 71.96 14.81 37.04 夏至草属 Lagopsis 夏至草 Lagopsis supina P 29.63 22.22 14.81 14.81 大戟科 Euphorbiaceae 大戟属 Euphorbia 地锦草 Euphorbia humifusa A 29.63 29.63 19.75 铁苋菜属 Acalypha 铁苋菜 Acalypha australis A 22.22 29.63 143.21 22.22 豆科 Fabaceae 胡枝子属 Lespedeza 胡枝子 Lespedeza bicolor S 74.07 决明属 Senna 决明 Senna tora A 103.70 米口袋属 Gueldenstaedtia 少花米口袋 Gueldenstaedtia verna P 59.26 苜蓿属 Medicago 苜蓿 Medicago sativa P 88.89 148.15 禾本科 Poaceae 地毯草属 Axonopus 地毯草 Axonopus compressus P 29.63 狗尾草属 Setaria 狗尾草 Setaria viridis A 18.52 216.67 56.30 122.22 黑麦草属 Lolium 黑麦草 Lolium perenne P 14.81 马唐属 Digitaria 马唐 Digitaria sanguinalis A 287.04 119.75 48.68 140.74 穇属 Eleusine 牛筋草 Eleusine indica A 55.56 129.29 154.07 67.72 早熟禾属 Poa 早熟禾 Poa annua A 79.01 77.04 196.83 29.63 虎耳草科 Saxifragaceae 虎耳草属 Saxifraga 虎耳草 Saxifraga stolonifera P 44.44 29.63 夹竹桃科 Apocynaceae 鹅绒藤属 Cynanchum 萝藦 Cynanchum rostellatum V 14.81 堇菜科 Violaceae 堇菜属 Viola 堇菜 Viola arcuata P 19.75 紫花地丁 Viola philippica P 14.81 14.81 88.89 锦葵科 Grewia 扁担杆属 Grewia 扁担杆 Grewia biloba S 14.81 苘麻属 Abutilon 苘麻 Abutilon theophrasti A 14.81 14.81 田麻属 Corchoropsis 田麻 Corchoropsis crenata A 14.81 菊科 Asteraceae 飞蓬属 Erigeron 小蓬草 Erigeron canadensis A 22.22 一年蓬 Erigeron annuus AB 14.81 鬼针草属 Bidens 鬼针草 Bidens pilosa A 14.81 蒿属 Artemisia 青蒿 Artemisia caruifolia A 59.26 29.63 44.44 黄鹌菜属 Youngia 黄鹌菜 Youngia japonica P 74.07 29.63 蓟属 Cirsium 蓟 Cirsium japonicum P 74.07 29.63 103.70 91.85 假还阳参属 Crepidiastrum 假还阳参 Crepidiastrum lanceolatum P 103.70 59.26 金光菊属 Rudbeckia 黑心金光菊 Rudbeckia hirta AB 14.81 菊三七属 Gynura 白子菜 Gynura divaricata P 59.26 苦苣菜属 Sonchus 长裂苣荬菜 Sonchus brachyotus A 14.81 22.22 苦苣菜 Sonchus oleraceus AB 31.75 47.41 14.81 24.69 苦荬菜属 Ixeris 中华苦荬菜 Ixeris chinensis P 113.58 44.44 鳢肠属 Eclipta 鳢肠 Eclipta prostrata A 14.81 牛膝菊属 Galinsoga 牛膝菊 Galinsoga parviflora A 22.22 29.63 14.81 蒲公英属 Taraxacum 蒲公英 Taraxacum mongolicum P 29.63 29.63 旋覆花属 Inula 旋覆花 Inula japonica P 14.81 苦木科 Simaroubaceae 臭椿属 Ailanthus 臭椿 Ailanthus altissima T 14.81 蓼科 Polygonaceae 千叶兰属 Muehlenbeckia 竹节蓼 Muehlenbeckia platyclada P 44.44 列当科 Orobanchaceae 地黄属 Rehmannia 地黄 Rehmannia glutinosa P 14.81 马齿苋科 Portulacaceae 马齿苋属 Portulaca 马齿苋 Portulaca oleracea A 74.07 48.48 276.54 72.22 木樨科 Oleaceae 梣属 Fraxinus 白蜡树 Fraxinus chinensis T 22.22 29.63 茜草科 Rubiaceae 耳草属 Hedyotis 耳草 Hedyotis auricularia P 14.81 29.63 拉拉藤属 Galium 车轴草 Galium odoratum P 14.81 蔷薇科 Rosaceae 委陵菜属 Potentilla 朝天委陵菜 Potentilla supina AB 207.41 51.03 44.44 37.04 茄科 Solanaceae 茄属 Solanum 龙葵 Solanum nigrum A 14.81 忍冬科 Caprifoliaceae 忍冬属 Lonicera 金银忍冬 Lonicera maackii S 14.81 桑科 Moraceae 构属 Broussonetia 构 Broussonetia papyrifera T 14.81 桑属 Morus 桑 Morus alba T 29.63 14.81 14.81 莎草科 Cyperaceae 莎草属 Cyperus 莎草 Cyperus rotundus P 14.81 水蜈蚣属 Kyllinga 短叶水蜈蚣 Kyllinga brevifolia P 29.63 44.44 14.81 薹草属 Carex 薹草 Carex spp. P 29.63 88.89 十字花科 Brassicaceae 独行菜属 Lepidium 独行菜 Lepidium apetalum AB 29.63 88.89 14.81 37.86 荠属 Capsella 荠 Capsella bursa-pastoris AB 14.81 诸葛菜属 Orychophragmus 诸葛菜 Orychophragmus violaceus AB 29.63 64.20 石竹科 Caryophyllaceae 繁缕属 Stellaria 繁缕 Stellaria media AB 14.81 29.63 488.89 漆姑草属 Sagina 漆姑草 Sagina japonica AB 14.81 天门冬科 Asparagaceae 沿阶草属 Ophiopogon 沿阶草 Ophiopogon bodinieri P 138.27 14.81 55.56 通泉草科 Mazaceae 通泉草属 Mazus 通泉草 Mazus pumilus A 14.81 44.44 14.81 125.93 无患子科 Sapindaceae 栾属 Koelreuteria 栾 Koelreuteria paniculata T 29.63 44.44 22.22 14.81 槭属 Acer 元宝槭 Acer truncatum T 59.26 苋科 Amaranthaceae 沙冰藜属 Bassia 地肤 Bassia scoparia A 74.07 藜属 Chenopodium 藜 Chenopodium album A 29.63 32.59 39.51 48.15 苋属 Amaranthus 凹头苋 Amaranthus blitum A 29.63 44.44 旋花科 Convolvulaceae 打碗花属 Calystegia 打碗花 Calystegia hederacea A 14.81 虎掌藤属 Ipomoea 牵牛 Ipomoea nil A 14.81 荨麻科 Urticaceae 雾水葛属 Pouzolzia 雾水葛 Pouzolzia zeylanica P 14.81 苎麻属 Boehmeria 苎麻 Boehmeria nivea S 29.63 29.63 74.07 44.44 鸭跖草科 Commelinaceae 鸭跖草属 Commelina 鸭跖草 Commelina communis A 14.81 银杏科 Ginkgoaceae 银杏属 Ginkgo 银杏 Ginkgo biloba T 14.81 罂粟科 Papaveraceae 紫堇属 Corydalis 紫堇 Corydalis edulis A 14.81 鸢尾科 Iridaceae 鸢尾属 Iris 鸢尾 Iris tectorum P 29.63 紫草科 Boraginaceae 附地菜属 Trigonotis 附地菜 Trigonotis peduncularis B 107.41 69.63 212.35 80.00 酢浆草科 Oxalidaceae 酢浆草属 Oxalis 酢浆草 Oxalis corniculata P 19.75 37.04 98.77 59.26 注:A.一年生草本;B.二年生草本;AB.一或二年生草本;P.多年生草本;V.藤本;S.灌木;T.乔木。Notes: A, annual herb; B, biennial herb; AB, annual or biennial herb; P, perennial herb; V, vine; S, shrub; T, tree.
下载: 导出CSV
表 3 不同类型城市公园之间土壤种子库、地上植被Sørensen相似性系数
Table 3 Sørensen similarity coefficients of soil seed banks and aboveground vegetation species among different types of urban parks
公园类型 土壤种子库间物种相似性
Species similarity among soil seed banks地上植被间物种相似性
Species similarity among aboveground vegetations综合公园
Comprehensive park郊野公园
Country park文化遗址公园
Cultural heritage park综合公园
Comprehensive park郊野公园
Country park文化遗址公园
Cultural heritage park郊野公园
Country park0.64 0.62 文化遗址公园
Cultural heritage park0.64 0.57 0.60 0.59 社区公园
Community park0.68 0.56 0.57 0.65 0.64 0.60
下载: 导出CSV
-
[1] Cheng Y Y, Zhang J G, Wei W, et al. Effects of urban parks on residents’ expressed happiness before and during the COVID-19 pandemic[J/OL]. Landscape and Urban Planning, 2021, 212: 104118[2021−10−25]. https://doi.org/10.1016/j.landurbplan.2021.104118.
[2] 陶晓丽, 陈明星, 张文忠, 等. 城市公园的类型划分及其与功能的关系分析: 以北京市城市公园为例[J]. 地理研究, 2013, 32(10): 1964−1976. Tao X L, Chen M X, Zhang W Z, et al. The classification of urban parks and its relationship with functions: a case study of urban parks in Beijing[J]. Geographical Research, 2013, 32(10): 1964−1976.
[3] Özgüner H. Cultural differences in attitudes towards urban parks and green spaces[J]. Landscape Research, 2011, 36: 599−620. doi: 10.1080/01426397.2011.560474
[4] Wang M, Zhang H, Fan S, et al. A zoning-based solution for hierarchical forest patch mosaic in urban parks[J/OL]. Urban Forestry & Urban Greening, 2021, 65: 127352[2022−12−23]. https://doi.org/10.1016/j.ufug.2021.127352.
[5] Morelli F, Benedetti Y, Su T, et al. Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing’s urban parks: effects of land use and vegetation structure[J]. Urban Forestry & Urban Greening, 2017, 23: 84−92.
[6] Wu W, Ding K. Optimization strategy for parks and green spaces in Shenyang City: improving the supply quality and accessibility[J/OL]. International Journal of Environmental Research and Public Health, 2022, 19(8): 4443[2023−01−12]. https://doi.org/10.3390/ijerph19084443.
[7] Brown G, Rhodes J, Dade M. An evaluation of participatory mapping methods to assess urban park benefits[J]. Landscape and Urban Planning, 2018, 178: 18−31. doi: 10.1016/j.landurbplan.2018.05.018
[8] Mata L, Garrard G E, Fidler F, et al. Punching above their weight: the ecological and social benefits of pop-up parks[J]. Frontiers in Ecology and the Environment, 2019, 17: 341−347. doi: 10.1002/fee.2060
[9] 康冰, 王得祥, 崔宏安, 等. 秦岭山地油松群落更新特征及影响因子[J]. 应用生态学报, 2011, 22(7): 1659−1667. doi: 10.13287/j.1001-9332.2011.0236 Kang B, Wang D X, Cui H A, et al. Regeneration characteristics and related affecting factors of Pinus tabulaeformis secondary forests in Qinling Mountains[J]. Chinese Journal of Applied Ecology, 2011, 22(7): 1659−1667. doi: 10.13287/j.1001-9332.2011.0236
[10] 李小双, 彭明春, 党承林. 植物自然更新研究进展[J]. 生态学杂志, 2007, 26(12): 2081−2088. doi: 10.13292/j.1000-4890.2007.0364 Li X S, Peng M C, Dang C L. Research progress on natural regeneration of plants[J]. Chinese Journal of Ecology, 2007, 26(12): 2081−2088. doi: 10.13292/j.1000-4890.2007.0364
[11] Harper J L. Population biology of plants[M]. London: Academic Press, 1977.
[12] Roberts H A. Seed banks in soils[J]. Advances in Applied Biology, 1981, 6: 1−55.
[13] 李彦娇, 包维楷, 吴福忠. 岷江干旱河谷灌丛土壤种子库及其自然更新潜力评估[J]. 生态学报, 2010, 30(2): 399−407. Li Y J, Bao W K, Wu F Z. Soil seed bank and natural regeneration potential of shrubland in dry valleys of Minjiang River[J]. Acta Ecologica Sinica, 2010, 30(2): 399−407.
[14] 卢正宽, 刘贺永, 蹇述莲, 等. 森林草原过渡带持久土壤种子库沿降水梯度的格局[J]. 应用生态学报, 2022, 33(9): 2363−2370. Lu Z K, Liu H Y, Jian S L, et al. Pattern of persistent soil seed bank along precipitation gradient in forest-steppe transition zone[J]. Chinese Journal of Applied Ecology, 2022, 33(9): 2363−2370.
[15] 王东丽, 郭莹莹, 谢伟, 等. 风沙黄土区排土场不同恢复类型植物群落与土壤种子库特征[J]. 水土保持研究, 2022, 29(5): 171−177. Wang D L, Guo Y Y, Xie W, et al. Community and soil seed bank characteristics of different vegetation restoration types in aeolian sandy loess dumps[J]. Research of Soil and Water Conservation, 2022, 29(5): 171−177.
[16] 陈颖颖, 吴自荣, 潘萍, 等. 飞播马尾松林土壤种子库的萌发特征及其与土壤理化性质的关系[J]. 土壤通报, 2016, 47(1): 92−97. Chen Y Y, Wu Z R, Pan P, et al. The germination characteristics of soil seed bank and its relationship with doil properties in aerially-seeded Pinus massoiana plantations[J]. Chinese Journal of Soil Science, 2016, 47(1): 92−97.
[17] 李国旗, 邵文山, 赵盼盼, 等. 荒漠草原区4种植物群落土壤种子库特征及其土壤理化性质[J]. 生态学报, 2019, 39(17): 6282−6292. Li G Q, Shao W S, Zhao P P, et al. Analysis of soil seed bank characteristics and soil physical and chemical properties of four plant communities in a desert steppe region[J]. Acta Ecologica Sinica, 2019, 39(17): 6282−6292.
[18] Basto S, Thompson K, Phoenix G, et al. Long-term nitrogen deposition depletes grassland seed banks[J/OL]. Nature Communications, 2015, 6(1): 6185[2022−12−25]. https://www.nature.com/articles/ncomms7185.
[19] Zhao D J, Sun M Q, Xue Y W, et al. Spatial variations of plant species diversity in urban soil seed banks in Beijing, China: Implications for plant regeneration and succession[J/OL]. Urban Forestry & Urban Greening, 2023, 86: 128012[2023−10−30]. https://doi.org/10.1016/j.ufug.2023.128012.
[20] Zhao D J, Yang Q M, Sun M Q, et al. Urbanization and greenspace effect on plant biodiversity variations in Beijing, China[J/OL]. Urban Forestry & Urban Greening, 2023, 89: 128119[2023−10−30]. https://doi.org/10.1016/j.ufug.2023.128119.
[21] Monteiro M V, Doick K J, Handley P, et al. The impact of greenspace size on the extent of local nocturnal air temperature cooling in London[J]. Urban Forestry & Urban Greening, 2016, 16: 160−169.
[22] 仇宽彪. 北京市五环内城市植被格局及公园绿地生态服务功能价值初步研究[D]. 北京: 中国林业科学研究院, 2011. Qiu K B. Primary study on the urban vegetation pattern and the ecological services valuation of parks within the 5th Ring Road in Beijing[D]. Beijing: Chinese Academy of Forestry, 2011.
[23] 中华人民共和国建筑部. 城市绿地分类标准: CJJ/T85−2017[S]. 北京: 中国建筑工业出版社, 2017. Ministry of Construction of the People’s Republic of China. Classification standard of urban green space: CJJ/T85−2017[S]. Beijing: China Architecture and Building Press, 2017.
[24] 北京市园林绿化局. 北京市公园分类分级管理办法[EB/OL]. (2022−04−15)[2023−03−03]. https://www.beijing.gov.cn/zhengce/gfxwj/202210/W020221008609059716277.pdf. Beijing Municipal Forestry and Parks Bureau. Measures of Beijing for the Administration of Parks by Classification and Classification[EB/OL]. (2022−04−15)[2023−03−03]. https://www.beijing.gov.cn/zhengce/gfxwj/202210/W020221008609059716277.pdf.
[25] 王子尧, 高宇, 王向荣. 北京中心城区城市公园使用状况与影响因素量化研究[J]. 北京规划建设, 2020(2): 88−93. Wang Z Y, Gao Y, Wang X R. A quantitative study on the use status and influencing factors of urban parks in central Beijing[J]. Beijing Planning Review, 2020(2): 88−93.
[26] Warr S J, Kent M, Thompson K. Seed bank composition and variability in five woodlands in south-west England[J]. Journal of Biogeography, 1994, 21(2): 151−168. doi: 10.2307/2845469
[27] Bigwood D W, Inouye D W. Spatial pattern analysis of seed banks: an improved method and optimized sampling[J]. Ecology, 1988, 2: 497−507.
[28] 刘静逸, 牛艳东, 郭克疾, 等. 南洞庭湖杨树清理迹地恢复初期土壤种子库特征及其与土壤因子的关系[J]. 应用生态学报, 2020, 31(12): 4042−4050 doi: 10.13287/j.1001-9332.202012.005 Liu J Y, Niu Y D, Guo K J, et al. Characteristics of soil seed bank and their correlations with soil factors in the early restoration period of Populus deltoides cutting slash in Lake South Dongting, China[J]. Chinese Journal of Applied Ecology, 2020, 31(12): 4042−4050. doi: 10.13287/j.1001-9332.202012.005
[29] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2002: 146−185. Lu R K. Methods for agricultural chemical analysis of soil[M]. Beijing: China Agricultural Science and Technology Press, 2002: 146−185.
[30] Gross K L. A comparison of methods for estimating seed numbers in the soil[J]. Journal of Ecology, 1990,78(4): 1079−1093.
[31] Wang J, Huang L, Ren H, et al. Regenerative potential and functional composition of soil seed banks in remnant evergreen broad-leaved forests under urbanization in South China[J]. Community Ecology, 2015, 16: 86−94. doi: 10.1556/168.2015.16.1.10
[32] Vieira M D, Overbeck G E. Small seed bank in grasslands and tree plantations in former grassland sites in the South Brazilian Highlands[J]. Biotropica, 2020, 52: 775−782. doi: 10.1111/btp.12785
[33] 罗超, 郭小平, 冯昶栋, 等. 乌海周边土壤种子库特征及其与地上植被和土壤因子的关系[J]. 草业学报, 2021, 30(11): 13−28. Luo C, Guo X P, Feng C D, et al. The characteristics of the soil seed bank in Wuhai and surrounding areas and the relationship with vegetation and soil factors[J]. Acta Prataculturae Sinica, 2021, 30(11): 13−28.
[34] 胡安. 滩羊轮牧对典型草原土壤种子库的作用[D]. 兰州: 兰州大学, 2021. Hu A. Rotational grazing of Tan-Sheep affects soil seed bank of typical steppe[D]. Lanzhou: Lanzhou University, 2021.
[35] Bengtsson J. Which species? What kind of diversity? Which ecosystem function? Some problems in studies of relations between biodiversity and ecosystem function[J]. Applied Soil Ecology, 1998, 10: 191−199. doi: 10.1016/S0929-1393(98)00120-6
[36] 李晓鹏, 董丽, 关军洪,等. 北京城市公园环境下自生植物物种组成及多样性时空特征[J]. 生态学报, 2018, 38(2): 581−594. Li X P, Dong L, Guan J H, et al. Temporal and spatial characteristics of spontaneous plant composition and diversity in a Beijing urban park[J]. Acta Ecologica Sinica, 2018, 38(2): 581−594.
[37] 刘庆艳, 王国栋, 姜明, 等. 三江平原沟渠土壤种子库特征及其与地上植被的关系[J]. 植物生态学报, 2014, 38(1): 17−26 doi: 10.3724/SP.J.1258.2014.00002 Liu Q Y, Wang G D, Jiang M, et al. Characteristics of soil seed banks and their relationships with aboveground vegetation in ditches in the Sanjiang Plain[J]. Chinese Journal of Plant Ecology, 2014, 38(1): 17−26. doi: 10.3724/SP.J.1258.2014.00002
[38] Decocq G, Valentin B, Toussaint B, et al. Soil seed bank composition and diversity in a managed temperate deciduous forest[J]. Biodiversity & Conservation, 2004, 13: 2485−2509.
[39] Pearson T R H, Burslem D F R P, Mullins C E, et al. Germination ecology of neotropical pioneers: interacting effects of environmental conditions and seed size[J]. Ecology, 2002, 10: 2798−2807.
[40] Sitvertown J W. Introduction to plantation population ecology[M]. London: Longman, 1982: 126−135.
[41] 张姝萌. 上海城市废弃地杂草种子库季相动态与城乡差异[D]. 上海: 华东师范大学, 2017. Zhang S M. Seasonal dynamics of weed seed bank in wasteland of Shanghai and its difference along urban-rural gradient[D]. Shanghai: East China Normal University, 2017.
[42] 张博, 高尚军, 殷鸣放, 等. 城市近郊林土壤种子库研究[J]. 辽宁林业科技, 2009(3): 5−9. Zhang B, Gao S J, Yin M F, et al. Studies on soil seed bank of forest in suburbs[J]. Liaoning Forestry Science and Technology, 2009(3): 5−9.
[43] 伏凯. 北京山区自然边坡土壤种子库特征研究[D]. 北京: 北京林业大学, 2016. Fu K. Soil seed bank of typical slope in Beijing mountain area[D]. Beijing: Beijing Forestry University, 2016.
[44] 牛瑞芳, 吴铁航, 柴宝峰. 黄土高原亚高山草地退化对土壤种子库的影响[J]. 草地学报, 2021, 29(5): 972−981. Niu R F, Wu T H, Chai B F. Effects of subalpine grassland degradation on soil seed bank in the Loess Plateau[J]. Acta Agrestia Sinica, 2021, 29(5): 972−981.
[45] Paluch J G. Ground seed density patterns under conditions of strongly overlapping seed shadows in Abies alba Mill. stands[J]. European Journal of Forest Research, 2011, 130: 1009−1022. doi: 10.1007/s10342-011-0486-4
[46] Thompson K. Small-scale heterogeneity in the seed bank of an acidic grassland[J]. Journal of Ecology, 1986, 74: 733−738. doi: 10.2307/2260394
[47] Pekas K M, Schupp E W. Influence of aboveground vegetation on seed bank composition and distribution in a Great Basin Desert sagebrush community[J]. Journal of Arid Environments, 2013, 88: 113−120. doi: 10.1016/j.jaridenv.2012.08.013
[48] 贺士元, 邢其华, 尹祖棠,等. 北京植物志[M]. 北京: 北京出版社, 1984. He S Y, Xing Q H, Yin Z T, et al. Flora of Beijing[M]. Beijing: Beijing Publishing Group, 1984.
[49] Tedoradze G, Nakhutsrishvili G, Seip M, et al. Terrain impacts the composition of the persistent soil seed bank: a case study of steep high mountain grasslands in the Greater Caucasus, Georgia[J]. Phytocoenologia, 2020, 50: 47−63. doi: 10.1127/phyto/2019/0321
[50] Zhang D J, Zhang J, Yang W Q, et al. Plant and soil seed bank diversity across a range of ages of Eucalyptus grandis plantations afforested on arable lands[J]. Plant and Soil, 2014, 376: 307−325. doi: 10.1007/s11104-013-1954-z
[51] Thompson K, Grime J P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats[J]. Journal of Ecology, 1979, 67: 893−921. doi: 10.2307/2259220
[52] White E, Tucker N, Meyers N, et al. Seed dispersal to revegetated isolated rainforest patches in North Queensland[J]. Forest Ecology and Management, 2004, 192: 409−426. doi: 10.1016/j.foreco.2004.02.002
[53] Klimkowska A, Diggelen R V, Held S D, et al. Seed production in fens and fen meadows along a disturbance gradient[J]. Applied Vegetation Science, 2009, 12: 304−315. doi: 10.1111/j.1654-109X.2009.01024.x
[54] Guo Y L, Li Y F, Li J X, et al. Comparison of aboveground vegetation and soil seed bank composition among three typical vegetation types in the karst regions of Southwest China[J/OL]. Agronomy, 2022, 12: 1871[2022−12−23]. https://doi.org/10.3390/agronomy12081871.
[55] 孙映通, 夏礼庆, 林琳, 等. 香格里拉亚高山森林带退化群落土壤种子库特征与土壤理化性质分析[J]. 云南大学学报(自然科学版), 2020, 42(5): 1014−1026. Sun Y T, Xia L Q, Lin L, et al. Characteristics of soil seed bank and soil physicochemical properties of degraded communities in Shangri-la subalpine forest belt[J]. Journal of Yunnan University (Natural Sciences Edition), 2020, 42(5): 1014−1026.
[56] 冯璐, 刘京涛, 韩广轩, 等. 黄河三角洲滨海湿地地下水位变化对土壤种子库特征的影响[J]. 生态学报, 2021, 41(10): 3826−3835. Feng L, Liu J T, Han G X, et al. Effects of groundwater level fluctuation characteristics of soil seed banks in coastal wetlands of the Yellow River Delta[J]. Acta Ecologica Sinica, 2021, 41(10): 3826−3835.
[57] Valdés-Rodríguez O A, Sánchez-Sánchez O, Pérez-Vázquez A. Effects of soil texture on germination and survival of non-toxic Jatropha curcas seeds[J]. Biomass and Bioenergy, 2013, 48: 167−170. doi: 10.1016/j.biombioe.2012.10.025
-
期刊类型引用(8)
1. 马丹,汤志伟,马小玉,邵尔辉,黄达沧. 基于GEE的中国不同生态系统林火驱动力研究. 应用科学学报. 2024(04): 684-694 .  百度学术
百度学术
2. 张吕成,孙志超,董灵波. 基于Landsat时间序列数据的火烧迹地识别与恢复效果评价. 森林工程. 2024(05): 8-16 . 百度学术
3. 侯波,李倩倩,杨艳蓉,张乐英. 基于MODIS数据的2003—2020年西南地区林火随地形因子的动态变化. 生态科学. 2024(05): 131-137 . 百度学术
4. 刘海新,钱以临,孔俊杰,张灿,刘韦志. 2003—2019年内蒙古FIRMS_MODIS植被火点时空变化. 林业科技情报. 2023(01): 1-8 . 百度学术
5. 崔阳,狄海廷,邢艳秋,常晓晴,单炜. 基于MODIS数据的2001—2018年黑龙江省林火时空分布. 南京林业大学学报(自然科学版). 2021(01): 205-211 . 百度学术
6. 吴立志,陈振南,张鹏. 基于随机森林算法的城市火灾风险评估研究. 灾害学. 2021(04): 54-60 . 百度学术
7. 孙宝军. 内蒙古电力系统自然灾害链分析. 灾害学. 2020(04): 8-12+47 . 百度学术
8. 潘霞,汪季,高永,王祯仪. 基于MODIS数据的阿拉善盟植被指数变化的地形分异性. 生态环境学报. 2019(02): 226-234 . 百度学术
其他类型引用(9)

计量
- 文章访问数: 434
- HTML全文浏览量: 74
- PDF下载量: 66
- 被引次数: 17
