
Effects of urban heat island intensity on leaf water-relevant traits of greening tree species with different leaf textures
-
摘要:目的
了解两种不同质地叶片水分相关性状对城市热岛的响应,揭示不同质地叶片对城市热岛的适应方式是否有所区别。
方法在北京市4个典型热岛强度地块中,以常见纸质叶和革质叶绿化树种为对象,研究与树木水分过程相关的叶脉、气孔性状以及叶表皮厚度的变化。
结果(1) 纸质叶的叶脉密度、气孔密度、气孔长度、最大气孔导度、上表皮厚度在强热岛区相对于强冷岛区表现出增大的趋势,叶脉直径在强热岛区相对于强冷岛区表现出减小的趋势;(2)革质叶的叶脉密度、叶脉直径、气孔长度在强热岛区相对于强冷岛区表现出增大的趋势,气孔密度、最大气孔导度、上表皮厚度在强热岛区相对于强冷岛区表现出减小的趋势;(3)不同质地叶片表现出了不同的水分相关性状相关性,但两者均表现出气孔密度与最大气孔导度呈正相关(p < 0.05),两种质地叶片通过调整水分相关性状之间不同的关系来适应城市热岛的胁迫。
结论城市热岛会显著影响树木叶片的水分相关性状,不同质地的叶片应对城市热岛环境所采取的适应策略有所区别。纸质叶采取了加大水分供应,提高蒸腾作用,帮助植物体降温的策略;革质叶采取了提高水分的运输,减小蒸腾水分耗散的策略。两种叶片质地树种的叶脉与气孔均存在协同进化的现象。对于处于较高热岛强度的城市公园,应参考水分相关性状等指标选择耐高温树种,并且在高温季节采取加大灌溉量和人工降温等措施。
Abstract:ObjectiveThis paper aims to explore the response of water content correlation between two different leaf textures to urban heat islands, and to reveal whether there are differences in the adaptation methods of varied leaf textures to urban heat islands.
MethodIn four typical heat island intensity plots in Beijing, common chartaceous and coriaceous leaf greening tree species were used as objects to study the changes in leaf vein, stomatal traits, and leaf epidermal thickness related to tree water processes.
Result(1) The leaf vein density, stomatal density, stomatal length, maximum stomatal conductivity, and upper epidermal thickness of chartaceous leaves showed an increasing trend in the strong heat island area compared with the strong cold island area, while the leaf vein diameter showed a decreasing trend in the strong heat island area compared with strong cold island area. (2) The vein density, vein diameter, and stomatal length of coriaceous leaves showed an increasing trend in the strong heat island area compared with strong cold island area, while the stomatal density, maximum stomatal conductivity, and upper epidermal thickness showed a decreasing trend in the strong heat island area compared with strong cold island area. (3) Different leaf textures showed varied water related correlations, but both showed a positive correlation between stomatal density and maximum stomatal conductance (p < 0.05). The two types of leaf textures adapted to urban heat island stress by adjusting the different relationships between water related traits.
ConclusionUrban heat islands significantly affect the water related traits of tree leaves, and different textures of leaves adopt different adaptation strategies to the urban heat island environment. Chartaceous leaves adopt a strategy of increasing water supply, improving transpiration, and helping plants cool down. The coriaceous leaves adopt a strategy of increasing water transportation and reducing transpiration water dissipation. There is a phenomenon of co-evolution between the leaf veins and stomata of two tree species with different leaf textures. For urban parks with high heat island intensity, high-temperature resistant tree species should be selected based on indicators such as water correlation, and measures such as increasing irrigation and artificial cooling should be taken during the high-temperature season.
-
松栎柱锈菌(Cronartium orientale)是导致樟子松(Pinus sylvestris var. mongolica)、兴凯湖松(Pinus takahasii)和马尾松(Pinus massoniana)等2针松瘤锈病的森林有害生物,是一种转主寄生菌,其春孢子阶段寄生在松树枝干部[1-2]。当松树被松栎柱锈菌侵染致病后,其病原菌就会长期寄生在松树枝干部,并且病部肿瘤会逐年增大,给林分健康和林木生长造成极大危害[3]。黄酮类化合物具有较高的生物活性,是一种强抗氧化剂,能有效清除自由基从而延缓衰老和阻止细胞衰老、癌变,对人类心脑血管疾病也有很好的治疗和预防效果,具有较高的医疗和保健作用[4-8]。已有较多研究成果表明松科植物树皮与松针中黄酮类化合物含量丰富,具有较好的开发利用潜力[9-10]。国内学者如李石清等对南方红豆杉(Taxus chinensis var. mairei)枝叶总黄酮的研究、冯慧英等对巴山冷杉(Abies fargesii)总黄酮的研究、王飞等对青扦(Picea wilsonii)针叶总黄酮的研究等均表明其易提取且具有较强的抗氧化能力[11-13]。
担子菌门(Basidiomycota)的许多菌类代谢产物已经被开发利用,关于担子菌的研究已经成为当代生命科学、食品科学和医药领域的热点[14-15]。锈菌目(Uredinales)隶属于担子菌门,目前却尚未发现任何关于锈菌目真菌开发利用的研究,这也表明我们对于锈菌目真菌的潜在价值知之甚少。本研究选择了锈菌目中寄生在松科植物上的易于采集的松栎柱锈菌春孢子为研究对象,进行了松栎柱锈菌春孢子黄酮类化合物(以下简称CQF)的提取与体外抗氧化活性的研究,旨在为锈菌目菌物开发利用、森林有害生物开发利用和提升与拓宽森林有害生物防治理念提供科学依据。
2. 材料与方法
2.1 研究材料与预处理
松栎柱锈菌春孢子于2015年5月中旬采自大兴安岭地区,过80目筛后于45℃烘干至恒质量,密封冷藏。所用试剂盒均购自南京建成生物研究所;芦丁标准品,购自上海源叶生物科技有限公司;其他试剂均为国产分析纯;KQ-600B型超声波仪,昆山市超声波仪器有限公司;分析天平(感应量0.0001g);Gene Quant 1300型分光光度计,美国GE公司。
2.2 测定波长的选择与标准曲线的建立
精密称取芦丁标准品10.0mg,使用体积分数为75%的乙醇溶液配制成50mL 0.2mg/mL的标准溶液。取上述标准溶液稀释为质量浓度为0.04,0.06,0.08,0.10,0.12,0.14,0.16,0.18,0.20mg/mL的芦丁溶液,分别吸取5mL上述各质量浓度芦丁溶液,置于1,2,3,4,5,6,7,8,9号共9支25mL容量瓶中,分别加入5%亚硝酸钠(NaNO2)溶液1.0mL,摇匀后室温下静置6min,再分别加入10%硝酸铝(Al(NO3)3)溶液1.0mL,摇匀后室温下静置6min,再加入1mol/L的氢氧化钠(NaOH)溶液8mL,用75%乙醇溶液定容至25mL。显色15min后取5号容量瓶进行紫外全波长扫描,以确定吸收峰。用蒸馏水调零后在300~900nm范围内的扫描,截取其在400~650nm范围内的吸光度变化示意图,结果如图 1所示。可知经过本方法处理过的黄酮类化合物在500~520nm处出现了明显的吸收峰,因此判断本方法能够用于CQF的测定。为减小红移和蓝移对检测结果的影响,选择510nm作为测定波长。
不同质量浓度芦丁标准品溶液按照本方法操作后,在510nm处测定各质量浓度对应的吸光度A,以芦丁标准品质量浓度(mg/mL)为横坐标,吸光度A值为纵坐标,绘制标准曲线,结果如图 2所示。标准曲线的回归方程为y=2.5617x-0.001,回归方程的相关系数R2=0.9995,说明该检测方法可行,所得回归方程能够用来测定CQF质量浓度。
2.3 单因素试验
设定松栎柱锈菌春孢子黄酮类化合物的初始提取条件为:提取温度A=40℃、料液比B=1:40、提取时间C=1h、乙醇体积分数D=50%(v/v);每次试验所用春孢子粉200mg;超声频率40kHz,超声功率600W。将所得提取液5000r/min离心8min,吸取上层清液按照2.2的方法进行测定样品吸光值A。根据2.2中所得芦丁标准曲线:y=2.5617x-0.001推算CQF提取液中CQF的质量浓度,并通过式(1)计算CQF的得率。采用控制变量法依次考察该提取条件下上述4个单因子对CQF得率的影响。
T=NnVm×100% (1) 式中:T为CQF得率,%;N为根据标准曲线计算出的CQF的质量浓度,mg/mL;n为稀释倍数;V为提取液的体积,mL;m为供试春孢子质量,mg。
2.4 得率建模试验
在单因子考察试验基础上,根据中心组合实验(central composite design)原理使用软件Design-Expert 8.0.6对提取温度(A),料液比(B),提取时间(C),乙醇体积分数(D)这4个因子较优的水平进行了CQF得率的响应面建模试验。完成建模试验后使用建模试验数据建立CQF得率关于以上4个影响因素的二次多项式回归方程数学模型,对所得模型及各个影响因子进行方差分析。通过软件Design-Expert 8.0.6与所得数学模型得出最优CQF提取条件,并验证优化结果。建模试验各因素水平与实际值对应关系见表 1。
表 1 建模试验因子水平表Table 1. Factors and levels of flavonoid extraction experiment水平
Level影响因子Influencing factor 温度Temperature(A)/℃ 料液比Solid-liquid ratio(B)/(g·mL-1) 时间Time(C)/min 乙醇体积分数
Ethanol contentration(D)/%-2 35 1:100 20 50 -1 45 1:120 30 60 0 55 1:140 40 70 1 65 1:160 50 80 2 75 1:180 60 90 2.5 CQF抗氧化能力测定
除CQF的还原力与DPPH·清除能力之外,清除超氧阴离子(O2-·)活性、清除羟基自由基(·OH)活性和总抗氧化活性均采用从南京建成生物研究所购买的对应的试剂盒测定,并严格按照试剂盒使用说明执行。本文所有CQF体外抗氧化活性测定实验均以抗坏血酸(Vc)做阳性对照,用以评价CQF的体外抗氧化活性。
2.5.1 CQF还原力测定
用体积分数为50%的乙醇溶液将在建模试验中所收集的CQF溶液稀释为原来质量浓度的0.2、0.4、0.6、0.8和1.0倍,分别向装有1.0mL各质量浓度CQF溶液的试管中加入0.2mol/L的pH 6.6的磷酸缓冲溶液2.5mL,快速混匀后加入质量分数1%的铁氰化钾溶液2.5mL并在50℃下水浴20min。迅速冷却反应液并加入质量分数10%的三氯乙酸溶液2.5mL,充分混匀,3000r/min离心10min后取上清液3mL,依次加入3mL蒸馏水和1.0mL的0.1%的三氯化铁溶液,充分混匀后静置10min,于700nm波长下测定溶液吸光度。吸光度越大还原能力也越大,计算公式如(2)。
A700=A样品−A空白 (2) 2.5.2 DPPH·清除能力测定
用95%乙醇溶液配置DPPH·浓度为0.2mmol/L的乙醇溶液,置于冰箱中避光冷藏备用。测定方法:测定管用2mL DPPH·乙醇溶液中加入2mL各质量浓度的CQF样品溶液,CQF质量浓度设置梯度与2.5.1相同。对照管用95%乙醇代替DPPH·溶液;空白管用蒸馏水代替CQF溶液。混匀器混匀后避光反应30min。波长调至517nm处用0.5mL蒸馏水和2.5mL 95%的乙醇调零,然后测定反应液吸光值A,每个质量浓度设置3个平行实验,结果取平均值。DPPH·自由基的清除率通过公式(3)计算得出。
DPPH⋅清除率=A0−(A1−A2)A0×100% (3) 式中:A0为2mL DPPH·乙醇溶液+2mL超纯水的吸光值;A1为2mL DPPH·乙醇溶液+2mL样品溶液后的吸光值;A2为2mL样品溶液+2mL超纯水的吸光值。
3. 结果与分析
3.1 单因子试验
3.1.1 乙醇体积分数对CQF得率的影响
由图 3可知,在其他影响因子不变的条件下,乙醇体积分数在50%~90%范围内变化导致CQF的得率先逐渐增大,当乙醇体积分数增至70%的时候,得率最大,但是仍然不足0.5%。当乙醇体积分数超过70%以后,随着体积分数的增加导致CQF得率骤降并逐渐趋于稳定。黄酮类化合物难溶于水,易溶于有机溶剂,水和乙醇可以以任意比例互溶。在水中加入不同量的乙醇,溶液的极性也随之不同,因而对某种或某类物质的溶解能力也不相同。根据所得实验结果绘制的折线图可以看出乙醇体积分数为70%是较优水平。
![]() 图 3 乙醇体积分数对黄酮类化合物(CQF)得率的影响Figure 3. Effects of ethanol concentration on extraction rate of flavonoids
图 3 乙醇体积分数对黄酮类化合物(CQF)得率的影响Figure 3. Effects of ethanol concentration on extraction rate of flavonoids3.1.2 料液比对CQF得率的影响
根据图 4可以看出在其他条件不变的情况下,随着料液比的减小,CQF得率在不断升高,并逐渐趋于平稳,在料液比达到1:140以后,CQF得率增长幅度变小,得率增幅减小,因此选择1:140为较优水平。这一结果与CQF在50%乙醇溶液中的溶解度相关,当料液比较大时,CQF只有少部分溶于了乙醇溶剂,此时的提取液处于一种近饱和状态;随着提取液的增多,也就是料液比减小,未溶出的CQF部分也逐渐溶出,导致CQF的得率上升。当料液比减小到一定程度时,能溶于50%乙醇溶液的部分溶解殆尽,所以这时的得率逐渐趋于稳定,靠近极值。
![]() 图 4 料液比对CQF得率的影响Figure 4. Effects of solid-liquid ratio on extraction rate of flavonoids
图 4 料液比对CQF得率的影响Figure 4. Effects of solid-liquid ratio on extraction rate of flavonoids3.1.3 提取温度对CQF得率的影响图
在其他试验条件不变的条件下,考察了温度在15~75℃之间变化对CQF得率的影响,通过考察发现CQF得率先随着温度升高而增大,随着温度升高至65℃以后又逐渐降低。当提取液温度较低时,分子获得的能量相对较少,分子运动缓慢,有效提取CQF耗费的时间增长,这一点可以通过分子扩散现象很好地解释。但是,温度的变化同样会影响溶剂分子之间的作用力,从而影响溶剂溶解溶质的能力,导致CQF得率也随之发生变化。也就是说温度的变化同时影响着CQF分子运动速率与溶剂,因而CQF得率受此二者的交互作用影响,CQF得率随提取温度的变化如图 5所示。
![]() 图 5 提取温度对CQF提取率的影响Figure 5. Effects of extraction temperature on extraction rate of flavonoids
图 5 提取温度对CQF提取率的影响Figure 5. Effects of extraction temperature on extraction rate of flavonoids3.1.4 提取时间对CQF得率的影响
在其他试验因子不变的条件下,考察提取时间对CQF得率的影响,结果显示提取时间对CQF得率的影响微弱,时间在20~60min变化时,CQF得率的变化始终处在0.46%~0.468%之间。CQF分子溶解到提取液中需要一定的时间,所以在提取时间40min以内时,CQF得率随着时间增加而增高,随着提取时间的延长,CQF的得率反而逐渐开始降低。这或许是因为部分原生质体内的CQF分子进入了细胞壁,因为细胞壁本身就可以容纳黄酮类化合物,另一方面,松栎柱锈菌春孢子表面具有油球,对黄酮类化合物具有一定的溶解能力。除此之外,随着溶剂分子运动时间的增加,乙醇分子扩散增多引起溶剂中乙醇浓度降低,从而导致CQF提取率也降低。CQF得率随提取时间的变化如图 6所示。
![]() 图 6 提取时间对CQF提取率的影响Figure 6. Effects of extraction time on extraction rate of flavonoids
图 6 提取时间对CQF提取率的影响Figure 6. Effects of extraction time on extraction rate of flavonoids3.2 响应面法分析
3.2.1 建模试验设计及结果
使用软件Design-Expert 8.0.6对提取温度(A),料液比(B),提取时间(C),乙醇体积分数(D)这4个因子的较优水平进行CQF得率的响应面建模试验。试验因子水平设计如表 1所示,中心组合实验设计与结果见表 2。
表 2 中心组合实验设计与结果Table 2. Design and results of central composite design试验号
Run No.温度
Temperature(A)/℃料液比
Solid-liquid ratio(B)/(g·mL-1)时间
Time(C)/min乙醇体积分数
Ethanol concentration(D)/%得率
Yield(T)/%1 0 -2 0 0 1.6329 2 1 -1 -1 1 1.0071 3 -1 1 1 1 0.9619 4 1 1 1 -1 1.4116 5 2 0 0 0 1.2952 6 0 0 0 0 1.1149 7 -1 1 1 -1 1.1555 8 0 0 0 0 1.1586 9 1 -1 1 1 1.1196 10 0 0 0 0 1.0930 11 1 1 -1 -1 1.3928 12 0 0 0 2 0.7542 13 0 0 0 0 1.1477 14 -1 -1 1 1 0.9697 15 1 -1 -1 -1 1.2507 16 -1 -1 -1 -1 1.1524 17 -1 -1 -1 1 0.8713 18 1 1 1 1 1.1805 19 -1 1 -1 1 0.9993 20 0 2 0 0 1.1453 21 -1 -1 1 -1 1.1055 22 0 0 -2 0 1.2133 23 1 -1 1 -1 1.3959 24 0 0 2 0 0.1969 25 1 1 -1 1 1.0555 26 0 0 0 -2 1.1040 27 0 0 0 0 1.1258 28 -2 0 0 0 1.3675 29 0 0 0 0 1.1477 30 -1 1 -1 -1 1.2304 3.2.2 数学模型的建立与分析
以CQF的得率为响应值,通过使用软件Design-Expert 8.0.6对得率T与4个影响因子回归拟合之后,得到CQF关于4个影响因子的数学模型方程:
T=1.13+0.051A−0.019B−0.071C−0.11D+1.171e−3AB+0.029AC−0.015AD−0.017BC−3.513e−3BD+0.016CD+0.059A2+0.073B2−0.098C2−0.042D2 对该数学模型进行方差分析(表 3)可知该模型回归显著(P<0.05),失拟项不显著(P=0.149),说明未知因素对CQF得率影响较小,模型拟合效果良好。在我们设置的4因子水平梯度范围内,乙醇体积分数D对于CQF得率的影响显著(P<0.05),这与黄酮类化合物易溶于有机溶剂难溶于水的化学性质密不可分。由F值可得出各个影响因子对CQF得率的影响程度大小顺序为:乙醇体积分数(D)>提取时间(C)>提取温度(A)>料液比(B)。
表 3 数学模型方差分析Table 3. Parameter estimate of regression model and variance analysis方差来源
Variance source平方和Sum
f squares自由度
df均方
Mean squareF值
F valueP值
P value显著性
Significance模型Model 1.150 14 0.082 7.469 0.043 1 显著Significant 温度Temperature(A) 0.062 1 0.062 1.520 0.237 1 料液比Solid-liquid ratio(B) 0.009 1 0.009 0.210 0.650 1 时间Time(C) 0.120 1 0.120 2.900 0.109 1 乙醇体积分数Ethanol concentration(D) 0.290 1 0.290 7.010 0.018 3 AB 0.000 1 0.000 0.001 0.981 9 AC 0.013 1 0.013 0.320 0.577 2 AD 0.004 1 0.004 0.093 0.765 2 BC 0.005 1 0.005 0.120 0.736 6 BD 0.000 1 0.000 0.005 0.945 7 CD 0.004 1 0.004 0.100 0.756 6 A2 0.095 1 0.095 2.300 0.149 8 B2 0.150 1 0.150 3.580 0.078 1 C2 0.260 1 0.260 6.380 0.023 3 D2 0.048 1 0.048 1.160 0.297 5 残差Residual 0.620 15 0.041 失拟项Lack of fit 0.610 10 0.061 0.017 0.149 不显著Not significant 纯误差Pure error 0.003 5 0.001 总误差Total error 1.770 29 3.2.3 交互项对CQF得率的影响
响应面是以响应值与影响响应值的因子构建起了空间三维图形,可以让我们更直观的判断出两两不同影响因子交互作用对响应值的影响趋势以及响应值的变化范围。响应面图中颜色的变化表示响应值(CQF得率)的变化,颜色变化快,响应值变化也快,一般较低的响应值使用蓝色表示,较高的响应值用红色表示,其他颜色均为中间过渡颜色。同时,在一定方向和范围内响应面3D图形中坡度越大表示响应值变化的越快,即对CQF得率的影响更为显著。这一点在等高线图形中也有很好的体现,当某条线段在垂直其中一条轴的方向上移动时,在一定范围内与它所相交的等高线越多,则说明在该范围内某一因子对响应值的影响越显著。等高线表示在此条线上所有试验方案都能够得到相同的响应值,即虽然等高线所对应的试验条件不同,但是能够产生相同的实验结果。4个影响因子的两两之间交互作用对CQF得率的影响示意图见图 7。
![]() 图 7 各因素交互作用对CQF得率影响的响应面与等高线图Figure 7. Response surface and contour plots for the interaction effects of various factors on extraction rate of CQF
图 7 各因素交互作用对CQF得率影响的响应面与等高线图Figure 7. Response surface and contour plots for the interaction effects of various factors on extraction rate of CQF3.2.4 最佳提取工艺及验证
根据在3.2.2中对模型进行的方差分析,已经得知我们拟合出的回归方程模型显著,失拟项不显著,说明该方程是适合用于CQF得率与提取工艺的数学模型,所以我们可以借助于该方程确定最佳的CQF提取工艺。经软件Design-Expert 8.0.6分析,得知模型预测的最佳提取工艺为:提取温度65℃,料液比1:120,提取时间38min,提取液乙醇体积分数60%。在此条件下有对该优化结果进行了3次验证实验,3次验证实验所得出的CQF得率为1.382%,实际得率与在该条件下模型所预测的得率1.416%相差2.46%,这表明我们优化出来的CQF提取条件真实可靠,能够应用于实践。
3.3 抗氧化活性分析
3.3.1 CQF的还原力测定
还原力是评价物质抗氧化能力的重要指标之一,其原理是具有较强还原力的物质将Fe3+还原为Fe2+,Fe2+在酸性环境中能与FeCl3产生有颜色的普鲁士蓝。在该检测体系下物质的还原力越强,对应的吸光值也越大。根据图 8可以看出,CQF与Vc的还原力与质量浓度均呈现很好的线性关系,拟合度均超过0.998。在图示浓度范围内,CQF和Vc的质量浓度与还原力均表现出较好的量效关系,相同质量浓度条件下CQF的还原力更强,而且CQF质量浓度与还原力的线性方程斜率更大,量效关系更为明显。在图示范围内,不论是量效关系还是还原力,CQF都强于Vc,说明CQF具备很好的还原力。
3.3.2 CQF对DPPH·的清除能力
在图 9中,根据受试物质量浓度与DPPH·清除率的散点图,我们可以看出Vc对DPPH·的清除能力较CQF强,并迅速逼近清除率最大值。在散点图的基础上,可以得出CQF与Vc清除DPPH·的IC50分别为0.0492mg/mL和0.0335mg/mL,也就是说对CQF对DPPH·清除能力较Vc差。但是随着CQF质量浓度的逐步增大,CQF对DPPH·清除能力也逐步增强,量效关系越来越明显。对于该种人工合成自由基,CQF的反应不够灵敏。
![]() 图 9 受试物清除DPPH·能力的比较Figure 9. Comparison of DPPH· radical scavenging capacity of CQF and Vc
图 9 受试物清除DPPH·能力的比较Figure 9. Comparison of DPPH· radical scavenging capacity of CQF and Vc3.3.3 CQF总抗氧化能力
生物活性物质总抗氧化能力是指活性物质清除各种自由基的有效总和。虽然已有多种测定抗氧化活性的方法,但这些方法只是从某个角度来衡量天然抗氧化物的活性。事实上,抗氧化作用涉及到多种作用机制,它涉及一系列的问题,目前尚未形成标准方法[16]。本实验测定总抗氧化能力采用的南京建成生物研究所试剂盒原理为FRAP法,在该种检测方法下,由图 10可以看出CQF与Vc的总抗氧化能力相当。根据散点图经计算得知,CQF的半抑制率IC50为0.0303mg/mL,而Vc的半抑制率为0.0240mg/mL,可以看出Vc的总抗氧化能力较CQF强,但是CQF与Vc趋势线在0.0654mg/mL处相交。整体来看,两种物质达到最大清除率都不需要较大浓度,量效关系相近,其总抗氧化能力并不具有很大的差异。所以说,CQF的总抗氧化能力还是非常理想的。
3.3.4 CQF对·OH的清除能力
羟基自由基(·OH)属于一种强氧化剂(E0=2.80V,3.06V),氧化能力远大于一般氧化剂,是机体中最活跃的一种活性氧分子,具有很强的破坏性。它几乎可以和活细胞中的任何生物分子发生反应而造成损伤,而且反应速度极快[17]。因此,清除机体内过多的·OH具有非常重要的生物学意义。通过与Vc的对比测试,我们可以看出Vc在质量浓度0.016 ~0.080mg/mL范围内呈现线性变化规律,但对·OH较强的清除能力远低于CQF。由图 11可以非常明显的看出Vc与CQF在·OH清除能力上的差距。虽然Vc具有很强的还原性,不仅能够直接清除·OH,还能够阻断Fenton反应,从而减少体系中的·OH,但是CQF对·OH的敏感程度与清除能力依然远胜Vc。经测定,CQF对·OH清除能力的IC50仅为0.0129mg/mL,因为Vc对·OH的清除能力远小于CQF,故未予测定与计算其对·OH的半抑制率。
3.3.5 CQF对O2-·的清除能力
超氧阴离子O2-·经过一系列反应可以生成其他氧自由基,是大多数氧自由基的母体,能够攻击细胞DNA,具有较强的毒性,所以清除生物机体内过多的O2-·具有重要意义。理论上来讲,CQF能够提供电子从而有效清除自由基,但本课题在测定CQF清除O2-·能力时却发现了截然相反的实验结果。由图 12可知,两种受试物对清除O2-·都具有一定的量效关系,但是测定结果不在同一象限内。这也就是说在该中检测体系下受试物CQF非但没有清除O2-·,反而促进了O2-·的产生。所使用试剂盒检测原理为模拟机体中黄嘌呤与黄嘌呤氧化酶反应系统,黄嘌呤在黄嘌呤酶的催化下转化为尿酸,在此过程中能够产生O2-·,然后通过衡量O2-·的量来推算加入反应体系的物质时清除还是产生O2-·。黄酮类化合物作为电子提供物质,不会产生O2-·,但是CQF提取液是未经过除杂的,一些会对该反应体系产生影响的物质可能一并被提取了,比如说嘌呤或黄嘌呤氧化酶以及一些自氧化物质,这些物质生成O2-·的能力远超黄酮类物质清除O2-·的能力,所以才产生了如图 12所示的测定结果[18-20]。这一问题没有得到有效解决,所以尚不清楚导致该检测结果的具体原因,需要在后续工作中进一步研究导致该结果的具体原因与解决方法。黄嘌呤酶氧化法目前是测定O2-·最完善最普遍的方法,但是这种方法应该对待测样品设置一定的要求或合适的处理条件以排除干扰因子,需要我们进一步完善这种测定方法。
4. 结论与讨论
从微生物孢子中提取黄酮类化合物的例子尚不多见,在此背景下,本研究采用控制变量法逐个考察了能够影响CQF得率的4个因子。然后使用软件Design-Expert 8.0.6设计了建模试验,完成建模试验之后利用可靠的CQF得率数学模型得到了适用的最优CQF提取条件:提取温度65℃,料液比1:120,提取时间38min,提取液乙醇体积分数60%,在该提取条件下CQF得率为1.382%。虽然松栎柱锈菌春孢子具有厚且坚硬的几丁质细胞壁,能够有效阻止细胞内含物的流失以抵抗不良环境因素,但是我们使用一般植物总黄酮提取的方法并辅以超声振荡也提取到了CQF,说明醇提取法同样能够用于其他孢子总黄酮的提取并且达到了预期的CQF得率。总黄酮得率既取决于提取方法,也取决于实验材料总黄酮含量。一般来说针叶树实验材料总黄酮得率高于菌物但不足5%。闫琳娜通过对落叶松(Larix gmelinii)松针黄酮的研究发现落叶松松针总黄酮含量较高且在不同提取条件下得率都能达到6%以上[21];孙琼优化了在菌物中黄酮类化合物含量相对较高的杏鲍菇(Pleurotus eryngii)黄酮类化合物的提取条件,但是得率仍然不足3%[22]。同时我们也看到了黄嘌呤酶氧化法在实际应用中暴露出来的问题,没能有效测定出CQF清除O2-·的能力,需要进一步明确并排除干扰因子或者改进O2-·检测方法。
黄晓冬等对阔叶树种白桦(Betula platyphylla)活体寄生真菌桦褐孔菌(Inonotus obliquus)总黄酮的研究表明其具有较强的体内外抗氧化活性[23],Boonsong等研究了5种食用菌总酚与类黄酮的提取与抗氧化活性,发现乙醇溶液更适合菌物总酚的提取,其提取物均表现出较强的抗氧化活性,然而很多的食用菌不是活体寄生或不含有黄酮类物质[24]。整体来看,对于针叶树总黄酮的研究成果与热情远胜担子菌,而活体寄生的担子菌总黄酮的研究很鲜见。本研究所选研究对象将针叶树与担子菌门菌物总黄酮的研究很好地有机结合,旨在为锈菌目菌物开发利用、森林有害生物开发利用和转变森林有害生物防治理念提供科学依据。迄今为止,人类对锈菌的认知还很不完善,尚未看到国内外任何关于锈菌开发利用的报道。本研究首次完成了CQF的提取与体外抗氧化活性测定工作,发现CQF具有很强的还原力,还原力较Vc强;其次CQF的总抗氧化能力和DPPH·清除能力与Vc相当;CQF对·OH具有非常好的清除作用,这一能力远胜Vc,与Treml等对黄酮类化合物清除·OH能力的研究现状综述相一致[26]。体外试验结果为体内试验提供了一定的科学依据,这表明CQF具有开发利用的潜在价值。
长久以来,人类对于有害生物的认识是不全面的,往往只是单纯的从治理的角度去对待,总是尝试使用各种各样的手段降低或控制有害生物的种群数量。森林有害生物确实为林业生产活动和森林健康产生了很多负面的影响,如果说发现并证实某种有害生物有其独特的利用价值,那么它的某种功能或价值就能在一定程度上抵偿它所造成的损失,从而引导我们转变传统的有害生物防治理念。
-
![]()
图 2 热岛强度对不同叶片质地树木叶片水分相关性状的影响
CL为革质叶树种,CHL为纸质叶树种,ATS为所有树种。大写字母表示同一热岛梯度上的两种质地叶片性状在p < 0.05 水平上差异显著,小写字母则表示同种叶片质地树木叶片性状在不同热岛梯度间在 p < 0.05 水平上存在显著差异。*和***分别代表纸质叶与革质叶的性状指标间在p < 0.05和p < 0.001水平存在显著差异。下同。CL, coriaceous leaf species; CHL, chartaceous leaf species; ATS, all tree species. Capital letters indicate significant differences in leaf traits of tree species with coriaceous leaf and chartaceous leaf in the same heat island gradient at the level of p < 0.05, while lowercase letters indicate significant differences among different heat island gradients for leaf traits of tree species with the same leaf texture at the level of p < 0.05. * and *** represent significant differences between mean values of chartaceous and coriaceous leaves at the levels of p < 0.05 and p < 0.001, respectively. Same as below.
Figure 2. Effects of heat island intensity on water-relevant traits of tree leaves with different leaf textures
![]()
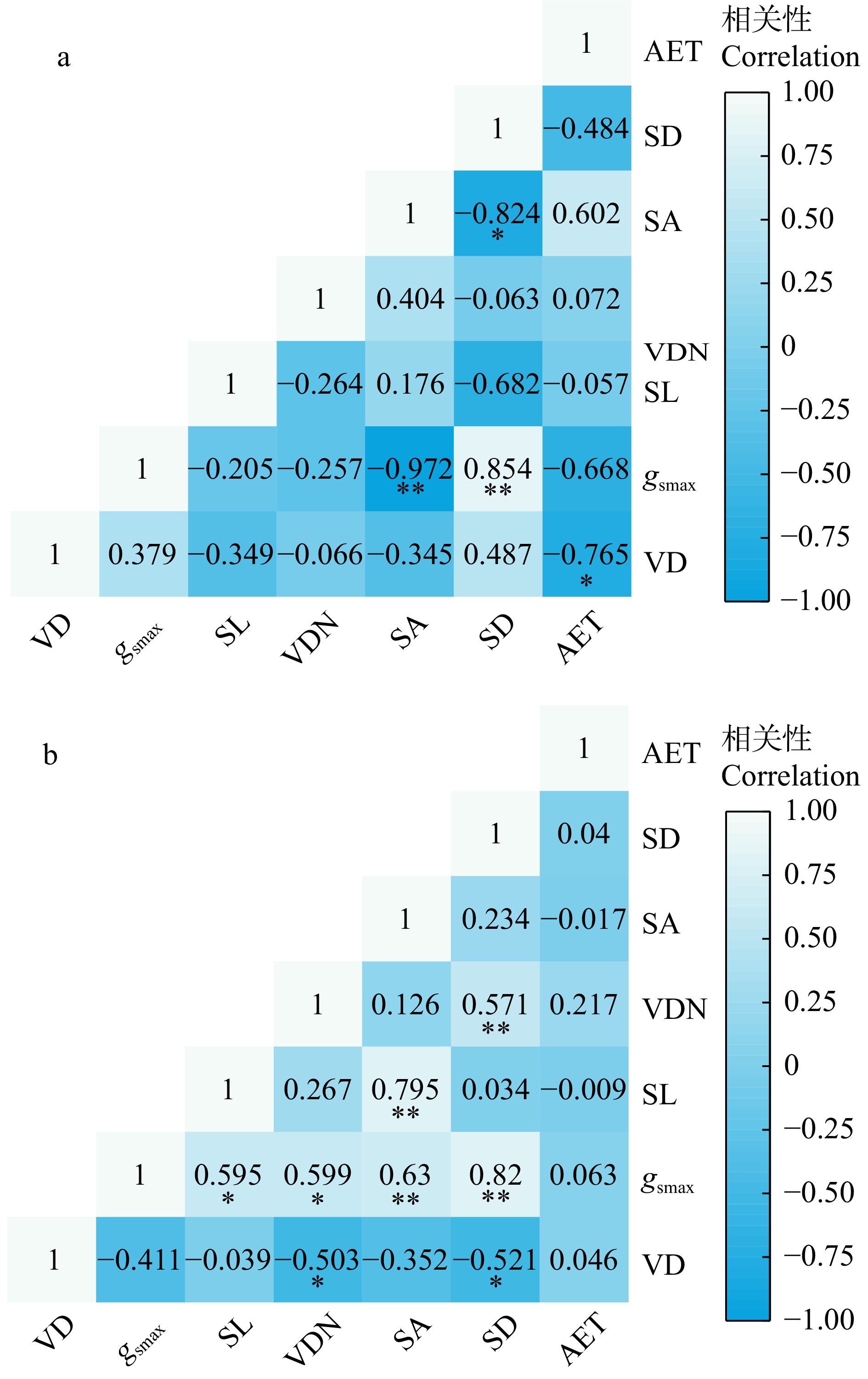
图 3 革质叶(a)和纸质叶(b)树种叶片水分相关性状的Pearson相关性
*和**分别代表纸质叶与革质叶的性状指标在p < 0.05和 p < 0.01水平存在显著差异。VD. 叶脉直径;gsmax. 最大气孔导度;SL. 气孔长度;VDN. 叶脉密度;SA. 气孔聚集度;SD. 气孔密度;AET. 上表皮厚度。下同。* and ** represent significant differences between mean values of coriaceous and chartaceou leaves at the levels of p < 0.05 and p < 0.01, respectively. VD, vein diameter; gsmax, maximum stomatal conductance to water vapour; SL, stomatal length; VDN, vein density; SA, stomatal aggregation; SD, stomatal density; AET, adaxial epidermis thickness. Same as below.
Figure 3. Pearson correlations of leaf water related traits in tree species with coriaceous (a) and chartaceou (b) leaves
![]()
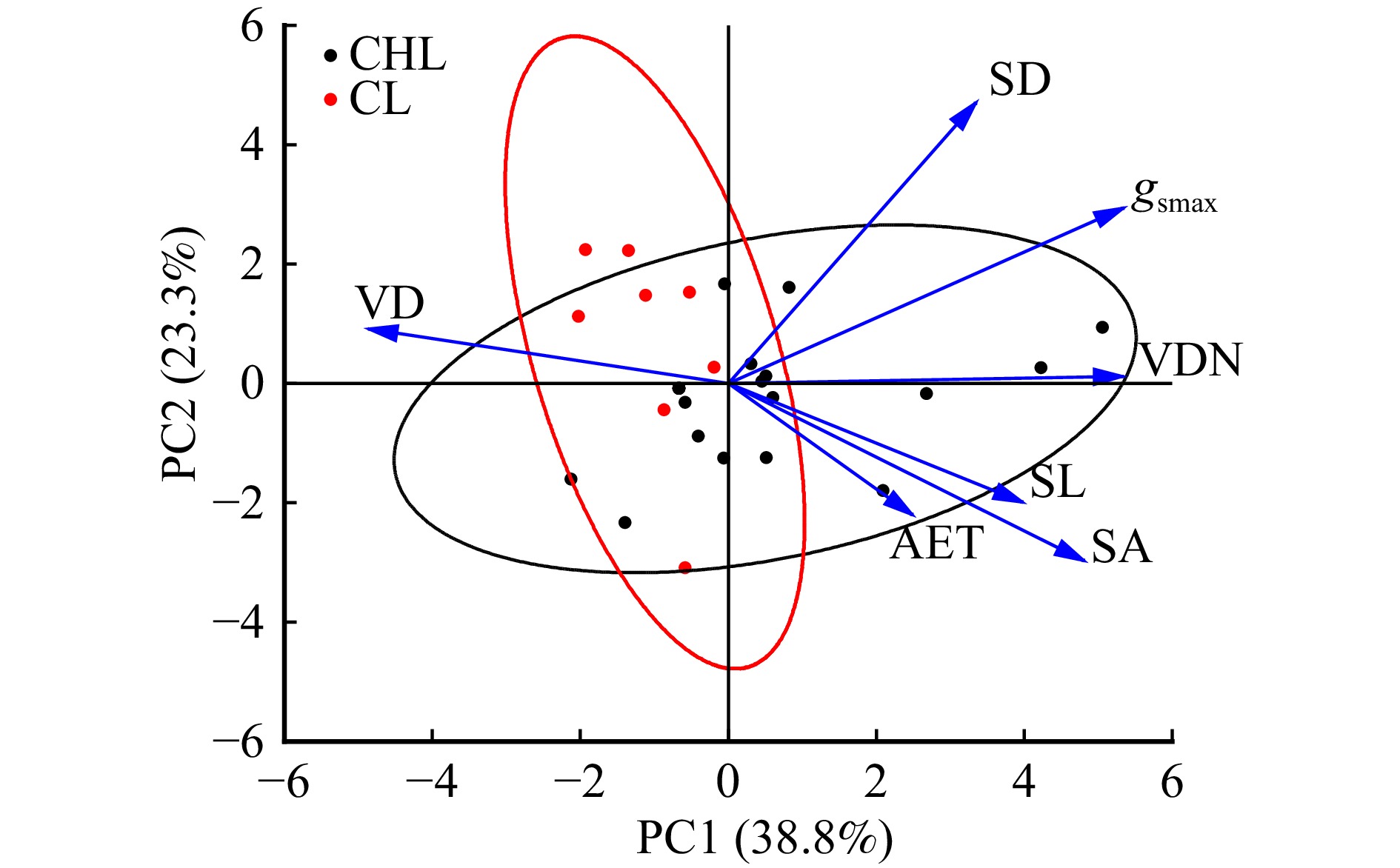
图 4 全部树种叶片水分相关功能性状的主成分分析
Figure 4. Principal component analysis of water-relevant traits of leaves of all tree species
表 1 热岛强度和叶片质地对叶片水分相关功能性状影响的方差分析
Table 1 Variance analysis on effects of heat island intensity and leaf textures on water-relevant traits of leaves of tree species
项目
Item指标
Index叶脉直径
Vein
diameter叶脉密度
Vein
density气孔密度
Stomatal
density气孔聚集度
Stomatal
aggregation最大气孔导度
Maximum stomatal
conductance气孔长度
Stomatal
length上表皮厚度
Adaxial epidermis
thickness热岛强度
Heat island intensityF 8.503 14.156 0.436 0.366 1.617 2.458 0.505 p < 0.001 < 0.001 0.727 0.778 0.184 0.062 0.679 叶片质地
Leaf textureF 5.716 21.591 0.041 1.882 1.783 5.922 29.794 p 0.017 < 0.001 0.840 0.173 0.182 0.015 < 0.001 变异系数
Variation coefficient/%57.98 28.38 22.61 5.24 25.71 15.25 25.54  下载: 导出CSV
下载: 导出CSV
-
[1] Esperon-Rodriguez M, Baumgartner J B, Beaumont L J, et al. Climate-change risk analysis for global urban forests[J]. Cold Spring Harbor Laboratory, 2021. DOI: 10.1101/2021.05.09.443030.
[2] Esperon-Rodriguez M, Tjoelker M G, Lenoir J, et al. Climate change increases global risk to urban forests[J]. Nature Climate Change, 2022, 12(10): 950−955. doi: 10.1038/s41558-022-01465-8
[3] Valpine D, Perry H. Plant responses to experimental warming in a montane meadow[J]. Ecology, 2001, 82(3): 637−648. doi: 10.1890/0012-9658(2001)082[0637:PRTEWI]2.0.CO;2
[4] Penuelas J, Filella I. Responses to a warming world[J]. Science, 2001, 294: 793−795. doi: 10.1126/science.1066860
[5] Moser A, Uhl E, Thomas R, et al. Effects of climate and the urban heat island effect on urban tree growth in Houston[J]. Open Journal of Forestry, 2017, 7(4): 428−445. doi: 10.4236/ojf.2017.74026
[6] Smith I A, Dearborn V K, Hutyra L R. Live fast, die young: accelerated growth, mortality, and turnover in street trees[J]. PLoS One, 2019, 14(5): e0215846. doi: 10.1371/journal.pone.0215846
[7] Han H, Xi B, Wang Y, et al. Lack of phenotypic plasticity in leaf hydraulics for 10 woody species common to urban forests of North China[J]. Tree Physiology, 2022, 42(6): 1203−1215. doi: 10.1093/treephys/tpac003
[8] Esperon-Rodriguez M, Ordoñez C, van Doorn N S, et al. Using climate analogues and vulnerability metrics to inform urban tree species selection in a changing climate: the case for Canadian cities[J]. Landscape and Urban Planning, 2022, 228: 104578. doi: 10.1016/j.landurbplan.2022.104578
[9] Brodribb T J, Jordan G J. Internal coordination between hydraulics and stomatal control in leaves[J]. Plant, Cell & Environment, 2008, 31(11): 1557−1564.
[10] Sack L, Scoffoni C. Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future[J]. New Phytologist, 2013, 198(4): 983−1000. doi: 10.1111/nph.12253
[11] 狄丽俊. 水稻不同叶位叶片气孔导度对CO2浓度和温度升高的响应[D]. 南京: 南京农业大学, 2018. Di L J. Response of stomatal conductance of rice leaves at different leaf positions to elevated carbon dioxide concentration and temperature[D]. Nanjing: Nanjing Agricultural University, 2018.
[12] Zhu J, Zhu H, Cao Y, et al. Effect of simulated warming on leaf functional traits of urban greening plants[J]. BMC Plant Biology, 2020, 20(1): 139. doi: 10.1186/s12870-020-02359-7
[13] 陈丽华, 宋丽华, 谢云. “灵武长枣”生长和叶片解剖结构对大气增温的响应[J]. 北方园艺, 2020(20): 1−8. doi: 10.11937/bfyy.20194812 Chen L H, Song L H, Xie Y. Responses of elevated temperature on growth and leaf anatomy of jujube cultivar ‘Lingwu Long Jujube’[J]. Northern Horticulture, 2020(20): 1−8. doi: 10.11937/bfyy.20194812
[14] 朱济友, 于强, 徐程扬, 等. 植物功能性状及其叶经济谱对硬化地表的响应[J]. 农业机械学报, 2019, 50(3): 204−211. Zhu J Y, Yu Q, Xu C Y, et al. Response of plant functional traits and its leaf economics spectrum to urban pavement[J]. Transactions of the Chinese Society for Agricultural Machinery, 2019, 50(3): 204−211.
[15] Funk J L, Cornwell W K. Leaf traits within communities: context may affect the mapping of traits to function[J]. Ecology, 2013, 94(9): 1893−1897. doi: 10.1890/12-1602.1
[16] Lenoir J, Bertrand R, Comte L, et al. Species better track climate warming in the oceans than on land[J]. Nature Ecology & Evolution, 2020, 4(8): 1−16.
[17] 岳阳, 韦柳端, 徐程扬, 等. 不同叶片质地行道树枝叶功能性状对冠下硬化地表覆盖的响应[J]. 北京林业大学学报, 2022, 41(23): 9492−9501. doi: 10.12171/j.1000-1522.20210262 Yue Y, Wei L D, Xu C Y, et al. Response of functional characters of street trees with different leaf textures to impervious land coverage under canopy[J]. Journal of Beijing Forestry University, 2022, 41(23): 9492−9501. doi: 10.12171/j.1000-1522.20210262
[18] 潘莹萍, 陈亚鹏. 叶片水力性状研究进展[J]. 生态学杂志, 2014, 33(10): 2834−2841. Pan Y P, Chen Y P. Recent advances in leaf hydraulic traits[J]. Chinese Journal of Ecology, 2014, 33(10): 2834−2841.
[19] 刘宏伟. 两种不同生态系统中植物叶片功能性状及养分再吸收比较研究[D]. 重庆: 西南大学, 2014. Liu H W. The comparative study on leaf functional traits and nutrient resorption in two different ecosystems of Chongqing[D]. Chongqing: Southwest University, 2014.
[20] 刘施含, 曹银贵, 贾颜卉, 等. 北京市海淀区热岛特征及形成原因分析[J]. 资源与产业, 2020, 22(5): 86−95. Liu S H, Cao Y G, Jia Y H, et al. Analysis on the characteristics and causes of heat island in Haidian District of Beijing[J]. Resources and Industry, 2020, 22(5): 86−95.
[21] Buckley T N. How do stomata respond to water status?[J]. New Phytologist, 2019, 224(1): 21−36. doi: 10.1111/nph.15899
[22] Selvi F, Massimo B. Leaf surface and anatomy in Boraginaceae tribe Boragineae with respect to ecology and taxonomy[J]. Flora, 2001, 196(4): 269−285. doi: 10.1016/S0367-2530(17)30056-7
[23] He N, Liu C, Tian M, et al. Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functions[J]. Functional Ecology, 2018, 32(1): 10−19. doi: 10.1111/1365-2435.12934
[24] 李和平. 植物显微技术[M]. 北京: 科学出版社, 2009. Li H P. Plant microscopy technology[M]. Beijing: Science Press, 2009.
[25] Zhang S B, Guan Z J, Sun M, et al. Evolutionary association of stomatal traits with leaf vein density in Paphiopedilum, Orchidaceae[J]. PLoS One, 2012, 7(6): e40080. doi: 10.1371/journal.pone.0040080
[26] 徐程扬. 紫椴幼苗、幼树对光的响应与适应研究[D]. 北京: 北京林业大学, 2005. Xu C Y. Responses and adaptations of Tilia Amurensis seedlings and saplings to light rgimes[D]. Beijing: Beijing Forestry University, 2005.
[27] 李乐, 曾辉, 郭大立. 叶脉网络功能性状及其生态学意义[J]. 植物生态学报, 2013, 37(7): 691−698. doi: 10.3724/SP.J.1258.2013.00072 Li L, Zeng H, Guo D L. Leaf venation functional traits and their ecological significance[J]. Chinese Journal of Plant Ecology, 2013, 37(7): 691−698. doi: 10.3724/SP.J.1258.2013.00072
[28] Givnish T J. Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constraints[J]. New Phytologist, 1987, 106(S1): 131−160. doi: 10.1111/j.1469-8137.1987.tb04687.x
[29] Zhang Y J, Cao K F, Lawren S, et al. Extending the generality of leaf economic design principles in the cycads, an ancient lineage[J]. New phytologist, 2015, 206(2): 817−829. doi: 10.1111/nph.13274
[30] 王颖, 许艺, 陈秉初, 等. 探究环境因素对植物气孔开闭的影响[J]. 生物学通报, 2019, 54(1): 39−42. Wang Y, Xu Y, Chen B C, et al. The effect of environmental factors on stomatal opening and closing of plants[J]. Bulletin of Biology, 2019, 54(1): 39−42.
[31] 白万鹏, 李虎军, 刘林波, 等. 植物气孔与角质层蜡质响应非生物胁迫的研究进展[J]. 安徽农业科学, 2020, 48(22): 14−18, 46. Bai W P, Li H J, Liu L B, et al. Research progress of plant stomata and cuticular wax in response to abiotic stress[J]. Journal of Anhui Agricultural Sciences, 2020, 48(22): 14−18, 46.
[32] Hasanuzzaman M, Davies N W, Shabala L, et al. Residual transpiration as a component of salinity stress tolerance mechanism: a case study for barley[J]. BMC Plant Biology, 2017, 17(1): 107. doi: 10.1186/s12870-017-1054-y
[33] Premachandra G S, Hirofumi S, Kounosuke F, et al. Leaf water relations, osmotic adjustment, cell membrane stability, epicuticular wax load and growth as affected by increasing water deficits in sorghum[J]. Journal of Experimental Botany, 1992, 43(12): 1569−1576. doi: 10.1093/jxb/43.12.1569
[34] 朱广龙, 魏学智. 酸枣叶片结构可塑性对自然梯度干旱生境的适应特征[J]. 生态学报, 2016, 36(19): 6178−6187. Zhu G L, Wei X Z. Leaf morphological plasticity of Ziziphus jujuba var. spinosa in response to natural drought gradient ecotopes[J]. Acta Ecologica Sinica, 2016, 36(19): 6178−6187.
[35] 朱玉, 黄磊, 郑云普, 等. 高温对高丛越橘叶片气孔特征和气体交换参数的影响[J]. 果树学报, 2016, 33(4): 444−456. Zhu Y, Huang L, Zheng Y P, et al. Effects of high temperatures on leaf stomatal traits and gas exchanges of highbush blueberries[J]. Journal of Fruit Science, 2016, 33(4): 444−456.
[36] 郑云普, 徐明, 王建书, 等. 玉米叶片气孔特征及气体交换过程对气候变暖的响应[J]. 作物学报, 2015, 41(4): 601−612. doi: 10.3724/SP.J.1006.2015.00601 Zheng Y P, Xu M, Wang J S, et al. Responses of the stomatal traits and gas exchange of maize leaves to climate warming[J]. Acta Agronomica Sinica, 2015, 41(4): 601−612. doi: 10.3724/SP.J.1006.2015.00601
[37] 闫润杰, 李菲, 张茜茜, 等. 秸秆还田下不同水肥对玉米气孔形态特征及其分布格局的影响[J]. 节水灌溉, 2021(10): 71−77. Yan R J, Li F, Zhang X X, et al. Effects of irrigation and fertilization on the morphological traits and distribution pattern of stomata of maize leaves under straw returning[J]. Water Saving Irrigation, 2021(10): 71−77.
[38] Hetherington A M, Woodward F I. The role of stomata in sensing and driving environmental change[J]. Nature, 2003, 42: 901−908.
[39] Zhang S B, Sun M, Cao K F, et al. Leaf photosynthetic rate of tropical ferns is evolutionarily linked to water transport capacity[J]. PLoS One, 2014, 9(1): e84682. doi: 10.1371/journal.pone.0084682
[40] 司怀通. 红树林植物气孔对环境因子的响应特点[D]. 南宁: 广西大学, 2017. Si H T. The characteristics of stomatal response to environmental factors in mangrove forest[D]. Nanning: Guangxi University, 2017.
[41] 张亚, 杨石建, 孙梅, 等. 基部被子植物气孔性状与叶脉密度的关联进化[J]. 植物科学学报, 2014, 32(4): 320−328. Zhang Y, Yang S J, Sun M, et al. Stomatal traits are evolutionarily associated with vein density in basal angiosperms[J]. Plant Science Journal, 2014, 32(4): 320−328.
[42] 李林鑫, 陶长铸, 林景泉, 等. 不同杉木无性系叶片解剖结构对大气增温的响应[J]. 生态学报, 2022, 42(20): 8385−8397. Li L X, Tao C Z, Lin J Q, et al. Response of needle anatomical structure of different Chinese fir clones to atmospheric warming[J]. Acta Ecologica Sinica, 2022, 42(20): 8385−8397.
[43] 王姝, 周道玮. 植物表型可塑性研究进展[J]. 生态学报, 2017, 37(24): 8161−8169. Wang S, Zhou D W. Research on phenotypic plasticity in plants: an overview of history, current status, and development trends[J]. Acta Ecologica Sinica, 2017, 37(24): 8161−8169.
[44] Qin M, Hao L, Sun L, et al. Climatic controls on watershed reference evapotranspiration varied during 1961−2012 in southern China[J]. Journal of the American Water Resources Association, 2019, 55(1): 189−208. doi: 10.1111/1752-1688.12714
[45] Almeida A C, Landsberg J J. Evaluating methods of estimating global radiation and vapor pressure deficit using a dense network of automatic weather stations in coastal Brazil[J]. Agricultural & Forest Meteorology, 2003, 118(34): 237−250.
[46] Williams L E, Baeza P. Relationships among ambient temperature and vapor pressure deficit and leaf and stem water potentials of fully irrigated, field-grown grapevines[J]. American Journal of Enology & Viticulture, 2007, 58(2): 173−181.
[47] 黄志刚, 李锋瑞, 曹云, 等. 南方红壤丘陵区杜仲人工林土壤水分动态[J]. 应用生态学报, 2007, 18(9): 1937−1944. Huang Z G, Li F R, Cao Y, et al. Dynamics of soil water under Eucommia ulmoides plantation in hilly red soil region of southern China[J]. Chinese Journal of Applied Ecology, 2007, 18(9): 1937−1944.
[48] Wright I J, Reich P B, Cornelissen J H C, et al. Modulation of leaf economic traits and trait relationships by climate[J]. Global Ecology Biogeography, 2010, 15(5): 411−421.
[49] 陈展宇. 旱稻抗旱解剖结构及其生理特性的研究[D]. 长春: 吉林农业大学, 2008. Chen Z Y. Research on anatomic structure of drought resistance and physiological characteristic of upland rice[D]. Changchun: Jilin Agricultural University, 2008.














计量
- 文章访问数: 264
- HTML全文浏览量: 49
- PDF下载量: 21
