
Screening of water-saving and sink-enhancing plant species in arid and semi-arid regions of Northwest China
-
摘要:目的
为改善西北地区的生态环境,践行可持续发展理念,促进地区生态修复与节水增汇,有必要开展西北干旱半干旱地区生存的灌草植物适宜性研究。
方法以现场勘察和文献调研当地优势植物为基础,选择10种草本植物和7种灌木为研究对象,进行田间试验,测定它们的生长性状指标、光合生理指标、生物量和碳储量指标,运用层次分析法分别对草本植物和灌木进行节水增汇的综合评价,以此筛选适生植物种类。
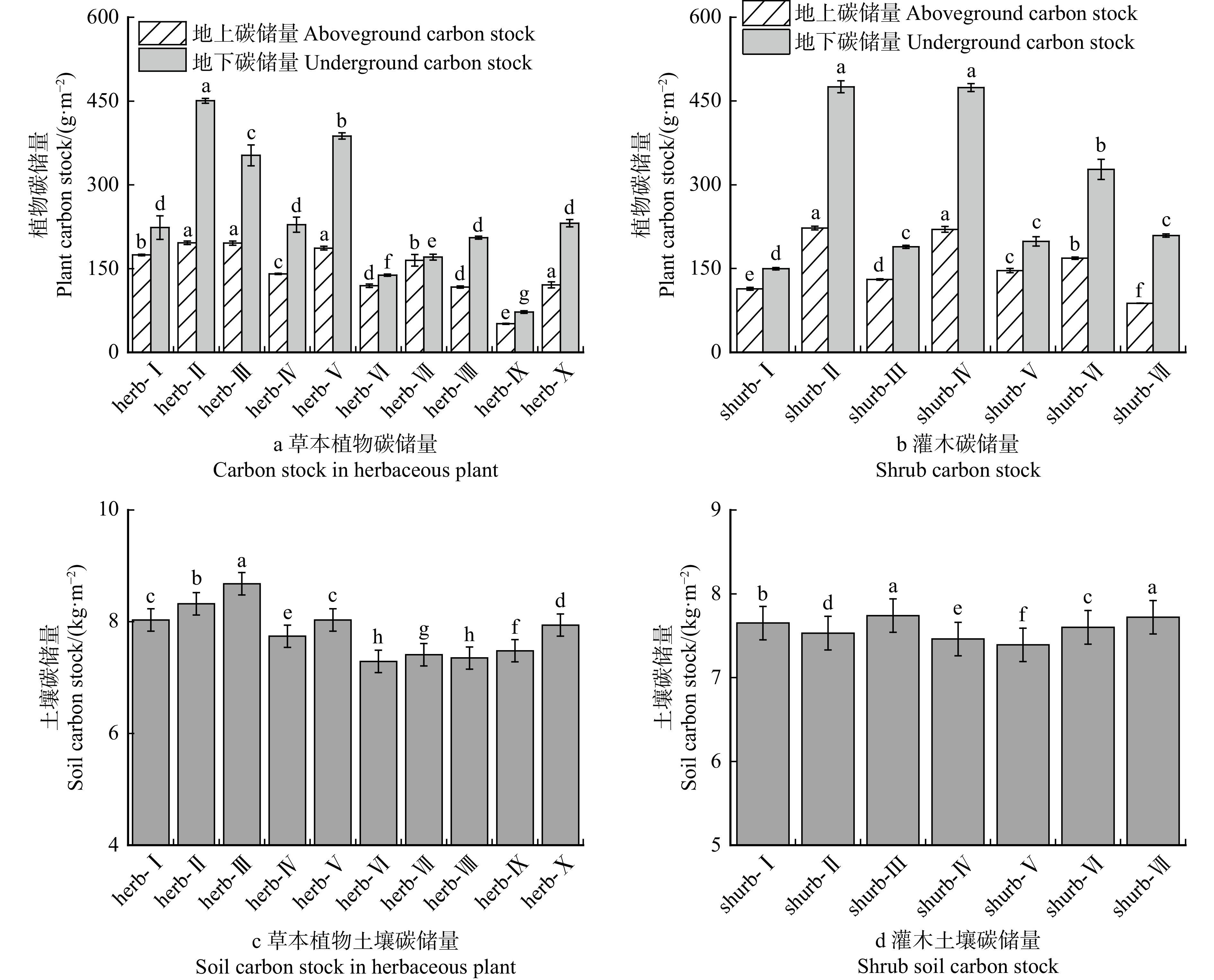
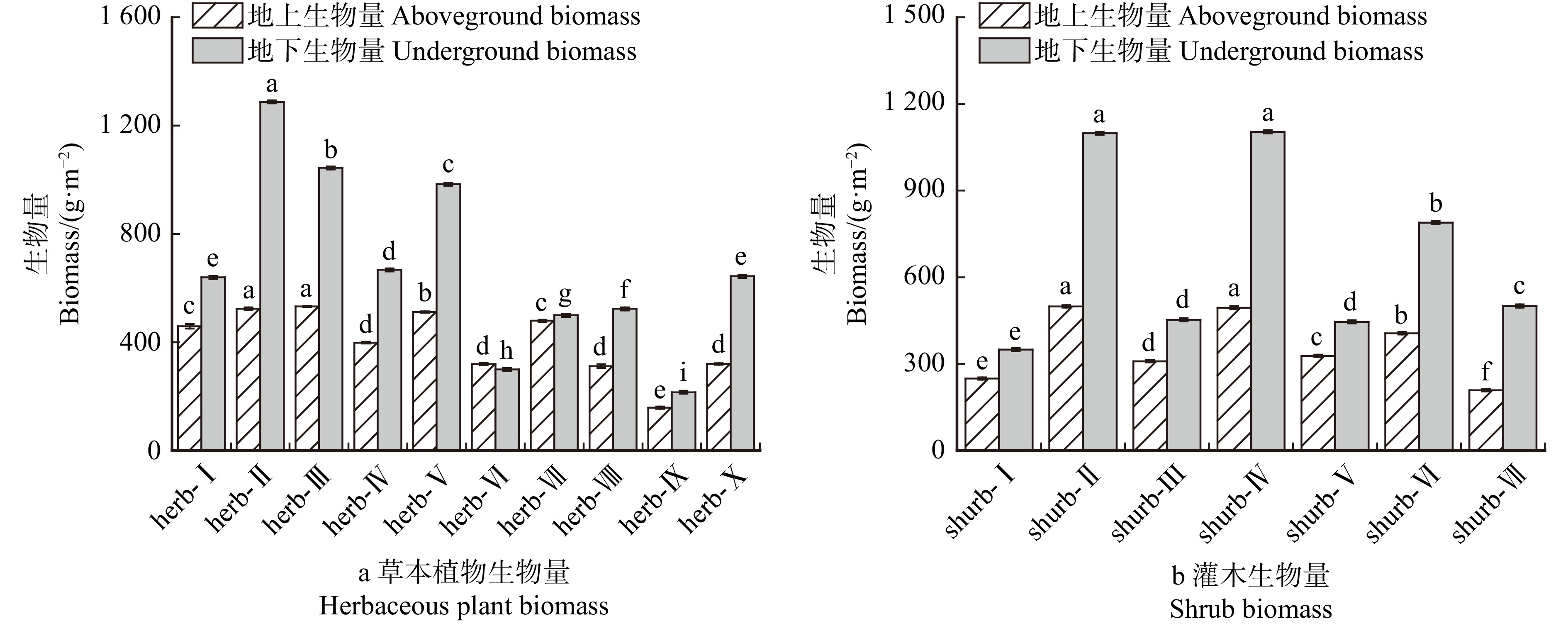
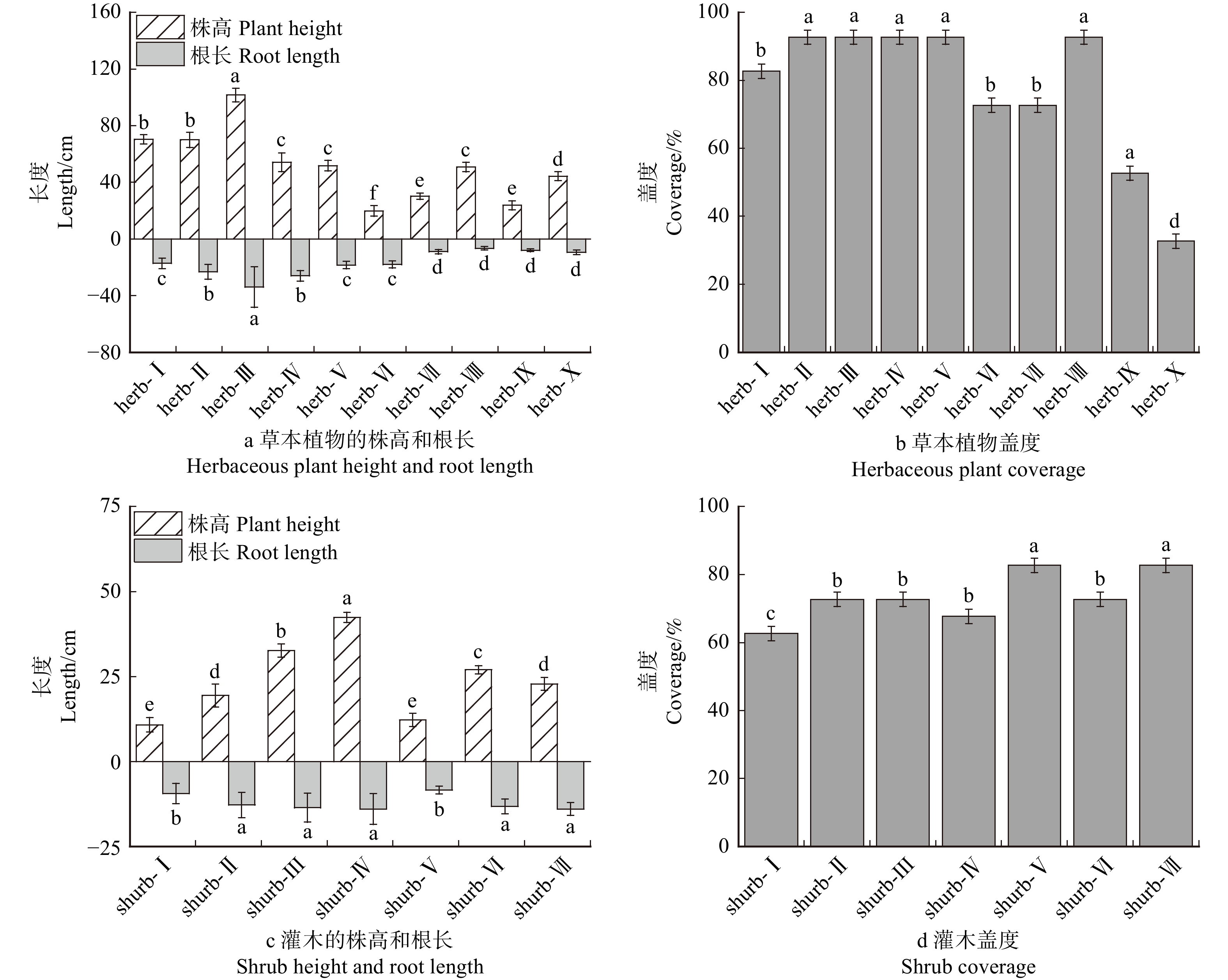
结果(1)草本植物中,高冰草、沙打旺和早熟禾的水分利用效率较高,灌木中沙地柏、柠条锦鸡儿和紫穗槐的水分利用效率较高;(2)草本植物中总碳储量较高的是紫花苜蓿、草木樨和沙打旺,灌木总碳储量较高的是紫穗槐、柠条锦鸡儿和沙棘;(3)节水增汇草本植物排序从高到低依次为紫花苜蓿、 草木樨、 沙打旺、 蜀葵 、 披碱草、 沙蒿、高冰草、高羊茅、黑麦草、早熟禾,节水增汇灌木排序从高到低依次为紫穗槐、柠条锦鸡儿、沙棘、沙地柏、胡枝子、沙柳、花棒。
结论本研究筛选出了适宜在西北干旱半干旱地区生存的节水增汇植物种类,为该地区植物配置提供参考,为推动生态环境改善、地区生态重建和碳汇增加提供科技支撑。
Abstract:ObjectiveThe study of shrub and grass plants suitable for survival in the arid and semi-arid regions of Northwest China can not only improve the ecological environment of Northwest China, but also conform to the concept of sustainable development, which is of far-reaching significance for the ecological restoration and the water-saving and carbon-sink-enhancing work in the region.
MethodBased on the field survey and literature research of local dominant plants, this study selected ten herbaceous plants and seven shrubs as research objects. Field experiments were conducted, then their growth trait indexes, photosynthesis physiological indexes, biomass and carbon storage indexes were tested. The hierarchical analysis method was applied to comprehensively evaluate the water-saving and carbon-sink-enhancing performance of herbaceous plants and shrubs, respectively, so as to screen out the suitable plant species.
Result(1) In herbaceous plants, water-use efficiencies of Agropyron elongatum, Astragalus laxmannii and Poa annua were high. In shrubs, water-use efficiencies of Juniperus sabina, Caragana korshinskii, and Amorpha fruticosa were high. (2) In herbaceous plants, total carbon stocks of Medicago sativa, Melilotus officinalis, and Astragalus laxmannii were high. In shrubs, total carbon stocks of Amorpha fruticosa, Caragana korshinskii, and Hippophae rhamnoides were high. (3) Water-saving and carbon-sink-enhancing herbs ranked in descending order of Medicago sativa > Melilotus officinalis > Astragalus laxmannii > Alcea rosea > Elymus dahuricus > Artemisia desertorum > Agropyron elongatum > Festuca elata > Lolium perenne > Poa annua. Water-saving and arbon-sink-enhancing shrubs ranked in descending order of Amorpha fruticosa > Caragana korshinskii > Hippophae rhamnoides > Juniperus sabina > Lespedeza bicolor > Salix cheilophila > Corethrodendron scoparium.
ConclusionThe study has screened out water-saving and carbon-sink-enhancing plant species suitable for survival in the arid and semi-arid regions of Northwest China. It can not only provide a basis for the plant configuration in the region, and improve ecological environment, but also provide scientific and technological support for ecological reconstruction and increase of carbon sinks in the region.
-
湿地因其高生产力和低温厌氧环境而使土壤积累大量有机碳,故被认为是重要的碳汇,对全球碳循环具有积极影响[1−2]。湿地仅占地球陆地表面的5% ~ 8%,却储存了全球陆地碳的20% ~ 30%,因此在减缓气候变化方面的作用备受关注[2−3]。然而,由于湿地碳储量存在较大的空间异质性[4−6],其在局地、区域和全球尺度上均存在较大不确定性[7−8]。因此,揭示湿地碳储量的空间分异规律及其形成机制将有助于减少这些不确定性。
湿地碳储量依赖于植被碳输入和土壤碳分解两者的长期平衡,且受气候、水文、土壤、植被和微地形等多重因子的显著影响[9−12]。气候是决定湿地土壤碳库的关键因素,其中温度和降水直接或间接控制土壤有机碳分解速率[13]。同时,厌氧条件和较高植被生产力是促进土壤有机碳积累的关键因素[14−15]。水文状况作为湿地生态系统的主要驱动力,直接影响湿地的理化特性,特别是氧的可利用性,进而调控土壤有机碳分解速率[16−18]。此外,水文过程还直接影响植物群落的结构和分布[19−21],以及沿水分环境梯度分布的各类湿地植物的生长、生物量积累和碳固定[22]。植被是湿地碳储量和固碳的主要影响因素[23−24],群落优势种的变化能够改变土壤中有机物的质和量,进而影响湿地土壤碳库[23−25]。微地形通过影响湿地的小气候、水文、土壤理化特性,调控湿地碳累积过程,对湿地土壤有机碳空间分布起着至关重要作用[26−27]。湿地生态系统还存在着密切的碳氮耦合关系[28],通常湿地生产力受到氮的限制[29]。氮的可利用性的增加能够提高湿地生产力和生物量积累,进而增加湿地的碳固存[30]。此外,氮含量及其迁移转化过程显著影响湿地生态系统的结构和功能[29]。因此,了解湿地氮储量的空间分布将有助于认识其固碳过程[31−33]。研究湿地碳氮储量沿水分环境梯度的空间变化,有助于揭示其空间异质性的形成机制,为湿地生态系统的管理和保护提供科学依据。
从景观尺度上看,北方温带地区的湖泊湿地储存了大量的碳[34],其中80%的碳储存在湖岸泥炭湿地和沉积物中。这些泥炭湿地和沉积物仅占整个景观面积约1/3(泥炭湿地和沉积物各占20%和13%),却储存碳库总量的80%以上[35]。因此,量化湖泊湿地碳储量具有重要意义。湖岸湿地(湖滨带)是湖泊水域与陆地之间的过渡区域[36],是湖泊湿地重要组成部分[37]。水文状况是湖滨带建立和维持的控制性因素。其中水位波动塑造了湖滨带时空范围和功能差异[38],尤其是决定了植被沿水陆过渡性分布(湿生向中生和旱生过渡),引起局地碳氮储量沿水分环境梯度产生空间分异[32,39]。例如:我国北方温带干旱区高原湖泊湿地土壤有机碳储量沿水分环境梯度呈降低趋势,主要受水文过程和植被类型所影响[40−41];而亚热带湿润区平原湖泊沿岸湿地土壤碳储量随水分梯度呈增加趋势,主要受人为施氮所影响[42]。目前国内有关湖滨带碳氮储量沿水分环境梯度空间分异方面的研究比较缺乏,已有的研究结果在不同气候区存在较大差异性,且多集中在土壤层和碳储量方面。因此,有必要从生态系统尺度上揭示各气候区湖滨带碳氮储量沿水分环境梯度空间分异规律及其形成机制。
长白山园池沿岸湿地具有湖滨带和高海拔区的气候敏感性特征[38],了解这些地区的生态系统碳氮储量特征对全球气候变化的潜在反馈具有重要意义。本研究以长白山园池沿岸沿水分环境梯度分布的典型天然沼泽类型为对象,通过测定各沼泽类型的植被、土壤和凋落物的碳氮储量以及相关环境因子(温度、水位和土壤理化性质等),量化各沼泽类型的生态系统碳氮储量及其分配格局,揭示其沿过渡带水分环境梯度的空间分异规律及形成机制,以便为温带高海拔区湖泊湿地碳汇管理提供依据。
1. 研究区概况与研究方法
1.1 研究区概况
研究地点位于长白山园池国家级湿地自然保护区(42°01′54″N,128°26′02″E),海拔1 250 m。该区受长白山地区特殊的大陆性山地气候的影响,冬季漫长寒冷,夏季短暂暖湿,年平均气温在−7 ~ 3 ℃之间,年平均降水量700 ~ 1 400 mm。本研究调查的园池湖泊湿地是形成时间较短的雨养型泥炭沼泽湿地,位于长白山火山东坡,是火山喷发后所形成的火山口湖演变而来,距今已有900多年[43]。沿园池湖岸至高地水分环境梯度依次分布着芦苇沼泽、草丛沼泽、杜香沼泽、落叶松泥炭藓沼泽、落叶松藓类沼泽和落叶松苔草沼泽6种沼泽类型。研究区土壤类型主要为山地沼泽土、草甸土和棕色针叶林土,主要地带性植被为寒温带针叶林,其中长白落叶松(Larix olgensis)是主要的乔木树种,细叶杜香(Ledum palustre)是典型的灌木,芦苇(Phragmites australis)、修氏苔草(Carex schmidti)、泥炭藓(Sphagnum palustre)等草本植物构成了湿地的底层植被。试验地位于国家级自然保护区内部,海拔较高且无人为活动干扰,是研究生态系统碳、氮储量沿环境梯度空间分布格局及其形成机制的理想场所。
1.2 研究方法
1.2.1 样地设置
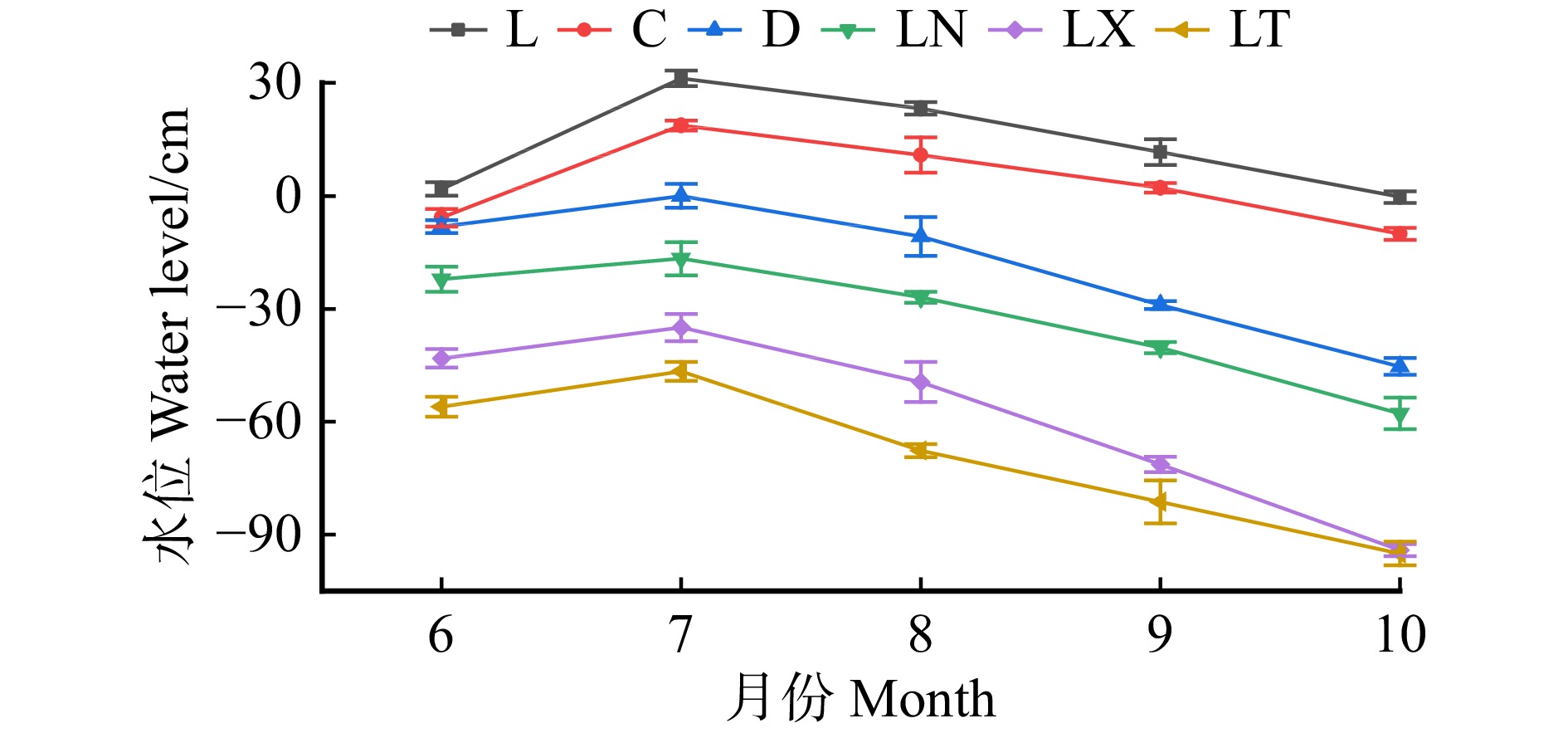
于2021年5月进行样地设置,结合研究区的微地形环境特点及不同植被类型的空间分布状况,沿园池湖岸至高地环境梯度设置3条重复样带,每个样带均包括6种沼泽群落类型(芦苇沼泽L、草丛沼泽C、杜香沼泽D、落叶松泥炭藓沼泽LN、落叶松藓类沼泽LX、落叶松苔草沼泽LT),共设置了18个20 m × 30 m标准地。于 2021年5月上旬(生长季初期)对标准样地进行了每木调查。于 2021年6月上旬(土壤化冻后)至10月上旬(土壤冻结前),每月对样地进行1次水位观测,并记录水位变化情况。森林湿地类型的林分状况见表1,各沼泽类型的土壤理化性质见表2,各沼泽类型的水位变化情况见图1。
表 1 长白山园池沿岸3种森林湿地类型林分特征Table 1. Stand characteristics of three forest wetland types along lakeshore in Yuanchi, Changbai Mountain湿地类型
Wetland type树种
Tree species胸高断面积/(m2·hm−2)
Basal area/(m2·ha−1)平均胸径
Mean DBH/cm密度/(株·hm−2)
Density/(plant·ha−1)胸径范围
DBH range/cmLN 落叶松 Larix olgensis 14.90 9.25 1 933 4 ~ 18 LX 落叶松 Larix olgensis 26.28 11.08 2 094 4 ~ 26 LT 落叶松 Larix olgensis 22.75 12.59 1 522 4 ~ 26 注:LN.落叶松泥炭藓沼泽;LX.落叶松藓类沼泽;LT.落叶松苔草沼泽。下同。Notes: LN, Larix olgensis-Sphagnum magellanicum swamp; LX, Larix olgensis-moss swamp; LT, Larix olgensis-Carex schmidtii swamp. The same below. 表 2 长白山园池沿岸6种湿地土壤理化性质Table 2. Soil physical and chemical properties of six wetland types along lakeshore in Yuanchi, Changbai Mountain湿地类型
Wetland type水位
Water level/cm水位波动幅度
Water level fluctuation
amplitude/cm土壤温度
Soil
temperature/℃有机质
Organic
matter/(g·kg−1)全氮
Total
nitrogen/(g·kg−1)全磷
Total
phosphorus/(g·kg−1)L 13.50 ± 1.94A 31.50± 1.00D 6.35± 0.25A 302.11 ± 22.13A 10.33 ± 0.49A 0.44 ± 0.01B C 3.13 ± 1.82B 28.83± 0.57D 6.10 ± 0.14A 263.94 ± 8.68B 10.00 ± 0.37A 0.49 ± 0.06B D −18.67 ± 1.53C 45.33 ± 4.37BC 4.24 ± 0.11B 228.34 ± 14.46C 5.24 ± 0.26B 0.61 ± 0.10A LN −32.80 ± 2.73D 42.63 ± 2.97C 3.94 ± 0.26C 208.65 ± 14.42C 4.93 ± 0.97B 0.41 ± 0.03B LX −58.67 ± 1.52E 60.17 ± 5.11A 3.92 ± 0.06C 47.78 ± 5.89D 1.00 ± 0.08C 0.18 ± 0.04C LT −69.33 ± 1.39F 48.33 ± 5.61B 3.96 ± 0.09BC 55.81 ± 4.65D 1.17 ± 0.14C 0.21 ± 0.02C 注:表中数据为平均值 ± 标准差;L.芦苇沼泽;C.草丛沼泽;D.杜香沼泽。不同大写字母表示不同湿地类型之间差异显著(P < 0.05)。下同。Notes: data in the table are mean ± SD. L, Phragmites australis swamp; C, tussock swamp; D, Ledum palustre swamp. Different capital letters mean significant difference among different wetland types (P < 0.05). The same below. ![]() 图 1 长白山园池沿岸6种湿地类型的水位变化情况Figure 1. Water level changes of six wetland types along lakeshore in Yuanchi, Changbai Mountain
图 1 长白山园池沿岸6种湿地类型的水位变化情况Figure 1. Water level changes of six wetland types along lakeshore in Yuanchi, Changbai Mountain1.2.2 植被碳储量测定
乔木层生物量测定:采用相对生长方程法,首先对各沼泽类型标准地中胸径 ≥ 4 cm的林木进行每木检尺,然后利用各样地胸径生长数据结合现有长白山区森林沼泽群落建群种落叶松相对生长方程[44],估算各样地乔木层生物量。灌木层和草本层生物量测定:采用收获法,在每个标准地的中心和四角设置5个2 m × 2 m样方测定灌木层生物量,并随机选择10个1 m × 1 m样方测定草本层生物量。在具体测定过程中,分别对地上枝叶和地下根系进行生物量鲜质量测定,并对枝叶和根系进行取样,在70 ℃下烘干至恒质量,得到灌木层和草本层的生物量干质量。于秋季落叶前(10月前),在各样地中分别设置5个50 cm × 50 cm的样方,收集其中全部凋落物装入封口袋,带回实验室,在70 ℃下烘干至恒质量,得到凋落物生物量的干质量。植被碳、氮储量的测定:利用碳/氮分析仪MultiN/C 3100和HT 1300 Soilds Module,通过1 300 ℃干烧法测定各层的有机碳含量,采用间断分析仪AQ400测定各层全氮含量。最后,将各组分的生物量分别乘以碳含量和氮含量,可以得到乔木层、灌木层、草本层和凋落物层的碳储量和氮储量,将4部分加和即可获得植被层的碳、氮储量。
1.2.3 土壤碳氮储量测定
在每个标准样地中心,以“品”字形设置3个土壤剖面,由于各沼泽类型土壤深度不同,取样深度均至母质层。确定土壤剖面后,用土壤环刀(100 cm3)每间隔10 cm作为一取样层,土样用铝盒封装,带回实验室在烘箱105 ℃下烘24 h后,测定其土壤密度;同时在同一土层深度取约500 g土样装入样品袋,带回实验室风干后,取出大于2 mm 的根系或岩石后,在70 ℃下烘干24 h,研磨粉碎后过2 mm土壤筛,利用同上方法测定土壤有机碳含量和全氮含量,并利用以下公式计算土壤有机碳储量。某一土层i的有机碳密度(SOCi,kg/m2)的计算公式[45]为
SOCi=CiDiEi(1−Gi)/100 式中:Ci为土壤有机碳含量(g/kg),Di为土壤密度(g/cm3),Ei为土层厚度(cm),Gi为直径大于2 mm的石砾所占的体积百分比(%),i为某一土层,t为所有土层的总和。土壤剖面由k层组成,那么该剖面的碳密度(SOCt,kg/m²)如下,并利用相同方法计算土壤全氮储量。
SOCt=k∑i=1SOCi=k∑i=1CiDiEi(1−Gi)/100 1.2.4 数据处理
数据使用 SPSS 22.0软件进行单因素方差分析(one-way ANOVA),采用最小显著差异法(least significant difference,LSD)分析不同数据组间的差异性,显著性水平设置为α = 0.05。利用相关关系与多元逐步分析对生态系统组分碳氮储量与环境因子的关系进行分析,并筛选主要因子,使用Origin 2023进行制图。
2. 结果和分析
2.1 园池沿岸沼泽湿地植被碳氮储量及其分配格局
园池湖岸至高地依次分布的6种沼泽类型的植被碳、氮储量存在显著差异(表3)。植被碳储量分布在2.17 ~ 69.98 t/hm2之间,其中,落叶松苔草沼泽和落叶松藓类沼泽(LT和LX)相对较高(显著高于其他4种沼泽0.8 ~ 31.2倍,P < 0.05);落叶松泥炭藓沼泽和杜香沼泽(LN和D)居于中等(显著高于其他2种沼泽1.4 ~ 16.7倍,且LN显著高于D 4.3倍);芦苇沼泽和草丛沼泽(L和C)相对较低(且C显著高于L 0.4倍)。植被氮储量分布在0.058 ~ 0.940 t/hm2,也是LT和LX相对较高(显著高于其他4种沼泽0.8 ~ 15.2倍); LN和D居中(显著高于其他2种沼泽0.6 ~ 7.0倍,且LN显著高于D 1.8倍);L和C相对较低(且C显著高于L 0.8倍)。因此,长白山高海拔区园池沿岸沼泽湿地的植被碳储量和氮储量沿湖岸至高地水分环境梯度均呈递增规律性(LT ≈ LX > LN > D > C > L),仅是碳储量递增幅度大于氮储量。
表 3 长白山园池沿湖岸至高地分布的6种湿地类型的植被碳氮储量Table 3. Vegetation carbon and nitrogen storage of six wetland types along the environmental gradient from lakeshore to upland in Yuanchi, Changbai Mountain指标
Index层次
Layer湿地类型 Wetland type L C D LN LX LT 生物量/(t·hm−2)
Biomass/(t·ha−1)乔木层 Tree layer 71.49 ± 28.71B 135.28 ± 14.98A 141.72 ± 13.01A 灌木层 Shrub layer 11.41 ± 2.00A 4.58 ± 1.61B 3.96 ± 1.17B 4.08 ± 1.46B 草本层 Herb layer 4.14 ± 0.35A 5.57 ± 0.87A 2.35 ± 0.44B 3.46 ± 0.97B 3.40 ± 0.57B 3.28 ± 0.50B 凋落物层 Litter layrt 1.09 ± 0.27C 1.62 ± 0.54B 1.71 ± 0.53B 2.89 ± 0.60A 2.61 ± 0.59A 2.98 ± 0.57A 植被 Vegetation 5.24 ± 0.39E 7.18 ± 0.08D 15.47 ± 1.42C 82.42 ± 12.53B 145.24 ± 14.04A 152.05 ± 13.67A 碳含量 Carbon
content/(g·kg−1)乔木层 Tree layer 462.37 ± 4.16B 472.64 ± 7.78AB 479.51 ± 0.81A 灌木层 Shrub layer 476.97 ± 5.59A 477.18 ± 6.10A 466.81 ± 6.65AB 463.56 ± 7.04B 草本层 Herb layer 421.34 ± 3.00C 422.80 ± 5.84C 465.55 ± 15.65A 455.63 ± 6.48AB 457.46 ± 13.17A 445.52 ± 8.19B 凋落物层 Litter layer 377.07 ± 6.32B 403.13 ± 8.41B 461.55 ± 11.49A 448.95 ± 16.30A 461.56 ± 34.09A 411.41 ± 37.01B 植被 Vegetation 399.20 ± 1.83D 412.97 ± 6.92C 468.02 ± 0.19A 461.03 ± 3.31AB 464.62 ± 7.73A 450.00 ± 8.52B 碳储量/(t·hm−2)
Carbon storage/
(t·ha−1)乔木层 Tree layer 32.92 ± 7.73B 62.22 ± 6.75A 65.35 ± 5.95A 灌木层 Shrubd layer 5.40 ± 0.87A 2.35 ± 1.01B 1.86 ± 0.55B 1.92 ± 0.70B 草本层 Herb layer 1.75 ± 0.14B 2.36 ± 0.38A 1.11 ± 0.16C 1.74 ± 0.84B 1.56 ± 0.28BC 1.48 ± 0.24BC 凋落物层 Litter layer 0.41 ± 0.06C 0.64 ± 0.19B 0.79 ± 0.22B 1.33 ± 0.09A 1.20 ± 0.31A 1.23 ± 0.30A 植被 Vegetation 2.17 ± 0.16E 3.00 ± 0.04D 7.30 ± 0.62C 38.34 ± 5.92B 66.85 ± 2.37A 69.98 ± 7.63A 碳储量分配比
Carbon storage
allocation/%乔木层 Tree layer 85.10 ± 7.61 93.00 ± 1.27 93.37 ± 0.20 灌木层 Shrub layer 73.72 ± 5.49 6.53 ± 3.91 2.83 ± 0.65 2.74 ± 0.27 草本层 Herb layer 81.03 ± 1.08 78.51 ± 4.44 15.33 ± 3.42 4.85 ± 3.18 2.37 ± 0.60 2.10 ± 0.20 凋落物层 Litter layer 18.97 ± 1.08 21.49 ± 4.44 10.95 ± 2.09 3.52 ± 0.59 1.80 ± 0.13 1.78 ± 0.45 氮含量
Nitrogen content/
(g·kg−1)乔木层 Tree layer 11.12 ± 0.47A 12.07 ± 0.26A 11.95 ± 0.76A 灌木层 Shrub layer 12.63 ± 0.35B 14.33 ± 1.69B 17.60 ± 2.28A 15.55 ± 0.58AB 草本层 Herb layer 9.71 ± 0.38BC 13.62 ± 0.09A 7.83 ± 0.28C 11.14 ± 2.55B 15.03 ± 1.37A 14.44 ± 0.46A 凋落物层 Litter layer 12.62 ± 0.89B 15.78 ± 1.29A 17.52 ± 5.15A 10.84 ± 1.12B 10.73 ± 1.62B 13.72 ± 1.87AB 植被 Vegetation 11.16 ± 0.26B 14.70 ± 0.67A 12.66 ± 1.57B 11.86 ± 0.83B 13.86 ± 1.23AB 13.91 ± 0.46AB 氮储量/(t·hm−2)
Nitrogen storage/
(t·ha−1)乔木层 Tree layer 0.340 ± 0.093B 0.735 ± 0.09A 0.802 ± 0.10A 灌木层 Shrub layer 0.121 ± 0.02A 0.052 ± 0.02B 0.053 ± 0.02B 0.050 ± 0.02B 草本层 Herb layer 0.044 ± 0.01B 0.081 ± 0.01A 0.018 ± 0.00C 0.044 ± 0.02B 0.049 ± 0.01B 0.047 ± 0.01B 凋落物层 Litter layer 0.014 ± 0.01C 0.025 ± 0.01B 0.030 ± 0.01B 0.032 ± 0.00B 0.028 ± 0.01B 0.041 ± 0.01A 植被 Vegetation 0.058 ± 0.01E 0.107 ± 0.00D 0.169 ± 0.01C 0.467 ± 0.06B 0.865 ± 0.08A 0.940 ± 0.11A 氮储量分配比
Nitrogen storage
allocation/%乔木层 Tree layer 71.75 ± 12.28 84.80 ± 2.51 85.24 ± 1.34 灌木层 Shrub layer 71.57 ± 4.72 11.43 ± 5.30 6.19 ± 1.44 5.31 ± 0.28 草本层 Herb layer 76.01 ± 0.86 76.26 ± 6.20 10.59 ± 2.00 9.95 ± 5.93 5.73 ± 1.40 5.05 ± 0.74 凋落物层 Litter layer 23.99 ± 0.86 23.74 ± 6.20 17.84 ± 2.78 6.88 ± 1.06 3.27 ± 0.28 4.40 ± 1.10 各沼泽类型植被碳、氮储量在垂直空间上的分配规律也不同,芦苇沼泽和草丛沼泽的碳、氮储量以草本层占优势地位(78.51% ~ 81.03%和76.01% ~ 76.26%),而凋落物层占次要地位(18.97% ~ 21.49%和23.74% ~ 23.99%);杜香沼泽的碳、氮储量以灌木层占优势地位(73.72%和71.57%),草本层和凋落物层占比较低(26.28%和28.43%);而3种森林沼泽(LN、LX和LT)碳、氮储量均以乔木层占优势(85.10% ~ 93.37%和71.75% ~ 85.24%),灌木层、草本层和凋落物层占比较低(6.63% ~ 14.90%和14.76% ~ 28.25%)。因此,园池沿岸沼泽湿地的植被碳、氮储量存在明显的垂直空间分异规律性(即沿水分环境梯度依次分布的草类沼泽(L和C)、灌丛沼泽(D)和森林沼泽(LN、LX和LT)的碳、氮储量均以优势植被层占主体地位)。
2.2 园池沿岸沼泽湿地土壤碳氮储量及其分配格局
园池沿湖岸至高地水分环境梯度依次分布的6种沼泽类型的土壤碳、氮储量存在显著差异(表4)。土壤碳储总量分布在67.45 ~ 243.21 t/hm2之间,其中,芦苇沼泽(L)和草丛沼泽(C)相对较高(显著高于其他4种沼泽0.5 ~ 2.6倍,且L显著高于C 0.4倍,P < 0.05);杜香沼泽和落叶松泥炭藓沼泽居于中等(两者相近且显著高于其他2种沼泽0.5 ~ 0.7倍);而落叶松藓类沼泽和落叶松苔草沼泽相对较低且相近。其土壤氮储量分布在2.26 ~ 13.53 t/hm2,也是L和C相对较高(显著高于其他4种沼泽2.0 ~ 4.5倍,且L显著高于C 0.2倍);D和LN居中等(两者相近且显著高于其他2种沼泽0.5 ~ 0.8倍);而LX和LT相对较低且相近。因此,园池沿岸沼泽湿地的土壤碳、氮储量沿湖岸至高地水分环境梯度总体上均呈阶梯式递减规律性(L > C > D ≈ LN > LX ≈ LT),且氮储量降低幅度要大于碳储量。
表 4 长白山园池沿湖岸至高地分布的6种湿地类型的土壤碳氮储量Table 4. Soil carbon and nitrogen storage of six wetland types along the environmental gradient from lakeshore to upland in Yuanchi, Changbai Mountain指标
Index土层深度
Soil depth/cm湿地类型 Wetland type L C D LN LX LT 土壤密度
Bulk density/
(g·cm−3)0 ~ 10 0.14 ± 0.01Bd 0.14 ± 0.01Bd 0.10 ± 0.01Cd 0.14 ± 0.02Bc 0.74 ± 0.07Aa 0.66 ± 0.08Aa 10 ~ 20 0.14 ± 0.01Dde 0.15 ± 0.01Dd 0.33 ± 0.03Cc 0.40 ± 0.06Bb 0.75 ± 0.08Aa 0.78 ± 0.06Aa 20 ~ 30 0.14 ± 0.04De 0.32 ± 0.05Cc 0.66 ± 0.06Ba 0.60 ± 0.03Ba 0.72 ± 0.03ABa 0.78 ± 0.07Aa 30 ~ 40 0.49 ± 0.04Bc 0.55 ± 0.10ABb 0.52 ± 0.04Bb 0.47 ± 0.12Bab 0.69 ± 0.05Aa 0.67 ± 0.11Aa 40 ~ 50 0.91 ± 0.04Aa 0.78 ± 0.03Ba 50 ~ 60 0.58 ± 0.03Ab 0.60 ± 0.03Ab 60 ~ 70 0.49 ± 0.02c 平均值
Mean0.41 ± 0.01B 0.42 ± 0.01B 0.40 ± 0.01B 0.40 ± 0.01B 0.72 ± 0.01A 0.72 ± 0.01A 碳含量
Carbon content/
(g·kg−1)0 ~ 10 344.50 ± 47.70ABa 350.70 ± 11.42Ab 319.80 ± 14.35ABa 310.15 ± 23.15Ba 60.86 ± 6.39Ca 78.09 ± 7.42Ca 10 ~ 20 346.38 ± 21.23Ba 386.70 ± 4.94Aa 158.94 ± 20.96Cb 143.29 ± 23.10Cb 25.24 ± 4.15Db 23.03 ± 4.37Db 20 ~ 30 349.60 ± 11.96Aa 125.43 ± 19.16Bc 41.73 ± 9.89Cc 22.15 ± 2.24Cc 14.34 ± 4.80Cc 18.42 ± 2.21Cb 30 ~ 40 112.59 ± 13.90Ab 25.47 ± 4.33Bd 9.33 ± 1.61Cd 8.52 ± 1.83Cc 10.43 ± 1.69Cc 9.95 ± 1.36Cc 40 ~ 50 38.55 ± 12.56Ac 17.45 ± 5.46Bde 50 ~ 60 21.07 ± 2.68Ad 12.86 ± 1.48Be 60 ~ 70 13.97 ± 2.92e 平均值 175.24 ± 12.84A 153.10 ± 5.04A 132.45 ± 8.38B 121.03 ± 8.36C 27.72 ± 3.41D 32.37 ± 2.69D Mean 碳储量/(t·hm−2)
Carbon storage/
(t·ha−1)0 ~ 10 46.07 ± 3.09ABa 46.89 ± 0.37ABb 32.92 ± 3.67Cb 40.71 ± 4.55Bb 37.15 ± 2.54BCa 48.10 ± 2.85Aa 10 ~ 20 46.97 ± 7.27Ba 54.75 ± 2.62Aa 53.75 ± 7.95Aa 53.92 ± 1.03Aa 16.52 ± 5.22Cb 11.53 ± 0.93Cb 20 ~ 30 49.09 ± 2.95Aa 39.44 ± 6.36Ac 25.25 ± 8.05Bc 10.80 ± 1.46Cc 8.13 ± 2.37Dc 9.62 ± 1.01Cb 30 ~ 40 54.39 ± 7.67Aa 13.40 ± 1.57Bd 2.98 ± 0.80Dd 2.61 ± 0.35Dd 5.64 ± 1.69Cd 4.84 ± 1.38Cc 40 ~ 50 31.97 ± 9.48Ab 11.80 ± 3.41Bd 50 ~ 60 10.59 ± 0.82Ac 5.11 ± 0.75Be 60 ~ 70 4.14 ± 0.81Ad 总计
Total243.21 ± 24.43A 171.39 ± 9.93B 114.90 ± 4.60C 108.03 ± 3.71C 67.45 ± 5.28D 74.08 ± 2.62D 氮含量
Nitrogen content/
(g·kg−1)0 ~ 10 22.92 ± 0.64Aa 23.45 ± 1.37Aa 12.78 ± 0.69Ba 12.73 ± 1.47Ba 2.59 ± 0.06Ca 3.30 ± 0.67Ca 10 ~ 20 19.82 ± 1.17Ba 25.12 ± 1.37Aa 5.82 ± 1.42Cb 5.11 ± 0.78Cb 0.59 ± 0.12Eb 0.70 ± 0.07Db 20 ~ 30 20.92 ± 0.79Aa 6.79 ± 0.77Bb 1.09 ± 0.03Cc 1.32 ± 0.19Cc 0.41 ± 0.10Db 0.36 ± 0.05Dc 30 ~ 40 5.75 ± 0.54Ab 2.50 ± 1.94Bc 0.93 ± 0.26Cc 0.55 ± 0.14Dc 0.38 ± 0.09Db 0.33 ± 0.03Dc 40 ~ 50 1.52 ± 0.01Ac 1.33 ± 0.53Ac 50 ~ 60 1.05 ± 0.14Ad 0.85 ± 0.21Bc 60 ~ 70 0.88 ± 0.16e 平均值
Mean10.41 ± 0.31A 10.01 ± 0.37A 5.16 ± 0.49B 4.93 ± 0.49B 1.00 ± 0.02C 1.17 ± 0.14C 氮储量/(t·hm−2)
Nitrogen storage/
(t·ha−1)0 ~ 10 3.07 ± 0.05Aa 3.13 ± 0.12Aa 1.31 ± 0.22Cb 1.66 ± 0.11Cb 1.60 ± 0.28Ca 2.04 ± 0.41Ba 10 ~ 20 2.69 ± 0.29Ba 3.56 ± 0.30Aa 1.94 ± 0.34Ca 1.93 ± 0.22Ca 0.39 ± 0.13Db 0.35 ± 0.03Db 20 ~ 30 2.94 ± 0.16Aa 2.14 ± 0.27Bb 0.65 ± 0.06Cc 0.64 ± 0.11Cc 0.24 ± 0.06Db 0.19 ± 0.02Dc 30 ~ 40 2.77 ± 0.07Aa 1.27 ± 0.82Bc 0.29 ± 0.08Cd 0.18 ± 0.09Cd 0.20 ± 0.20Cb 0.16 ± 0.04Cc 40 ~ 50 1.27 ± 0.07Ab 0.90 ± 0.34Acd 50 ~ 60 0.53 ± 0.05Ac 0.34 ± 0.08Bd 60 ~ 70 0.26 ± 0.06d 总计
Total13.53 ± 0.28A 11.33 ± 1.17B 4.20 ± 0.34C 4.42 ± 0.08C 2.44 ± 0.20D 2.74 ± 0.46D 注:不同小写字母表示相同湿地类型内各土层间差异显著(P < 0.05)。Note: different lowercase letters indicate significant differences among soil layers within the same wetland types (P < 0.05). 各沼泽类型土壤碳、氮储量的垂直分布格局也不同。芦苇沼泽可分为7个土壤层(按10 cm划分),草丛沼泽可分为6层,而其他沼泽可分为4层。并且土壤碳储量随土壤深度增加的变化趋势也不同。芦苇沼泽土壤碳储量在0 ~ 40 cm土壤层较高,而后各层依次递减,呈先恒定后递减的变化趋势;草丛沼泽土壤碳储量在0 ~ 30 cm较高(10 ~ 20 cm层最高),而后各层依次递减,呈先增后降趋势;杜香沼泽和落叶松泥炭藓沼泽则在0 ~ 20 cm土壤层碳储量较高(10 ~ 20 cm层最高),而后各层依次递减,两者也呈先增后降趋势;而落叶松藓类沼泽和落叶松苔草沼泽仅在0 ~ 10 cm土壤层碳储量较高,而后各层依次递减,呈递减趋势。同时,这6种沼泽中有4种(L、D、LN和LT)土壤氮储量的垂直分布格局与其土壤碳储量相一致,仅草丛沼泽和落叶松藓类沼泽土壤氮储量的垂直分布格局与其碳储量略有不同,草丛沼泽土壤氮储量呈先恒定后递减型,落叶松藓类沼泽呈上层高其他层低且恒定型。因此,园池沿岸6种沼泽类型土壤碳、氮储量垂直分布格局分别存在3种类型(先恒定后递减型(L)、先增后降型(C、D和LN)和递减型(LX和LT))和4种类型(先恒定后递减型(L和C)、先增后降型(D和LN)、先高后降且恒定型(LX)和递减型(LT))。
2.3 园池沿岸沼泽湿地生态系统碳氮储量及其分配格局
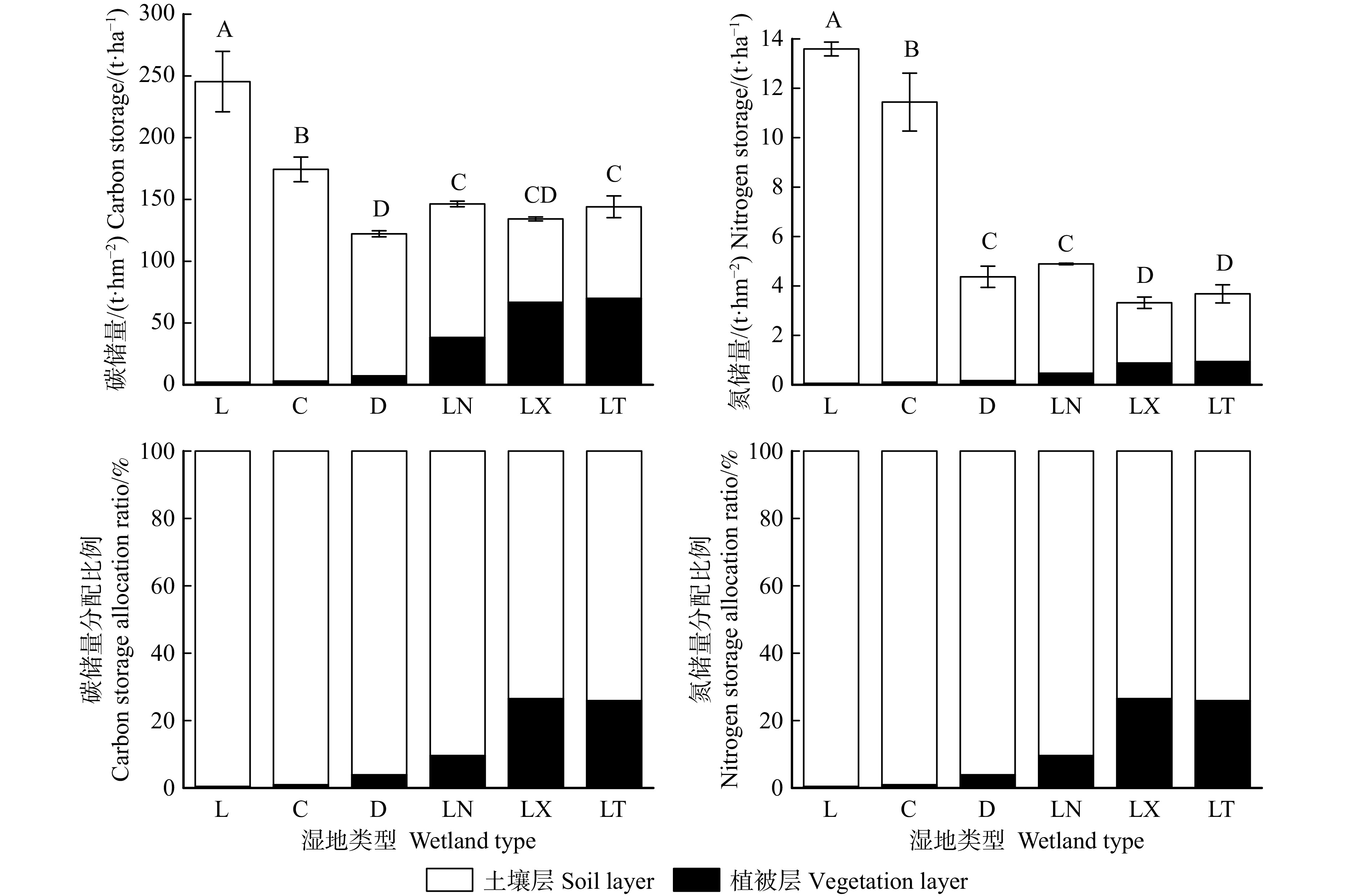
园池沿岸6种沼泽类型的生态系统碳、氮储量同样存在显著差异性(图2)。生态系统碳储量分布在122.20 ~ 245.38 t/hm2之间,其中,芦苇沼泽最高(显著高于其他5种沼泽0.4 ~ 1.0倍,P < 0.05);草丛沼泽较高(显著高于其他4种沼泽0.2 ~ 0.4倍);3种森林沼泽(LN、LX和LT)居中等(LN和LT显著高于D 17.9% ~ 19.7%,P < 0.05;LX略高于D 9.8%,P > 0.05);而杜香沼泽则最低。其生态系统氮储量分布在3.31 ~ 13.58 t/hm2,其中,芦苇沼泽和草丛沼泽相对较高(两者显著高于其他4种沼泽1.3 ~ 3.1倍,且L显著高于C 0.2倍);杜香沼泽和落叶松泥炭藓沼泽居于中等(两者显著高于其他2种沼泽0.2 ~ 0.5倍);而落叶松藓类沼泽和落叶松苔草沼泽则最低。因此,长白山园池沿岸沼泽湿地的生态系统碳储量沿湖岸至高地水分环境梯度呈先降低后升高且恒定的变化规律性,而其生态系统氮储量却呈阶梯式递减规律性。
![]() 图 2 长白山园池沿湖岸至高地环境梯度生态系统碳、氮储量及其分配不同大写字母表示不同湿地生态系统碳氮储量差异显著(P < 0.05)。Different capital letters mean significant difference between wetland types of environmental carbon and nitrogen storage (P < 0.05).Figure 2. Carbon and nitrogen storage and distribution along the environmental gradient from lakeshore to upland inYuanchi, Changbai Mountain
图 2 长白山园池沿湖岸至高地环境梯度生态系统碳、氮储量及其分配不同大写字母表示不同湿地生态系统碳氮储量差异显著(P < 0.05)。Different capital letters mean significant difference between wetland types of environmental carbon and nitrogen storage (P < 0.05).Figure 2. Carbon and nitrogen storage and distribution along the environmental gradient from lakeshore to upland inYuanchi, Changbai Mountain同时,各沼泽类型的生态系统碳、氮储量分配格局也不同。芦苇沼泽和草丛沼泽的生态系统碳、氮储量均以土壤占绝对优势地位(98.3% ~ 99.1%和99.1% ~ 99.6%),植被仅占极次要地位;杜香沼泽和落叶松泥炭藓沼泽仍以土壤占优势(73.9% ~ 94.0%和90.4% ~ 96.1%),但植被碳、氮储量占比却有所增大;而落叶松藓类沼泽和落叶松苔草沼泽的生态系统碳储量和氮储量分配格局明显不同,两者的生态系统碳储量为土壤和植被占比相近(50.3% ~ 51.5%和48.5% ~ 49.7%),生态系统氮储量却以土壤占优势(73.5% ~ 74.1%),植被氮储量占比约为1/4(25.9% ~ 26.5%)。因此,园池沿岸6种沼泽类型的生态系统碳、氮储量分配格局沿湖岸至高地水分环境梯度呈现出土壤碳、氮储量占比呈递减规律性,而植被碳、氮储量占比却呈递增规律性。
2.4 园池沿岸沼泽湿地生态系统碳氮储量的主控因子
园池沿岸6种沼泽类型的生态系统碳、氮储量主控因子不同(表5)。处于湖岸至高地环境梯度下部生境地段上的芦苇沼泽和草丛沼泽的生态系统碳储量主要受水位促进(可解释其变差的91.7%);中部生境地段上杜香沼泽和落叶松泥炭藓沼泽的生态系统碳储量主要受水位抑制(88.2%);处于上部生境地段上落叶松藓类沼泽和落叶松苔草沼泽的生态系统碳储量主要受水位波动幅度抑制(65.1%)。因此,园池沿岸沼泽湿地生态系统碳储量的主控因子沿湖岸至高地水分环境梯度发生了有规律性的变化(下部生境地段的水位促进、中部的水位抑制和上部的水位波动幅度抑制)。
表 5 不同生境下湿地碳氮储量与环境因子的逐步多元线性回归分析Table 5. Stepwise multiple linear regression analysis of carbon and nitrogen storage and environment factors of wetlands in different habitats生境类型
Habitat type指标
Index水位
Water level水位波动幅度
Water level
fluctuation
amplitude有机质
Organic
matter全氮
Total N全磷
Total P截距
InterceptR2 P F 下部生境
(长期水淹生境,L和C)
Lower habitat (permanently
flooded habitat, L and C)植被碳储量
Vegetation carbon storage−0.075** 3.205*** 0.869 < 0.01 34.033 土壤碳储量
Soil carbon storage6.975** 149.304*** 0.920 < 0.01 58.127 生态系统碳储量
Ecosystem carbon storage6.900** 152.509*** 0.917 < 0.01 56.017 中部生境
(季节性水淹生境,D和LN)
Intermediate habitat
(seasonally flooded
habitat, D and LN)植被碳储量
Vegetation carbon storage−2.010** −28.899 + 0.813 < 0.01 22.743 土壤碳储量
Soil carbon storage38.798** 91.644*** 0.847 < 0.01 28.616 生态系统碳储量
Ecosystem carbon storage−1.616** 92.705*** 0.882 < 0.01 38.400 上部生境
(无积水生境,LX和LT)
Upper habitat (no standing
water habitat, LX and LT)植被碳储量
Vegetation carbon storage135.469* 41.533* 0.804 < 0.05 21.459 土壤碳储量
Soil carbon storage0.734* 32.768* 0.894 < 0.05 19.600 生态系统碳储量
Ecosystem carbon storage−0.848* 185.136*** 0.651 < 0.05 10.336 下部生境
(长期水淹生境,L和C)
Lower habitat
(permanently flooded
habitat, L and C)植被氮储量
Vegetation nitrogen storage−0.04** 0.118*** 0.869 < 0.01 34.286 土壤氮储量
Soil nitrogen storage0.221* 10.592*** 0.809 < 0.05 22.216 生态系统氮储量
Ecosystem nitrogen storage0.217* 10.710*** 0.800 < 0.05 21.047 中部生境
(季节性水淹生境,D和LN)
Intermediate habitat
(seasonally flooded
habitat, D and LN)植被氮储量
Vegetation nitrogen storage−0.20** −0.186 + 0.837 < 0.05 26.586 土壤氮储量
Soil nitrogen storage−0.078** 0.007* 6.050** 0.963 < 0.05 65.463 生态系统氮储量
Ecosystem nitrogen storage−0.088* 8.418** 0.760 < 0.05 16.820 上部生境
(无积水生境, LX和LT)
Upper habitat (no standing
water habitat, LX and LT)植被氮储量
Vegetation nitrogen storage1.802* 0.551** 0.721 < 0.01 13.894 土壤氮储量
Soil nitrogen storage2.335* 0.056 0.680 < 0.05 11.646 生态系统氮储量
Ecosystem nitrogen storage2.239* 1.068 0.697 < 0.05 12.506 注:+表示在P < 0.1水平上显著;*表示在P < 0.05水平上显著;**表示在P < 0.01水平上显著;***表示在P < 0.001水平上显著。Notes: + indicates significance at P < 0.1 level; * indicates significance at P < 0.05 level; ** indicates significance at P < 0.01 level; *** indicates significance at P < 0.001 level. 同时,下部生境地段上的芦苇沼泽和草丛沼泽的生态系统氮储量主要受水位所促进(80.0%);中部生境地段上杜香沼泽和落叶松泥炭藓沼泽的生态系统氮储量主要受水位波动幅度所抑制(76.0%);上部生境地段上落叶松藓类沼泽和落叶松苔草沼泽的生态系统氮储量主要受土壤全氮所促进(69.7%)。因此,园池沿岸沼泽湿地生态系统氮储量的主控因子沿湖岸至高地水分环境梯度同样发生了有规律性的变化(下部的水位促进、中部的水位波动幅度抑制和上部的土壤全氮促进)。
3. 结论与讨论
3.1 湖泊沿岸沼泽湿地的植被碳氮储量
温带长白山高海拔区园池沿岸沼泽湿地的植被碳、氮储量沿湖岸至高地水分环境梯度呈递增规律,这与寒温带大兴安岭森林沼泽[46]和温带长白山低海拔区森林沼泽[47]的植被碳储量沿过渡带呈递增规律一致。
由于微地形改变了湿地水文状况,引起植物群落类型沿环境梯度过渡性分布,进而影响湿地植物生长、生物量积累和碳固定[20−22]。园池湖岸至高地同样存在着微地形引起的水分环境梯度(生长季平均水位由13.5 cm(−0.3 ~ 31.2 cm)逐渐降低至−69.3 cm(−95.0 ~ −46.7 cm))(表2和图1),植物长期适应该水分环境,形成了沿水分环境梯度依次出现的芦苇沼泽、草丛沼泽、杜香沼泽、落叶松泥炭藓沼泽、落叶松藓类沼泽和落叶松苔草沼泽的植被分布格局。其中,L和C长期处于水淹状态,限制了乔木和灌木的生存,群落结构不完整,故导致两者的生物量及碳氮储量较低;随着微地形升高,水位逐渐下降,灌木成为群落优势种,使其生物量和碳氮储量得到提高;随着微地形进一步升高,水位降低,3种森林沼泽的群落结构趋于完整(乔、灌和草),生物量积累更高,但因LN积水程度较重,落叶松生长缓慢(平均胸径仅9.25 cm)(表1),其植被碳氮储量显著低于LX和LT。故园池湖岸沼泽湿地的植被碳、氮储量呈递增趋势。此外,由于植被各器官在生长过程中对氮的需求和利用不同[48−49],使其乔木层氮储量在植被层的占比低于其碳储量占比,故导致植被碳储量沿环境梯度递增幅度大于其氮储量。
长白山高海拔区园池沿岸沼泽湿地植被碳储量(2.17 ~ 66.98 t/hm2)要低于同区低海拔森林沼泽(3.18 ~ 112.16 t/hm2)[47],但与寒温带森林沼泽(3.60 ~ 62.18 t/hm2)[50]相近,且3种森林沼泽的植被碳储量(38.34 ~ 66.98 t/hm2)与北方森林(40.0 ~ 64.0 t/hm2)[51]相近,而其他3种沼泽类型(2.17 ~ 7.30 t/hm2)则远低于其下限值;植被氮储量(0.06 ~ 0.94 t/hm2)沿水分环境梯度呈递增趋势,处于环境梯度中下部生境的草类沼泽和灌丛沼泽(0.06 ~ 0.17 t/hm2)远低于全球植被平均氮储量估计值(0.50 t/hm2)[52],而处于中上部生境的3种森林沼泽(0.47 ~ 0.94 t/hm2)一般要高于全球植被平均氮储量估计值[52](LX和LT高出73.0% ~ 88.0%,仅LN与其相近),也是3种森林沼泽氮储量相对较高。故园池沿岸各沼泽类型中森林沼泽的植被固碳、氮能力相对较强。
3.2 湖泊沿岸沼泽湿地土壤碳氮储量
长白山高海拔区园池沿岸沼泽湿地的土壤碳、氮储量沿湖岸至高地环境梯度呈阶梯式递减规律。这与长白山森林沼泽[47]、大兴安岭森林沼泽[46]、青藏高原湿地[53]以及温带干旱区湖泊湿地[40]沿低地至高地水分环境梯度总体上呈递减趋势不同。
这主要源于微地形改变了土壤水文状况和植被优势种分布[26],控制了湿地土壤有机物的输入和输出,进而影响湿地土壤碳积累[30]。芦苇沼泽(L)和草丛沼泽(C)长期处于水淹状态(生长季平均水位13.5 cm和3.1 cm)(表2),土壤以厌氧反应空间占绝对优势,抑制了微生物对土壤的分解和矿化作用,有利于泥炭形成,进而增加了土壤有机碳、氮的积累[54−56];杜香沼泽(D)和落叶松泥炭藓沼泽(LN)因所处生境地势升高,生长季水位降低(−18.7 cm和−32.8 cm),有氧反应空间和时间增加,有机碳和氮的分解速率加快[57],形成的泥炭层较薄(20 cm)且相近,故两者的土壤碳、氮储量居中等水平;而落叶松藓类沼泽(LX)和落叶松苔草沼泽(LT)所处地势最高,生长季水位较低(−58.7 cm和−69.3 cm),土壤中有氧反应空间占绝对优势,加速了土壤有机碳和氮分解[57],仅在土壤上部形成泥炭层(10 cm),使两者土壤碳、氮储量较低且相近。因此,园池沿岸沼泽湿地土壤碳、氮储量沿湖岸至高地环境梯度呈阶梯式递减趋势。
至于湖岸沼泽湿地土壤碳氮储量垂直空间分布格局存在多种类型的原因,也源于各沼泽类型在水分环境梯度上所处的位置不同,芦苇沼泽(L)积水较深,且水淹时间长,上部泥炭几乎不受有氧环境干扰,使其上部土壤的碳、氮储量较高且恒定,而下部土壤层随着深度增加,有机物输入逐渐减少,土壤碳氮储量也逐层递减,故呈先恒定后递减型分布;草丛沼泽(C)、杜香沼泽(D)和落叶松泥炭藓沼泽(LN)因所处生境积水逐渐变浅,水淹时间变短,且水位波动比较明显,表层泥炭经常处于有氧/厌氧环境之间的转换,使得表层泥炭分解加快,碳氮储量降低,故三者土壤碳氮储量呈先增后递减型分布;而落叶松藓类沼泽(LX)和落叶松苔草沼泽(LT)因地表已无积水环境,且水位波动主要发生在土壤深层,故两者土壤碳、氮储量随土壤深度呈递减型分布。
本研究发现长白山高海拔区园池沿岸沼泽湿地土壤碳储量(67.45 ~ 243.21 t/hm2)低于该区低海拔区森林沼泽(296.3 ~ 824.5 t/hm2)[47],但与我国东北地区平均土壤碳储量(105 t/hm2)[58]相比,芦苇沼泽和草丛沼泽提高了63.2% ~ 131.6%,杜香沼泽和落叶松泥炭藓沼泽略高2.9% ~ 9.4%,而两种森林沼泽(LX和LT)则降低了41.7% ~ 55.7%;土壤氮储量(2.44 ~ 13.53 t/hm2)与全球土壤平均氮储量(9.41 t/hm2)[52]相比,芦苇沼泽和草丛沼泽提高了20.4% ~ 43.8%,而其他4种沼泽类型则降低了53.0% ~ 74.1%。这是因为相比于大部分地区的土层深度(0 ~ 100 cm),中、上生境地段的4种沼泽的土壤发育时间周期短,土层深度明显更低(0 ~ 40 cm),导致其土壤碳氮储存能力相对较弱。而芦苇沼泽和草丛沼泽土层深度尽管也较低(0 ~ 70 cm),但由于其厌氧环境充足,有利于泥炭形成,因此它们的土壤碳氮储存能力相对较强。
3.3 湖泊沿岸沼泽湿地生态系统碳氮储量
长白山高海拔区园池沿岸沼泽湿地生态系统碳储量沿水分环境梯度呈先降再升后恒定型分布,这明显不同于寒温带大兴安岭森林沼泽[46]和同区低海拔森林沼泽[47]生态系统碳储量沿过渡带环境梯度呈恒定型或递减型分布;而生态系统氮储量沿水分环境梯度呈阶梯式递减型分布。
生态系统碳储量沿水分环境梯度的变化主要归因于微地形对湿地水文状况的调控,进而影响了植物群落建立和分布。这种影响不仅体现在湿地植被的生物量积累和碳固定[20−22],还显著作用于湿地土壤有机物的输入和输出及土壤的碳积累[30−31]。此外,生态系统碳储量沿过渡带的变化趋势取决于植被碳储量和土壤碳储量两者沿环境梯度递变趋势的权衡,同时受到微地形坡度大小的影响[46−47]。大兴安岭森林沼泽,地势坡度相对平缓(4° ~ 5°),土壤碳储量递减趋势与植被碳储量递增趋势相近,使得生态系统碳储量呈恒定型分布[46−47]。而在长白山森林沼泽,地势坡度相对较大(8° ~ 10°),土壤碳储量递减趋势大于植被碳储量递增趋势,加之生态系统碳储量以土壤占优势,因此生态系统碳储量沿过渡带呈递减趋势[46−47]。尽管本研究中的长白山高海拔园池沿岸沼泽湿地地势坡度也相对较缓(3° ~ 4°),但其生态系统碳储量沿湖岸至高地水分环境梯度变化趋势却兼有上述两种特征(下半段递减及上半段恒定)。这主要因为下半段的3种沼泽(芦苇沼泽、草丛沼泽和杜香沼泽)受湖泊水位波动影响强烈,厌氧反应空间不断减少且持续时间逐渐变短,导致泥炭层明显变薄,土壤碳储量的递减趋势明显大于植被碳储量的递增趋势,从而使得下半段的生态系统碳储量呈递减趋势。而处于上半段的3种森林沼泽因地势相对较高,受水位波动影响较小,土壤碳储量递减趋势与植被碳储量递增趋势相近,因此3种森林沼泽的生态系统碳储量保持相对恒定。此外,本研究还发现,长白山高海拔园池沿岸沼泽湿地生态系统氮储量沿环境梯度变化规律与其碳储量不同。这是因为氮在植被生物量中的占比远低于碳[48−49],植被的氮存储能力相对较弱,导致土壤碳储量在生态系统氮储量中占主导地位。因此,生态系统氮储量沿环境梯度呈阶梯式递减特征。综上所述,园池沿岸湿地生态系统碳、氮储量沿湖岸至高地的水分环境梯度空间分异,主要由微地形引起的水位梯度和植被类型分布共同控制,进而影响各生境地段的碳氮积累过程。
长白山高海拔区园池沿岸沼泽湿地生态系统碳储量(122.20 ~ 245.38 t/hm2)与北方森林生态系统碳储量(125 ~ 149 t/hm2)相近[51],其中芦苇沼泽和草丛沼泽较大幅度高于其上限值(15.0% ~ 64.7%)。此外,与全球陆地生态系统的平均氮储量(9.91 t/hm2)[52]相比,芦苇沼泽和草丛沼泽(11.44 ~ 13.58 t/hm2)提高了15.4% ~ 37.0%,而其他沼泽类型则有较大幅度降低(53.3% ~ 64.9%)。相比于其他地区的湿地类型,园池沿岸各沼泽类型的土壤层深度更低。因此,园池沿岸这6种沼泽类型中仅有芦苇沼泽和草丛沼泽在生态系统尺度上的碳、氮存储能力相对较强,在湖泊湿地碳汇管理中应重点加以保护。
3.4 湖泊沿岸沼泽湿地生态系统各组分碳氮储量的主控因子
园池沿岸沼泽湿地生态系统碳、氮储量的主控因子沿湖岸至高地水分环境梯度发生转换。在环境梯度下部生境地段,芦苇沼泽和草丛沼泽长期处于水淹环境,生态系统碳储量以土壤碳储量占绝对优势,且水位越深土壤有机质分解越缓慢,有利于土壤碳的积累[53−55],故其生态系统碳储量受水位所促进;在中部生境地段,杜香沼泽和落叶松泥炭藓沼泽处于季节性水淹环境,土壤碳储量相近,生态系统碳储量的差异性主要来自于植被碳储量,而后者受水位所抑制(群落结构完整性不同),故其生态系统碳储量受水位抑制;在上部生境地段,由于落叶松苔草沼泽和落叶松藓类沼泽处于非水淹环境(水位波动发生在地下),且两者生态系统碳储量中土壤和植被占比相近,又因为植被碳储量受土壤全磷含量促进(磷为植物生长限制性营养元素可促进植物生长[59−60]),土壤碳储量受土壤有机质促进(既是土壤碳储量主要来源,也能提高植物生产力及增加土壤碳输入[61]),当水位波动幅度增大,一方面增加了有氧反应空间,促进有机质分解[62],导致土壤有机质含量降低。另一方面,土壤淋溶作用增强,也会导致土壤磷的损失(表2),故水位波动幅度抑制了其生态系统碳储量。
此外,园池沿岸沼泽生态系统氮储量的主控因子与碳储量不同。处于下部生境地段上芦苇沼泽和草丛沼泽的生态系统氮储量受水位促进,且影响机制与其碳储量相同。在中部生境地段上杜香沼泽和落叶松泥炭藓沼泽,生态系统氮储量受水位波动幅度的抑制,这可能是因为水位波动幅度增大,导致上部土壤颗粒物质随淋溶损失较大,土壤密度降低(表4),从而导致土壤氮储量降低,进而降低生态系统氮储量。在上部生境地段上落叶松藓类沼泽和落叶松苔草沼泽,生态系统氮储量受土壤全氮促进,源于其生态系统氮储量以土壤氮储量占优势,且土壤全氮又是土壤氮储量的主要来源,故两者的生态系统氮储量取决于土壤氮含量。因此,园池沿岸沼泽湿地生态系统碳、氮储量沿湖岸至高地水分环境梯度的空间分异主控因子略有不同,其生态系统碳储量主要受水文状况(水位或水位波动幅度)控制,而氮储量则同时受水文状况(水位或水位波动幅度)和土壤特性的控制。
-
![]()
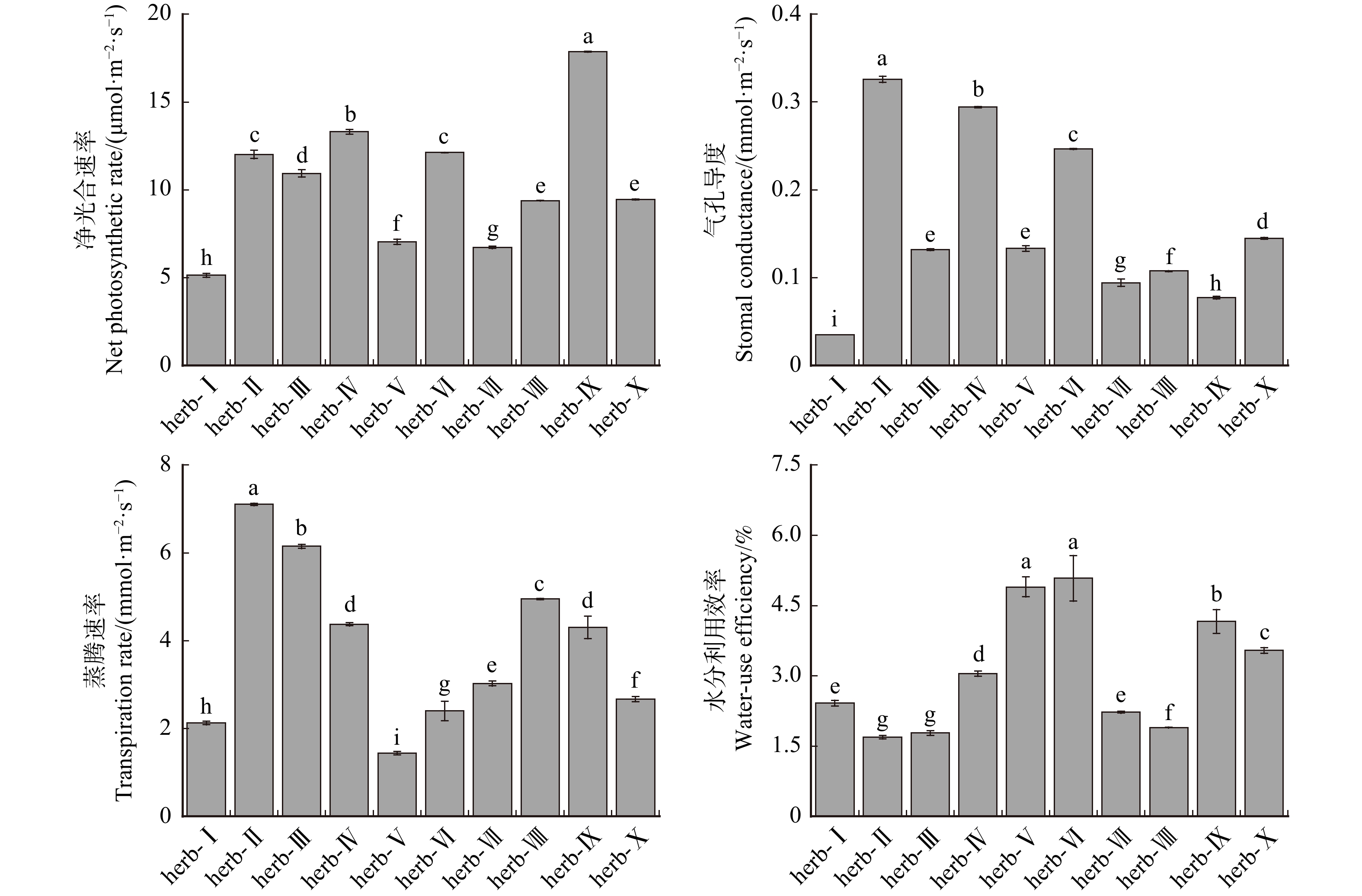
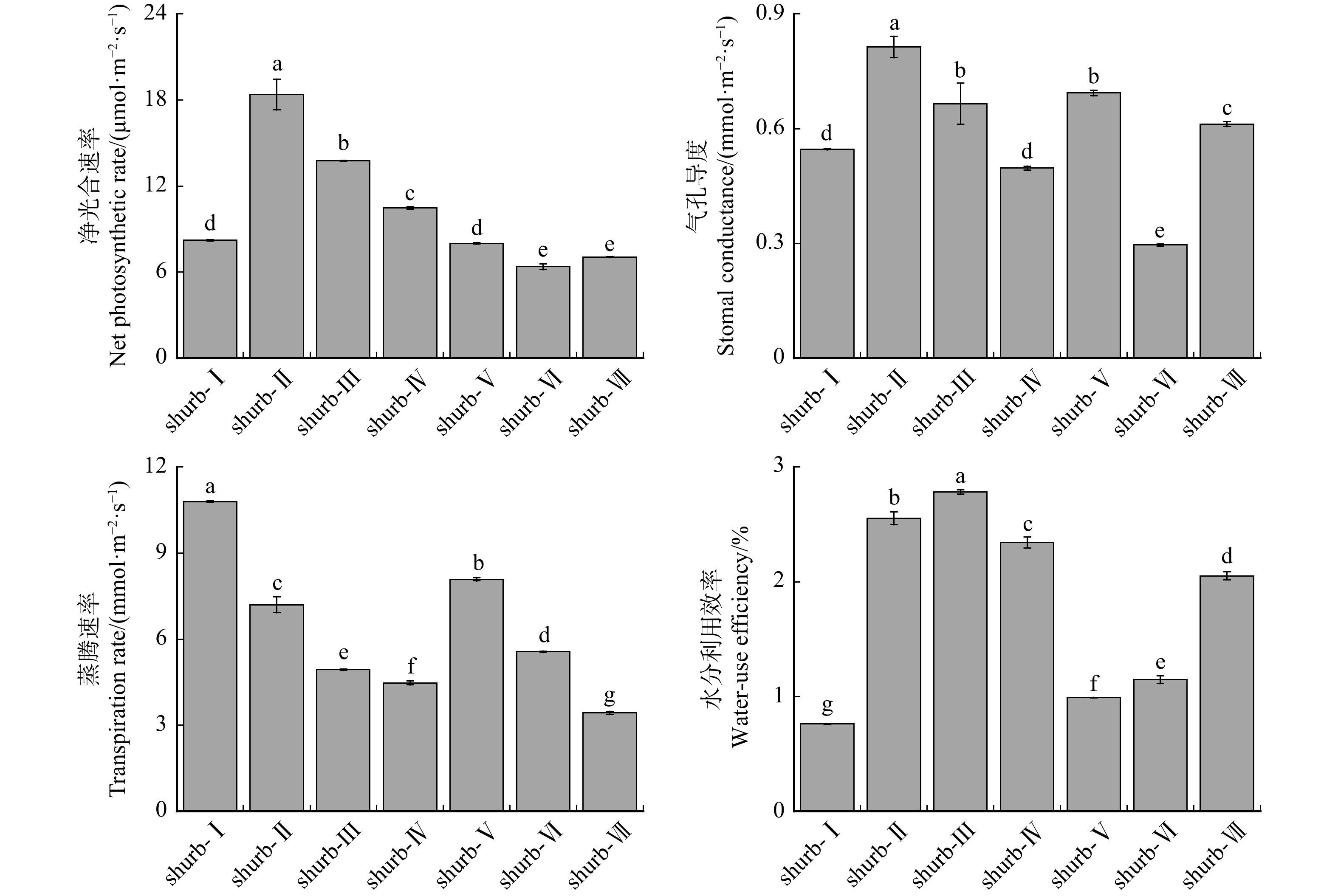
图 1 不同灌草植物的生理性状指标
herb-Ⅰ.披碱草,herb-Ⅱ.紫花苜蓿,herb-Ⅲ.草木樨,herb-Ⅳ.蜀葵,herb-Ⅴ.沙打旺,herb-Ⅵ.高冰草,herb-Ⅶ.高羊茅,herb-Ⅷ.黑麦草,herb-Ⅸ.早熟禾,herb-Ⅹ.沙蒿;shurb-Ⅰ.花棒,shurb-Ⅱ.柠条锦鸡儿,shurb-Ⅲ.沙地柏,shurb-Ⅳ.紫穗槐,shurb-Ⅴ.胡枝子,shurb-Ⅵ.沙棘,shurb-Ⅶ.沙柳。柱子上方不同小写字母表示不同植物间差异显著(P < 0.05)。下同。herb-Ⅰ is Elymus dahuricus, herb-Ⅱ is Medicago sativa, herb-Ⅲ is Melilotus officinalis, herb-Ⅳ is Alcea rosea, herb-Ⅴ is Astragalus laxmannii, herb-Ⅵ is Agropyron elongatum, herb-Ⅶ is Festuca elata, herb-Ⅷ is Lolium perenne, herb-Ⅸ is Poa annua, herb-Ⅹ is Artemisia desertorum;shurb-Ⅰ is Corethrodendron scoparium, shurb-Ⅱ is Caragana korshinskii, shurb-Ⅲ is Juniperus sabina, shurb-Ⅳ is Morpha fruticosa, shurb-Ⅴ is Lespedeza bicolor, shurb-Ⅵ is Hippophae rhamnoides, shurb-Ⅶ is Salix cheilophila. Different lowercase letters above the columns indicate significant differences between plants (P < 0.05). Same as below.
Figure 1. Physiological trait indicators of different shrub and herbaceous plants
![]()
图 2 不同草本植物的光合生理指标
Figure 2. Photosynthetic physiological indicators of different herbaceous plants
表 1 矿区土壤理化性质
Table 1 Physical and chemical properties of soil in mining area
pH值
pH value土壤密度
Soil density/
(g·cm−3)有机质
Organic matter/
(g·kg−1)全氮
Total nitrogen/
(g·kg−1)速效磷
Available phosphorus/
(mg·kg−1)全磷
Total phosphorus/
(g·kg−1)速效钾
Available potassium/
(mg·kg−1)全钾
Total potassium/
(g·kg−1)8.12 ± 0.02 1.39 ± 0.27 3.25 ± 0.30 0.60 ± 0.06 3.99 ± 0.16 0.19 ± 0.04 116 ± 12.48 4.36 ± 0.22  下载: 导出CSV
下载: 导出CSV
表 2 试验材料基本情况
Table 2 General situation of experimental materials
植物种
Plant species科名
Family name生活型
Life form植物种
Plant species科名
Family name生活型
Life form柠条锦鸡儿
Caragana korshinskii豆科
Leguminosae灌木
Shrub沙打旺
Astragalus laxmannii豆科
Leguminosae多年生草本
Perennial herb紫穗槐
Amorpha fruticosa豆科
Leguminosae落叶灌木
Deciduous shrub早熟禾
Poa annua禾本科
Gramineae一年生草本
Annual herb胡枝子
Lespedeza bicolor豆科
Leguminosae直立灌木
Upright shrub高羊茅
Festuca elata禾本科
Gramineae多年生草本
Perennial herb花棒
Corethrodendron scoparium豆科
Leguminosae半灌木
Shrubbery黑麦草
Lolium perenne禾本科
Gramineae多年生草本
Perennial herb沙棘
Hippophae rhamnoides胡颓子科
Elaeagnaceae落叶灌木
Deciduous shrub披碱草
Elymus dahuricus禾本科
Gramineae多年生草本
Perennial herb沙柳
Salix cheilophila杨柳科
Salicaceae灌木
Shrub高冰草
Agropyron elongatum禾本科
Gramineae多年生草本
Perennial herb沙地柏
Juniperus sabina柏科
Cupressaceae匍匐灌木
Creeping shrub蜀葵
Alcea rosea锦葵科
Malvaceae两年生草本
Biennial herb草木樨
Melilotus officinalis豆科
Leguminosae一年生或二年生草本
Annual or biennial herb沙蒿
Artemisia desertorum菊科
Compositae多年生草本
Perennial herb紫花苜蓿
Medicago sativa豆科
Leguminosae多年生草本
Perennial herb
下载: 导出CSV
表 3 草本植物层次分析法筛选表
Table 3 Hierarchical analysis screening table for herbaceous plants
目标层
Target level准则层
Standardized layer指标层
Indicator layer方案层
Programme level节水增汇植物筛选
Water-saving and sink-enhancing
plant screening生长状况
Growth condition (0.109 1)株高 Plant height (0.020 6) 蜀葵 Alcea rosea ( 0.0893 )
黑麦草 Lolium perenne (0.0503 )
披碱草 Elymus dahuricus (0.0800 )
沙蒿 Artemisia desertorum (0.0672 )
沙打旺 Astragalus laxmannii (0.1731 )
紫花苜蓿 Medicago sativa (0.2038 )
草木樨 Melilotus officinalis (0.1750 )
高羊茅 Festuca elata (0.0557 )
高冰草 Agropyron elongatum (0.0614 )
早熟禾 Poa annua (0.0442 )根长 Root length (0.008 8) 盖度 Coverage (0.079 7) 碳储量
Carbon stock (0.350 9)地上碳储量
Aboveground carbon stock (0.117 0 )地下碳储量
Underground carbon stock (0.117 0 )土壤碳储量
Soil carbon stock (0.117 0 )光合生理
Photosynthetic physiology (0.189 1)蒸腾速率
Transpiration rate (0.034 4)气孔导度
Stomal conductance (0.013 5)净光合速率
Net photosynthetic rate (0.044 7)水分利用效率
Water use efficiency (0.096 4)生物量
Biomass (0.350 9)地上生物量
Aboveground biomass (0.175 5)地下生物量
Underground biomass (0.175 5)注:括号内数据为指标得分。下同。Notes: data in parentheses represent the scores of indicators. Same as below.
下载: 导出CSV
表 4 灌木层次分析法筛选表
Table 4 Hierarchical analysis screening table for shrubs
目标层
Target level准则层
Standardized layer指标层
Indicator layer方案层
Programme level节水增汇植物筛选
Water-saving and sink-enhancing
plant screening生长状况
Growth condition (0.1091 )株高 Plant height ( 0.0206 )沙地柏 Juniperus sabina ( 0.1122 )
胡枝子 Lespedeza bicolor (0.0876 )
花棒 Corethrodendron scoparium (0.0676 )
柠条锦鸡儿 Caragana korshinskii (0.2405 )
紫穗槐 Amorpha fruticose (0.2613 )
沙棘 Hippophae rhamnoides0.1464 )
沙柳 Salix cheilophila (0.0844 )根长 Root length ( 0.0088 )盖度 Coverage ( 0.0797 )碳储量
Carbon stock (0.3509) 地上碳储量
Aboveground carbon stock (0.117 0 )地下碳储量
Underground carbon stock (0.117 0 )土壤碳储量
Soil carbon stock (0.117 0 )光合生理
Photosynthetic
physiology (0.1891 )蒸腾速率
Transpiration rate (0.0344 )气孔导度
Stomal conductance (0.0135 )净光合速率
Net photosynthetic rate (0.0447 )水分利用效率
Water use efficiency (0.0964 )生物量
Biomass (0.350 9)地上生物量
Aboveground biomass (0.175 5)地下生物量
Underground biomass (0.175 5)
下载: 导出CSV
-
[1] 关中美, 郝成元. 我国干旱半干旱地区脆弱生态系统及其退化成因[J]. 生态经济, 2013(9): 158−162. doi: 10.3969/j.issn.1671-4407.2013.09.035 Guan Z M, Hao C Y. The fragile ecosystem types in arid and semi-arid region of China and their degradation causes[J]. Ecological Environment, 2013(9): 158−162. doi: 10.3969/j.issn.1671-4407.2013.09.035
[2] 吴秦豫, 姚喜军, 梁洁, 等. 鄂尔多斯市煤矿区植被覆盖改善和退化效应的时空强度[J]. 干旱区资源与环境, 2022, 36(8): 101−109. Wu Q Y, Yao X J, Liang J, et al. Spatial and temporal intensity of vegetation cover improvement and degradation in coal mining areas of Erdos City[J]. Journal of Arid Land Resources and Environment, 2022, 36(8): 101−109.
[3] 周航. 桂林石灰岩矿山采损生态脆弱区复绿植物筛选与配置[D]. 昆明: 桂林理工大学, 2023. Zhou H. Screening and allocation of reforestation plants in the ecologically fragile area of limestone mining damage in Guilin, China[D]. Kunming: Guilin University of Technology, 2023.
[4] 吴金山, 张景欢, 李瑞杰, 等. 植物对干旱胁迫的生理机制及适应性研究进展[J]. 山西农业大学学报(自然科学版), 2017, 37(6): 452−456. Wu J S, Zhang J H, Li R J, et al. The plant’s physiological mechanism and adaptability to drought stress[J]. Journal of Shanxi Agricultural University (Natural Science Edition), 2017, 37(6): 452−456.
[5] Hetherington A M, Woodward F I. The role of stomata in sensing and driving environmental change[J]. Nature, 2003, 424: 901−908. doi: 10.1038/nature01843
[6] Lake J A, Woodward F I. Response of stomatal numbers to CO2 and humidity: control by transpiration rate and abscisic acid[J]. New Phytol, 2008, 179: 397−404. doi: 10.1111/j.1469-8137.2008.02485.x
[7] 余林辉, 蔡晓腾, 徐萍, 等. 植物抗旱节水: 从实验室到田间[J]. 中国科学: 生命科学, 2017, 47(1): 145−154. doi: 10.1360/N052016-00341 Yu L H, Cai X T, Xu P, et al. Drought-resistant and water-saving plants: from laboratory to field[J]. Scientia Sinica Vitae, 2017, 47(1): 145−154. doi: 10.1360/N052016-00341
[8] Deng L F, Yuan H R, Xie J, et al. Herbaceous plants are better than woody plants for carbon sequestration[J/OL]. Resources Conservation and Recycling, 2022, 184[2023−09−28]. https://doi.org/10.1016/j.resconrec.2022.106431.
[9] Yang Y, Tilman D, Furey G, et al. Soil carbon sequestration accelerated by restoration of grassland biodiversity[J]. Nature Communications, 2019, 10(1): 718. doi: 10.1038/s41467-019-08636-w
[10] Qu Y, Mu P, Zhang H, et al. Mapping QTLs of root morphological traits at different growth stages in rice[J]. Genetica, 2008, 133: 187−200. doi: 10.1007/s10709-007-9199-5
[11] Comas L H, Becker S R, Cruz V M V, et al. Root traits contributing to plant productivity under drought[J]. Frontiers in Plant Science, 2013, 4: 442.
[12] 尤云楠, 朱燕峰, 陈浮, 等. 植被类型对露天矿复垦土壤微生物固碳潜力及路径的影响[J]. 生态与农村环境学报, 2023, 39(9): 1170−1179. You Y N, Zhu Y F, Chen F, et al. Effects of vegetation types on the potential and pathway of microbial carbon sequestration in reclaimed soil of open-pit mine[J]. Journal of Ecology and Rural Environment, 2023, 39(9): 1170−1179.
[13] 王新宇, 高英志. 禾本科/豆科间作促进豆科共生固氮机理研究进展[J]. 科学通报, 2020, 65(专1): 142−149. Wang X Y, Gao Y Z. Advances in the mechanism of cereal/legume intercropping promotion of symbiotic nitrogen fixation[J]. Chinese Science Bulletin, 2020, 65(Spec.1): 142−149.
[14] 郝新忠, 郜超, 张继平, 等. 煤矿开采对榆林毛乌素沙地植被与地下水的影响[J]. 陕西林业科技, 2019, 47(4): 28−31. doi: 10.3969/j.issn.1001-2117.2019.04.007 Hao X Z, Gao C, Zhang J P, et al. Effect of coal mining on vegetation and groundwater in Mu Us Sandy Land of Yulin[J]. Shannxi Forest Science and Technology, 2019, 47(4): 28−31. doi: 10.3969/j.issn.1001-2117.2019.04.007
[15] 侯健. 西湾露天煤矿建设项目环境评价及生态保护策略研究[D]. 杨凌: 西北农林科技大学, 2018. Hou J. The construction project study of environmental assessment and ecological protection strategy for Xiwan Opencast Coal Mine[D]. Yangling: Northwest A&F University, 2018.
[16] 白晓霞, 郭志军, 李沛, 等. 陕北旱区榆林市公园绿地植物群落调查[J]. 贵州农业科学, 2021, 49(5): 116−122. doi: 10.3969/j.issn.1001-3601.2021.05.020 Bai X X, Guo Z J, Li P, et al. Investigation on park greenbelt plant community in Yulin City of arid area in northern Shananxi[J]. Guizhou Agricultural Sciences, 2021, 49(5): 116−122. doi: 10.3969/j.issn.1001-3601.2021.05.020
[17] 项元和, 于晓杰, 魏勇明. 露天矿排土场生态修复与植被重建技术[J]. 中国水土保持科学, 2013, 11(增刊1): 48−54. Xiang Y H, Yu X J, Wei Y M. Techniques ecological restoration and plant restoration on spoil dump of modern strip mine[J]. Science of Soil and Water Conservation, 2013, 11(Suppl. 1): 48−54.
[18] Ye S. Research progress on vegetation ecological restoration of abandoned land in coal mine area[J/OL]. Iop Conference Series: Earth and Environmental Science, 2021, 781(3): 032002[2023−09−28]. https://doi.org/10.1088/1755-1315/781/3/032002.
[19] 张永娥, 余新晓, 陈丽华, 等. 北京西山侧柏林冠层不同高度处叶片水分利用效率[J]. 应用生态学报, 2017, 28(7): 2143−2148. Zhang Y E, Yu X X, Chen L H, et al. Foliar water use efficiency of platycladus orientalis of different canopy heights in Beijing Western Mountains area, China[J]. Chinese Journal of Applied Ecology, Jul, 2017, 28(7): 2143−2148.
[20] Deng L, Han Q S, Zhang C, et al. Above-ground and below-ground ecosystem biomass accumulation and carbon sequestration with Karagana korshinskii Kom. plantation development[J]. Land Degradation & Development, 2017, 28(3): 906−917.
[21] 舒洋. 大兴安岭落叶松林碳储量监测及碳层分配特征研究[D]. 呼和浩特: 内蒙古农业大学, 2017. Shu Y. Study on the distribution characteristics of carbon layer and carbon storage monitoring in Larix gmelini forest in Daxing’anling[D]. Hohhot: Inner Mongolia Agricultural University, 2017.
[22] 于欢, 魏天兴, 陈宇轩, 等. 黄土丘陵区典型人工林土壤有机碳储量的分布特征[J]. 北京林业大学学报, 2023, 45(12): 100−107. doi: 10.12171/j.1000-1522.20230097 Yu H, Wei T X, Chen Y X, et al. Distribution characteristics of soil organic carbon stocks in typical plantations in loess hilly region of northwestern China[J]. Journal of Beijing Forestry University, 2023, 45(12): 100−107. doi: 10.12171/j.1000-1522.20230097
[23] 严慧玲, 刘东明, 李作恒, 等. 层次分析法在河北省太行山区植被恢复植物种类筛选中的运用[J]. 植物研究, 2015, 35(5): 751−758. doi: 10.7525/j.issn.1673-5102.2015.05.019 Yan H L, Liu D M, Li Z H, et al. Analytic hierarchy process in plant species selection of revegetation in the Taihang Mountains in Hebei Province[J]. Bulletin of Botanical Research, 2015, 35(5): 751−758. doi: 10.7525/j.issn.1673-5102.2015.05.019
[24] 赵楠, 梁倩雨, 郭丽丽, 等. 基于AHP的榆林市木本彩叶植物综合评价[J]. 陕西农业科学, 2023, 69(3): 110−116. doi: 10.3969/j.issn.0488-5368.2023.03.023 Zhao N, Liang Q Y, Guo L L, et al. Comprehensive evaluation of woody color-leafed plants in Yulin based on AHP[J]. Shaanxi Jpural of Agricultural Sciences, 2023, 69(3): 110−116. doi: 10.3969/j.issn.0488-5368.2023.03.023
[25] 于晓燕, 宋宇辰, 魏光普, 等. 基于层次分析和模糊综合评判的尾矿库植物群落评价[J]. 稀土, 2020, 41(5): 70−79. Xu X Y, Song Y C, Wei G P, et al. Assessment of plant community around Baotou Lree Tailings Reservoir based on AHP and fuzzy comprehensive evaluation method[J]. Chinese Rare Earths, 2020, 41(5): 70−79.
[26] 廖立国, 谭正洪, 蒋龙, 等. 基于过程模型的泰国热带森林碳通量动态模拟解析[J]. 热带生物学报, 2021, 12(3): 279−288. Liao L G, Tan Z H, Jiang L, et al. Dynamic analysis of carbon flux of tropical forest in Thailand based on process model[J]. Journal of Tropical Biology, 2021, 12(3): 279−288.
[27] 渠春梅, 韩兴国, 苏波, 等. 云南西双版纳片断化热带雨林植物叶片δ ~ (13)C值的特点及其对水分利用效率的指示[J]. 植物学报, 2001, 43(2): 186−192. Qu C M, Han X G, Su B, et al. The characteristics of foliarδ13C values of plants and plant water use efficiency indicated byδ13C values in two fragmented rainforests in Xishuangbanna, Yunnan[J]. Acta Botanica Sinica, 2001, 43(2): 186−192.
[28] 王芳, 汪左, 黄静, 等. 安徽省农田水分利用效率时空特征及其与气候因子的关系[J]. 生态学报, 2018, 38(17): 6268−6279. Wang F, Wang Z, Huang J, et al. Spatio-temporal distribution characteristics of cropland water use efficiency and its relationship with climate in Anhui Province[J]. Acta Ecologica Sinica, 2018, 38(17): 6268−6279.
[29] 李机密, 黄儒珠, 王健, 等. 陆生植物水分利用效率[J]. 生态学杂志, 2009, 28(8): 1655−1663. Li J M, Huang R Z, Wang J, et al. Water use efficiency of terrestrial plants[J]. Chinese Journal of Ecology, 2009, 28(8): 1655−1663.
[30] 张琳婷, 姜德刚, 刘建辉, 等. 基于光合特性的海岛植被修复适生植物的筛选[J]. 应用海洋学学报, 2021, 40(3): 428−436. doi: 10.3969/J.ISSN.2095-4972.2021.03.008 Zhang L T, Jiang D G, Liu J H, et al. Screening of adaptable plants for island vegetation restoration based on photosynthetic characteristics[J]. Journal of Applied Oceanography, 2021, 40(3): 428−436. doi: 10.3969/J.ISSN.2095-4972.2021.03.008
[31] 张娜, 张巍, 陈玮, 等. 大连市6种园林树种的光合固碳释氧特性[J]. 生态学杂志, 2015, 34(10): 2742−2748. Zhang N, Zhang W, Chen W, et al. Carbon sequestration and oxygen release capabilities of six garden tree species in Dalian[J]. Chinese Journal of Ecology, 2015, 34(10): 2742−2748.
[32] Comstock J P. Hydraulic and chemical signalling in the control of stomatal conductance and transpiration[J]. Journal of Experimental Botany, 2002, 53: 195−200. doi: 10.1093/jexbot/53.367.195
[33] Schachtman D P, Goodger J Q D. Chemical root to shoot signaling under drought[J]. Trends in Plant Science, 2008, 13(6): 281−287. doi: 10.1016/j.tplants.2008.04.003
[34] Sah S K, Reddy K R, Li J. Abscisic acid and abiotic stress tolerance in crop plants[J]. Frontiers in Plant Science, 2016, 7: 571.
[35] 符亚儒, 张继平, 董强, 等. 陕北沙区煤矸石废弃地的植被恢复技术研究[J]. 西北林学院学报, 2012, 27(5): 178−183. doi: 10.3969/j.issn.1001-7461.2012.05.34 Fu Y R, Zhang J P, Dong Q, et al. Vegetation restoration technologies in the coal gangue waste lands in sandy areas of northern Shaanxi[J]. Journal of Northwest Foresty University, 2012, 27(5): 178−183. doi: 10.3969/j.issn.1001-7461.2012.05.34
[36] 李晋川, 王翔, 岳建英, 等. 安太堡露天矿植被恢复过程中土壤生态肥力评价[J]. 水土保持研究, 2015, 22(1): 66−71, 79. Li J C, Wang X, Yue J Y, et al. Evaluation on soil ecologic fertility during vegetation succession in Antaibao Open Pit[J]. Research of Soil and Water Conservation, 2015, 22(1): 66−71, 79.
[37] 陈来红, 马万里. 霍林河露天煤矿排土场植被恢复与重建技术探讨[J]. 中国水土保持科学, 2011, 9(4): 117−120. doi: 10.3969/j.issn.1672-3007.2011.04.021 Chen L H, Ma W L. Discussion on vegetation restoration and reconstruction in dumping site of open coal mine in Huolinhe[J]. Science of Soil and Water Conservation, 2011, 9(4): 117−120. doi: 10.3969/j.issn.1672-3007.2011.04.021
[38] 许爱云, 曹兵, 谢云. 干旱风沙区煤炭基地12种草本植物对干旱胁迫的生理生态响应及抗旱性评价[J]. 草业学报, 2020, 29(10): 22−34. doi: 10.11686/cyxb2020114 Xu A Y, Cao B, Xie Y. Physiological-ecological responses of tweive herbaceous plant species under drought stress and evaluation of their drought resistance when planted in coal producting basis in arid windy and sandy areas[J]. Acta Prataculturae Sinica, 2020, 29(10): 22−34. doi: 10.11686/cyxb2020114
[39] 宋曦, 王金成, 井明博, 等. 紫花苜蓿对陇东黄土高原油污土壤场地生态修复的综合响应[J]. 草业科学, 2019, 36(7): 1754−1764. Song X, Wang J C, Jing M B, et al. Comprehensive response of Medicago sativa when used to ecologically remediate a site in the eastern region of the Loess Plateau contaminated with different concentrations of oil[J]. Pratacultural Science, 2019, 36(7): 1754−1764.
-
期刊类型引用(1)
1. 卜弘毅,朱正杰,崔素珍,张明,李成之,许志敏,李典鹏,曹学城,孙天一,雷礼纲. 洪泽湖不同湿地类型表层土壤与沉积物碳氮含量特征分析. 湿地科学与管理. 2025(01): 45-51 .  百度学术
百度学术
其他类型引用(0)
计量
- 文章访问数: 170
- HTML全文浏览量: 18
- PDF下载量: 27
- 被引次数: 1
