
Overexpression of poplar PtoXTH34 gene to enhance drought resistance of tobacco
-
摘要:目的
探究杨树木葡聚糖内转糖苷酶/水解酶基因PtoXTH34在植物响应干旱胁迫过程中发挥的功能。
方法将杨树PtoXTH34基因在烟草中异源表达,并进行干旱胁迫,分析过表达烟草表型以及光合作用、抗逆等指标的变化。
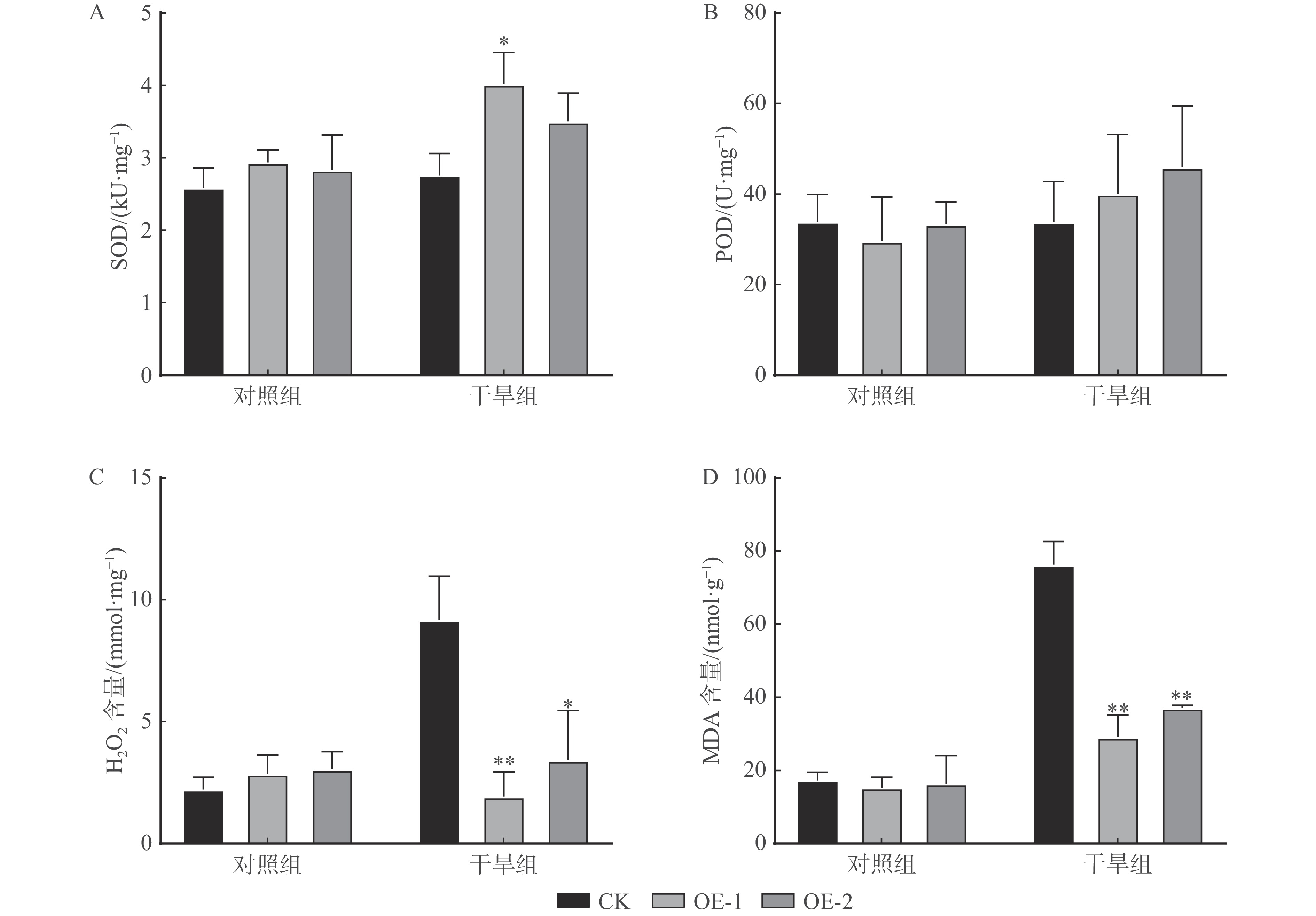
结果(1)过表达PtoXTH34基因能够显著降低离体烟草叶片的失水率。(2)干旱处理14 d后,对照植株出现明显的枯萎现象,而过表达PtoXTH34烟草叶片生长良好,仍为绿色,且过表达PtoXTH34植株的株高和茎直径均显著高于对照植株。(3)干旱处理14 d后,对照植株的净光合速率、气孔导度和蒸腾速率均趋近于0,而过表达PtoXTH34烟草的各项光合指标均极其显著高于对照植株,且高于干旱处理前。(4)干旱处理14 d后,对照植株体内的H2O2和MDA含量均出现大幅上升,且显著高于过表达PtoXTH34植株,对照植株出现活性氧积累现象,且叶片损伤程度较大,而过表达PtoXTH34植株并未出现此现象。
结论过表达PtoXTH34烟草通过降低叶片失水率,提升光合作用,清除活性氧积累,进而提高植株的抗旱性。本研究为揭示杨树PtoXTH34的基因功能奠定了基础,丰富了林木分子育种的基因资源。
Abstract:ObjectiveThis study explored the effects of xyloglucan endoglycosidase/hydrolase gene PtoXTH34 in response to drought stress in poplar.
MethodThe PtoXTH34 of poplar was heterologously expressed in tobacco to analyze the changes in phenotype and parameters related to photosynthesis and stress resistance of transgenic tobacco under drought stress.
Result(1) Overexpression of PtoXTH34 gene can significantly reduce the rate of water loss in isolated tobacco leaves. (2) After 14 d of drought treatment, the control tobacco showed obvious wilting, while the leaves of overexpressing PtoXTH34 plants grew well and remained green. The height and stem diameter of plants overexpressing PtoXTH34 were significantly higher than those of control. (3) After 14 d of drought treatment, the net photosynthetic rate, stomatal conductivity, and transpiration rate of control were close to 0, while the photosynthetic indexes of overexpressing PtoXTH34 tobacco were significantly higher than those of control, and higher than before drought treatment. (4) After 14 d of drought treatment, both H2O2 and MDA contents in control increased significantly and were significantly higher than those in overexpressing PtoXTH34 plants, and the control plants showed accumulation of reactive oxygen species and significant leaf damage, while the overexpressed PtoXTH34 plants did not exhibit this phenomenon.
ConclusionOverexpression of PtoXTH34 in tobacco enhances drought resistance of plants by reducing water-loss rate of leaves, improving photosynthesis, and clearing accumulation of reactive oxygen species. This study lays the foundation for gene function of PtoXTH34 and enriches genetic resources for molecular breeding of trees.
-
Keywords:
- poplar /
- gene expression /
- PtoXTH34 /
- drought resistance /
- photosynthesis /
- tobacco
-
近年来海绵城市在国家的支持引导下发展迅猛,出台了一系列技术指导文件,并设立试点城市,逐步进行探索 [1]。我国海绵城市研究的热点主要集中在水体污染、城市排水、生态修复、低影响开发、水文安全、绿色城市等方向[2]。自2014年启动海绵城市国家计划以来,海绵城市试点城市的城市雨水管理规划实践需要大量的资金投入,由于海绵城市的建设是建立在技术与经济的可行性之上的,因此有必要对海绵城市建设的成本和效益开展研究[3]。
相关研究表明,相比于传统雨洪管理技术,LID的应用可降低15% ~ 80%的开发成本[4]。中国城市LID设施的成本效益存在区域差异[5],可以基于成本效益优化LID的布局方式[6]。LID措施的经济效益主要来自减少暴雨径流排放许可费的直接效益和控制雨洪、减少雨污排放管道和后续水质净化费用等间接效益[7-8],以及雨水径流通过LID措施如下沉式绿地入渗回馈地下水,也可以间接产生经济效益[9],并且LID措施可在场地清理、管道安装和防腐上节约大量成本[10]。
综观之前的研究,国内关于海绵城市经济效益方面还有欠缺,目前国内对于公园绿地径流收集灌溉利用效益成本的研究较少,通常绿地灌溉方面的研究都没有与海绵城市的建设标准相结合,有一部分海绵城市经济研究中是以雨水灌溉利用作为效益的一部分,但又缺乏雨水径流收集的测算。
半湿润地区的降雨量小于蒸发量,绿地面临缺水的问题。半湿润地区的降雨多集中于夏季,夏季又是园林植物需水量较大的季节,因此在夏季收集径流用于植物灌溉,这一策略符合半湿润地区的降雨特征和园林植物的生理需要[11]。但是,这个问题在经济上是否可行和如何做到,让收集利用设施能够达到经济效益的最大化,用定性分析无法得出准确结论,需要用定量的方法做出详细的测算和深入的研究。
1. 研究区域概况
1.1 半湿润地区海绵城市建设的特殊性
半湿润地区是指干燥度为1 ~ 1.5的地区,包括大部分东北平原、华北平原以及黄土高原南部和青藏高原东南部,气候类型主要是温带季风气候和少部分的高原高山气候[11]。
半湿润地区具有明显的降雨特征,年降水量在400 ~ 800 mm,降雨主要集中在夏季,易造成短时间内的城市洪涝灾害,而在其他时间降雨量较少,水资源紧缺。因此,径流的收集与植物灌溉利用的需要存在时间差异,海绵绿地收集利用的可能性与效率相对较低[11]。
“渗、滞、蓄、净、用、排”是海绵城市背景下绿地建设策略的多种手段,在降雨量一定的前提下不同技术手段需由设计师进行选择,收集的径流量增加,渗透的径流量就会相应地减少,因此应根据绿地的立地条件来选择绿地建设的技术手段,将问题聚焦,将目标明确,针对半湿润地区场地的问题采用“渗、滞、蓄、净、用、排”中某一主导功能作为主要技术手段,场地内LID体系的功能将更加符合绿地自身需求[12-14]。
1.2 研究区域
迁安市位于河北省唐山市东北部,属于典型的半湿润地区气候,年平均气温10.5 ℃,年平均降雨量672.4 mm,雨季集中在夏季[15]。研究区域位于迁安市中心城区,总面积为371.870 hm2,其中东侧场地及外源场地面积为155.37 hm2,西侧场地面积216.505 hm2。按照迁安市相关部门要求,设计场地选取惠民大街与惠昌大街间的带状空间,面积为26.745 hm2,汇水区域位于滨湖东路东侧,面积为155.37 hm2,灌溉区域为东西侧场地范围内的公园绿地,面积为155.410 hm2(图1)。根据地勘显示滨湖东路绿地下层的土壤表层是层厚约为0.5 m的素填土;第2层是层厚约为0.6 m的中砂;第3层是层厚约为0.7 m的稍密卵石;第4层为密实卵石。表层耕土呈现松散状态,下层土壤渗透性能较好[13]。
2. 数据与方法
2.1 数据说明
本文研究区域为迁安市滨湖东路绿地,研究数据包括迁安市2014—2021年逐日降雨量数据和逐日温度数据(来源于中国地面国际交换站气候资料日值数据集)、2022—2024年逐日降雨量数据和逐日温度数据(通过cmip6预测得出)。城市雨洪管理LID技术的各项建造设计参数和成本数据主要参考文献包括住建部印发的《海绵城市建设技术指南——低影响开发雨水系统构建(试行)》[16]、《室外排水设计标准》[17]、《迁安市海绵城市专项规划(2015—2030)(修编稿)》[18]、《迁安市城市排水(雨水)防涝综合规划说明书》[19]、《迁安市中心城区雨水工程规划图》[20]、《国家建筑标准设计图集10SS705:雨水综合利用》[21]和《国家建筑标准设计图集05S804:矩形钢筋混凝土蓄水池》[22]。
2.2 研究方法
本文运用的主要研究方法包括控制变量法、容积法、对比分析法、统计分析法和文献查阅法。首先利用气象资料及相关文献资料分析研究区域的降雨类型及特征,然后依据城市雨洪管理LID技术相关规范及指南文件,确定设计情景下的不同雨水调蓄池数量1 ~ 27个共27种方案(根据研究区域径流调蓄容积和单个雨水调蓄池容积确定方案);结合逐日降雨数据与逐日温度,计算园林植物灌溉净需水量、研究区域内园林植物总灌溉需水量;结合逐日降雨数据和径流系数,计算研究区域内雨水径流量,再根据设置的不同方案(不同数量的雨水调蓄池)得出雨水调蓄池收集量;最后计算研究区域内自来水灌溉成本和利用收集的雨水径流与自来水共同灌溉成本,得出后者相对于前者的节约成本。统计2014—2024年的计算结果,确定最佳方案。
2.2.1 调蓄容积计算
滨湖东路绿地设计以满足绿地基本功能为前提,以消纳内、外部雨水径流为目的,场地内部设置的“蓄用系统”是以利用为主要功能的低影响开发雨水系统[13,23-24]。
雨水径流控制量是雨洪管理视角下的滨水绿地空间布局的重要依据,根据国家和地方的海绵城市建设相关文件确定其径流控制率、设计降雨量,以此确定绿地的径流调蓄容积,为低影响开发体系预留相应的“海绵空间”[16]。低影响开发设施计算采用容积法[16]:
V=10HφF 式中:V为设计调蓄容积(m3),H为设计降雨量(mm),φ为综合雨量径流系数,F为汇水面积(hm2)。
设计中采用的综合径流计算公式[24]:
φ=(∑Sψ)/S 式中:S为面积,ψ为雨量径流系数。
根据《海绵城市建设技术指南》中提供的各汇水面雨量径流系数φ的取值范围,绿地取值为0.15、非铺砌的土路面取值为0.30、混凝土或沥青路面及广场取值为0.80[16]。
根据《迁安市海绵城市专项规划(2015—2030)(修编稿)》[18]滨湖东路绿地年径流总量控制率,场地外部为85%(设计降雨量为42.6 mm)、场地内部为76%(设计降雨量为29.6 mm)。根据《迁安市城市排水(雨水)防涝综合规划说明书》[19]和迁安市中心城区雨水工程规划图[20],东侧内、外部汇水面积共计155.37 hm2,计算得到东侧场地低影响开发雨水系统调蓄容积为40 361.71 m3(表1)。其中:内部径流量为1 185.48 m3,外部径流量为39 176.23 m3。外部径流均由管网流入场地内,且外部径流高程较低、流速较快。如果由绿地承接外部径流容易导致绿地遭受冲刷,也不利于地表景观的营造,因此由雨水收集设施直接收纳径流。
表 1 滨湖东路东侧绿地雨水径流控制量Table 1. Amount of rainwater runoff control on the east side of Binhu East Road设计降雨量
Design rainfall/mm汇水区域名称
Name of catchment area汇水面积/hm2
Catchment area/ha综合雨量径流系数Comprehensive rainfall
runoff coefficient设计调蓄容积
Design storage capacity/m3总计
Total/m342.6 外部汇水面(汇流入东侧场地)External catchment surface (flow into the east site) 地块1 Plot 1 15.30 0.15 ~ 0.80 4 330.93 40 361.71 地块2 Plot 2 60.42 0.15 ~ 0.80 18 118.41 地块3 Plot 3 35.10 0.15 ~ 0.80 10 660.65 滨湖东路
Binhu East Road10.00 0.80 3 408.00 阜安大街
Fu’an Street3.40 0.80 1 158.72 惠兴大街
Huixing Street4.40 0.80 1499.52 29.6 内部汇水面
Internal catchment surface滨湖东路绿地(东侧)
Binhu East Road greenland (east side)26.75 0.15 1 185.48 2.2.2 植物灌溉需水量计算
园林植物灌溉需水量是指为了满足植物净需水量和为抵消水量分布不均匀而施加于园林植物的水量。园林植物需水量通常通过园林系数法进行计算,公式 [25]为:
PWR=ET0×KL 式中:PWR为园林植物需水量(mm/d),ET0为参考作物腾发量(mm/d),KL为园林系数。
参考作物腾发量(ET0)通常通过区域常规观测气象数据作为依据(月降雨量/日数),在气象数据资料缺失的情况下也可利用平均气温对参考作物腾发量进行预测。
ET0=0.187t+2.6757 式中:t为月平均温度(℃)。
园林系数(KL)计算公式为:
KL=KsKdKmc 式中:Ks为园林植物种类因子;Kd为园林植物密度因子;Kmc为园林小气候因子。
将不同类型植物的各因子值(表2)带入园林系数计算公式中,可以得到各园林植物的园林系数(表3)。
表 2 各类园林植物系数相关指标Table 2. Indexes related to the coefficients of various garden plants植被类型
Vegetation type种类因子
Type factor(Ks)密度因子
Density factor(Kd)小气候因子Microclimate factor(Kmc) 长势好
Good growth长势一般
Average growth长势差
Bad growth长势好
Good growth长势一般
Average growth长势差
Bad growth长势好
Good growth长势一般
Average growth长势差
Bad growth乔木 Tree 0.90 0.50 0.20 1.30 1.00 0.50 1.40 1.00 0.50 灌木 Shrub 0.70 0.50 0.20 1.10 1.00 0.50 1.30 1.00 0.50 地被植物
Ground-cover plant0.90 0.50 0.20 1.10 1.00 0.50 1.20 1.00 0.50 乔−灌−草
Tree-shrub-grass0.90 0.50 0.20 1.30 1.00 0.60 1.40 1.00 0.50 冷季型草
Cool season grass0.80 1.00 1.00 0.60 1.20 1.00 0.80 注:表2来源于参考文献[25]。Note: Tab. 2 is cited from reference [25]. 园林植物的需水量在自然条件下主要包括有效降雨量和人工灌溉补水量,因此单位面积园林植物灌溉需水量,即单位面积园林植物净需水量。其计算公式[26]如下:
PWRnet=Ki(PWR日−Re日) 式中:PWRnet为植物净需水量(m3);Ki为浇灌补偿系数,取Ki = 2.7[26];PWR日为植物日需水量(m3);Re日为日有效降雨量(mm/d)。
有效降雨量指的是植物在灌溉季节内实际上能够利用的降雨量。日有效降雨量计算表达式:
Re日=Rh日×RF日 式中:Rh日为历史降雨量(mm/d);RF日为有效雨量系数(%),通常为0.50 ~ 1.00。
由于基于长时段平均历史数据预测未来降雨的不确定性,通常在规划园林植物的用水需求时,可以认为不超过50%的历史降雨量是有效降雨,故研究范围内应取0.50作为有效降雨量系数[25]。
将滨湖东路绿地东、西侧的内、外场地范围内的公园绿地作为本次研究的灌溉区域,共155.41 hm2,根据单位面积植物灌溉净需水量计算公式可以确定灌溉区域植物灌溉总需水量。迁安市相对湿度为62%,夏季最热月(7月份)气温为25.2 ℃;参考作物腾发量(ET0)最大范围为5.08 ~ 6.35 mm/d。由于缺少气象数据,通过月平均气温可以预测参考作物腾发量(ET0),同时根据不同园林植物类型确定园林系数(KL)(表3),由此可确定灌溉区域2014—2024年间每周的园林植物灌溉需水量。
表 3 各园林植物的园林系数Table 3. Landscape coefficients of each garden plant植被类型
Vegetation type种类因子
Type factor (Ks)密度因子
Density factor(Kd)小气候因子Microclimate factor(Kmc) 园林系数
Garden coefficient(KL)乔木 Tree 0.90 1.30 1.40 1.64 灌木 Shrub 0.70 1.10 1.30 1.00 地被植物
Ground-cover plant0.90 1.10 1.20 1.19 乔−灌−草
Tree-shrub-grass0.90 1.30 1.40 1.64 冷季型草
Cool season grass0.80 1.00 1.20 0.96 注:表3来源于参考文献[25]。Note: Tab. 3 is cited from reference [25]. 根据每周园林植物灌溉需水量计算可得2014年园林植物灌溉需水量为865 115.94 m3;2015年园林植物灌溉需水量为851 546.84 m3;2016年园林植物灌溉需水量为798 995.80 m3;2017年园林植物灌溉需水量为996 174.02 m3;2018年园林植物灌溉需水量为889 347.78 m3;2019年园林植物灌溉需水量为630 720.81 m3;2020年园林植物灌溉需水量为784 066.80 m3;2021年园林植物灌溉需水量为646 583.98 m3;2022年园林植物灌溉需水量为623 288.49 m3;2023年园林植物灌溉需水量为741 634.92 m3;2024年园林植物灌溉需水量为589 183.65 m3。
2.2.3 雨水调蓄池径流利用量计算
雨水调蓄池是一种雨水收集设施,收集的径流可用于园林植物灌溉。蓄水池设计参考国家建筑标准设计图集10SS705《雨水综合利用》 [21]和国家建筑标准设计图集05S804《矩形钢筋混凝土蓄水池》[22],采用钢筋混凝土结构,为了将园区分为等量的灌溉面积,雨水调蓄池采用统一容积和规格,容积为1500 m3,规格为25 000 mm(长) × 15 000 mm(宽) × 4 000 mm(高)。雨水调蓄池直接与雨水管网相连,超量径流可以溢流到下一个蓄水池中。因为无法确定雨水调蓄池个数变化对节约成本的影响,需设置多个方案进行比选。根据场地调蓄容积40 361.71 m3和单个雨水调蓄池容积1500 m3得出需要27个调蓄池,因此设计27个方案;依次增加调蓄池个数,最后对其结果进行统计分析,得出最优方案。
雨水调蓄池收集量与汇水区域雨水径流量相关。当汇水区域雨水径流量小于雨水调蓄池总容积时,雨水调蓄池收集量 = 汇水区域雨水径流量;当汇水区域雨水径流量大于雨水调蓄池总容积时,雨水调蓄池收集量 = 雨水调蓄池总容积。雨水径流量计算采用容积法:
V′=10H′φ′F′ 式中:V′为雨水径流量(m3);H′为降雨量(mm);F′为汇水面积(hm2);φ′为公园绿地径流系数,根据《室外排水设计标准》[17],φ′取值0.1 ~ 0.2。
雨水调蓄池径流利用量由雨水调蓄池收集量和植物灌溉需水量共同决定,当雨水调蓄池收集量大于植物灌溉需水量时,雨水调蓄池利用量 = 植物灌溉需水量;当雨水调蓄池收集量小于植物灌溉需水量时,雨水调蓄池利用量 = 雨水调蓄池收集量。
2.2.4 成本计算
收集每次降雨的径流,径流回收后经过简单的过滤,利用收集的径流进行植物灌溉,从而减少自来水灌溉,节约市政供水,减少经济支出[27]。
计算研究区域自来水灌溉成本和利用收集的雨水径流与自来水共同灌溉成本。
自来水灌溉成本的计算公式为:
T1=W1×w 式中:T1为所有都用自来水灌溉所需的成本(元);W1为园林植物灌溉量(m3);w为迁安市水费,迁安市当前水费为5.05元/m3 。
计算利用收集的雨水径流与自来水共同灌溉成本的公式为:
T2=T3+T4 式中:T2为利用收集的雨水径流与自来水共同灌溉成本(元);T3为利用收集的雨水径流灌溉成本(元);T4为总需水量−利用径流灌溉的量,即剩余部分的自来水灌溉成本(元),当收集的雨水径流小于植物灌溉需水量时,剩余部分采用自来水灌溉。
T3=(W3/Q)×N×p 式中:W3为调蓄池径流利用量(m3);Q为4G50KDFZ-B自控自吸泵流量,取10 m3/h;N为4G50KDFZ-B自控自吸泵功率,取7.5 kW;p为迁安市电费(元/(kW·h)),迁安市当前电费为0.52元/(kW·h)。
T4=W4×w 式中:T4为当收集的雨水径流小于植物灌溉需水量时剩余部分采用自来水灌溉的成本;W4为园林植物灌溉量(m3),除去收集的雨水径流灌溉部分。
根据下面公式计算节约成本:
T=T1−T2 式中:T为节约成本(元)。
2.2.5 建设及维护成本
LID设施的建设需要资金投入作为支撑,因此成本是LID建设的重要考量因素[3]。LID设施的造价参考本项目的预算清单,每个调蓄池配备4G50KDFZ-B自控自吸泵,该型号水泵参数如下:Q(流量) = 10 m3/h,N(功率) = 7.5 kW。建设成本根据迁安市滨湖东路集雨型绿地的预算清单,雨水调蓄池建设单价为388 690.7元,自吸自控泵建设单价6 750.0元。
LID设施的维护方式和次数参考《海绵城市建设技术指南》[16],雨水调蓄池内易有沉积,每年需两次清淤、检修,每次清淤约有50 mm淤泥堆积,自吸自控泵需对潜水轴检修。维护成本根据河北省预算定额中维护价格取费,每立方米清淤单价需442.0元,自吸自控泵每次检修需635.0元。研究区域内LID设施造价维护情况见表4。
表 4 LID设施及其配件造价维护表Table 4. Cost maintenance table of lid facilities and their accessories名称
Name建设单价/元
Construction unit price/CNY维护单价/(元·a−1)
Maintenance unit price/(CNY·year−1)雨水调蓄池 Rainwater storage tank 388 690.70 16 575.00 自控自吸泵 Self-control self-priming pump 6 750.00 1 270.00 3. 结论与讨论
3.1 结 论
通过控制变量——雨水调蓄池的个数,从1个雨水调蓄池开始依次增加1个,直到27个雨水调蓄池;搭建模型计算2014—2024年共11年的结果,最终得出27个方案的年平均调蓄池收集量、年平均径流利用率和年平均节约成本(表5、图2)。
表 5 2014—2024年年平均调蓄池收集量与节约成本Table 5. Annual average collection volume of regulation and storage tank and cost savings from 2014 to 2024方案
Plan年平均调蓄池收集量Average annual storage pool collection/m3 年平均径流利用率
Average annual runoff utilization rate/%年平均节约成本/(元·a−1)
Average annual cost saving/(CNY·year−1)方案1 Plan 1 34 123.38 44.95 71 908.34 方案2 Plan 2 58 951.48 36.84 101 114.15 方案3 Plan 3 78 271.25 30.35 109 667.28 方案4 Plan 4 93 944.53 25.63 110 661.16 方案5 Plan 5 106 598.61 22.66 110 661.16 方案6 Plan 6 116 944.00 20.69 110 661.16 方案7 Plan 7 125 803.95 19.24 110 661.16 方案8 Plan 8 133 404.79 18.14 110 661.16 方案9 Plan 9 139 876.88 17.31 110 661.16 方案10 Plan 10 145 352.78 16.66 110 661.16 方案11 Plan 11 150 115.53 16.14 110 661.16 方案12 Plan 12 154 119.79 15.72 110 661.16 方案13 Plan 13 157 221.16 15.42 110 661.16 方案14 Plan 14 160 084.80 15.18 110 661.16 方案15 Plan 15 162 763.84 14.96 110 661.16 方案16 Plan 16 165 218.39 14.76 110 661.16 方案17 Plan 17 167 532.34 14.58 110 661.16 方案18 Plan 18 169 215.16 14.47 110 661.16 方案19 Plan 19 170 851.52 14.35 110 661.16 方案20 Plan 20 172 450.50 14.25 110 661.16 方案21 Plan 21 173 464.57 14.17 110 661.16 方案22 Plan 22 174 277.92 14.12 110 661.16 方案23 Plan 23 174 893.48 14.07 110 661.16 方案24 Plan 24 175 358.54 14.04 110 661.16 方案25 Plan 25 175 629.89 14.02 110 661.16 方案26 Plan 26 175 766.26 14.01 110 661.16 方案27 Plan 27 175 902.62 14.00 110 661.16 ![]() 图 2 2014—2024年年平均调蓄池收集量与节约成本Figure 2. Annual average collection volume of regulation and storage tank and cost savings from 2014 to 2024
图 2 2014—2024年年平均调蓄池收集量与节约成本Figure 2. Annual average collection volume of regulation and storage tank and cost savings from 2014 to 2024年平均调蓄池收集量随调蓄池个数的增加而增大,最大年平均调蓄池收集量为方案27(175 902.62 m2)。年平均径流利用率随调蓄池个数的增加而降低,最大年平均径流利用率为方案1(44.95%),见图2。方案1(设计1个雨水调蓄池)的年平均径流灌溉节约成本为71 908.34元;方案2(设计2个雨水调蓄池)的年平均节约成本为101 114.15元;方案3(设计3个雨水调蓄池)的年平均节约成本为109 667.28元;方案4(设计4个雨水调蓄池)的年平均节约成本为110 661.16元;方案5(设计5个雨水调蓄池)至方案27(设计27个雨水调蓄池)的年平均节约成本不再增加,皆为110 661.16元。
由计算结果可知,从方案4开始,随着雨水调蓄池的增加年平均节约成本不再变化,取方案1到方案4计算建设成本、维护成本和节约成本之间的关系。
假设第N年,建设成本(C1) + 维护成本(C2) = 节约成本(C3),根据公式C1 + C2 × N = C3 × N,计算得出结果。方案1:约第7.4年时,建设成本(C1) + 维护成本(C2) = 节约成本(C3),从第8年开始效益为54 063.34元/年。方案2:约第12.1年时,建设成本(C1) + 维护成本(C2) = 节约成本(C3),从第13年开始效益为65 424.15元/年。方案3:约第21.2年时,建设成本(C1) + 维护成本(C2) = 节约成本(C3),从第22年开始效益为56 132.28元/年。方案4:约第40.3年时,建设成本(C1) + 维护成本(C2) = 节约成本(C3),从第41年开始效益为39 281.16元/年。从最快获得效益出发,建议选择方案1。从长期效益出发,建议选择年效益更多的方案2。
3.2 讨 论
半湿润地区的降雨具有极强的季节性,其径流收集与植物灌溉需水无法做到完全匹配。如果将绿地定位为收集径流并利用径流进行植物灌溉,本文建议按照植物的单周最大灌溉需水量作为雨水收集池的总容量,在满足灌溉需求的条件下减少建设资金的投入,提高雨水收集设施的利用率。
由于研究条件所限,本文未能获取到现场径流的实测数据,数据的精确度未经过比对核算;径流储存设施的成本会根据材料选择的不同产生很大差异,本文仅根据施工预算清单中的造价进行成本测算,未能针对不同材料的造价进行比对。本次研究效益计算中未考虑通货膨胀等经济因素的影响,实际节约灌溉费用与雨水收集利用建设费用持平的时间可能比研究结果更长。海绵城市的建设要求为城市绿地增添了新的功能,也带来了一些问题与困惑,关于在半湿润地区回收雨水径流用于植物灌溉的可能性,从经济方面的考量可以提供一个新的思路,帮助设计师和决策者更加理性地面对雨水径流的处理问题,从有益于绿地的角度推动海绵城市建设健康发展。
-
![]()
图 1 PtoXTH34转基因烟草的获得过程
A ~ F. 愈伤组织分化出芽;G ~ J.转基因烟草生根培养。
Figure 1. Acquisition process of PtoXTH34 transgenic tobacco
![]()
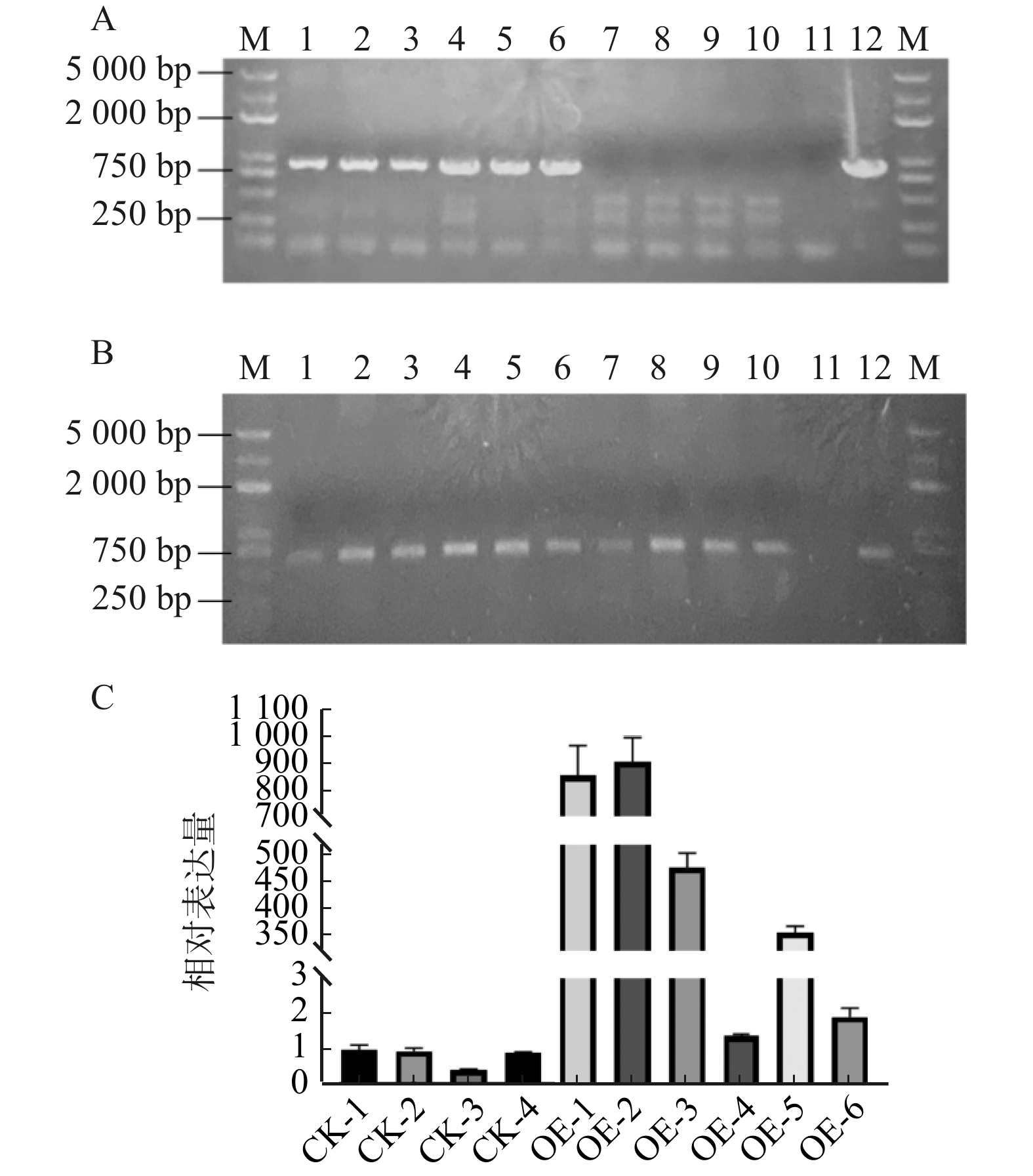
图 2 转基因烟草的筛选与鉴定
A. PtoXTH34基因结果;B. GFP基因结果;C. PtoXTH34基因表达量结果。1 ~ 6. PtoXTH34转基因株系组;7 ~ 10. 对照株系组;11. 阴性对照,12. 阳性对照。CK-1 ~ CK-4为对照组,OE-1 ~ OE-6为过表达组。
Figure 2. Screening and identification of transgenic tobacco
![]()
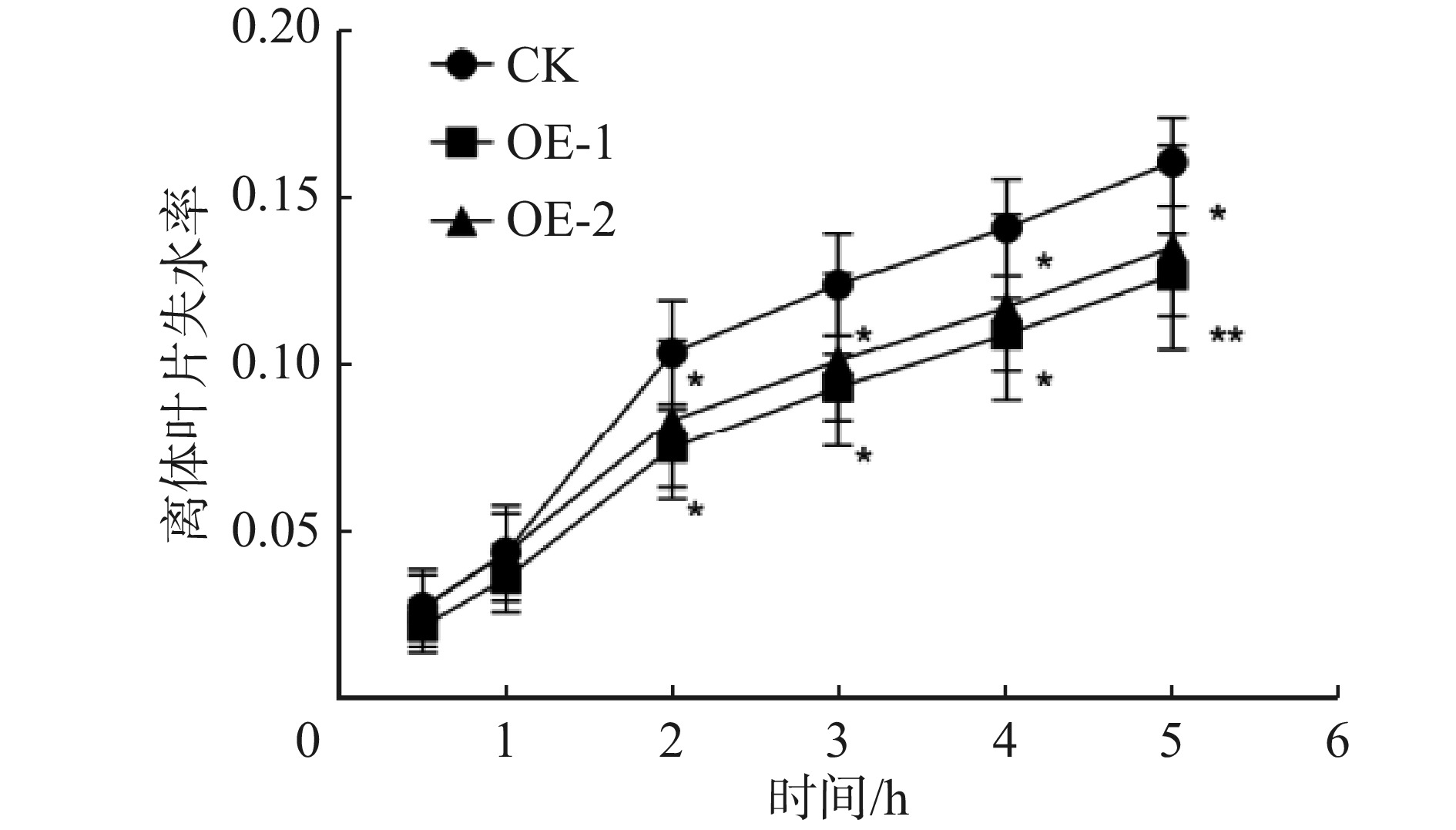
图 3 离体叶片失水率结果
*表示在0.05水平上存在显著差异;**表示在0.01水平上存在极显著差异。
Figure 3. Results of leaf water loss rate in vitro
![]()
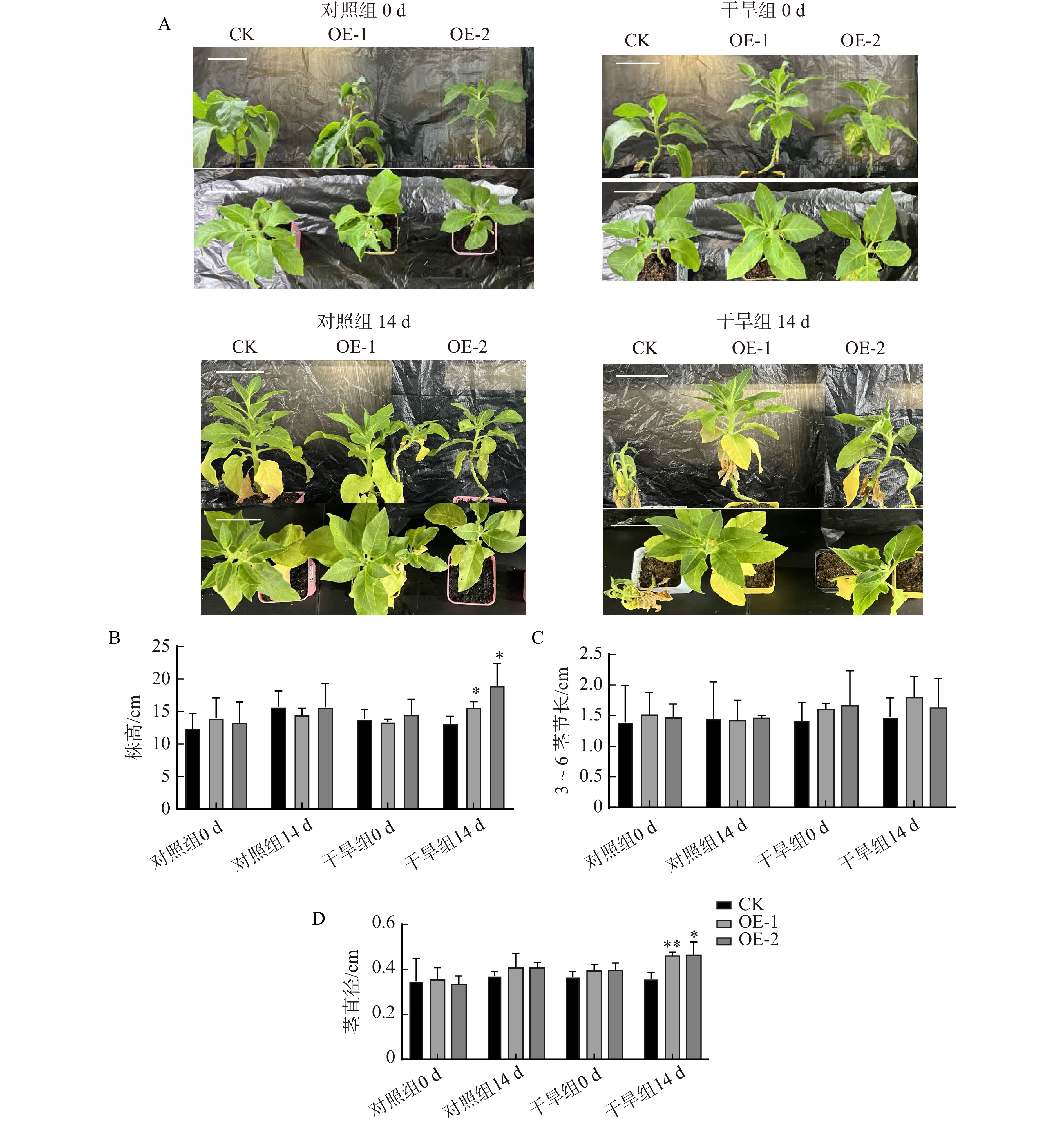
图 4 过表达PtoXTH34烟草表型分析
A. 对照组和干旱组处理前后植株形态(bar = 7 cm);B. 植物株高;C. 3 ~ 6茎节长;D. 茎直径结果
Figure 4. Phenotypic analysis of overexpressing PtoXTH34 tobacco
![]()
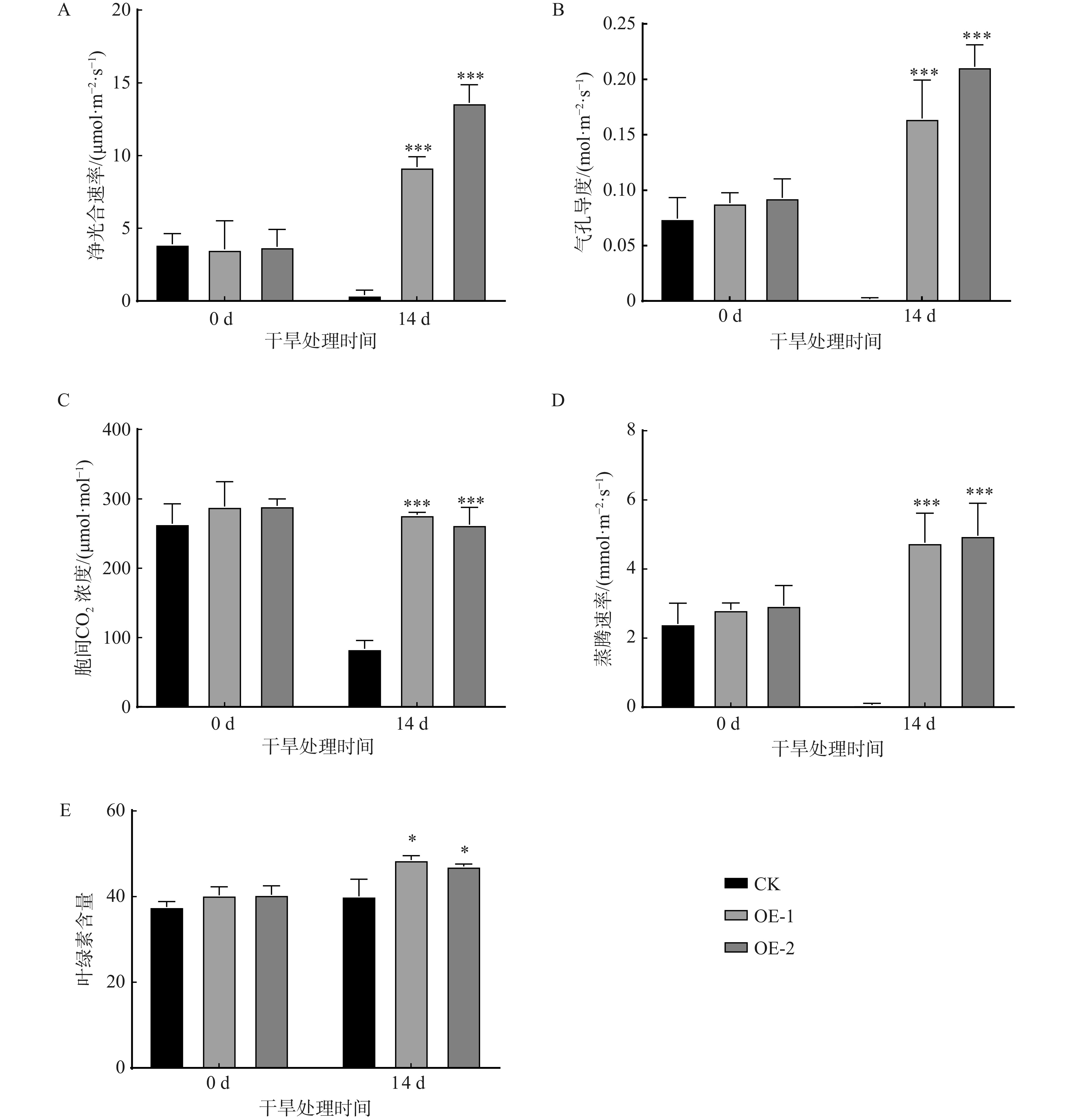
图 5 过表达PtoXTH34烟草光合指标(A ~ D)和叶绿素含量(E)测定结果
***表示在0.001水平上存在极其显著差异。
Figure 5. Measuring results of photosynthetic indicators (A−D) and chlorophyll content (E) in overexpressing PtoXTH34 tobacco
![]()
图 6 过表达PtoXTH34烟草抗逆生理指标测定结果
Figure 6. Measurement results of physiological indicators of stress resistance in overexpressing PtoXTH34 tobacco
表 1 本研究所用引物及序列
Table 1 Primers and sequences used in this study
基因名称 基因ID 正向引物(5′—3′) 反向引物(5′—3′) PtoXTH34 KY653707 AAACATGGCTGCTGCTTATC GAGCTTGCAGGGCTTCTTC GFP LOC107769044 GTGAGCAAGGGCGAGGAG CCAGCAGGACCATGTGAT PtoXTH34-RT KY653707 TCCTCGTGGATGATGTGCC CTGTCTTCTCGAGTCCACCC EF-1α LOC107788982 TGAGATGCACCACGAAGCTC CCAACATTGTCACCAGGAAGTG  下载: 导出CSV
下载: 导出CSV
-
[1] Cosgrove D J. Growth of the plant cell wall[J]. Nature Reviews Molecular Cell Biology, 2005, 6(11): 850−861. doi: 10.1038/nrm1746
[2] Rose J K C, Braam J, Fry S C, et al. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: current perspectives and a new unifying nomenclature[J]. Plant and Cell Physiology, 2002, 43(12): 1421−1435. doi: 10.1093/pcp/pcf171
[3] Baumann M J, Eklöf J M, Michel G, et al. Structural evidence for the evolution of xyloglucanase activity from xyloglucan endo-transglycosylases: biological implications for cell wall metabolism[J]. The Plant Cell, 2007, 19(6): 1947−1963. doi: 10.1105/tpc.107.051391
[4] 宣云, 赵竑绯, 郭肖颖, 等. 植物细胞壁重构酶木葡聚糖内转糖苷酶/水解酶(XTH)的研究进展[J]. 中国农学通报, 2016, 32(18): 83−88. Xuan Y, Zhao H F, Guo X Y, et al. Plant cell wall, remodeling enzyme xyloglucan endotransglucosylase/hydrolase (XTH)[J]. China Agriculture Bulletin, 2016, 32(18): 83−88.
[5] 陈燕. 龙眼三维基因组构建及细胞壁修饰基因BGAL9和XTH22在体胚发生早期的功能研究[D]. 福州: 福建农林大学, 2023. Chen Y. Three-dimensional genome construction in longan and the function of cell wall modification genes BGAL9 and XTH22 during early somatic embryogenesis[D]. Fuzhou: Fujian Agriculture and Forestry University, 2023.
[6] Han Y, Han S, Ban Q, et al. Overexpression of persimmon DkXTH1 enhanced tolerance to abiotic stress and delayed fruit softening in transgenic plants[J]. Plant Cell Reports, 2017, 36(4): 583−596. doi: 10.1007/s00299-017-2105-4
[7] Takeda T, Furuta Y, Awano T, et al. Suppression and acceleration of cell elongation by integration of xyloglucans in pea stem segments[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(13): 9055−9060.
[8] 王迪. 苹果砧木杂交后代MdXTH15、MdXTH23基因响应盐胁迫功能验证[D]. 阿拉尔: 塔里木大学, 2023. Wang D. MdXTH15, MdXTH23 of the hybrid offspring of apple rootstock functional validation of genes in response to salt stress[D]. Aral: Tarim University, 2023.
[9] 马怀龙. 甜菜木葡聚糖内转糖苷酶基因BvXTH1的克隆及功能分析[D]. 呼和浩特: 内蒙古农业大学, 2018. Ma H L. Cloning and functional analysis of xyloglucan endotransglucosylase gene BvXTH1 from sugar beets (Beta vulgaris L.)[D]. Hohhot: Inner Mongolia Agricultural University, 2018.
[10] Cho S K, Kim J E, Park J A, et al. Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants[J]. FEBS Letters, 2006, 580(13): 3136−3144. doi: 10.1016/j.febslet.2006.04.062
[11] Jiang Y, Li Y, Lu C, et al. Isolation and characterization of Populus xyloglucan endotransglycosylase/hydrolase (XTH) involved in osmotic stress responses[J]. International Journal of Biological Macromolecules, 2020, 155: 1277−1287. doi: 10.1016/j.ijbiomac.2019.11.099
[12] Johansson P, Brumer Ⅲ H, Baumann M J, et al. Crystal structures of a poplar xyloglucan endotransglycosylase reveal details of transglycosylation acceptor binding[J]. The Plant Cell, 2004, 16(4): 874−886. doi: 10.1105/tpc.020065
[13] 韩彦莎. 胡杨XTH调控烟草盐诱导肉质化及缓解重金属胁迫的机理研究[D]. 北京: 北京林业大学, 2013. Han Y S. Populus euphratica XTH mediates salinity-induced leaf succulence and alleviates heavy metal stress in tobacco plants[D]. Beijing: Beijing Forestry University, 2013.
[14] Han Y, Wang W, Sun J, et al. Populus euphratica XTH overexpression enhances salinity tolerance by the development of leaf succulence in transgenic tobacco plants[J]. Journal of Experimental Botany, 2013, 64(14): 4225−4238. doi: 10.1093/jxb/ert229
[15] Han Y , Sa G, Sun J , et al. Overexpression of Populus euphratica xyloglucan endotransglucosylase/hydrolase gene confers enhanced cadmium tolerance by the restriction of root cadmium uptake in transgenic tobacco[J]. Environmental and Experimental Botany, 2014, 100: 74−83.
[16] Cheng Z, Zhang X, Yao W, et al. Genome-wide identification and expression analysis of the xyloglucan endotransglucosylase/hydrolase gene family in poplar[J]. BMC Genomics, 2021, 22(1): 804. doi: 10.1186/s12864-021-08134-8
[17] 余凌翔, 鲁韦坤, 张加云, 等. 烤烟叶片光合速率日变化特征及其影响因素分析[J]. 气象与环境科学, 2021, 44(5): 79−86. Yu L X, Lu W K, Zhang J Y, et al. Study on diurnal variation characteristics of photosynthetic rate in tobacco leaves and its influence factors[J]. Meteorology and Environmental Science, 2021, 44(5): 79−86.
[18] 韩彦莎, 仪慧兰. 过表达胡杨XTH基因能够提高烟草抗旱性[J]. 中国生物化学与分子生物学报, 2016, 32(8): 919−925. Han Y S, Yi H L. Over-expression of Populus euphratica XTH gene enhances drought tolerance of tobacco[J]. Chinese Journal of Biochemistry and Molecular Biology, 2016, 32(8): 919−925.
[19] 陈龙, 张沿政, 李永光, 等. GmXTH23基因的克隆及抗旱性鉴定[J]. 江西农业大学学报, 2020, 42(5): 898−905. Chen L, Zhang Y Z, Li Y G, et al. Cloning of GmXTH23 gene and identification of its drought resistance[J]. Journal of Jiangxi Agricultural University, 2020, 42(5): 898−905.
[20] Han J, Liu Y, Shen Y, et al. A surprising diversity of xyloglucan endotransglucosylase/hydrolase in wheat: new in sight to the roles in drought tolerance[J]. International Journal of Molecular Sciences, 2023, 24(12): 9886. doi: 10.3390/ijms24129886
[21] 沈少炎, 吴玉香, 郑郁善. 植物干旱胁迫响应机制研究进展: 从表型到分子[J]. 生物技术进展, 2017, 7(3): 169−176. Shen S Y, Wu Y X, Zheng Y S. Review on drought response in plants from phenotype to molecular[J]. Biotechnology Progress, 2017, 7(3): 169−176.
[22] 刘波, 池明, 曹梦琦, 等. 过表达马铃薯StuPPO9基因对烟草抗旱能力的影响[J]. 作物学报, 2024, 50(9): 1−12. Liu B, Chi M, Cao M Q, et al. Impact of potato StuPPO9 gene overexpression on drought resistance in Nicotiana benthamiana[J]. [J]. Journal of Crop Sciences, 2024, 50(9): 1−12.
[23] 王依萍. 细叶百合LpWRKY49基因的克隆与抗盐和抗旱功能分析[D]. 哈尔滨: 东北林业大学, 2024. Wang Y P. Cloning and functional analysis of salt and drought resistance of the LpWRKY49 gene in Lilium pumilum[D]. Harbin: Northeast Forestry University, 2024.
[24] 陈简村, 史莹莹, 何栋, 等. 4种报春苣苔对干旱胁迫的生理响应及抗旱性评价[J]. 安徽农业大学学报, 2021, 48(5): 757−762. Chen J C, Shi Y Y, He D, et al. Physiological response and drought resistance evaluation of four kinds of primulina[J]. Journal of Anhui Agricultural University, 2021, 48(5): 757−762.
[25] 党明青, 王京平, 冉昆, 等. 泰山海棠抗旱基因MhDREB2A的克隆与功能鉴定[J]. 沈阳农业大学学报, 2022, 53(4): 462−468. Dang M Q, Wang J P, Ran K, et al. Cloning and functional idenfication of drought-resistant related gene in Malus hupehensis[J]. Journal of Shenyang Agricultural University, 2022, 53(4): 462−468.
[26] 周琪, 冯燕茹, 李嵩, 等. 小麦TaXTH-7A基因的克隆及抗旱性鉴定[J]. 农业生物技术学报, 2019, 27(9): 1521−1532. Zhou Q, Feng Y R, Li S, et al. Cloning and drought resistance identification of TaXTH-7A gene in wheat (Triticum aestivum)[J]. Journal of Agricultural Biotechnology, 2019, 27(9): 1521−1532.
[27] 李波, 曾琴, 赵丹, 等. 杜仲EuERD16基因的克隆及功能分析[J]. 基因组学与应用生物学, 2023, 42(4): 373−383. Li B, Zeng Q, Zhao D, et al. Cloning and function analysis of EuERD16 gene in Eucommia ulmoides[J]. Genomics and Applied Biology, 2023, 42(4): 373−383.
[28] 徐松华. 逆境条件下植物体内活性氧代谢研究进展[J]. 安徽农学通报, 2021, 27(21): 29−32. Xu S H. Research advances of reactive oxygen species in plants under environmental stress[J]. Anhui Agriculture Bulletin, 2021, 27(21): 29−32.
-
期刊类型引用(8)
1. 张元梅,孙桂丽,鲁艳,李利,张志浩,张栋栋. 昆仑山北坡两种优势荒漠灌木的生物量预测模型. 干旱区研究. 2024(02): 284-292 .  百度学术
百度学术
2. 赵秀英,庞志,于凤强,张建华,杨建军,郝彦卿,白小琴,杨蓉,黄鹏,谢丕青,王佳濛,杨海峰. 柠条锦鸡儿种子产量预测模型研究. 山东农业大学学报(自然科学版). 2024(04): 557-565 . 百度学术
3. 郭玉东,张秋良,陈晓燕,张榕,宝朝鲁门,阿日宾巴雅尔,斯庆毕力格,王颖. 库布齐沙漠地区人工灌木林生物量模型构建. 西北农林科技大学学报(自然科学版). 2022(04): 74-82 . 百度学术
4. 段菁,云海英,李鑫,滕飞,纳青. 基于CASA模型的内蒙古地区草地生产力估测. 林业调查规划. 2022(03): 133-138+155 . 百度学术
5. 朱雅丽,张景路,张绘芳,地力夏提·包尔汉,廉佳佳. 天山中部锦鸡儿生物量分析及模型构建. 林业资源管理. 2022(05): 129-135 . 百度学术
6. 刘江,袁勤,张立欣,代香荣,刘向阳,丁茹,叶丽娜. 库布齐沙漠北缘不同人工灌木林地土壤肥力质量状况. 西北林学院学报. 2021(02): 46-53 . 百度学术
7. 黄小娟,侯扶江. 高寒典型草原主要物种的株高和盖度预测种群和群落地上生物量. 生态学报. 2021(12): 4942-4952 . 百度学术
8. 张殿岱,王雪梅. 塔里木盆地北缘绿洲-荒漠过渡带典型植物地上生物量估测. 生态学杂志. 2019(10): 3211-3220 . 百度学术
其他类型引用(7)
















计量
- 文章访问数: 208
- HTML全文浏览量: 47
- PDF下载量: 67
- 被引次数: 15
