
Isolation and functional analysis of CYC2d orthologous genes from several plants of the tribe Anthemideae
-
摘要: CYC同源基因作为控制花瓣对称性形成的关键基因,在菊科头状花序中主要调控着舌状花(两侧对称)的生长发育。菊科植物中舌状花的有或无及其分子调控机制和演化过程一直备受关注。本研究从花型不同的川甘亚菊、戈壁短舌菊和神农香菊中分别同源克隆了转录因子基因CYC2d。3个序列与菊花CmCYC2d基因氨基酸序列的比对结果显示其同源性均超过90%,且均含有保守的TCP和R结构域。半定量RT-qPCR结果显示,CYC2d在地被菊品种‘毛香玉’幼嫩花序中的表达量最高,而在川甘亚菊和戈壁短舌菊中的表达量非常微弱。因此,进一步通过实时荧光定量检测了CmCYC2d在‘毛香玉’6个发育时期舌状花和管状花中的表达情况,结果表明, CmCYC2d各个时期管状花中的表达量均很低,而在相应时期舌状花中的表达均很丰富。在不同花型的杂交F1代优株中,CmCYC2d也主要在不同位置的舌状花中高表达。通过农杆菌转化重组质粒pSUPER1300-CmCYC2d-GFP在烟草表皮细胞瞬时表达,亚细胞定位结果显示其定位于表皮细胞核。分别在野生型拟南芥和tcp1突变体(SALK-022364)中过表达CmCYC2d基因的结果表明,转基因阳性株系的营养生长受到抑制,花期延迟,花瓣大小和排列均发生了变化,使原本辐射对称的花瓣呈现两侧对称的趋势。从研究结果可知, 转录因子基因CmCYC2d对菊花舌状花的发育有重要调控作用。本研究为菊科舌状花演化的分子调控机制研究奠定了基础。Abstract: As the important gene of regulating flower symmetry, cyc-like proteins have been shown to mainly regulate the identity and development of ray floret (bilaterally symmetrical) in Asteraceae. The presence or absence of ray floret in Asteraceae and its molecular regulation mechanism as well as the evolutionary process have been highly concerned. Orthologous genes of CYC2d from Ajania potaninii, Brachanthemum titovii and Chrysanthemum indicum var. aromaticum were obtained by homology-based cloning. Their sequence alignment and conserved motif analysis were performed with the amino sequence of CmCYC2d, respectively. The results showed that their homology was more than 90% and all these proteins contained the conserved TCP and R domains. Furthermore, according to the result of semi-quantitative RT-PCR assay, CYC2d was strongly expressed in the young inflorescence of the groundcover chrysanthemum 'Mao xiangyu', while slightly expressed in that of A.potaninii and B.titovii. Therefore, the transcription levels of CmCYC2d were examined in ray and disc florets of 'Mao xiangyu' at six developing stages using quantitative real-time PCR. The results indicated that it was weakly expressed in disc florets of all stages, while highly expressed in ray florets of the corresponding stages. Moreover, in three F1 progenies with various whorls of ray florets, the CmCYC2d was expressed at much higher levels in ray florets of different whorls than in disc florets. The recombinant plasmid pSUPER1300-CmCYC2d-GFP was transiently expressed into the epidermal cells of Nicotiana benthamiana by agrobacterium-mediated transformation, and subcellular localization analysis revealed that the CmCYC2d protein mainly localized into the nucleus of epidermal cells. Furthermore, CmCYC2d was overexpressed in wild type Arabidopsis and the TCP1 mutant used the floral-dip method. The results showed that the vegetative growth and the flowering time of the positive transgenic lines were repressed and postponed. Moreover, the size and arrangement of the petals seemed to be changed, making the petal arrangement showed bilateral symmetry from original radial symmetry. These results indicate that the transcription factor CmCYC2d is essential in regulating ray floret identity in chrysanthemum. Our study lays a foundation for the research of molecular mechanisms for the evolutionary process of ray floret in Asteraceae.
-
Keywords:
- flower symmetry /
- Chrysanthemum /
- Anthemideae /
- CmCYC2d /
- ray floret /
- gene expression
-
噪声会增加劳动者的生理和心理负荷,引起工作能力下降,甚至能够导致职业听力损失和心率变异[1-3]。2012年我国实施的GBZ/T229.4—2012《工作场所职业病危害作业分级第4部分:噪声》[4]规定了噪声危害级别,目的是保护劳动者不受过度噪声的干扰或伤害。园林工人用来割除杂草、藤蔓的割灌机在作业时会产生较大的噪声。叶仲基等[5]对台湾割灌作业中使用打草绳和2齿刀片产生的噪声进行了数据调查;莫秋云等[6-7]基于人体脑波和心率变异对噪声综合评价方法进行了研究;Hinchcliffe[8]研究了噪声对听力、心率以及疲劳的影响;GB/T 5390—2013规定了以内燃机为动力的便携式手持操作林业和园林机械A计权声功率级和耳旁噪声的测定方法[9]。但是,园林工人在实际操作割灌机时,会根据不同的现场地形和切割对象变化来选择适当的刀具进行作业,由于机器老化、地形差异、切割刀具和对象的变化,割灌机作业产生的噪声与企业检测结果会有明显差异。在园林作业现场,对园林工人每个工作日操作割灌机作业的噪声暴露剂量和噪声频谱进行实际调查和记录,分析噪声对园林工人脑电波的影响,对于提高园林工人的职业安全,减少职业病的发生和提升割灌机具的设计生产水平具有现实意义。
1. 材料与方法
1.1 割灌作业现场噪声检测实验
1.1.1 实验对象和设备
选取18名年龄在40~50岁的男性园林工人, 平均身高174 cm,平均体重65.2 kg,受试者均能够熟练操作割灌机,身体健康无疾病,实验前一天无反常情绪影响,休息充分。
背负式割灌机选用国内使用率较高的BG140型四冲程背负式割灌机,净质量9.5 kg,排量37.7 mL,工作杆长度1 330 mm;侧挂式割灌机选用国内使用率较高的CG140型四冲程侧挂式割灌机,排量37.7 mL,净质量8.5 kg,工作杆长度1 330 mm,燃油为93#汽油,刀具选用圆形尼龙打草绳(直径2.4 mm、长度150 mm)、2齿锰钢菱形刀片(长305 mm、宽90 mm、厚2 mm)、4齿锰钢刀片(直径255 mm、厚2 mm)、40齿锰钢刀片(直径255 mm、厚2 mm)。割灌机均已正常使用1年以上,非出厂新机。噪声声级计选用台湾泰仕牌TES-1355型噪音计声级计。音频分析仪选用恒升HS5671A型噪声频谱分析仪,符合IEC1260和GB/T3241—1998对倍频程滤波器和1/3倍频程滤波器的要求。
1.1.2 实验方法
在园林作业现场,采用对园林工人实际测量的方式进行实验检测,并进行噪声采集。实验于2015年6月在哈尔滨某生态公园进行,分为噪声声级检测和噪声频谱检测两部分,并按照割灌机种类、刀具型号进行分类。根据调查,园林工人在作业中针对现场地形和切割对象的变化,常用刀具的对应情况如表 1所示。
表 1 园林工人常用刀具与作业环境对照Table 1. Comparison of common tools and operating environment for garden workers打草绳
Trimmer line2齿刀片
Two-tooth cutting tool4齿刀片
Four-tooth cutting tool40齿刀片
Forty-tooth cutting tool地形
Terrain切割对象
Cutting object地形
Terrain切割对象
Cutting object地形
Terrain切割对象
Cutting object地形
Terrain切割对象
Cutting object背负式割灌机
Knapsack brush cutter起伏地面
Undulating ground高度≤300 mm低软嫩草
Low soft tender grass with height≤300 mm起伏地面
Undulating ground高度≤300 mm
多种类杂草Variety of weeds with height≤300 mm起伏地面
Undulating ground高度≥300 mm多种类高韧杂草
Variety of high tough weeds with height≥300 mm起伏地面
Undulating ground直径15~50 mm较坚硬灌木
Harder shrub with ϕ15-50 mm侧挂式割灌机
Shoulder-hanging brush cutter平整草坪
Smooth lawn高度≤300 mm低软嫩草
Low soft tender grass with height≤300 mm平整地面
Flat ground高度≤300 mm多种类杂草
Variety of weeds with height≤300 mm平整地面
Flat ground高度≥300 mm多种类高韧杂草
Variety of high tough weeds with height≥300 mm平整地面
Flat ground直径15~50 mm较坚硬灌木
Harder shrub with ϕ15-50 mm1) 噪声声级检测。将噪声声级计固定于割灌机作业人员腰间,再将麦克风固定于人员颈后(侧挂式割灌机为右侧)衣领处。门槛声压位准采用80 dB。噪声声级检测实验的目的是测量作业者整个工作日的耳旁噪声暴露声级和暴露剂量。在实验过程中,对作业时间、地点、检测时间、割灌机型式、割灌机型号、刀具型式、环境温湿度、草的长度、背景噪声、噪声声级计型号等项目内容进行记录,检测完毕后,对声级计检测的噪声剂量与8小时加权平均分贝数值进行记录。
2) 噪声频谱检测。根据GB10000—1988中国成年人人体尺寸国家标准[10],人耳距离地面高度约为1 500 mm,将音频分析仪固定于距离地面1 500 mm的支架上,实验测量时间为5 min,分析频率范围在100~6 000 hz,采用1/3倍频。噪声频谱检测实验的目的是调查割灌机作业时噪声的主要集中频率与频率的分布。对割灌机怠速空转(1 500~2 000 r/min)、低速作业(2 000~4 000 r/min)和高速作业(4 000~6 500 r/min)的噪声频谱分别进行检测记录,然后对结果进行对比分析。
1.2 人体脑电测试
1.2.1 实验对象和设备
选取18名年龄在40~50岁的男性园林工人, 平均身高174 cm,平均体重65.2 kg,受试者均能够熟练操作割灌机,身体健康无疾病,实验前一天无反常情绪影响,休息充分。。
脑电测试仪采用荷兰ANT Neuro公司生产的64通道无线脑电肌电系统(Ultra-Mobile EEG & EMG Recording Platform)。噪声声级计、音频分析仪同上文。
1.2.2 实验方法
脑电信号与人的精神状态有密切联系,且准确度高、客观性强,在疲劳检测领域有广泛应用。Berger把脑波分成α波、β波、θ波、δ波[11],在β波时人的注意力集中或精神紧张,脑电中的α波受到抑制或消失,部分人可出现θ波,而在精神放松时α波明显增强,但精神放松到一定阶段而出现倦意时,α波则渐渐解体,同时θ波活动增强。近年,国外研究资料[12-13]也表明:脑电θ波的变化与人的心理活动和情感变化密切相关。随着睡眠的加深,α波随之逐步消失,而且逐步被高振幅的慢波所代替,此时,在脑电图中以θ波或δ波为主。如果人处于昏迷状态时,脑电图将全部出现δ波。
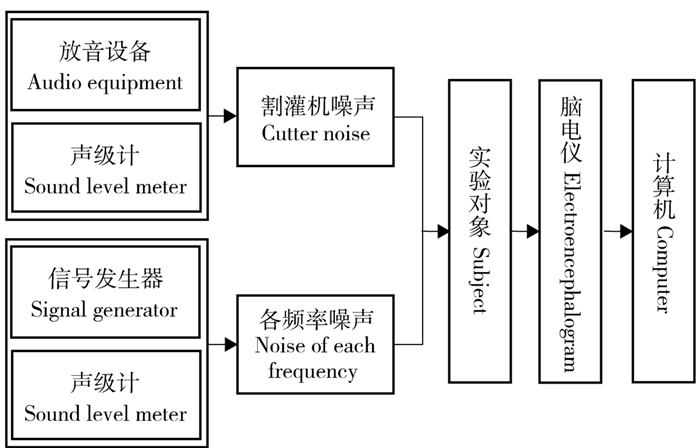
本研究选取人体脑电作为噪声对人体生理及心理负荷影响的评价指标。将1.1割灌作业现场噪声检测实验中采集的噪声录音文件进行回放,为了充分考虑实际噪声和单频噪声对人体影响, 以便于对不同单频噪声的对比分析, 选择了两类噪声进行实验:1)现场噪声,工人使用割灌机不同刀具在低速和高速作业切割时的噪声,取平均值90 dB;2)不同频率下的单频噪声,频率与声级如表 2所示。实验时间为5 min,为了保证实验的可对比性,将实验时间分为初始安静阶段(60 s)、噪声暴露阶段1(90 s)、噪声暴露阶段2(90 s)和噪声消除阶段(60 s),之后根据脑电的频谱变化更准确地分析噪声对疲劳程度的影响。实验地点为东北林业大学人机工程实验室。实验系统框图如图 1所示。实验对象保持坐姿,对头皮酒精消毒后,涂抹脑电传导胶,佩戴脑电帽,如图 2所示。闭目,与噪声源距离0.2 m,开始实验,实验过程中实验对象只受有无噪声变化的影响,其他环境因素不变。
表 2 不同频率下的单频噪声对照Table 2. Single frequency noise control at different frequencies频率
Frequency/Hz800 1 200 2 000 3 150 4 000 声级Noise level/dB 80 80 80 80 80 85 85 85 85 85 90 90 90 90 90 2. 结果与分析
2.1 噪声声级
对18名实验对象进行5个工作日的噪声监测调查,以每人每工作日的测量数据为一组,共计90组数据,其中背负式割灌机数据51组,侧挂式割灌机数据39组。对每组数据进行统计计算,结果如表 3所示。
表 3 割灌机噪声检测实验结果Table 3. Experimental results of noise level detection in brush cutters打草绳
Trimmer line2齿刀片
Two-tooth cutting tool4齿刀片
Four-tooth cutting tool40齿刀片
Forty-tooth cutting tool噪声剂量
Noise dose/%8小时加权平均分贝
8-hour time-weighted average decibel/dB噪声剂量
Noise dose/%8小时加权平均分贝
8-hour time-weighted average decibel/dB噪声剂量
Noise dose/%8小时加权平均分贝
8-hour time-weighted average decibel/dB噪声剂量
Noise dose/%8小时加权平均分贝
8-hour time-weighted average decibel/dB背负式割灌机Knapsack brush cutter 44.90 84.40 69.55 88.28 75.11 90.40 77.76 92.37 侧挂式割灌机Shoulder-hanging brush cutter 41.66 82.13 50.52 85.03 64.80 86.09 70.00 90.75 由表 3可见,背负式割灌机和侧挂式割灌机在使用不同刀具进行作业时产生的噪声剂量不同。背负式割灌机在使用打草绳、2齿刀片、4齿刀片和40齿刀片作业时,其8小时加权平均分贝分别为84.40<88.28<90.4<92.37 dB;侧挂式割灌机分别为82.13<85.03<86.09<90.75 dB。在使用相同刀具时,背负式割灌机作业人员的8小时加权平均分贝值都高于使用侧挂式割灌机的。
在噪声声级检测实验得出的90组数据中,共有54组数据的耳旁噪声8小时加权平均分贝超过85 dB,达到GBZ/T229.4—2012《工作场所职业病危害作业分级第4部分:噪声》[4]规定的轻度噪声危害级别;共有32组数据超过90 dB,达到中度噪声危害级别。
2.2 噪声频谱
对51组背负式割灌机和39组侧挂式割灌机使用不同刀具在怠速空转、低速作业和高速作业状态中的声级和频率分别进行检测记录,对每组数据进行统计计算,结果如表 4所示。
表 4 割灌机噪声频谱检测实验结果Table 4. Experimental results of noise spectrum detection for brush cutter打草绳
Trimmer line2齿刀片
Two-tooth cutting tool4齿刀片
Four-tooth cutting tool40齿刀片
Forty-tooth cutting tool声级
Noise level/dB频率
Frequency/Hz声级
Noise level/dB频率
Frequency/Hz声级
Noise level/dB频率
Frequency/Hz声级
Noise level/dB频率
Frequency/Hz背负式割灌机
Knapsack brush cutter怠速
Idle speed74.58 800 75.73 630~2 000 75.83 630~2 000 76.96 630~2 000 低速
Low speed80.43 800 81.52 800~2 000 84.2 800~2 000 85.81 2 000 高速
High speed92.68 800~4 000 88.05 500~2 000 90.77 500~2 000 91.93 500~2 000 侧挂式割灌机
Shoulder-hanging brush cutter怠速
Idle speed77.32 500~2 000 77.51 500~800 78.33 2 000 77.63 2 000 低速
Low speed84.72 800~3 150 84.16 800 86.41 2 000 89.31 2 000 高速
High speed92.66 2 500~4 000 94.83 2 500~4 000 90.76 4 000 93.01 4 000 由噪声频谱检测实验结果可见:在怠速空转时,背负式割灌机产生的噪声频谱数据集中在630~2 000 hz,声级数据集中在74.58~76.96 dB;侧挂式割灌机产生的噪声频谱数据集中在500~2 000 hz,声级数据集中在77.32~78.33 dB。检测数据符合GBZT229.4—2012规定,但是作业人员在割草时不可能使用怠速操作。
在低速作业时,背负式割灌机产生的噪声频谱数据集中在800~2 000 hz,声级数据集中在80.43~85.81 dB;侧挂式割灌机产生的噪声频谱数据集中在800~3 150 hz,声级数据集中在84.16~89.31 dB。与怠速空转时相比,噪声频谱数据和声级数据都有所增加,有多组声级数据超过了GBZT229.4—2012规定的轻度危害值85 dB,特别是在使用40齿刀片对较坚硬的灌木进行修剪时,声级数据更是达到了89.31 dB,接近了中度危害的临界值90 dB。
在高速作业时,背负式割灌机产生的噪声频谱数据集中在500~4 000 hz,声级数据集中在88.05~92.68 dB;侧挂式割灌机产生的噪声频谱数据集中在2 500~4 000 hz,声级数据位于90.76~94.83 dB。与怠速空转和低速作业相比较,高速作业时的噪声频谱数据和声级数据都有所增加,侧挂式割灌机的声级数据最大值达到了94.83 dB,已接近重度危害的临界值95 dB。
根据GBZT229.4—2012规定的分级管理原则,在目前的作业条件下,很可能对劳动者的听力产生不良影响,必须采取噪声控制措施,减少作业者噪声暴露时间,作业时佩戴有效的耳塞、耳罩等个人防护用品,以避免噪声危害。
2.3 割灌机作业噪声对园林工人脑波的影响
图 3截取了一例实验对象不同状态下在电极O1处的脑电数据样本,每个样本时间为6 s,共3 000个数据点。
对图 3中的脑电进行快速傅立叶变换(FFT), 得到其相应的频谱图(图 4)。
有研究证明:脑电α波和β波频带的平均功率谱密度比值R可作为衡量作业疲劳度的脑电特征量[14]。设信号在频带h的平均功率谱密度G(h)为:
G(h)=∫fufdp(f)dffu−fd (1) 式中:fu为频带h的上限,Hz;fd为h频带的下限,Hz;p(f)为信号的功率谱密度,W/Hz。令:
R=G(h1)G(h2) (2) 式中:h1为α波频带;h2为β波频带。
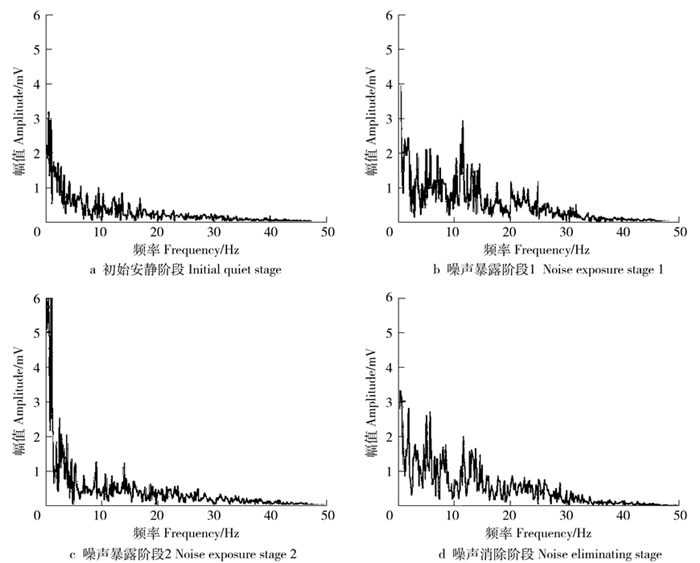
表 5为18个实验对象在初始安静阶段、噪声暴露阶段1、噪声暴露阶段2和噪声消除阶段时O1电极处的脑电α波频带(10~13 hz)和β波频带(18~22 hz)的R值统计数据。
表 5 18名实验对象的脑电R值Table 5. Electroencephalogram R value of 18 test subjects初始安静阶段
Initial quiet stage噪声暴露阶段1
Noise exposure phase 1噪声暴露阶段2
Noise exposure phase 2噪声消除阶段
Noise eliminating phase平均值Average 1.27 3.69 3.15 1.73 标准差Standard deviation 0.09 0.50 0.30 0.15 在噪声暴露阶段1,脑电α波和β波的R平均值为3.69,对比初始安静阶段,R值有明显提高;在噪声暴露阶段2,R平均值为3.15,相比噪声暴露阶段1,共有9例实验对象的R值有所回落,其原因可能是生理对噪声有所适应,情绪紧张度下降;在噪声消除阶段,实验对象的R值迅速下降,说明脑电对噪声反应敏感,减少噪声即可降低噪声对脑波反映出的心理负荷影响。
3. 结论与讨论
本研究在园林作业现场,对园林工人每个工作日操作割灌机作业的噪声暴露剂量和噪声频谱进行了实际测量和记录;并根据已获得的割灌机作业现场噪声录音文件,对18名实验对象进行了脑电测量实验,分析了噪声对园林工人脑电的影响,强调了被忽略的噪声影响劳动安全的心理负荷问题。具体结论如下:
1) 在园林工人操作割灌机进行绿化作业时,有半数以上操作者的耳旁噪声暴露剂量处于超标状态,达到了轻度或中度噪声危害等级。割灌机低速作业产生的噪声已达轻度危害等级,高速作业产生的噪声频率已达到中度危害等级。
2) 园林绿化部门必须加大对割灌机的设计研究投入,减少割灌机本身的振动和噪声。根据噪声特性,选择适宜的防护用具,阻止噪声传播,减少噪声对作业者产生的影响。
3) 由人体脑电实验得出的数据样本可知:割灌机作业产生的噪声对人体脑波变化的影响差异性显著,脑电特征量或可作为衡量噪声负荷与作业疲劳度的影响因子。
4) 脑电信号具有易干扰、复杂性的特点,本实验是以模拟噪声现场环境数据为基础的,是否还有预测效果更好的脑电特征量指标,尚需进一步的对比分析,且需要在真实工况中进一步验证结论的有效性。
-
![]()
图 1 地被菊‘毛香玉’花序的6个生长阶段
Figure 1. Six inflorescence developing stages of the groundcover chrysanthemum 'Mao xiangyu'
![]()
图 2 六倍体菊杂交F1代3个优株F1-1、F1-2和F1-3
Figure 2. Three F1 progenies of two hexaploid chrysanthemum: F1-1, F1-2 and F1-3
![]()
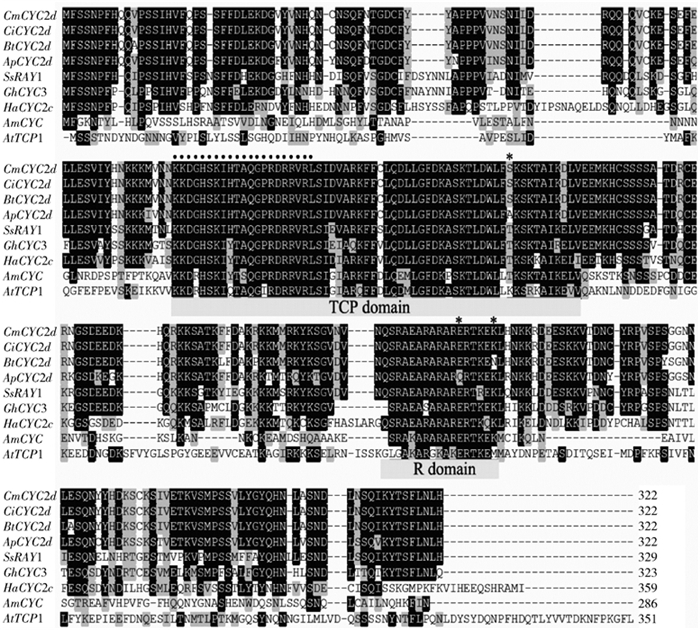
图 3 CYC2d同源基因氨基酸序列比对
*为川甘亚菊和戈壁短舌菊与菊花CYC2d基因在TCP和R结构域有差异的位点;黑圆点表示bHLH的核定位信号区。
Figure 3. Alignment of amino acid sequence of CYC2d homologues
* indicates the different sites in TCP and R domain of CYC2d proteins among Ajania potaninii, Brachanthemum titovii and Chrysanthemum; The black circle indicates the nuclear localization signal site.
![]()
图 4 半定量RT-PCR检测CYC2d在4种材料花芽中的表达
MXY为‘毛香玉’,Ci为神农香菊,Ap为川甘亚菊,Bt为戈壁短舌菊。
Figure 4. Expression of CYC2d in floral buds of four plants detected by semi-quantitative RT-PCR
MXY, 'Mao xiangyu'; Ci, C. indicum var. aromaticum; Ap, A. potaninii; Bt, B. titovii.
![]()
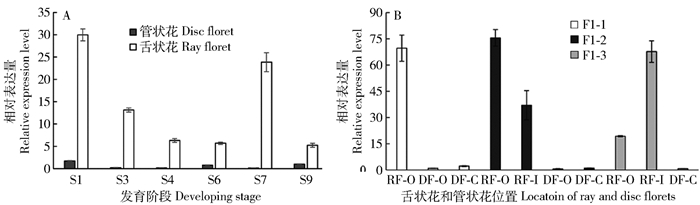
图 5 CmCYC2d在‘毛香玉’舌状花和管状花中的时空表达模式
A:小花不同发育时期CmCYC2d基因的表达分析。B: CmCYC2d在3个F1优株不同位置小花中的表达分析,其中:RF-O为外轮舌状花;RF-I为内轮舌状花;DF-O为外轮管状花;DF-C为中心管状花。
Figure 5. Temporal and spatial expression patterns of CmCYC2d in ray and disc florets of 'Mao xiangyu'
A: expression analysis of CmCYC2d during different stages of floret development. B: expression analysis of CmCYC2d at different floret locations of three F1 progenies. Tissues are outer ray florets (RF-O), inner ray florets (RF-I), outer disc florets (DF-O) and central disc florets (DF-C).
![]()
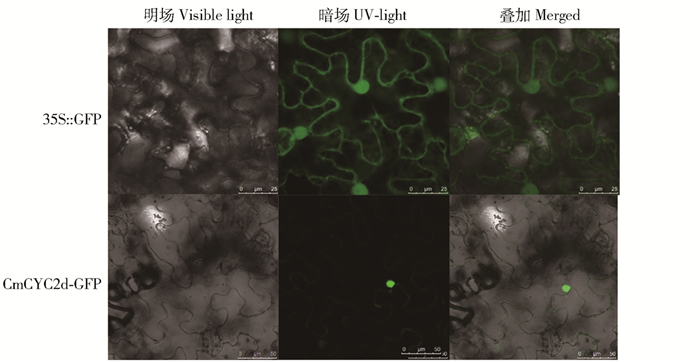
图 6 CmCYC2d蛋白在烟草叶片表皮细胞中的亚细胞定位分析
Figure 6. Subcellular localization of CmCYC2d in tobacco epidermal cells
![]()
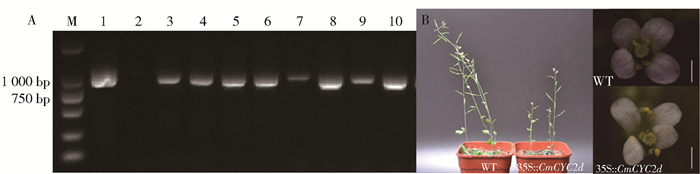
图 7 转基因拟南芥的PCR鉴定(A)及表型分析(B)
A: M,DNA Marker; 1和2分别为质粒和野生型对照; 3~10为转基因株系;B: CmCYC2d超表达影响了植株的营养、生殖生长及花瓣排列方式。标尺:1 mm。
Figure 7. PCR identification (A) and phenotype analysis (B) of CmCYC2d overexpressing Arabidopsis
A: M, DNA marker; 1 and 2 for plasmid and wild-type controls; 3-10 for transgenic lines. B: ectopic expression of CmCYC2d in Arabidopsis affecting vegetative and reproductive growth, as well as the arrangement of petals. Scale bars: 1 mm.
![]()
图 8 转基因拟南芥tcp1突变体的PCR鉴定及表型分析
A:M,DNA Marker 2000; 1为转空载对照; 2~11为CmCYC2d基因超表达tcp1突变体株系。B:CmCYC2d超表达抑制了根系的生长。C:CmCYC2d超表达推迟了花期。D和E:CmCYC2d超表达和空载株系的花。
Figure 8. PCR identification and phenotype analysis of CmCYC2d overexpressing Arabidopsis of tcp1 mutant
A: M, DNA marker; 1 for transgenic plants with empty vector; 2-11 for CmCYC2d overexpression tcp1 mutant lines. B: ectopic expression of CmCYC2d affecting growth of roots in positive transgenic lines; C: ectopic expression of CmCYC2d affecting flowering. D, E: flower types of the CmCYC2d overexpression lines and empty lines.
表 1 本研究所用引物序列
Table 1 Primer sequences used in this study
引物名称
Primer name引物序列(5′-3′)
Primer sequence (5′-3′)用途Use CYC2d -F1 ATGTTTTCCTCGAACCCTTTTCAT 同源克隆及半定量
RT-PCRHomology-based cloning and semi-quantitative RT-PCRCYC2d-R1 CTAGTGTAAATTTAGGAAACTTGTGTAC CnActin-F1 CACCTCTAAATCCTAAGGCTAACAG 半定量
RT-PCRSemi-quantitative RT-PCRCnActin-R1 GAACAATGGATGGGCCAGACTC CnActin-F2 CTGACAGGATGAGCAAGGAAATCAC 荧光定量
PCRQuantitative RT-PCRCnActin-R2 GAACAATGGATGGGCCAGACTC CmCYC2d-F1 TCCTCGAACCCTTTTCATCAACAG CmCYC2d-R1 GCTGCCTGTCCAAAATATTGCTGT CmCYC2d-F2 CCAAATCGACTCTAGAATGTTTTCCTCGAACCCTTTTC 亚细胞定位载体构建
Construction of pSUPER1300-CmCYC2d-GFPCmCYC2d-R2 TACCGGATCCACTAGTGTAGTGTAAATTTAGGAAACTTGTGTAC CmCYC2d-F3 GGACTCTTGACCATGGCTATGTTTTCCTCGAACCCTTTTC 转基因载体构建
Construction of pCAMBIA1304-CmCYC2dCmCYC2d-R3 CTTCTCCTTTACTAGTGTAGTGTAAATTTAGGAAACTTGTGTAC p1304-F1 ACACGGGGGACTCTTGAC 转基因植株鉴定
PCR identification of transgenic insertionp1304-R1 CAACAAGAATTGGGACAACTC p1300-F1 TCATAACCAATCTCGATACACCA p1300-R1 CTGAACTTGTGGCCGTTTACG 注:下划线处为酶切位点。Note:underlines are enzyme recognition sites.  下载: 导出CSV
下载: 导出CSV
-
[1] GUSTAFSSON Å. Linnaeus' Peloria: the history of a monster[J]. Theoretical and Applied Genetics, 1979, 54(6): 241-248. doi: 10.1007/BF00281206
[2] LUO D, CARPENTER R, COPSEY L, et al. Control of organ asymmetry in flowers of Antirrhinum[J]. Cell, 1999, 99(1): 367-376. doi: 10.1016-S0092-8674(00)81523-8/
[3] LUO D, CARPENTER R, VINCENT C, et al. Origin of floral asymmetry in Antirrhinum[J]. Nature, 1996, 383: 794-799. doi: 10.1038/383794a0
[4] CUBAS P, VINCENT C, COEN E. An epigenetic mutation responsible for natural variation in floral symmetry[J]. Nature, 1999, 401: 157-161. doi: 10.1038/43657
[5] CUBAS P, LAUTER N, DOEBLEY J, et al. The TCP domain: a motif found in proteins regulating plant growth and development[J]. The Plant Journal, 1999, 18(2): 215-222. doi: 10.1046/j.1365-313X.1999.00444.x
[6] GAO Q, TAO J H, YAN D, et al. Expression differentiation of CYC-like floral symmetry genes correlated with their protein sequence divergence in Chirita heterotricha (Gesneriaceae)[J]. Development Genes and Evolution, 2008, 218(7): 341-351. doi: 10.1007/s00427-008-0227-y
[7] YANG X, PANG H B, LIU B L, et al. Evolution of double positive autoregulatory feedback loops in CYCLOIDEA2 clade genes is associated with the origin of floral zygomorphy[J]. The Plant Cell, 2012, 24(5): 1834-1847. doi: 10.1105/tpc.112.099457
[8] FENG X, ZHAO Z, TIAN Z, et al. Control of petal shape and floral zygomorphy in Lotus japonicus[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(13): 4970-4975. doi: 10.1073/pnas.0600681103
[9] REARDON W, GALLAGHER P, NOLAN K M, et al. Different outcomes for the MYB floral symmetry genes DIVARICATA and RADIALIS during the evolution of derived actinomorphy in Plantago[J]. The New Phytologist, 2014, 202(2): 716-725. doi: 10.1111/nph.12682
[10] KIM M, CUI M L, CUBAS P, et al. Regulatory genes control a key morphological and ecological trait transferred between species[J]. Science, 2008, 322: 1116-1119. doi: 10.1126/science.1164371
[11] CHAPMAN M A, LEEBENS-MACK J H, BURKE J M. Positive selection and expression divergence following gene duplication in the sunflower CYCLOIDEA gene family[J]. Molecular Biology and Evolution, 2008, 25(7): 1260-1273. doi: 10.1093/molbev/msn001
[12] BROHOLM S K, TAHTIHARJU S, LAITINEN R A, et al. A TCP domain transcription factor controls flower type specification along the radial axis of the Gerbera (Asteraceae) inflorescence[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(26): 9117-9122. doi: 10.1073/pnas.0801359105
[13] CHAPMAN M A, TANG S, DRAEGER D, et al. Genetic analysis of floral symmetry in Van Gogh's sunflowers reveals independent recruitment of CYCLOIDEA genes in the Asteraceae[J/OL]. PLoS Genet, 2012, 8(3): e1002628[2016-09-16]. http://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1002628.
[14] FAMBRINI M, SALVINI M, BASILE A, et al. Transposon-dependent induction of Vincent van Gogh's sunflowers: exceptions revealed[J]. Genesis, 2014, 52(4): 315-327. doi: 10.1002/dvg.22743
[15] JUNTHEIKKI-PALOVAARA I, TÄHTIHARJU S, LAN T, et al. Functional diversification of duplicated CYC2 clade genes in regulation of inflorescence development in Gerbera hybrida (Asteraceae)[J]. The Plant Journal, 2014, 79(5): 783-796. doi: 10.1111/tpj.12583
[16] GARC S H M P, SPENCER V M R, KIM M. Control of floret symmetry by RAY3, SvDIV1B and SvRAD in the capitulum of Senecio vulgaris [J/OL]. Plant Physiology, 2016, 10[2016-09-16]. http://www.plantphysiol.org/content/early/2016/05/12/pp.16.00395.abstract.
[17] HELARIUTTA Y, ELOMAA P, KOTILAINEN M, et al. Cloning of cDNA coding for dihydroflavonol-4-reductase (DFR) and characterization of dfr expression in the corollas of Gerbera hybrida var. regina (Compositae)[J]. Plant Molecular Biology, 1993, 22(1): 183-193.
[18] MANASSERO N G, VIOLA I L, WELCHEN E, et al. TCP transcription factors: architectures of plant form[J]. Biomolecular Concepts, 2013, 4(2): 111-127. https://www.ncbi.nlm.nih.gov/pubmed/25436570
[19] TAHTIHARJU S, RIJPKEMA A S, VETTERLI A, et al. Evolution and diversification of the CYC/TB1 gene family in Asteraceae: a comparative study in Gerbera (Mutisieae) and sunflower (Heliantheae)[J]. Molecular Biology and Evolution, 2012, 29(4): 1155-1166. doi: 10.1093/molbev/msr283
[20] HUANG D, LI X, SUN M, et al. Identification and characterization of CYC-like genes in regulation of ray floret development in Chrysanthemum morifolium [J]. Frontiers in Plant Science, 2016, 7[2016-09-16]. http://journal.frontiersin.org/article/10.3389/fpls.2016.01633.
[21] CITERNE H L, PENNINGTON R T, CRONK Q C. An apparent reversal in floral symmetry in the legume Cadia is a homeotic transformation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(32): 12017-12020. doi: 10.1073/pnas.0600986103
[22] TAKEDA T, SUWA Y, SUZUKI M, et al. The OsTB1 gene negatively regulates lateral branching in rice[J]. The Plant Journal, 2003, 33(3): 513-520. doi: 10.1046/j.1365-313X.2003.01648.x
[23] AGUILAR-MARTNEZ J A, POZA-CARRI N C, CUBAS P. Arabidopsis BRANCHED1 acts as an integrator of branching signals within axillary buds[J]. The Plant cell, 2007, 19(2): 458-472. doi: 10.1105/tpc.106.048934
[24] COSTA M M, FOX S, HANNA A I, et al. Evolution of regulatory interactions controlling floral asymmetry[J]. Development, 2005, 132(22): 5093-5101. doi: 10.1242/dev.02085
[25] HILEMAN L C. Bilateral flower symmetry: how, when and why?[J]. Current Opinion in Plant Biology, 2014, 17(3): 146-152. http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ0232220132/
[26] ALMEIDA J, ROCHETA M, GALEGO L. Genetic control of flower shape in Antirrhinum majus [J]. Development, 1997, 124(7): 1387-1392. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=HighWire000005683084
-
期刊类型引用(8)
1. 鹿林,张程凯,张慧. 自橡子壳中提取橡子壳棕色素的实验研究. 中国食品添加剂. 2024(10): 179-186 .  百度学术
百度学术
2. 靳子旋,张岩岩,杜黔运,徐静,赵余庆. 橡子化学成分及药理作用的研究进展. 中草药. 2023(24): 8301-8308 . 百度学术
3. 翟淑红,曹洪坤,余诗琴,朱斯豪. 红菜苔多酚超声提取工艺优化及其抗氧化活性研究. 农产品加工. 2023(22): 49-52+56 . 百度学术
4. 张博,李德海,王泽童,王楚雅,王怡雪. 橡子壳主要成分的生理功能及开发利用研究. 食品工业科技. 2022(07): 393-399 . 百度学术
5. 王荣芳,张子言,李德海. 酶解法对蒙古栎实壳提取物活性成分及抗氧化活性的影响. 北京林业大学学报. 2022(05): 150-160 . 本站查看
6. 豆佳媛,何志鹏,梁馨月,逯莉. 橡子中挥发油的提取及抗氧化性质研究进展. 广东化工. 2021(24): 55-56 . 百度学术
7. 黄艳,傅新征,吴琳珊,李烨. 锥栗壳色素抗氧化活性研究. 食品科技. 2019(02): 274-280 . 百度学术
8. 魏园园,侯盼盼,梁宗瑶,任维维,李珉梦,高鹏程,张建新,段旭昌. 栓皮栎橡子壳多酚的体外抗氧化与抑菌活性研究. 现代食品科技. 2019(09): 190-197+73 . 百度学术
其他类型引用(8)






计量
- 文章访问数: 2492
- HTML全文浏览量: 531
- PDF下载量: 41
- 被引次数: 16
