
Forest dynamic succession simulation based on individual plant growth model
-
摘要:
森林动态演替过程的计算模拟历来是森林生态学中的一个重要问题。不同于传统宏观式的模拟思路,本文提出了一种基于个体植物生长模型、自下而上的模拟方法。首先构建了参数化的个体植物生长模型,模型中考虑了植物内部属性、外部环境、邻域竞争3种对植物生长有影响的因素,综合模拟出植物在不同环境状况下的生长变化情况。在此基础上,对构成一片森林的大量植物个体进行遍历式、迭代化的计算,从而模拟出整片森林区域的生长演替现象。实验结果表明,本文方法可以模拟出植被在复杂外部环境影响下的动态变化效果,可表现出自然稀疏、物种竞争、物种演替等多种典型的植被演替现象。
Abstract:The calculation and simulation of forest dynamic succession is always a significant issue of forest ecology. Different from traditional macroscope simulation way, in this paper, we proposed a bottom-up simulation approach based on individual plant growth model. The parameterized individual plant growth model was constructed firstly considering three influencing factors on plant growth including plant internal attribute, external environment and neighborhood competition, so as to simulate the growth changes in various environments of plant. Under such circumstance, a large amount of plant individuals in a forest would be calculated by the ergodic and iterative way to simulate the growth succession phenomenon in whole forest region. The results of experiment show that the approach in this paper could simulate the dynamic changing effect influenced by complex external environment of vegetation, including various typical rules of vegetation succession such as natural thinning, species competition and species succession.
-
Keywords:
- individual plant /
- health index /
- forest dynamic succession /
- visualized simulation
-
随着我国城市建设规模的不断扩大,城市下垫面不透水铺装比例不断升高,城市洪涝问题日益严重,同时初期雨水的污染物对城市水体的水质也造成了威胁[1-3]。随着海绵城市相关政策的颁布和试点城市的改造运行,通过“渗、滞、蓄、净、用、排”六字方针解决城市雨洪管理问题,以自然积存、自然渗透、自然净化为目标,科学有效地利用城市管网系统和自然系统排水,进行“海绵城市”建设,其理论体系得到了不断的发展[4-5]。目前,对于海绵城市建设的研究多集中在我国中东部气候温和地区的市政道路、公园绿地、住宅区[6-9],对于高海拔严寒地区的海绵化建设少有研究。随着西部城市经济发展,人口、城市面积不断扩张和全球气候变化异常,越来越多的西部高海拔城市面临着严重缺水和雨洪灾害的双重挑战[10]。本文以西宁市安泰华庭小区海绵化改造项目设计与实施为例,结合SWMM模型验证小区海绵化设计与建设的合理性,并与实际监测数据对比验证LID措施的有效性,为该地区海绵城市建设提供案例借鉴和理论支持。
1. 项目研究概况
1.1 区域概况及自然条件分析
西宁市是青海省会,青藏高原东门户。在大气环流和高海拔地形综合作用下,夏季温度适宜多雨,冬季寒冷干燥。平均海拔2 261 m,属高海拔严寒地区[11]。中心城区位于湟水河河谷地带,西高东低、成“两山对峙、三山汇聚、四川相连”的主城山水小格局,建成区地势相对较平缓,绝大部分地区的地面坡度在5°以内。西宁市作为我国海绵城市建设的第二批试点城市,该试点区占地面积21.61 km2,主要位于海湖新区行政范围(图1)。试点区域建立“治山、理水、润城”的系统性技术路线,打造“山涵水、水融山、城依水、水衬城”的生态宜居海绵城市[4]。在城市中建筑与小区、城市广场、城市道路、绿地等的雨水径流通过低影响开发雨水设施的“渗、滞、蓄、净”作用后,收集一部分雨水,经净化后直接用于各城市地块。超量雨水经湟水河湿地公园净化、储存利用后,排入湟水河[12]。
![]() 图 1 西宁市海绵城市建设试点区域范围及项目位置图Figure 1. Pilot area of sponge city construction in Xining City and the site
图 1 西宁市海绵城市建设试点区域范围及项目位置图Figure 1. Pilot area of sponge city construction in Xining City and the site在推进试点工作中,西宁市乡规划和建设局从试点区内选择了不同类型的小区、公共建筑、公园、城市道路等项目进行示范性工程建设,通过示范项目的设计与实施,为后期的大规模海绵城市建设提供范例和依据。安泰华庭小区作为已完工的新小区入选示范项目,以探索此类典型用地与建设条件下海绵城市建设的思路与方法。
根据西宁市1985—2015年近30年的降雨量统计数据,市区内降水量周期性变化较明显,夏季暴雨多发,暴雨特点是降雨历时短、范围小、强度大,尤以7—8月最多。西宁的年均降雨量为410 mm,逐月平均降水量最大出现在7—8月,最少是12月份。降雨主要集中在汛期(5—9月),占全年降水量的85%;而1—2月、11—12月降雨量仅占全年的3%,逐月平均降雨量见图2所示。西宁全年平均降雨天数(降雨量2 mm以上)占总天数的12.5%。最高日降雨量发生在2007年8月26日,日降雨量为57.8 mm;超过20 mm的日降雨量共86 d,占降雨总天数的6.1%,基本以中小雨为主。西宁降雨多以单一前锋雨型为主,降雨峰值比例(r)多为0.375左右。
试点区域的土壤主要以厚黑黏淤土、厚黄淤土及厚黑淤土为主。土壤成分主要为粉土,属稍湿的弱透水层。通过对试点区内具有代表性的点位进行土壤渗透系数测定,得出试点区域内土壤渗透系数均大于5 × 10− 6 m/s,符合低影响开发设施的建设标准。
1.2 安泰华庭小区概况
安泰华庭小区占地总面积为10.65 hm2,位于五四西路以南,是一个2011年已建成入住的新建小区。小区被两条“T”字形交叉的道路分为I、II、III 3个分区。其中I、III两区为住宅用地,II区为商业办公用地。住宅建筑均为高层板式建筑,采用沿用地周边布置的方式,四周有1 ~ 2层底商,形成封闭的内院围合模式,T字内街为商业步行街。小区的交通采取了人车分流的设计,沿住宅外围设有车行道,直接进入小区的地下车库,内院道路均为人行通道。I、III区的景观设计均呈现相似的模式,环形主路兼有消防通道的功能,联系各楼座单元,楼间绿地内设游步道和休闲场地,中心花园以水系景观为主,滨水设活动广场。小区实行封闭式管理,围合式的建筑布局使得小区内安全且安静,具有较高的生活品质。
2. 海绵化改造方案
2.1 海绵化改造的基础条件分析
2.1.1 竖向及地表排水组织
小区整体地势为南高北低、西高东低,小区外围道路标高最低点2 282.29 m,最高点2 286.62 m,有4 m多的高差。小区建筑采用了错层设计,地下车库面积5.6 hm2,基本覆盖了I、III区的所有空地,使得内院景观空间均为车库顶覆土绿化,内部场地坡度与外围道路形成约2.5 m高差,采用挡土墙解决。内院空间的绿地通过微地形组织雨水至路侧,道路及场地的雨水通过坡度设置也集中到路侧。沿路两侧采用加盖的排水明沟引导雨水,在适当位置向下输送到车库顶板的砾石疏水层,再通过盲沟管收集排向外围的雨水管网。这样的排水方式节省了一些排水管道,但是雨水排放效率低,仅适用于降雨较少的西北地区[13]。车库顶板覆土厚度为0.8 ~ 1.0 m,土层较薄,但是回填的素土经过夯实,也解决了黄土的湿陷性问题,对雨水的渗透利用有利有弊[14-15]。小区外围的车行道和内街的雨水通过道路横、纵坡的设置,经雨水口排入雨水管网。小区地表排水方向及高程详见图3,下垫面及雨水管网情况详见图4。
2.1.2 建筑屋面排水组织
在高寒地区,高层建筑的屋面雨水优先考虑使用建筑内排水,有利于管道保温,保护管网不受冻害的影响[16]。小区的屋面落水管大部分使用了建筑内排水,仅在个别屋面及内街的商业建筑采用了暴露雨落管的建筑外排水。建筑内排水的雨落管直接引至地下车库,水平管横穿车库的侧墙,从地下进入市政雨水管。该设计使得雨水无法通过自流的方式进入绿地消纳,造成雨水利用的困难,需要在后期的海绵城市设计时着力解决该雨水问题。
2.1.3 排水管网条件
小区采用雨污分流的排水系统,沿小区东西两侧车行道和内街上各有一条雨水主干管,收集建筑屋面雨水和路面雨水,排入市政排水管网。小区雨水共有3个主要总排口,2号排口排放II区商务办公区建筑产生的雨水,1、3号排口分别排放I区、III区住宅用地部分的雨水。管网设计标准为2年一遇,管径DN400 ~ 600 mm。
2.1.4 其他条件
小区绿地内有小型景观水体,日常运行采用市政自来水作为水源。未来小区进行雨水收集后,该水体可以作为雨水回用的受纳水体。
小区内部道路铺装(图5)使用陶瓷面砖饰面,光滑的表面在冬季冰雪天气容易摔伤行人,迫切需要更换。小区的车行道(图6)是混凝土道路,在管网改造过程中有一定面积路面破拆的工作,也需要更换路面铺装形式。
![]() 图 5 改造前的路面铺装及路侧排水沟Figure 5. Pavement and roadside drainage ditch before reconstruction
图 5 改造前的路面铺装及路侧排水沟Figure 5. Pavement and roadside drainage ditch before reconstruction2.2 海绵化改造方案设计
2.2.1 总体目标要求
在《西宁市海绵城市建设项目系统性详细规划(2016—2018)》中,结合场地改造前的现状条件包括场地水文地质、现状下垫面条件、建筑及道路广场排水、车库顶板覆土深度等条件,根据低影响开发总体力度控制及LID设施区域配置要求,确定了不同地块年径流总量控制分解指标。统筹考虑自身径流控制、污染削减率与周边地块、水体等水量、水质衔接关系,确定了如下海绵化改造目标:
(1) 小区年径流总量控制率为88.8%,对应的设计降雨量为15.2 mm。
(2) 年径流污染物(以SS计算)总量削减率不低于44%。
(3)通过LID建设,提高场地的排涝标准至50年一遇。
2.2.2 雨水管理流程和设施选择
根据小区的基础条件和特点,结合西宁地区降雨量少且时空分布不均的问题,在海绵化改造中,着重建设雨水的“渗、滞、蓄”,在削减径流量的同时去除污染。设计中选择的主要技术方法包括:
(1) 利用小区路侧排水明沟对径流雨水进行截留和分配,引导至绿地中进行消纳和渗透。
(2) 使用透水材料改造小区的人行道、车行道等,截留雨水的同时改善行人车辆的通行安全。
(3) 根据小区现状地形情况细分汇水分区,在绿地中设置下凹绿地和雨水花园,消纳外排的屋面雨水和道路径流雨水。
(4) 无法通过雨水管断接方式处理的建筑屋面内排水以及外围道路的路面径流,通过在小区雨水管网的末端设置地下调蓄池进行截留调蓄。
(5) 在底商等低矮屋面实验性地设置托盘容器式屋顶绿化,削减屋面径流。
(6) 设置雨水回用装置,将调蓄池收集的雨水回灌绿地,补充景观水体,塑造跌水景墙景观等。
2.2.3 方案设计
通过分析雨水管网的布置情况、竖向设计及建筑排水方式等因素,将小区分为6个一级汇水分区,又细分成共计51个子分区进行雨水控制(图9)。分别统计各汇水子分区的下垫面覆盖情况。
根据《海绵城市建设指南》,海绵城市设施以径流总量和径流污染为控制目标进行设计时,设施具有的调蓄容积一般应满足“单位面积控制容积”的指标要求。设计调蓄容积采用容积法,计算公式如下:
Vx=10HφF (1) 式中:Vx为调蓄容积(m3);H为设计降雨量,取88.8%年径流总量控制率,对应设计降雨量15.2 mm;
φ 为综合雨量径流系数;F为汇水面积(hm2)。利用公式(1),以子汇水分区为单元分别计算各个汇水分区的调蓄容积,得出调控的目标径流量值(表1)。
表 1 下垫面情况及目标径流量计算Table 1. Calculation of the target runoff catchment volume下垫面类型
Type of underlying surface面积
Area/m2雨水径流系数
Runoff coefficient径流总量控制率
Runoff control rate/%对应降雨量
Rainfall /mm目标径流量
Target runoff catchment volume/m3绿地 Green land 无地下建筑绿地及有地下建筑绿地(覆土厚度 ≥ 0.5 m)
No underground building green land and green land above the underground building (thickness of soil ≥ 0.5 m)27 212.96 0.15 88.8 15.2 62.05 路面铺装 Pavement 砖 Brick 43 235.24 0.6 88.8 15.2 394.31 混凝土 Concrete 6 397.30 0.9 88.8 15.2 87.52 透水铺装 Permeable pavement 1 883.97 0.35 88.8 15.2 10.02 屋面 Roof top 硬屋面、未铺石子的平屋面
Hard, unpaved flat roofs26 359.19 0.9 88.8 15.2 360.59 水系 Water 1 451.64 1 88.8 15.2 22.06 合计 Total 106 540.31 936.55 在各子汇水分区内,充分考虑竖向标高、径流汇集方式、植草沟、排水明沟位置以及管网布置情况、现有植物栽植位置等,择地安排下凹绿地、透水铺装、屋面绿化等LID设施在源头进行雨水的渗透和消减[17]。其规模面积、形状、调蓄水深、雨水的入口及溢流口等均按照实际建造的可能性安排,使得各子汇水分区能够就地消纳目标径流量的水量。考虑到实际建设以及后期管理可能存在的变化,在设施安排时都至少提高了10%的安全余量[18],详见表2。安泰华庭小区LID设施布局详见图10。
表 2 子汇水分区目标径流量与设计消纳径流量对比Table 2. Comparison of the target runoff and designed runoff volumem3 汇水分区编号
Catchment area No.目标径流量
Target runoff volume设计消纳径流量
Designed volumeA 145.29 353.17 B 160.09 459.29 C 243.90 568.94 D 23.70 49.50 E 117.25 130.00 F 246.32 270.00 合计 Total 936.55 1 830.9 在方案设计中,下凹绿地是主要的雨水渗透利用技术措施,用来消纳可收集的雨落管雨水、绿地及道路铺装径流雨水。改造设计充分利用了小区原有的路侧明沟排水体系,按照子汇水分区的划分位置将明沟进行分段截断,将明沟汇流的雨水在截断处破口转向,流至下凹绿地中进行消纳。超过下凹绿地储水能力的超量溢流雨水则回流到截断处下游的明沟中,使得雨水在路侧明沟与下凹绿地之间不断往复进出,实现雨水的渗透利用和长距离输送(图11)。
在实施方案中,使用高强度抛丸混凝土透水砖(图12)替换小区环路的陶瓷路面砖,既改善了雨雪天气下的通行安全,又可以利用二者的厚度差调高路面,使得道路标高高于周围绿地,形成绿地的相对下凹,保证路面雨水全部进入绿地形成雨水花园(图13)。
在西宁地区,由于在严寒地区薄土层种植植物越冬问题的限制,屋顶绿化技术鲜见[19]。在本设计中尝试选择屋顶绿化技术,目的在于利用屋顶的土壤层进行雨水拦蓄利用。方案中选择底商的非上人屋面进行了少量的屋面绿化设计,采用15 cm厚人工配置土和景天科植物的组合(费菜Sedum aizoon、八宝景天Hylotelephium erythrostictum、胭脂红景天Sedum spurium)。经过全年一个生长周期的观察,植物能够安全越冬,夏季生长良好(图14),证明以拦蓄雨水为目标的屋顶绿化技术在西宁地区具有一定的实施可能性。在雨水花园中选择种植在西宁能越冬且在多个已实施项目中长势良好的地被植物,例如:水烛(Typha angustifolia)、鸢尾(Iris tectorum)、金娃娃萱草(Hemerocallis fulva ‘Golden Doll’)、荷兰菊(Symphyotrichum novi-belgii)、蓍草(Achillea millefolium)、辽东水蜡(Ligustrum obtusifolium)等;同时为了改善现状植物景观乔、灌、草层次交错杂乱的问题,设计中增加了底层灌木种类数量,其与上层的乔木共同组合成配套的景观,在沿道路两侧围合出较为完整的活动和休闲空间。
安泰华庭小区的建筑内排水雨落管雨水与外围车行道的径流雨水无法进入绿地进行消纳,如果舍弃这部分雨水会导致无法完成上位规划要求的就地消纳水量,因此在雨水管网接入大市政管网前设计了地下调蓄池(图15),满足了上位规划和雨水回用的比例的双重要求。设计中在地下调蓄池中安装了潜水泵,雨水回用管与小区灌溉管网连接,可引至绿化屋面、景观水系和雨水景观墙等部位,将雨水回用与灌溉、造景相结合。
3. 设计方案的SWMM模拟分析
3.1 模型的建立与模拟
3.1.1 模型参数率定
上述设计方案完成了上位规划对安泰华庭小区下达的雨水管理指标任务。为了验证设计的合理性,得到准确的数据分析,需要建立SWMM模型。模型共建立了49个子汇水区,78个节点,77条排水管道,管径400 ~ 600 mm之间,管网末端排放口C1、C3共计2个,其中C1口排放I、II区的雨水,C3口排放III区的雨水。安泰华庭小区的SWMM概化模型如图16所示。
SWMM模型相关参数子汇水区面积、汇水宽度和坡度根据研究区域基础数据直接计算获取;水文水力模块率定参数曼宁系数、地表洼蓄量和入渗模型参数,根据模型手册中的典型值和测试值以及《西宁市海绵城市建设项目系统性详细规划(2016—2018)》中的推荐参考值来进行参数调试,验证模型的可靠性,具体取值如表3;模型中的雨水干管管长由模型自动测量工具获取;雨水管道的曼宁系数根据手册经验值取0.013,雨水明渠的曼宁系数根据手册经验值取0.025;雨水传输采用运动波方程,下渗采用Horton模型[20-21]。设绿色屋顶和生态滞留池两类LID设施,设计参数取值范围见表4。
表 3 水文水力模块参数Table 3. Hydrological and hydraulic module parameters曼宁粗糙率 Manning roughness 地表洼蓄量 Depression storage Horton 渗透模型参数 Horton percolation model parameter 不透水粗糙率
N-imperv透水粗糙率
N-perv不透水洼蓄量
Des-imperv/mm透水洼蓄量
Des-perv/mm无低洼地不
透水区比例
Zero-imperv/%最大渗透率
Max. infil/
(mm·h− 1)最小渗透率
Min. infil/
(mm·h− 1)衰减常数
Decay
constant/h− 1干燥时间
Drying
time/d0.013/0.025 0.24 1.27 3.18 25 40 6 4.14 7 表 4 LID设施参数Table 4. LID facility parameters设施类型 Facility type 设施结构 Construction of the facility 设施参数 Facility parameter 取值 Value 生物滞留池 Bio-retention 表面层 Surface layer 存水高度 Deep of water/mm 200 表面坡度 Surface slope/% 0.4 土壤层 Soil layer 厚度 Thickness/mm 700 孔隙度 Porosity 0.2 排水层 Drainage layer 厚度 Thickness/mm 200 孔隙率 Porosity 0.35 屋顶绿化 Green roof 表面层 Surface layer 存水高度 Deep of water/mm 50 表面坡度 Surface slope/% 2 土壤层 Soil layer 厚度 Thickness/mm 150 孔隙度 Porosity 0.3 排水垫层 Drainage layer 厚度 Thickness/mm 0 孔隙比 Void ratio 0 干燥时间 Drying time/d 7 本研究的降雨模型采用芝加哥雨型对降雨强度进行时间尺度上的分配,根据《西宁市排水工程专项规划(2012—2030)》和《西宁市排水(雨水)防涝综合规划(2012—2030)》相关资料,西宁市暴雨强度公式采用由西安建筑科技大学1998年推导的公式,具体如下:
q=461.9(1+0.993lgP)(t+3)0.686 (2) 式中:q为暴雨强度(L/(s·hm2));P为重现期(a);T为降雨历时(min)。
根据西宁市气象局近5年(2010—2015年)监测的单场典型降雨统计中,西宁降雨多以单一前锋雨型为主,峰值多出现在r = 0.375左右,因此本文采用芝加哥雨型,推求西宁市历时2 h、1 min间隔的设计暴雨雨型。利用公式(2)推求P为2、5、10、20和50年重现期下的设计雨量,依据芝加哥雨型方法得到雨型分配结果[22-23]。西宁地区不同重现期降雨特征如表5所示。
表 5 不同重现期西宁地区降雨的特征Table 5. Characteristics of rainfall in Xining in different recurrence intervals重现期
Recurrence interval2 h 降雨量
Rainfall in two hours/mm平均雨强
Average rainfall intensity/(mm·min− 1)峰值雨强
Peak rainfall intensity/(mm·min− 1)2年一遇 2 years interval 16.18 0.14 1.69 5年一遇 5 years interval 21.11 0.18 2.21 10年一遇 10 years interval 24.83 0.21 2.59 20年一遇 20 years interval 28.56 0.24 2.98 50年一遇 50 years interval 33.48 0.28 3.5 3.1.2 改造前后不同重现期各排水口流量分析
通过SWMM模拟分析可知,在改造前后,C1和C3排水口的产流速率出现明显变化。从图17 ~ 21分析可得出以下结论:
![]() 图 17 改造前后2年一遇2 h降雨下C1和C3排水口流量变化Figure 17. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in two years recurrence interval before and after reconstruction
图 17 改造前后2年一遇2 h降雨下C1和C3排水口流量变化Figure 17. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in two years recurrence interval before and after reconstruction![]() 图 21 改造前后50年一遇2 h降雨下C1和C3排水口流量变化Figure 21. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in fifty years recurrence interval before and after reconstruction
图 21 改造前后50年一遇2 h降雨下C1和C3排水口流量变化Figure 21. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in fifty years recurrence interval before and after reconstruction(1) LID设施的建设使得小区绿地消纳雨水的能力增强,在2年一遇2 h降雨量略高于设计的15.2 mm,C1及C3排水口均无径流产生,且在5年一遇时仍无径流产生,说明设计的LID设施具有较好的减排效果,达到了上位规划要求的目标。
(2) 在10、20及50年重现期降雨条件下,改造前C1排水口的流量分别为0.145、0.145、0.145 m3/s,峰现时间分别为第50、52、53 min,改造后C1排水口的流量分别达到0.024、0.030、0.061 m3/s,峰现时间分别为第132、92、83 min。改造后峰值分别降低了83.45%、79.31%和57.39%,峰现时间延迟了82、40和30 min,达到了很好的削峰延时效果。
(3) 在10、20及50年重现期降雨条件下,改造前C3排水口的流量分别为0.094、0.094、0.094 m3/s,峰现时间分别为第57、59、44 min,改造后C3排水口的流量分别达到0.032、0.062、0.073 m3/s,峰现时间分别为第122、109、106 min。改造后峰值分别降低了65.96%、34.04%和22.34%,峰现时间延迟了65、50和62 min,达到了很好的削峰延时的效果。
(4) 改造前5年一遇及以上降雨情景下C1、C3排水口的流量峰值均没有变化,说明该径流量超过了管网排水能力,出现了溢流现象。2年一遇的C1、C3排水口的流量峰值分别为0.139和0.093 m3/s,基本接近管网的满流状态。
(5) 从峰值流量看,改造后50年重现期下小区C1排水口的流量为0.061 m3/s,C3排水口的流量为0.073 m3/s,均低于改造前2年重现期排放口的流量0.139和0.093 m3/s,故可以认为LID措施使得小区雨水的防洪排涝能力由2年一遇提高到50年一遇。
![]() 图 18 改造前后5年一遇2 h降雨下C1和C3排水口流量变化Figure 18. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in five years recurrence interval before and after reconstruction
图 18 改造前后5年一遇2 h降雨下C1和C3排水口流量变化Figure 18. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in five years recurrence interval before and after reconstruction![]() 图 19 改造前后10年一遇2 h降雨下C1和C3排水口流量变化Figure 19. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in ten years recurrence interval before and after reconstruction
图 19 改造前后10年一遇2 h降雨下C1和C3排水口流量变化Figure 19. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in ten years recurrence interval before and after reconstruction![]() 图 20 改造前后20年一遇2 h降雨下C1和C3排水口流量变化Figure 20. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in twenty years recurrence interval before and after reconstruction
图 20 改造前后20年一遇2 h降雨下C1和C3排水口流量变化Figure 20. Flow variation of C1 and C3 drainage outlets under 2-hour rainfall in twenty years recurrence interval before and after reconstruction3.2 改造实施后的监测数据分析
由于工期、投资等原因限制,安泰华庭小区目前仅在III区实施了海绵化改造,但是在C1、C3两个排水口均安装了雨水监测装置以监测雨水流量和SS浓度,为项目数据验证提供机会。
本文选取2018年5—6月间监测到的具有代表性的3场典型降雨(表6)作为数据样本,使用先前建立的SWMM模型对3场降雨的数据进行模拟,将模拟的产流速率与实测的数据进行对比,以验证SWMM模型参数选择的合理性以及海绵化改造的有效性。
表 6 3场降雨的特征数据Table 6. Characteristic data of three rainfall events降雨时间
Rainfall time总降雨量
Total rainfall depth/mm最大降雨强度
Max. rainfall intensity/(mm·min− 1)降雨时段平均雨强
Average rainfall intensity/(mm·min− 1)降雨等级
Rainfall class5月9日08:00—10日07:59
9 May 08:00−10 May 07:597.5 0.1 0.1 小雨
Light rain5月21日08:00—22日07:59
21 May 08:00−22 May 07:5910.5 0.1 0.1 中雨
Moderate rain6月7日08:00—8日07:59
7 June 08:00−8 June 07:5912 0.5 0.16 中雨
Moderate rain3.2.1 C1排水口的模拟与监测数据对比分析
利用3次降雨的实际监测值对安泰华庭小区C1排水口的SWMM模型模拟,将模拟结果(图22、23、24)与实测值(图25、26、27)分别进行对比,发现在C1排水口二者产流曲线基本吻合,模拟的峰值流量分别为107.82、130.14、347.58 m3/h,而实测值分别为106.515、190.247、338.928 m3/h,误差率在1.31%和2.49%之间,其中5月21日的峰值流量误差较大(31.59%)。降雨时间段内共计产生外排量模拟值分别为291.34、424.51、459.19 m3,而实测值为279.47、437.97、440.37 m3,误差率为3.17% ~ 4.10%之间。上述数据对比分析说明在安泰华庭小区建立的SWMM模型能够准确地模拟降雨产流状况,模型的参数选择符合实际情况。
![]() 图 22 5月9日降雨SWMM模型模拟C1排水口流量曲线Figure 22. Simulated flow curve of C1 outlet by SWMM model on 9 May
图 22 5月9日降雨SWMM模型模拟C1排水口流量曲线Figure 22. Simulated flow curve of C1 outlet by SWMM model on 9 May![]() 图 23 5月21日降雨SWMM模型模拟C1排水口流量曲线Figure 23. Simulated flow curve of C1 outlet by SWMM model on 21 May
图 23 5月21日降雨SWMM模型模拟C1排水口流量曲线Figure 23. Simulated flow curve of C1 outlet by SWMM model on 21 May![]() 图 24 6月7日降雨SWMM模型模拟C1排水口流量曲线Figure 24. Simulated flow curve of C1 outlet by SWMM model on 7 June
图 24 6月7日降雨SWMM模型模拟C1排水口流量曲线Figure 24. Simulated flow curve of C1 outlet by SWMM model on 7 June3.2.2 C3排水口的模拟与监测数据对比分析
利用3次降雨的实际监测值对安泰华庭小区III区的SWMM模型模拟,理论上3次降雨均不会在C3排水口产生径流,而实际监测数据发现在C3排水口3次降雨均发生溢流现象,有少量雨水外排(图28、29、30)。其中:5月9日C3号排口最大流量值为12.482 m3/h,监测时间段内共计产生外排水量为23.587 m3,平均SS浓度为56 mg/L;5月21日C3号排口最大流量值为31.482 m3/h,监测时间段内共计产生外排水量为42.070 m3,平均SS浓度为79 mg/L;6月7日C3号排水口最大流量值为58.944 m3/h,监测时间段内共计产生外排水量为26.717 m3,平均SS浓度为2 mg/L。
对比3次降雨的外排总量,发现其数值与降雨总量不是成比例关系,尤其是6月7日的最大雨强和平均雨强都比较大,而外排水量没有显著增加,说明监测到的雨水外排不是因降雨造成的。分析溢流产生的原因,应该是LID设施运行管理造成的。在安泰华庭的III区调蓄水池设计容量为300 m3,每次降雨结束后,调蓄池应及时清空库容,为调蓄雨水做准备。而在实际运行中,由于安泰华庭小区车库顶源源不断有土壤渗水排出,尤其是排入下凹绿地的雨水除了蒸发蒸腾的部分外,都会经由车库顶板的盲沟管进入雨水管网,汇流到调蓄池中,形成一个闭合循环。这是由原排水系统的设计带来的结果,无法通过海绵化改造消除。由于降雨前调蓄水池有一定的基本库容,导致3次小到中雨均出现了溢流现象。
3.2.3 C1与C3排水口的监测数据对比分析
通过对3次降雨C1和C3排水口的监测数据对比分析(表7),可以发现3次降雨过程中产流峰值、出流量均大幅度降低,充分说明了该项目通过海绵化的改造,有效地实现了雨水的减排。
表 7 C1和C3排放口的监测数据对比Table 7. Comparison of monitoring data for C1 and C3 outlet降雨时间
Rainfall time降雨量
Rainfall depth/mm排放口
Outlet汇水区面积/hm2
Catchment area/ha峰值流量
Peak flow rate/(m3·h− 1)外排总量
Discharge volume/m3单位面积外排量/(m3·hm− 2)
Unit area discharge/(m3·ha− 1)5月9日
9 May7.5 C1 5.69 106.515 279.471 49.1 C3 4.96 12.482 23.587 4.8 5月21日
21 May10.5 C1 5.69 190.247 437.967 77.0 C3 4.96 31.482 42.070 8.5 6月7日
7 June12 C1 5.69 338.928 440.366 77.4 C3 4.96 58.944 26.717 5.4 4. 结论与讨论
安泰华庭小区的海绵化改造项目从2016年7月开始,完成了项目设计、实施及后期维护管理的全过程。无论是SWMM理论模型验证还是实际监控监测检验,都证明该改造项目是成功的。尤其是两种不同来源的数据相互校验,增强了检验的可信度。
从模拟数据来看,安泰华庭的海绵化改造满足上位规划提出的设计要求。在5年一遇2 h降雨量排水口均无径流产生,设计的LID设施具有较好的减排效果,达到了上位规划要求的目标。在10、20及50年重现期降雨条件下,改造后的C1排水口的流量峰值分别降低了83.45%、79.31%和57.39%,峰现时间延迟了82、40和30 min;改造后C3排水口的流量峰值分别降低了65.96%、34.04%和22.34%,峰现时间延迟了65、50和62 min,达到了良好的削峰延时效果。改造后50年重现期下排水口的流量均低于改造前2年重现期排放口的流量,小区的排涝能力由2年一遇提高到50年一遇。
从监测数据来看,以2018年5月和6月3次实际降雨值验证未改造的C1排水口流量,验证结果发现模拟数据与实际监控数据能够很好地匹配,说明设计模型的参数率定合理,模拟结论具有较高的可信度。改造完成的区域相对于未改造区域的产流峰值、出流量均大幅度降低,充分说明了海绵化改造的实施,有效地削减了外排量,实现了削峰、延时、去污等综合目标。
在安泰华庭小区的海绵化改造项目的设计和实施过程中,也存在一些值得探讨的问题。在高海拔严寒地区,建筑内排水的雨落管设计使得屋面雨水无法引流至地面,海绵化改造的难度倍增。地下车库是当前解决小区停车问题的主要手段,如果车库顶覆土厚度不足也会带来雨水利用的困难,选择适合的位置安排下凹绿地和调蓄池成为设计的主要难点。上述问题应在小区规划和建筑设计阶段时提前布局安排,为海绵城市建设创造有利条件。在LID技术选择方面,在高寒地区使用屋顶绿化技术具有一定可行性,托盘容器式的屋顶绿化适用于各类非上人屋顶。在安泰华庭小区的改造费用中,地下调蓄池占比达到30%以上,高造价、高维护的特点使得地下调蓄池不应成为海绵城市建设设施选择的优先项。由于西宁处于黄河流域的上游区域,排入河道的雨水有较多的机会被下游地区所利用,减量与削峰的压力都不大。从雨洪管理系统的整体设计和工程造价方面综合考虑,雨水的储存利用不是西宁市海绵城市建设源头控制的最佳选择。湿陷性黄土、小区车库顶板覆土薄等自然及人为因素限制了渗透利用的实施,海绵城市设计的目标应着眼于滞留与净化。
-
![]()
图 2 某植物在温度环境因子中的适应区间和耐受区间
Figure 2. Adaptation range and tolerance interval of a plant in the temperature environmental factors
![]()
图 3 2种植物随温度变化的动态演替过程
Figure 3. Dynamic evolution of two kinds of plants with temperature
![]()
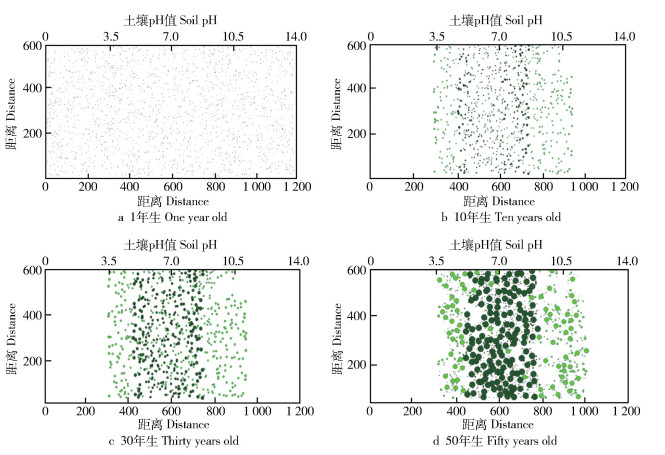
图 4 2种植物随土壤pH值变化的动态演替过程
Figure 4. Dynamic evolution of two kinds of plants with soil pH
![]()
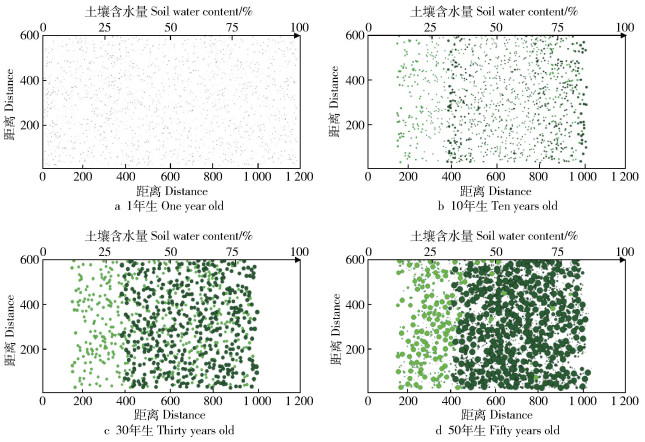
图 5 2种植物随土壤含水量变化的动态演替过程
Figure 5. Dynamic evolution of two kinds of plants with soil water content
![]()
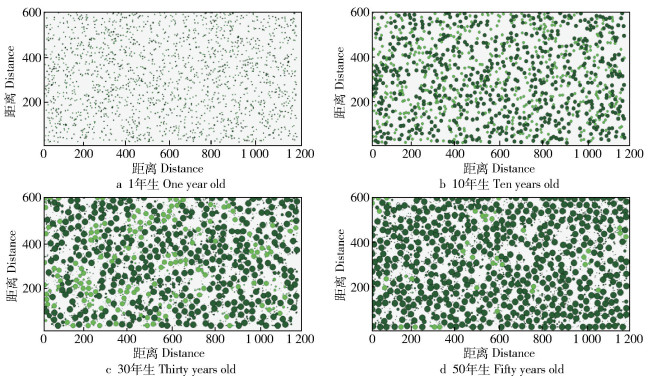
图 7 3种植物的动态演替过程
绿色植物代表马尾松; 红色植物代表杉木; 黄色植物代表白栎。
Figure 7. Dynamic evolution of three kinds of plants
Green plant represents wood of Pinus massoniana; red plant represents wood of Cunninghamia lanceolata; yellow plant represents wood of Quercus fabri.
表 1 2种植物的实验参数设置
Table 1 Parameter settings of two kinds of plants
参数 深绿色植物 浅绿色植物 Parameter Dark green plant Light green plant 温度权重系数Temperatureweight coefficient 1.00 0.25 温度适应区间Temperature adaptation interval/℃ 22 ~ 25 22 ~ 26 最低耐受温度Minimum tolerance temperature/℃ 12 1 最高耐受温度Maximum tolerance temperature/℃ 30 28  下载: 导出CSV
下载: 导出CSV
表 2 2种植物的土壤pH值实验参数设置
Table 2 Parameter settings in soil pH of two kinds of plants
参数Parameter 深绿色植物Dark green plant 浅绿色植物Light green plant 土壤pH值权重系数Weight coefficient of soil pH 1.00 0.25 土壤pH值适应区间Adaptation interval of soil pH 6.9 ~ 7.8 6.7 ~ 7.3 最低耐受土壤pH值Minimum tolerance of soil pH 4.0 4.5 最高耐受土壤pH值Maximum tolerance of soil pH 9.8 11.5
下载: 导出CSV
表 3 种植物的土壤含水量实验参数设置
Table 3 Parameter setting in soil water content of two kinds of plants
参数
Parameter深绿色植物
Dark green plant浅绿色植物
Light green plant土壤含水量权重系数Weight coefficient of soil water content 1.00 0.25 土壤含水量适应区间Adaptive range of soil water content/% 49~60 40~51 最低耐受土壤含水量Minimum tolerance of soil water content/% 32 18 最高耐受土壤含水量Maximum tolerance of soil water content/% 85 80
下载: 导出CSV
-
[1] 桑卫国, 马克平, 陈灵芝, 等.森林动态模型概论[J].植物学通报, 1999, 16(3):193 - 200. doi: 10.3969/j.issn.1674-3466.1999.03.001 SANG W G, MA K P, CHEN L Z, et al. A brief review on forest dynamics models[J]. Chinese Bulletin of Botany, 1999, 16(3):193 - 200. doi: 10.3969/j.issn.1674-3466.1999.03.001
[2] 刘永霞.北京山地油松林分生长过程数量化模拟研究[D].北京: 北京林业大学, 2007. LIU Y X. Study on quantitative simulation of growth process for Chinese pine stand in mountain areas of Beijing[D]. Beijing : Beijing Forestry University, 2007.
[3] 周丹卉, 贺红士, 孙国臣, 等.林窗模型及其在全球气候变化研究中的应用[J].生态学杂志, 2007, 26(8): 1303 - 1310. http://d.old.wanfangdata.com.cn/Periodical/stxzz200708028 ZHOU D H, HE H S, SUN G C, et al. Gap model and its application in studies of global climate change[J]. Chinese Journal of Ecology, 2007, 26(8): 1303 - 1310. http://d.old.wanfangdata.com.cn/Periodical/stxzz200708028
[4] 霍常富, 赵晓敏, 鲁旭阳, 等.林窗模型研究进展[J].世界林业研究, 2009, 22(6):43 - 48. http://d.old.wanfangdata.com.cn/Periodical/sjlyyj200906007 HUO C F, ZHAO X M, LU X Y, et al. Review on forest gap models[J].World Forestry Research, 2009, 22(6):43 - 48. http://d.old.wanfangdata.com.cn/Periodical/sjlyyj200906007
[5] 梁晓东, 叶万辉.林窗研究进展[J].热带亚热带植物学报, 2001, 9(4):355 - 364. doi: 10.3969/j.issn.1005-3395.2001.04.015 LIANG X D, YE W H.Advances in study on forest gaps[J].Journal of Tropical and Subtropical Botany, 2001, 9(4):355 - 364. doi: 10.3969/j.issn.1005-3395.2001.04.015
[6] 程肖侠.气候变化背景下中国东北森林的演替动态[D].北京: 中国科学院研究生院, 2007. http://cdmd.cnki.com.cn/Article/CDMD-80058-2007179153.htm CHENG X X. Succession dynamics of the forest in the northeast of China under the climate changing scenarios[D].Beijing: Graduate University of Chinese Academy of Sciences, 2007. http://cdmd.cnki.com.cn/Article/CDMD-80058-2007179153.htm
[7] YODA K, KIRA T, OGAWA H. Self-thinning in overcrowded pure stands under cultivated and natural conditions[J]. Journal of Biology, Osaka City University, 1963, 14: 107 - 129.
[8] GREEN D G. Modelling plants in landscapes[M]//MICHALEWICZ M T. Plants to ecosystems-advances in computational life sciences. Melboume: CSIRO Publishing, 1997: 85 - 96.
[9] BERGER U, HILDENBRANDT H, GRIMM V. Towards a standard for the individual based modeling of plant populations:self-thinning and the field of neighborhood approach[J]. Natural Resource Modeling, 2002, 15(1): 39 - 54. doi: 10.1111/j.1939-7445.2002.tb00079.x
[10] BERGER U, HILDENBRANDT H. A new approach to spatially explicit modeling of forest dynamics: spacing, ageing and neighborhood competition of mangrove trees[J]. Ecological Modeling, 2000, 132(3): 287 - 302. doi: 10.1016/S0304-3800(00)00298-2
[11] DEUSSEN O, HANRAHAN P, LINTERMANN B, et al. Realistic modeling and rendering of plant ecosystems[C]//Proceedings of the 25th Annual Conference on Computer Graphics and Interactive Techniques. New York: ACM Press, 1998: 275 - 286.
[12] LANE B, PRUSINKIEWICZ P. Generating spatial distributions for multilevel models of plant communities[C]//Proceedings of the Graphics Interface Calgary. Alberta: Canadian Human-Computer Communications Society Press, 2002: 69 - 80.
[13] ALSWEIS M, DEUSSEN O. Modeling and visualisation of symmetric and asymmetric plant competition[C]//Proceedings of Eurographics Workshop on Natural Phenomena'05. Dublin: Eurographics Association, 2005: 83 - 88.
[14] BENES B, GUERRERO J M S. Clustering in virtual plant ecosystems[C]//Proceedings of WSCG'04. Plzen : Union Agency-Science, 2004: 9 - 16.
[15] 单梁, 杨刚, 黄心渊.森林动态演替现象的可视化模拟[J].中国图象图形学报, 2013, 18(12): 1666 - 1675. doi: 10.11834/jig.20131216 SHAN L, YANG G, HANG X Y. Simulation of the dynamic evolution of spatial distribution of trees in the forest[J]. Journal of Image and Grafics, 2013, 18(12): 1666 - 1675. doi: 10.11834/jig.20131216
[16] 李清芳, 辛天蓉, 马成仓, 等. pH值对小麦种子萌发和幼苗生长代谢的影响[J].安徽农业科学, 2003, 31(2): 185 - 187. doi: 10.3969/j.issn.0517-6611.2003.02.007 LI Q F, XIN T R, MA C C, et al. Effect of pH value on wheat seed germination and seedlings growth and metabolism[J]. Journal of Anhui Agricultural Sciences, 2003, 31(2): 185 - 187. doi: 10.3969/j.issn.0517-6611.2003.02.007
-
期刊类型引用(7)
1. 丁金华,许艳秋,钱晶. 苏南水网地区水域景观破碎化时空演变特征及驱动因子研究——以吴江区为例. 西北林学院学报. 2024(01): 247-255 .  百度学术
百度学术
2. 陈实,陈丽捷,洪宇,刘金福,阙翔,李意敏,何东进,赵婧雯. 泉州湾湿地碳库生态安全评价及其障碍因素研究. 福建农林大学学报(自然科学版). 2024(05): 686-695 . 百度学术
3. 杨烜涵,付晖,秦煜姬,程恩起,陈圣天. 近20年来海口湿地景观格局演变及其驱动因子. 中国城市林业. 2023(03): 28-35 . 百度学术
4. 王琦,刘子刚,周隽伊. 三江平原沼泽湿地变化的影响因素及其空间效应. 中国人口·资源与环境. 2023(07): 44-54 . 百度学术
5. 赵红梅,毛欣,刘春雷,李亚松,刘林敬. 福建泉州湾海岸带MIS 3阶段以来的海侵—海退过程. 地质力学学报. 2023(04): 569-583 . 百度学术
6. 朱映辰,谭芳林,阙翔,洪宇,潘爱芳,刘金福. 多时间尺度下森林公园负离子变化特征及与温湿度关系研究. 西北林学院学报. 2023(06): 211-218+227 . 百度学术
7. 赵红梅,刘春雷,毛欣,毕志伟,刘哲,李亚松. 泉州湾海岸带全新世地层及沉积环境演化. 地层学杂志. 2022(04): 401-410 . 百度学术
其他类型引用(5)




































计量
- 文章访问数: 3814
- HTML全文浏览量: 737
- PDF下载量: 125
- 被引次数: 12
