
Probability distribution and selection of seed and fruit traits of different fruit types of Xanthoceras sorbifolium
-
摘要: 文冠果是无患子科单属种,其种群野生濒危,生境破碎化严重且具有自交不亲和性。文冠果有“千花一果”之称,为提高其产量,测量了4年生文冠果实生园1 051个单株的坐果序数、坐果数、果实质量、种子质量、壳质量、平均单粒质量共6个种实性状并对各种实性状按不同果型进行了单因素方差分析和分布拟合。结果表明:不同果型之间果实质量、种子质量、壳质量、平均单粒质量4个种实性状之间差异极显著,而坐果序数和坐果数2个种实性状之间差异不显著。所有种实性状均符合威布尔分布,其中果实质量、种子质量、壳质量、坐果序数和坐果数5个种实性状均符合左偏威布尔分布,说明丰产是个“小概率”事件;平均单粒质量近似正态分布,说明实验园整体种子大小较为稳定,极大值与极小值的出现概率均比较低。基于威布尔概率分布建立了早期性状选择的模型,模型预测大桃型、倒卵型、尖柱型、凸桃型、短桃型、柱型在种子质量和平均单粒质量上选择概率均比较高,可作为丰产果型,其中大桃型为丰产的最理想果型;小柱型、小桃型和短柱型的种子质量和平均单粒质量的选择概率较低,不适合作为丰产选优果型。因此可根据该分布的特性对不同果型进行选择,为进一步的育种选优提供理论支持。Abstract: Xanthoceras sorbifolium is the only species in this genus that wild-endangered in China, which may due to self-incompatible and habitat destruction severely. The paper aims to improve the yield since it was widely known as "a tree of thousand flower but one fruit". Six phenotypes for 1 051 individual trees in four-year-old seed orchard were measured in this study, including fruit number, infructescence number with fruit setting, fruit mass, seed mass, shell mass and average single grain mass. ANOVA analysis and distribution fitting based on different fruit types were carried out in all the traits. Results showed that there were significant differences in varied fruit type seeds and fruit traits, including fruit mass, seed mass, shell mass and average single grain mass, but not in the infructescence number and fruit number. All the seed and fruit traits were in accordance with Weibull distribution, the fruit mass, seed mass, shell mass, infructescence number and fruit number were in accordance with the left partial Weibull distribution, indicating that high yield was a "small probability" event; the average single grain mass was similar to the normal distribution, indicating that the seed size in the whole plantation was relative stable, the frequency of maximum and minimum probability was relatively low. The model of early trait selection was established based on Weibull probability distribution, indicating that big peach type, inverted egg type, pointed pillar type, convex peach type, short peach type and pillar type were higher in seed mass and average single grain mass, which can be used as high-yield fruit type, and the big peach type was the most ideal fruit type of high yield. Small pillar type, small peach type and short pillar type selection probability in seed mass and average single-grain mass were much lower, which was not suitable as a high yield fruit selection. Hence, the characteristics of the Weibull distribution can provide support for selecting excellent fruit types and further theory support of breeding selection.
-
多倍体育种是杨树(Populus spp.)遗传改良的重要途径。‘三毛杨’(P. tomentosa ‘Sanmaoyang’)、‘中林46’(P. × euramericana ‘Zhonglin-46’)、‘银中杨’(P. alba × P. berolinensis ‘Yinzhong’)、‘北林雄株1号’(P. × ‘Beilinxiongzhu 1’)、‘中怀1号’(P. × ‘Zhonghuai 1’)等三倍体品种均在我国林业生产中广泛利用[1-5],对推动我国林业产业升级、改善生态环境做出了重要贡献。为了指导林木多倍体品种选育,朱之悌[6]曾提出了“大群体,强选择”的育种策略,认为多倍体育种同样要基于大群体,依据主要目标性状对多倍体种质进行选择,经过遗传测定,才能形成表现优良的新品种。因此,在杨树多倍体品种选育过程中,对性状变异规律进行遗传分析十分必要。
就杨树多倍体育种而言,基于2n配子的有性多倍化途径综合了杂合效应和倍性效应,提供了更加丰富的遗传变异[7]。植物生长通过叶片的光合作用产生物质和能量[8],叶片大小和形状影响植物的气体交换和蒸腾作用[9],有关植物叶片和气孔性状的研究可间接反映植物的植物生长发育及生理代谢特征[10]。Liao等[11]分析了‘哲引3号杨’ × ‘北京杨’全同胞杂种三倍体和二倍体的苗期表型,发现三倍体群体整体具有生长和光合优势,其中也存在一些光合效率高但生长较差的个体和光合效率低但生长较好的个体,可见,异源三倍体杨树的性状变异规律非常复杂。已有研究表明,雌雄异株植物在叶面积、生长势、生物量等方面存在差异[12],性状表现也会受到性别的影响。然而,关于叶片及气孔性状的研究多集中在苗期植株的分析,尚缺少对于节间生长缓慢且已进入开花成熟期的杨树枝条上的短枝功能叶片及气孔性状的变异规律的研究,且对于异源三倍体杨树,性别效应对性状表现产生的影响尚不明确。因此,为进一步丰富杨树三倍体叶片及气孔性状的遗传变异特征,本研究以‘哲引3号杨’ × ‘北京杨’全同胞杂种二倍体和三倍体的10年生种质保存林为材料,对其大树短枝功能叶片及气孔性状变异规律进行系统研究,解析倍性、基因型和性别效应对叶片及气孔性状变异的影响大小,为杨树三倍体育种提供理论依据。
1. 材料与方法
1.1 试验材料
试验材料采集自河北威县的‘哲引3号杨’ × ‘北京杨’全同胞杂种二倍体和三倍体种质保存林,4株小区,3次重复,树龄10年,其中包括二倍体无性系17个(雌株7个,编号为D9、D24、D25、D34、D44、D45、D52;雄株10个,编号为D2、D3、D4、D11、D14、D20、D21、D38、D41、D43),三倍体无性系19个(雌株9个,编号为T6、T12、T15、T16、T19、T23、T26、T32、T40;雄株10个,编号为T13、T18、T22、T27、T30、T31、T33、T36、T37、T42)。每个无性系从每小区的4株中随机选择1株作为采样树,共3次重复,采集位于树冠中部北侧的无病虫害短枝功能叶片进行叶片长度、叶片宽度、叶面积、叶柄长度、叶缘锯齿数、气孔长度、气孔宽度、气孔密度等性状测量。
1.2 试验方法
1.2.1 短枝叶片性状测量
每株采样树随机选取30片形态完整的短枝叶片,每个无性系重复3株采样树,用CI-203便携式激光叶面积仪(美国CID)测量叶片长度、叶片宽度、叶面积和叶柄长度,并对叶缘锯齿数进行统计。
1.2.2 气孔性状
每株采样树随机选取5片形态完整的短枝叶片,每个无性系重复3株采样树,采用指甲油涂抹撕取法进行气孔性状观察[13]。每个叶片随机选择50个处于关闭状态的气孔,于Olympus BX51显微镜下测量气孔长度和气孔宽度;随机选择10个视野统计气孔密度。
1.2.3 数据统计分析
利用Excel整理数据,由于各性状数据均不满足正态分布或方差齐次,应用SPSS 17.0软件的非参数Kruskal−Wallis检验进行差异显著性分析。应用R语言对各性状进行Spearman相关分析和方差贡献率的计算,将方差分量的剩余项视为环境效应。
无性系重复力(R)计算公式:
R=σc2/(σc2+σe2/k) 式中:σc2为无性系方差,σe2为随机误差,k为每个无性系采集样本数的调和平均值[14]。
2. 结果与分析
2.1 青黑杨全同胞杂种短枝叶片及气孔性状基因型间的变异分析
青黑杨全同胞杂种植株短枝叶片及气孔各性状在无性系之间均存在极显著差异(表1)。无性系之间叶片性状变异较大,变异系数介于14.23% ~ 27.20%之间,其中锯齿数变异系数最高,叶片宽度变异系数最小;气孔性状变异系数在8.73% ~ 32.73%之间,其中气孔密度变异系数最高,气孔宽度变异系数最低,表明叶片宽度和气孔宽度是所测性状中较稳定的遗传特征。
表 1 ‘哲引3号杨’ × ‘北京杨’杂种二倍体和三倍体叶片及气孔性状无性系重复力及变异系数Table 1. Repeatability and variation coefficients of leaves and stomatal traits of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis性状 Trait 二倍体 Diploid 三倍体 Triploid P值 P value 无性系重复力 Repeatability of clone 变异系数 CV/% 变化范围
Variation range均值 ± 标准误 Mean ± SE 变化范围
Variation range均值 ± 标准误 Mean ± SE 倍性 Ploidy 性别Gender 基因型Genotype 叶片长度
Leaf length/cm6.26 ~ 9.35 7.52 ± 0.14 6.59 ~ 10.51 8.65 ± 0.18 < 0.01 0.86 < 0.01 0.903 15.23 叶片宽度
Leaf width/cm4.83 ~ 7.48 6.05 ± 0.13 5.45 ~ 7.92 6.85 ± 0.11 < 0.01 0.33 < 0.01 0.836 14.23 叶面积
Leaf area/cm217.76 ~ 39.92 27.32 ± 0.97 21.61 ~ 41.97 33.41 ± 1.09 < 0.01 0.72 < 0.01 0.789 25.32 叶柄长度
Petiole length/cm3.32 ~ 5.33 4.35 ± 0.10 3.28 ~ 5.95 4.48 ± 0.12 0.41 0.29 < 0.01 0.829 16.95 锯齿数
Serration number19 ~ 74 42.1 ± 2.0 30 ~ 55 38.4 ± 1.1 0.10 0.34 < 0.01 0.924 27.20 气孔长度
Stomatal length/μm20.67 ~ 28.29 23.58 ± 0.30 24.08 ~ 30.66 27.59 ± 0.30 < 0.01 0.79 < 0.01 0.954 11.13 气孔宽度
Stomatal width/μm12.82 ~ 16.17 14.53 ± 0.14 14.15 ~ 17.66 16.27 ± 0.16 < 0.01 0.28 < 0.01 0.952 8.73 气孔密度/(个·mm−2)
Stomatal density/
(number·mm−2)87 ~ 279 188.6 ± 8.1 95 ~ 191 128.5 ± 3.8 < 0.01 0.06 < 0.01 0.947 32.73 重复力作为一个重要的遗传参数,反映了无性系优良性状受特殊环境影响的程度[15],其大小还表明了性状稳定性的高低[16]。本研究中,叶片性状无性系重复力介于0.789 ~ 0.924之间,气孔性状无性系重复力介于0.947 ~ 0.960之间,气孔性状的遗传效应略大于叶片性状。可见,青黑杨全同胞杂种植株叶片和气孔性状受到强度的遗传控制,环境影响较小,有利于无性系的评价选择,更利于较为准确地解析各效应对性状变异的影响。
2.2 青黑杨全同胞杂种植株短枝叶片性状变异规律分析
不同倍性群体间比较,三倍体植株平均叶片长度、叶片宽度、叶面积均显著大于二倍体植株,呈现出一定的巨大性特征(表1,图1a、b);而叶柄长度和锯齿数性状则无显著差异(表1)。在不同性别间,尽管雄株的叶片宽度、叶面积和叶柄长度比雌株分别大3.14%、2.02%和3.70%,叶片长度和锯齿数分别小1.10%和1.97%,但是这些叶片性状在雌、雄株之间并未呈现显著差异。
![]() 图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis
图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis从叶片各性状相关分析可知(图2),叶面积与叶片长度、叶片宽度呈极显著的正相关,分别达0.86和0.94;叶柄长度与叶片长度、叶片宽度、叶面积之间呈显著弱正相关,与锯齿数相关关系不显著;锯齿数与叶片宽度、叶面积呈显著弱正相关,与叶片长度的相关关系不显著。进一步分析发现(表2),叶片长度、叶片宽度和叶面积与倍性水平呈极显著的正相关(r分别为0.44、0.43、0.38,P < 0.01),叶柄长度和锯齿数与倍性水平之间无显著相关性(分别为r = 0.09,P = 0.41和r = −0.17,P = 0.10),而性别与叶片性状之间均没有显著的相关性(P > 0.05)。
![]() 图 2 叶片和气孔性状的Spearman相关分析*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.Figure 2. Spearman correlation analysis of leaves and stomatal traits表 2 叶片及气孔性状与倍性、性别的相关性分析Table 2. Correlation analysis of leaves and stomatal traits with ploidy and gender
图 2 叶片和气孔性状的Spearman相关分析*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.Figure 2. Spearman correlation analysis of leaves and stomatal traits表 2 叶片及气孔性状与倍性、性别的相关性分析Table 2. Correlation analysis of leaves and stomatal traits with ploidy and gender项目
Item叶片长度
Leaf length叶片宽度
Leaf width叶面积
Leaf area叶柄长度
Petiole length锯齿数
Serration number气孔长度
Stomatal length气孔宽度
Stomatal width气孔密度
Stomatal density倍性 Ploidy r值 r value 0.44 0.43 0.38 0.09 −0.17 0.72 0.65 −0.60 P值 P value < 0.01 < 0.01 < 0.01 0.41 0.10 < 0.01 < 0.01 < 0.01 性别 Gender r值 r value −0.02 0.10 0.04 0.11 −0.10 0.03 −0.11 0.20 P值 P value 0.86 0.33 0.72 0.29 0.34 0.80 0.28 0.06 2.3 青黑杨全同胞杂种植株短枝叶气孔性状变异规律分析
三倍体植株的平均气孔长度和气孔宽度均极显著大于二倍体,而平均气孔密度极显著小于二倍体(表1,图1c、d)。在不同性别间比较发现,雄株群体的平均气孔长度和气孔密度均大于雌株,气孔宽度小于雌株,但是均为呈现显著差异。
从气孔各性状的相关分析可知(图2),气孔长度与气孔宽度呈极显著的正相关,达0.89,气孔密度与气孔长度和气孔宽度呈极显著的负相关,分别为−0.84和−0.76。进一步分析发现(表2),气孔长度和气孔宽度与倍性水平间均呈极显著正相关(r分别为0.72和0.65,P < 0.01),气孔密度与倍性水平呈极显著负相关(r = −0.60,P < 0.01),而性别与气孔性状之间均没有显著的相关性(P > 0.05),表明气孔性状与倍性水平密切相关,可作为鉴定杨树多倍体的有效指标。
2.4 影响青黑杨全同胞杂种叶片及气孔性状各效应的方差贡献率解析
为深入解析倍性、性别和基因型效应对各性状表现的影响程度,计算各效应的方差分量及方差贡献率后发现(图3),气孔性状的变异主要来源于倍性效应,其次是基因型效应和环境效应。其中,气孔长度的倍性效应方差分量在表型总变异中所占比例高达63.79%,其次是气孔宽度的57.05%,气孔密度的47.10%,表明这些性状受倍性水平的影响最大;叶片性状变异的主要来源于基因型效应,其中,锯齿数变异的基因型效应方差贡献率最高,达80.72%,其次是叶柄长度变异的基因型效应为61.66%,表明叶片性状主要受基因型的影响。性别效应对叶片及气孔性状的影响均很小,其中,性别效应对气孔密度的方差贡献率仅为0.06%,对锯齿数和叶柄长度变异的方差贡献率甚至均为0。
3. 讨 论
多倍化是植物进化和适应性改变的重要力量[17-18]。由于基因组剂量的增加所带来的倍性效应往往导致多倍体植物产生广泛的性状变异[19-20]。同源四倍体苏丹凤仙花(Impatiens walleriana)在叶面积、叶片厚度、子房宽度、花粉粒长度和气孔大小等性状上增加,花瓣总花青素含量较高,而株高、气孔保卫细胞密度和花数减少[21]。滇杨(P. yunnanensis)同源四倍体和嵌合体的苗期长枝叶分析发现,与叶片长度、宽度和气孔性状大于二倍体,而气孔密度则显著小于二倍体[22]。本研究发现青黑杨杂种的叶柄长度和锯齿数变异受倍性效应影响较小,而三倍体植株平均叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度均极显著大于二倍体,气孔密度则极显著小于二倍体,且气孔性状变异的倍性效应方差贡献率最高,表明倍性效应对短枝叶的性状表现同样产生重要影响。而且,叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度与倍性水平之间均存在极显著正相关性,气孔密度与倍性水平呈极显著负相关,叶柄长度和锯齿数与倍性水平无显著相关性,因此,在倍性检测时可通过叶片大小和气孔性状相结合作为初步鉴定。此外,值得注意的是,本研究测得的气孔长度和气孔宽度大小与王君等[23]测得的数据相比均偏小,可能是由于杨树长枝和短枝叶片的气孔长宽大小存在差异。
基因型差异也是影响植物性状的重要变异来源。白凤莹等[24]发现,受基因型的影响,不同天然三倍体毛白杨无性系间气孔性状的差异达到了极显著水平,其中存在部分个体表现出类似二倍体的特征。本研究发现,青黑杨全同胞杂种无性系间短枝叶片和气孔各性状均存在极显著差异,其中部分二倍体无性系的叶片和气孔性状表现甚至优于一些三倍体无性系,可见三倍体也并非株株皆优,无性系基因型对性状表现的影响也较大。
对于雌雄异株植物而言,植株性别会对表型产生一定的影响。相关研究表明,多年生植物雄株在叶片数量及叶面积、生长势、树冠体积、生物量等方面都超过雌株[12, 25]。陈珂等[26]发现成年银杏(Ginkgo biloba)雌株叶片长度、叶片宽度、叶面积显著小于雄株,而叶柄长度和气孔密度显著大于雄株。两年生青杨(P. cathayana)雌株叶片总数显著高于雄株,雌株叶片性状较雄株有优势[27];毛白杨雄株叶片长度和叶面积显著大于雌株,而叶片宽度则无显著差异[28]。本研究中,青黑杨全同胞杂种叶片和气孔性状在雌、雄株之间虽存在一定差异,但均未达显著性水平,性状变异与性别之间也不存在显著相关性,且性别效应方差贡献率几乎为0,说明性别对叶片及气孔性状的影响较小。
异源多倍化在植物遗传改良和种质创新中发挥着重要作用,特别是利用2n配子杂交的异源多倍化途径,结合了倍性优势和杂种优势,在杨树多倍体育种已取得了极大的成就[29]。Liao等[11]发现,杨树异源三倍体群体苗期整体具有生长和光合优势,但也存在一些光合效率高但生长较差的个体和光合效率低但生长较好的个体。本研究利用一个包含杂种二倍体和三倍体的青黑杨全同胞群体,证明了杨树经历异源多倍化后,倍性效应和基因型效应均对性状变异产生重要影响,而性别对性状变异的贡献相对较小,并初步明确了倍性、基因型和性别效应对功能叶片和气孔性状的贡献大小,为我们进一步开展多倍体性状变异的遗传解析提供了借鉴。显然,在进行杨树三倍体育种时,对大量变异材料进行“大群体,强选择”是进一步品种选育的必要环节,异源三倍体遗传变异的复杂性也为杨树三倍体选育提供了丰富的选择基础。
-
![]()
图 2 不同果型种子质量和平均单粒质量的差异
不同小写字母表示不同果型在P<0.01水平上差异显著。
Figure 2. Variations in seed yield and average single grain mass for different fruit types
Different lowercase letters mean significant difference among different fruit types at P < 0.01 level.
![]()
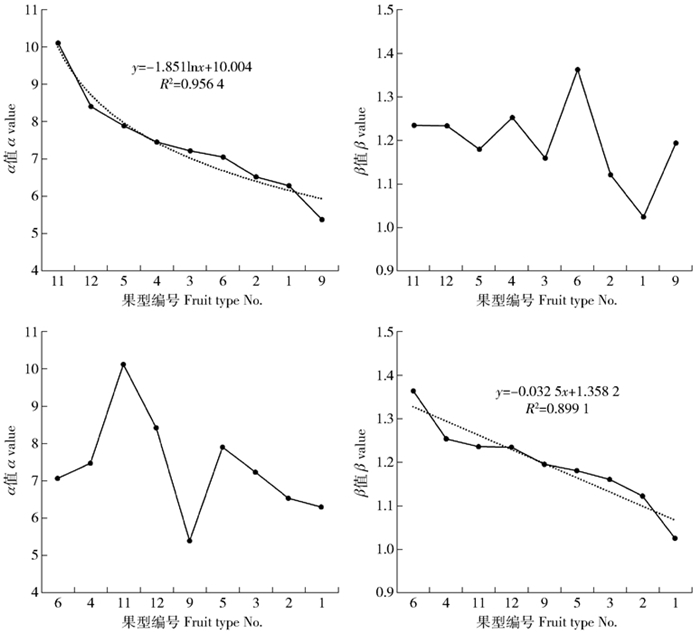
图 5 平均单粒质量威布尔参数变化趋势
Figure 5. Changing trend of Weibull parameters of average single grain mass
![]()
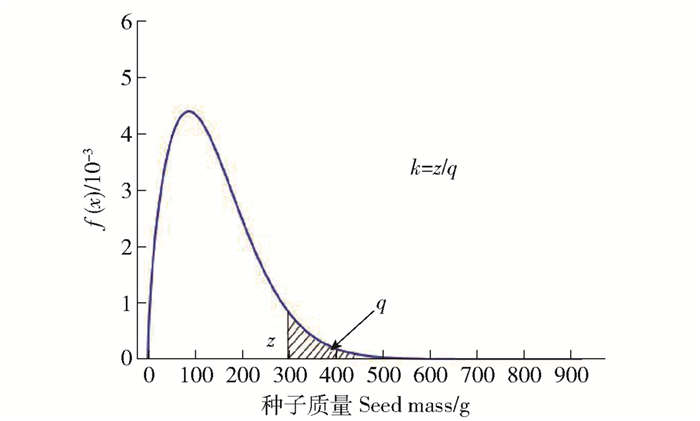
图 6 威布尔分布中k与z和q的关系
z为曲线在某一点的概率密度;q为选择部分的面积,即其所占的概率。下同。
Figure 6. Relationship between k and z and q in Weibull distribution
z means the probability density of curve at a certain point; q means the area of the chosen part, i. e. the probability. The same below.
![]()
图 7 种子质量和平均单粒质量概率累积分布曲线
Figure 7. Probability cumulative distribution curves of seed mass and average single grain mass
表 1 不同果型文冠果种实性状的均值与标准差
Table 1 Mean and standard deviation of phenotypic traits of different fruit types of Xanthoceras sorbifolium
果型
Fruit type株数
Trees number果实质量
Fruit mass/g种子质量
Seed mass/g壳质量
Shell mass/g平均单粒质量
Average single grain mass/g坐果序数
Infructescence number坐果数
Fruit number1 308 236.322±12.697 133.344±7.119 102.978±5.67 0.957±0.011 5.94±0.313 7.92±0.448 2 133 239.394±19.502 123.02±10.194 116.375±9.816 1.037±0.018 6.21±0.53 8.06±0.704 3 64 336.187±35.192 184.276±19.109 151.912±16.286 1.08±0.022 7.17±0.771 8.77±0.937 4 93 323.530±30.358 168.866±15.695 154.664±15.073 1.183±0.021 6.74±0.61 8.76±0.85 5 139 318.627±23.551 177.091±13.266 141.535±10.43 1.088±0.016 7.14±0.50 8.75±0.629 6 38 245.275±32.908 124.968±17.515 120.306±15.659 1.245±0.03 4.29±0.531 4.92±0.636 7 16 221.643±34.755 119.288±18.069 102.355±17.604 1.034±0.048 6.31±1.267 8.00±2.123 8 6 227.62±32.911 127.383±18.788 100.237±15.131 1.345±0.096 4.83±0.872 5.50±1.088 9 67 269.255±24.992 144.764±14.045 124.492±11.315 1.062±0.027 6.25±0.601 8.12±0.792 10 21 325.828±51.86 174.148±28.963 151.68±23.622 1.183±0.050 5.81±0.925 7.19±1.032 11 57 259.291±30.49 134.274±15.947 125.018±14.773 1.184±0.027 5.40±0.572 6.54±0.761 12 109 336.123±32.861 182.877±17.225 153.247±16.315 1.138±0.017 6.94±0.684 8.58±0.855 总体Total 1051 276.929±7.894 150.248±4.279 126.681±3.727 1.068±0.007 6.31±0.176 8.04±0.235 P — 0.002** 0.001** 0.001** 0.000** 0.215 0.312 注:*表示该列内各果型之间在P<0.05水平上差异显著,**表示该列内各果型之间在P<0.01水平上差异显著。Notes: * means significant difference among different fruits at P<0.05 level; ** means significant difference among different fruits at P<0.01 level.  下载: 导出CSV
下载: 导出CSV
表 2 不同果型种实性状的威布尔分布拟合
Table 2 Weibull distribution fitting of different fruit types
果型编号Fruit type No. 果实质量Total mass 种子质量Seed mass 粒数Grain number α β R2 α β R2 α β R2 1 1.744 7 238.907 3 0.900 1 1.609 6 139.911 7 0.923 5 1.519 9 151.782 5 0.941 9 2 1.711 6 249.666 7 0.942 6 1.729 2 130.624 7 0.923 1 1.698 6 133.602 7 0.951 9 3 1.573 7 337.515 3 0.808 4 1.460 4 213.911 7 0.775 6 1.466 3 189.655 7 0.762 2 4 1.433 5 381.180 7 0.949 0 1.469 4 194.908 5 0.969 6 1.376 5 180.380 8 0.884 5 5 1.814 321.281 6 0.923 8 1.745 8 180.106 9 0.943 6 1.752 5 173.967 8 0.951 4 6 2.346 5 225.220 3 0.917 4 2.340 5 112.779 1 0.898 9 2.111 9 96.160 4 0.939 5 9 1.647 7 334.577 3 0.971 0 1.505 3 182.665 0 0.919 3 1.501 0 164.926 8 0.949 3 11 1.080 7 257.100 0 0.971 2 1.107 8 131.864 9 0.974 0 1.075 4 115.099 6 0.957 8 12 1.731 5 310.185 3 0.917 9 1.732 1 161.522 9 0.898 1 1.665 0 156.194 1 0.948 7 果型编号Fruit type No. 单粒质量Single grain mass 坐果序数Fruit ordinal number 坐果数Fruit number α β R2 α β R2 α β R2 1 6.291 3 1.024 5 0.949 7 1.577 9 5.273 1 0.902 0 1.688 0 7.068 4 0.895 2 2 6.526 5 1.120 4 0.920 3 2.465 4 4.147 5 0.910 2 2.200 9 5.675 5 0.870 7 3 7.225 0 1.159 5 0.964 8 1.348 7 7.187 1 0.899 2 1.632 3 8.802 8 0.889 0 4 7.455 6 1.252 0 0.917 3 1.626 5 6.068 5 0.854 5 1.539 6 8.459 9 0.779 1 5 7.892 9 1.179 9 0.943 7 1.594 2 7.424 7 0.967 2 1.743 3 9.209 8 0.963 5 6 7.058 9 1.362 7 0.846 3 1.760 0 4.515 3 0.917 6 1.802 4 5.692 6 0.927 2 9 5.378 7 1.194 0 0.919 6 1.609 3 6.048 7 0.835 5 1.675 8 8.283 6 0.818 7 11 10.113 1 1.234 0 0.811 3 1.390 1 5.896 8 0.905 3 1.575 6 6.343 4 0.914 5 12 8.399 1 1.233 8 0.977 6 1.732 8 5.689 1 0.932 1 1.897 7 6.864 5 0.925 7
下载: 导出CSV
表 3 种实性状的选择概率与选择强度
Table 3 Selection of percentage and intensity of yield traits
q/% 果实质量
Fruit mass种子质量
Seed mass平均单粒质量
Average single grain mass壳质量
Shell massp/g k p/g k p/g k p/g k 90 75 0.002 1 40 0.004 0 0.81 0.821 7 36.0 0.004 4 70 156 0.003 4 84 0.006 3 0.98 2.129 0 73.0 0.007 4 50 233 0.004 5 127 0.008 1 1.10 3.791 9 108.0 0.009 9 40 275 0.005 0 150 0.009 0 1.15 4.735 0 127.0 0.011 1 30 324 0.005 5 178 0.010 0 1.20 5.857 1 148.0 0.012 5 20 386 0.006 2 213 0.011 1 1.26 7.474 1 176.0 0.014 1 10 478 0.007 2 266 0.012 8 1.34 10.166 0 216.0 0.016 4 5 560 0.008 0 313 0.014 1 1.40 12.653 4 251.0 0.018 3 4 585 0.008 2 326 0.014 5 1.42 13.582 8 262.0 0.018 9 3 616 0.008 5 344 0.014 9 1.44 14.566 0 276.0 0.019 6 2 657 0.008 9 368 0.015 6 1.46 15.605 3 294.0 0.020 5 1 725 0.009 5 407 0.016 6 1.51 18.464 9 323.0 0.022 0 0.8 746 0.009 7 419 0.016 9 1.52 19.084 1 331.0 0.022 4 0.6 772 0.009 9 435 0.017 3 1.53 19.719 8 343.0 0.023 0 0.4 809 0.010 2 456 0.017 8 1.55 21.041 9 358.0 0.023 7 0.2 866 0.010 7 490 0.018 6 1.58 23.157 2 383.0 0.024 9 0.1 921 0.011 1 522 0.019 3 1.61 25.439 3 407.0 0.026 1 注:p为选择值,k为选择强度。下同。Notes: p means selection value, k means selection intensity. The same below.
下载: 导出CSV
表 4 不同果型果实质量的选择概率和选择强度
Table 4 Selection of percentage and intensity of fruit mass for different fruit types
q/% 小柱型Small pillar type 小桃型Small peach type 尖柱型Pointed pillar type p/g k p/g k p/g k 90 66 0.002 4 68 0.002 4 82 0.001 8 70 133 0.004 0 138 0.003 9 176 0.002 8 50 194 0.005 3 203 0.005 2 268 0.003 6 40 228 0.006 0 238 0.005 8 320 0.004 0 30 266 0.006 8 279 0.006 5 381 0.004 4 20 315 0.007 7 331 0.007 3 458 0.004 9 10 386 0.008 9 407 0.008 5 574 0.005 6 5 449 0.010 0 475 0.009 5 679 0.006 2 4 468 0.010 3 495 0.009 8 710 0.006 3 3 491 0.010 7 521 0.010 1 750 0.006 5 2 523 0.011 2 555 0.010 6 804 0.006 8 1 574 0.012 0 610 0.011 3 892 0.007 2 q/% 大桃型Big peach type 短柱型Short pillar type 凸桃型Convex peach type p/g k p/g k p/g k 90 80 0.001 8 94 0.001 8 87 0.002 3 70 187 0.002 7 183 0.003 2 146 0.004 7 50 296 0.003 2 263 0.004 3 193 0.006 9 40 360 0.003 5 307 0.004 8 218 0.008 2 30 435 0.003 8 357 0.005 5 245 0.009 6 20 532 0.004 2 419 0.006 2 277 0.011 3 10 683 0.004 7 510 0.007 3 322 0.013 8 5 821 0.005 0 589 0.008 2 360 0.016 1 4 863 0.005 2 613 0.008 5 372 0.016 8 3 915 0.005 3 643 0.008 8 385 0.017 6 2 988 0.005 5 683 0.009 3 404 0.018 8 1 1 107 0.005 7 747 0.010 0 433 0.020 6 q/% 倒卵型Inverted egg type 短桃型Short peach type 柱型Pillar type p/g k p/g k p/g k 90 86 0.002 0 33 0.003 5 86 0.001 9 70 180 0.003 2 100 0.003 8 172 0.003 2 50 269 0.004 1 184 0.004 0 252 0.004 2 40 318 0.004 6 238 0.004 1 296 0.004 7 30 375 0.005 1 306 0.004 2 346 0.005 3 20 448 0.005 7 400 0.004 3 409 0.006 0 10 556 0.006 6 557 0.004 4 503 0.006 9 5 652 0.007 3 711 0.004 5 585 0.007 8 4 681 0.007 5 759 0.004 5 610 0.008 0 3 718 0.007 8 822 0.004 5 641 0.008 3 2 767 0.008 1 909 0.004 5 683 0.008 7 1 846 0.008 6 1 057 0.004 6 750 0.009 3
下载: 导出CSV
表 5 不同果型种子质量的选择概率与选择强度
Table 5 Selection of percentage and intensity of seed mass for different fruit types
q/% 小柱型Small pillar type 小桃型Small peach type 尖柱型Pointed pillar type p/g k p/g k p/g k 90 36 0.004 4 37 0.004 6 47 0.003 2 70 75 0.007 0 73 0.007 7 107 0.004 7 50 112 0.008 9 107 0.010 2 167 0.005 8 40 134 0.009 9 125 0.011 4 202 0.006 4 30 158 0.011 0 146 0.012 8 244 0.006 9 20 189 0.012 3 173 0.014 5 297 0.007 6 10 236 0.014 1 213 0.016 9 380 0.008 5 5 278 0.015 5 247 0.018 8 454 0.009 2 4 290 0.015 9 258 0.019 4 477 0.009 5 3 306 0.016 5 271 0.020 1 506 0.009 7 2 327 0.017 2 288 0.021 0 545 0.010 1 1 362 0.018 3 317 0.022 5 610 0.010 6 q/% 大桃型Big peach type 短柱型Short pillar type 凸桃型Convex peach type p/g k p/g k p/g k 90 43 0.003 5 51 0.003 4 44 0.004 7 70 98 0.005 2 101 0.005 6 74 0.009 6 50 153 0.006 4 147 0.007 5 97 0.013 8 40 185 0.007 0 172 0.008 4 110 0.016 3 30 222 0.007 6 201 0.009 4 123 0.019 0 20 270 0.008 4 238 0.010 7 139 0.022 4 10 345 0.009 4 291 0.012 5 162 0.027 6 5 412 0.010 2 339 0.013 9 181 0.032 0 4 433 0.010 5 353 0.014 4 187 0.033 5 3 459 0.010 8 371 0.014 9 194 0.035 2 2 494 0.011 1 394 0.015 6 203 0.037 4 1 552 0.011 7 433 0.016 8 218 0.041 1 q/% 倒卵型Inverted egg type 短桃型Short peach type 柱型Pillar type p/g k p/g k p/g k 90 42 0.003 8 18 0.006 5 45 0.003 5 70 93 0.005 7 53 0.007 4 90 0.005 9 50 144 0.007 1 96 0.007 9 132 0.007 8 40 173 0.007 8 123 0.008 1 155 0.008 8 30 208 0.008 6 157 0.008 3 181 0.009 9 20 252 0.009 5 204 0.008 5 214 0.011 2 10 319 0.010 7 281 0.008 8 262 0.013 0 5 380 0.011 7 356 0.009 1 305 0.014 5 4 398 0.011 9 380 0.009 1 318 0.014 9 3 421 0.012 3 410 0.009 2 334 0.015 5 2 453 0.012 7 452 0.009 3 356 0.016 2 1 505 0.013 5 524 0.009 5 391 0.017 4
下载: 导出CSV
表 6 不同果型平均单粒质量的选择概率与选择强度
Table 6 Selection of percentage and intensity of average single grain mass for different fruit types
q/% 小柱型Small pillar type 小桃型Small peach type 尖柱型Pointed pillar type p/g k p/g k p/g k 90 0.73 0.938 7 0.80 0.817 7 0.86 0.902 1 70 0.88 2.555 0 0.97 2.400 8 1.02 2.639 7 50 0.98 4.543 7 1.07 4.151 4 1.11 4.490 7 40 1.02 5.626 9 1.12 5.355 8 1.16 5.922 4 30 1.07 7.266 2 1.16 6.513 1 1.20 7.327 1 20 1.12 9.273 1 1.22 8.627 0 1.25 9.466 8 10 1.18 12.251 7 1.28 11.272 4 1.31 12.704 3 5 1.23 15.288 3 1.34 14.548 3 1.36 16.068 7 4 1.24 15.963 2 1.35 15.163 2 1.37 16.824 2 3 1.26 17.385 4 1.37 16.456 9 1.39 18.424 8 2 1.28 18.908 6 1.39 17.839 7 1.41 20.151 2 1 1.32 22.280 3 1.43 20.891 4 1.44 22.994 4 q/% 大桃型Big peach type 短柱型Short pillar type 凸桃型Convex peach type p/g k p/g k p/g k 90 0.94 0.838 7 0.90 0.907 9 1.00 0.784 2 70 1.10 2.337 1 1.05 2.656 3 1.19 2.271 9 50 1.20 4.118 8 1.14 4.706 9 1.30 3.898 5 40 1.25 5.372 3 1.18 5.982 3 1.36 5.134 7 30 1.29 6.594 4 1.22 7.542 2 1.41 6.400 4 20 1.34 8.445 1 1.26 9.437 5 1.47 8.253 2 10 1.41 11.760 1 1.32 13.037 9 1.54 10.960 9 5 1.46 14.750 0 1.37 16.878 6 1.60 13.837 5 4 1.47 15.419 2 1.38 17.752 9 1.62 14.926 2 3 1.49 16.834 8 1.39 18.665 5 1.64 16.085 5 2 1.51 18.358 6 1.41 20.611 7 1.66 17.319 0 1 1.55 21.758 2 1.44 23.855 2 1.70 20.023 9 q/% 倒卵型Inverted egg type 短桃型Short peach type 柱型Pillar type p/g k p/g k p/g k 90 0.80 0.751 2 1.00 0.893 7 0.95 0.886 4 70 1.00 2.018 0 1.12 2.535 2 1.10 2.650 7 50 1.13 3.463 8 1.20 4.780 2 1.19 4.767 6 40 1.19 4.353 0 1.23 5.997 7 1.23 6.101 3 30 1.25 5.408 8 1.27 8.047 5 1.27 7.746 2 20 1.31 6.652 1 1.30 9.972 2 1.32 10.330 9 10 1.40 8.917 7 1.35 14.102 5 1.37 13.630 2 5 1.47 11.058 2 1.39 18.437 7 1.42 17.804 1 4 1.49 11.737 0 1.40 19.691 7 1.43 18.759 9 3 1.52 12.815 0 1.41 21.021 1 1.44 19.759 7 2 1.55 13.967 8 1.42 22.429 7 1.46 21.898 4 1 1.60 16.065 3 1.45 27.173 7 1.49 25.481 0
下载: 导出CSV
表 7 不同果型壳质量的选择概率与选择强度
Table 7 Selection of percentage and intensity of shell mass for different fruit types
q/% 小柱型Small pillar type 小桃型Small peach type 尖柱型Pointed pillar type p/g k p/g k p/g k 90 41 0.004 0 45 0.003 7 58 0.003 0 70 87 0.006 3 88 0.006 4 114 0.005 2 50 130 0.008 1 127 0.008 6 165 0.006 9 40 154 0.009 0 149 0.009 8 193 0.007 8 30 183 0.010 1 173 0.011 0 225 0.008 8 20 218 0.011 2 203 0.012 5 264 0.010 0 10 272 0.012 8 248 0.014 7 322 0.011 7 5 320 0.014 2 287 0.016 5 373 0.013 2 4 335 0.014 6 299 0.017 0 389 0.013 6 3 353 0.015 1 313 0.017 6 408 0.014 1 2 378 0.015 7 333 0.018 5 433 0.014 8 1 418 0.016 7 365 0.019 9 474 0.015 9 q/% 大桃型Big peach type 短柱型Short pillar type 凸桃型Cenvex peach type p/g k p/g k p/g k 90 56 0.003 0 58 0.003 1 68 0.003 5 70 116 0.004 9 113 0.005 4 110 0.007 2 50 173 0.006 4 161 0.007 3 143 0.010 7 40 204 0.007 1 188 0.008 3 160 0.012 6 30 240 0.007 9 217 0.009 4 178 0.014 8 20 286 0.008 9 254 0.010 7 200 0.017 6 10 354 0.010 3 309 0.012 7 231 0.021 9 5 415 0.011 4 356 0.014 3 257 0.025 6 4 433 0.011 8 370 0.014 7 264 0.026 7 3 456 0.012 2 388 0.015 3 273 0.028 0 2 487 0.012 7 412 0.016 1 286 0.030 1 1 537 0.013 6 450 0.017 4 305 0.033 1 q/% 倒卵型Inverted egg type 短桃型Short peach type 柱型Pillar type p/g k p/g k p/g k 90 61 0.003 4 38 0.004 0 58 0.003 1 70 107 0.006 7 85 0.005 9 109 0.005 6 50 144 0.009 6 133 0.007 3 154 0.007 7 40 164 0.011 2 161 0.008 1 178 0.008 7 30 186 0.013 0 193 0.008 8 206 0.010 0 20 212 0.015 2 234 0.009 7 239 0.011 4 10 249 0.018 4 298 0.010 9 289 0.013 6 5 281 0.021 2 356 0.011 8 331 0.015 3 4 291 0.022 1 373 0.012 1 344 0.015 9 3 302 0.023 1 395 0.012 5 360 0.016 6 2 318 0.024 5 425 0.012 9 381 0.017 4 1 342 0.026 8 475 0.013 6 415 0.018 8
下载: 导出CSV
-
[1] 徐东翔.文冠果生物学[M].北京:科学出版社, 2010. XU D X. Xanthoceras sorbifolia biology[M]. Beijing: Science Press, 2010.
[2] 牟洪香.木本能源植物文冠果(Xanthoceras sorbifolia Bunge)的调查与研究[D].北京: 中国林业科学研究院, 2006. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y962144 MOU H X. Investigation and research on woody energy plant Xanthoceras sorbifolia Bunge[D]. Beijing: Chinese Academy of Forestry, 2006. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y962144
[3] 敖妍, 段劼, 于海燕, 等.文冠果研究进展[J].中国农业大学学报, 2012, 17(6): 197-203. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=zgnydxxb201206025 AO Y, DUAN J, YU H Y, et al. Research progress on Xanthoceras sorbifolia Bunge[J]. Journal of China Agricultural University, 2012, 17(6): 197-203. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=zgnydxxb201206025
[4] 侯元凯, 黄琳, 高巍, 等.文冠果幼树生长性状的相关性分析[J].经济林研究, 2012, 30(2): 52-55, 117. http://d.old.wanfangdata.com.cn/Periodical/jjlyj201202011 HOU Y K, HUANG L, GAO W, et al. Correlation of growth characters of young tree in Xanthoceras sorbifolia[J]. Nonwood Forest Research, 2012, 30(2): 52-55, 117. http://d.old.wanfangdata.com.cn/Periodical/jjlyj201202011
[5] 张萍, 王连军, 彭华.文冠果优良类型选择[J].中国林副特产, 2012(2): 10-12. doi: 10.3969/j.issn.1001-6902.2012.02.004 ZHANG P, WANG L J, PENG H. Excellent type selection of Xanthoceras sorbifolium Bunge[J]. Forest By-Product and Speciality in China, 2012(2): 10-12. doi: 10.3969/j.issn.1001-6902.2012.02.004
[6] 白金友, 李翠舫.文冠果优树无性系鉴定的研究[J].辽宁林业科技, 1988(3): 2-8. http://www.cnki.com.cn/Article/CJFDTotal-LNLK198803001.htm BAI J Y, LI C F. Identification of selected Xanthoceras sorbifolium Bunge excellent clones[J]. Liaoning Forestry Science and Technology, 1988(3): 2-8. http://www.cnki.com.cn/Article/CJFDTotal-LNLK198803001.htm
[7] 刘克武, 张海林, 张顺捷, 等.文冠果优良果型选择[J].中国林副特产, 2008(3): 15-18. doi: 10.3969/j.issn.1001-6902.2008.03.007 LIU K W, ZHANG H L, ZHANG S J, et al. Excellent fruit types selection of Xanthoceras sorbifolium Bunge[J]. Forest By-Product and Speciality in China, 2008(3): 15-18. doi: 10.3969/j.issn.1001-6902.2008.03.007
[8] 徐欢.文冠果花蕾期和盛花期花性状多尺度分析[D].北京: 北京林业大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10022-1014324798.htm XU H. Multiple-scale analysis on floral characteristics of Xanthoceras sorbifolium Bunge in bud and peak flowering stages[D]. Beijing: Beijing Forestry University, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10022-1014324798.htm
[9] 徐欢, 杨韫嘉, 樊简, 等.文冠果花性状多尺度分析[J].东北林业大学学报, 2014, 42(9): 126-137. doi: 10.3969/j.issn.1000-5382.2014.09.027 XU H, YANG Y J, FAN J, et al. Multiple scales analysis on floral characteristics of Xanthoceras sorbifolium Bunge[J]. Journal of Northeast Forestry University, 2014, 42(9): 126-137. doi: 10.3969/j.issn.1000-5382.2014.09.027
[10] 杨韫嘉.文冠果种实性状变异与根孽幼苗可塑性分析[D].北京: 北京林业大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10022-1014324796.htm YANG Y J. Variation of seeds, fruits characteristics and analysis of root cuttings plasticity in Xanthoceras sorbifolium Bunge[D]. Beijing: Beijing Forestry University, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10022-1014324796.htm
[11] 乐也, 王青, 杨韫嘉, 等.植物性状整合的不同软件PCA比较分析[J].中南林业科技大学学报, 2015, 35(9): 59-64. http://d.old.wanfangdata.com.cn/Periodical/znlxyxb201509010 YUE Y, WANG Q, YANG Y J, et al. Comparative analysis on plant traits by using two soft-wares of PCA method[J]. Journal of Central South University of Forestry & Technology, 2015, 35(9): 59-64. http://d.old.wanfangdata.com.cn/Periodical/znlxyxb201509010
[12] PIGLIUCCI M, MURREN C J, SCHLICHTING C D. Phenotypic plasticityand evolution by genetic assimilation[J]. Journal of Experimental Biology, 2006, 209(12): 2362-2367. doi: 10.1242/jeb.02070
[13] PUTENIKHIN V P. Phenotype analysis of Picea obovata in the southern Urals population structure[J]. Lesovedenie, 1997, 6: 37-49.
[14] 陈思羽, 杨辉, 韩姣, 等.长白山区核桃楸结实性状种源变异分析[J].北京林业大学学报, 2015, 37(12): 32-40. doi: 10.13332/j.1000-1522.20150278 CHEN S Y, YANG H, HAN J, et al. Provenance variation of seed traits of Juglans mandshurica in Changbai mountains, northeastern China[J]. Journal of Beijing Forestry University, 2015, 37(12): 32-40. doi: 10.13332/j.1000-1522.20150278
[15] 王文杰, 李文馨, 许慧男, 等.不同生境白屈菜(Chelidonium majus)生活史型特征及其与不同器官单宁、黄酮、生物碱含量的关系[J].生态学报, 2008, 28(11): 5228-5237. doi: 10.3321/j.issn:1000-0933.2008.11.004 WANG W J, LI W X, XU H N, et al. Characters of life cycle forms of Chelidonium majus populations in different habitats and their correlation to the contents of tannins, flavones and alkaloids in different organs[J]. Acta Ecologica Sinica, 2008, 28(11): 5228-5237. doi: 10.3321/j.issn:1000-0933.2008.11.004
[16] 于景华, 李德文, 庞海河, 等. UV-B辐射对南方红豆杉生活史型和紫杉烷类含量的影响[J].生态学报, 2011, 31(1): 75-81. http://d.old.wanfangdata.com.cn/Periodical/stxb201101009 YU J H, LI D W, PANG H H, et al. Effects of supplementary UV-B radiation on life cycle forms and the accumulation of taxanes of Taxus chinensis var. mairei[J]. Acta Ecologica Sinica, 2011, 31(1): 75-81. http://d.old.wanfangdata.com.cn/Periodical/stxb201101009
[17] 张振, 张含国, 周宇, 等.红松多无性系群体的种实性状变异研究[J].北京林业大学学报, 2015, 37(2): 67-78. doi: 10.13332/j.cnki.jbfu.2015.02.020 ZHANG Z, ZHANG H G, ZHOU Y, et al. Variation of seed characters in Korean pine (Pinus koraiensis) multi-clonal populations[J]. Journal of Beijing Forestry University, 2015, 37(2): 67-78. doi: 10.13332/j.cnki.jbfu.2015.02.020
[18] 黄雨洁.云南松针叶与油松种实性状的种群变异研究[D].北京: 北京林业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10022-1015319750.htm HUANG Y J. Population genetic variation of Pinus yunnanensis needle and Pinus tabuliformis cone and seed traits[D]. Beijing: Beijing Forestry University, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10022-1015319750.htm
[19] 师玮, 潘伯荣, 段士民, 等.蒙古沙拐枣(Calligonum mongolicum)与其相关种的果实形态差异性分析[J].中国沙漠, 2011, 31(1): 121-128. http://d.old.wanfangdata.com.cn/Periodical/zgsm201101019 SHI W, PAN B R, DUAN S M, et al. Difference of fruit morphological characters of Calligonum mongolicum and related species[J]. Journal of Desert Research, 2011, 31(1): 121-128. http://d.old.wanfangdata.com.cn/Periodical/zgsm201101019
[20] ZHAO Z, HE Y, WANG M, et al. Variations of flower size and reproductive traits in self-incompatible trollius ranunculoides (Ranunculaceae) among local habitats at alpine meadow[J]. Plant Ecology, 2007, 193(2): 241-251. doi: 10.1007/s11258-006-9262-9
[21] GRANDILLO S, KU H M, TANKSLEY S D. Identifying the loci responsible for natural variation in fruit size and shape in tomato[J]. Theoretical and Applied Genetics, 1999, 99(6): 978-987. doi: 10.1007/s001220051405
[22] CANTÍN C M, GOGORCENA Y, MORENO M Á. Phenotypic diversity and relationships of fruit quality traits in peach and nectarine[Prunus persica (L.) Batsch] breeding progenies[J]. Euphytica, 2010, 171(2): 211-226. doi: 10.1007/s10681-009-0023-4
[23] SHENG Z, JIN H, ZHANG C F, et al. Genetic analysis of fruit shape traits at different maturation stages in sponge gourd[J]. Biomedicine & Biotechnology, 2007, 8(5): 338-344. doi: 10.1631/jzus.2007.B0338
[24] 柴春山, 芦娟, 蔡国军, 等.文冠果人工种群的果实表型多样性及其变异[J].林业科学研究, 2013, 26(2): 181-191. doi: 10.3969/j.issn.1001-1498.2013.02.009 CHAI C S, LU J, CAI G J, et al. Fruit phenotypic diversity and variation of Xanthoceras sorbifolia artificial population[J]. Forest Research, 2013, 26(2): 181-191. doi: 10.3969/j.issn.1001-1498.2013.02.009
[25] LEGENDRE L, LEGENDRE P. Numerical ecology[M]. Amserdam: Elsevier Science, 1998: 194-198.
[26] LAWLESS J F. Statistical models and methods for lifetime data[M]. New York: A John Wiley & Sons, Inc., 1982.
[27] 刘兴聪, 牛玉玺.威布尔(Weibull)分布在洮河林区复层异龄林直径结构中的应用[J].甘肃农大学报, 1989(1): 81-91. http://www.cqvip.com/QK/94944X/198901/3001435481.html LIU X C, NIU Y X. Application of Weibull distribution in diameter structure of double layer uneven-aged forest in Taohe Forest Region[J]. Journal of Gansu Agricultural University, 1989(1): 81-91. http://www.cqvip.com/QK/94944X/198901/3001435481.html
[28] VAN BOEKEL M A. On the use of the Weibull model to describe thermal inactivation of microbial vegetative cells[J]. International Journal of Food Microbiology, 2002, 74(1-2): 139-159. doi: 10.1016/S0168-1605(01)00742-5
[29] SMITH R L. Weibull regression models for reliability data[J]. Reliability Engineering and System Safety, 1991, 34(1), 55-76. doi: 10.1016/0951-8320(91)90099-S
[30] 郭昭洋.柳杉人工纯林胸径、树高威布尔分布的拟合[J].江西林业科技, 1997(增刊1): 25-26. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=jxlykj1997z1008 GUO Z Y. Weibull distribution fitting of DBH and tree height in Gryptomeria fortunei Hooibrenk plantation[J]. Jiangxi Forestry Science and Technology, 1997(Suppl. 1): 25-26. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=jxlykj1997z1008
[31] 郭丽虹, 李荷云.桤木人工林林分胸径与树高的威布尔分布拟合[J].江西林业科技, 2000(增刊1): 26-27. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=jxlykj2000z1009 GUO L H, LI H Y. Weibull distribution of DBH and tree height in Alnus cremastogyne plantation[J]. Jiangxi Forestry Science and Technology, 2000(Suppl. 1): 26-27. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=jxlykj2000z1009
[32] 张鉴法.文冠果试种初报[J].新疆林业, 1981(增刊1): 2-7. http://www.cnki.com.cn/Article/CJFDTotal-XJLY1981S1001.htm ZHANG J F. Reported of Xanthoceras sorbifolium plant experimentally[J]. Forestry of Xinjiang, 1981(Suppl. 1): 2-7. http://www.cnki.com.cn/Article/CJFDTotal-XJLY1981S1001.htm
[33] 李付鹏, 王华, 伍宝朵, 等.可可果实主要农艺性状相关性及产量因素的通径分析[J].热带作物学报, 2014, 35(3): 448-453. doi: 10.3969/j.issn.1000-2561.2014.03.006 LI F P, WANG H, WU B D, et al. Correlation analysis among main pod agronomic traits and path analysis on yield factor in cacao[J]. Chinese Journal of Tropical Crops, 2014, 35(3): 448-453. doi: 10.3969/j.issn.1000-2561.2014.03.006
[34] FENG C Y, WANG H Y, LU N J, et al. Log-transformation and its implications for data analysis[J]. Shanghai Archives of Psychiatry, 2014, 26(2): 105-109. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=shjsyx201402009
[35] FAIRFIELD H. A discriminant function for plant selection[J]. Annals of Human Genetics, 1936, 7(3): 240-250. doi: 10.1111-j.1469-1809.1936.tb02143.x/
[36] 马育华.植物育种的数量遗传学基础[M].南京:江苏科学技术出版社, 1982. -
期刊类型引用(4)
1. 赵钰婷,陈冬瑶,杨柳,李晶楠,宁广亮,姜静. 白桦四倍体×紫雨桦二倍体杂交种子活力及杂种子代生长特性分析. 温带林业研究. 2025(01): 1-8 .  百度学术
百度学术
2. 马鸿文,任宇昕,龙羿辛,王楠,冯祥元,俞天泉,华晓琴,王君. 青黑杨杂种全同胞二倍体与三倍体长枝叶性状变异研究. 北京林业大学学报. 2024(01): 27-34 . 本站查看
3. 王舒扬,田力,周顺陶,储月娥,梅迪,袁佳秋,余延浩,洑香香. 多倍化对青钱柳叶形态、光合性能和次生代谢产物积累的影响. 林业科学. 2024(08): 120-131 . 百度学术
4. 卞瑶,耿丽妍,冯彦博,陈福燕,张雯,王宏伟. 不同进化历程下的异源四倍体小麦核型进化及其稳定性分析. 辽宁师范大学学报(自然科学版). 2023(03): 363-369 . 百度学术
其他类型引用(5)








计量
- 文章访问数: 5304
- HTML全文浏览量: 584
- PDF下载量: 64
- 被引次数: 9
