
Geographic variation of growth traits in early stage for Melia azedarach among different provenances
-
摘要:目的研究苦楝种源间生长性状的差异,揭示其地理变异规律,分析其气候生态学基础,为苦楝种子调拨提供理论依据,为广东省营造苦楝速生用材林提供技术参考。方法对15个省(自治区)的62个种源的树高、胸径、枝下高、干型、冠幅等6个性状进行观测统计,采用SPSS 20.0、SAS 9.0、Surfer 13.0与EXCEL 2011等数据分析软件对观测数据进行描述性统计分析、方差分析、Pearson相关分析、典型相关分析与趋势面分析。结果大部分苦楝种源在四会试验点的生长表现较好,且各性状在种源内及种源间均存在显著或极显著差异,其中冠幅的变幅最大,优劣单株相差23倍,在主要性状树高与胸径的表现上也分别相差8倍与12倍。苦楝的地理变异受纬度、经度和海拔的多重控制,以纬向变异为主,并具有一定的气候生态学基础,温度是制约苦楝生长的主导因子,光照、降水、气压等气候因子对苦楝各性状也有着不同程度的影响。聚类分析将62个种源分为5类,类别之间差异明显,类别之中符合物候区划。结论苦楝在种源间的变异存在明显的地理变异模式,由东向西,由北向南,苦楝的生长量变大,形质表现变佳,随着海拔上升,保存率下降。气候生态学基础则表现为来自极端气温与气压温和、温度高、降水多的种源更具生长优势。纬度是引起温度变化的主要因素,所以纬向变异这一模式与温度随纬度增加而递减这一规律密切相关。本研究后续将对四会种源试验林进行长期跟踪观测,并辅以分子遗传学分析,更全面探索苦楝地理变异规律。Abstract:ObjectiveThis paper aims to provide theoretical basis and technical reference for seed allocation and silviculture of fast growing timber plantation in Melia azedarach. The variance of growth traits among various provenances was estimated. Furthermore, the pattern of geographic variation was studied to reveal the underlying climatic and ecological foundations.MethodSix traits, including tree height, DBH, clear bole height, stem form, crown and survival rate of 62 provenances (15 provinces in China), were investigated, and the descriptive statistics analysis, variance analysis, Pearson correlation analysis, canonical correlation analysis and trend surface analysis were performed using SPSS 20.0, SAS 9.0, Surfer 13.0 and EXCEL 2011, respectively.ResultThe results showed that most provenances had well growth in Sihui Experimental Site of Southern China, and each trait had significant differences or highly significant differences among intra-provenances and inter-provenances. Tree height and DBH of the best one were 8 and 12 times of the worst one, respectively, and for crown it was 23 times. The geographic variation of Melia azedarach was influenced by multiple factors, including longitude, latitude and altitude, among which latitude plays the major role. The growth of Melia azedarach was mainly regulated by temperature, but also was affected by sunshine, precipitation and air pressure. The 62 provenances could be divided into 5 types. There were obvious differences between groups, and they followed the phenological division within a group.ConclusionThe variance of traits had obvious geographic variation patterns and certain eco-climate foundation. In the south-to-north distribution and the west-to-east distribution, growth gradually became better and stem-form quality was being higher. In the meantime, survival rate fell as altitude increased. The provenances with mildly extreme temperature and pressure, higher mean minimum temperature and more precipitation had growth advantages. Latitude is a key factor in temperature change, so the pattern of zonal variation and the regulation of temperature decrease are inseparable from the increase of latitude. To explore the Melia azedarach regularity of geographic variance compressively with the evidences of molecular genetic analyses, the further studies will pay continuous attention to the growth state in Sihui Experimental Site of southern China.
-
Keywords:
- Melia azedarach /
- provenance /
- geographic variation
-
苦楝(Melia azedarach)为楝科(Meliaceae)楝属(Melia)模式植物,其分布范围包括热带、亚热带及温带,在我国水平分布于18°~40°N,从东部的台湾、沿海各省,到西部的四川、云南保山,从北部的山西运城、河北保定、陕西渭南、甘肃陇南到南部的海南三亚,垂直分布于2 100 m以下,为常绿阔叶林、落叶林,是道路、农田、溪谷两旁的常见树种[1-4]。苦楝是速生用材树种,因其材性优良、花纹独特、光泽亮丽、抗虫耐腐,在家具、造船、乐器等多方面有着广泛的应用[5-7]。苦楝具有较好的抗虫作用,可作为优良的植物源农药,有良好的细胞毒作用,可以抗肿瘤、抗病毒及抗真菌等[8-11]。也是园林绿化和矿区植被恢复树种;此外,在农业与工业上还有多种用途[12-16]。
国内对苦楝种源遗传多样性及地理变异的研究多集中于叶片、果实、种子的表型性状与播种品质,苗木或林木的生长性状与生长节律等方面,均表明不同种源因适应不同生长环境而产生丰富的遗传差异,且地理变异趋势明显[1, 3, 6, 17-22],如程诗明等通过物候因子与地理因子的相关性分析,发现苦楝展叶期、叶芽开放期、展叶盛期自南向北,自西向东呈推迟的趋势,具有梯度变异[3];廖柏勇等通过趋势面分析得出苦楝果核和种子性状受到经纬度双重控制的结论[2]。
研究地理变异规律,分析其气候生态学基础,可为种子调拨等育种程序提供参考依据[23]。许多树种的生长性状或种实特征均能反映地理分布规律,如麻风树(Jatropha curcas)、无患子(Sapindus spp.)、乐昌含笑(Michelia chapensis)等,其地理变异规律可指导种源的初步区划及进一步的选优育种,给育种工作及经济价值的开发提供理论支撑[24-26]。本研究遵循布点均匀原则,采集全分布区内15个省(自治区)的62个种源,在广东四会布置种源试验,观测4年生苦楝的生长性状,统计保存率,进行数据分析,旨在研究苦楝种源间的性状差异,探讨与其相关的地理气候因子,分析地理变异规律及气候生态学基础,为加速遗传改良进程,进行速生材用的优良种源选择及广东地区引种提供理论依据。
1. 材料与方法
1.1 试验材料及设计
1.1.1 参试种源
采种母树均为当地自然分布,生长正常,无明显病虫害且发育成熟的苦楝,由于苦楝少有人工种植,能保证母树为当地种源。每个采种点要求母树至少30株以上,母树间距离100 m以上。每一采种点的母树种子育苗成功后,取等量苗木混合作为一个种源,参试种源共有62个,其地理信息详见表 1、图 1。
表 1 苦楝各种源编号及海拔Table 1. No. and elevation of Melia azedarach provenances编号No. 种源地Provenance 海拔Elevation/m 编号No. 种源地Provenance 海拔Elevation/m 编号No. 种源地Provenance 海拔Elevation/m 101 福建连城Liancheng, Fujian 351 422 广东阳山Yangshan, Guangdong 93 844 贵州黎平Liping, Guizhou 650 102 福建永安Yongan, Fujian 373 523 海南海口Haikou, Hainan 129 845 贵州罗甸Luodian, Guizhou 386 103 福建漳平Zhangping, Fujian 219 524 海南屯昌Tunchang, Hainan 160 846 贵州荔波Libo, Guizhou 467 204 江西赣县Ganxian, Jiangxi 115 525 海南五指山Wuzhishan, Hainan 341 847 贵州德江Dejiang, Guizhou 851 205 江西于都Yudu, Jiangxi 145 526 海南陵水Lingshui, Hainan 79 848 贵州麻江Majiang, Guizhou 1 048 206 江西遂川Suichuan, Jiangxi 103 527 海南琼海Qionghai, Hainan 32 849 贵州贵阳Guiyang, Guizhou 1 205 307 湖南东安Dongan, Hunan 252 628 广西桂林Guilin, Guangxi 166 850 贵州石阡Shiqian, Guizhou 496 308 湖南浏阳Liuyang, Hunan 137 629 广西融安Rongan, Guangxi 226 952 浙江绍兴Shaoxing, Zhejiang 181 309 湖南通道Tongdao, Hunan 442 630 广西三江Sanjiang, Guangxi 240 959 浙江临安Lin’an, Zhejiang 47 310 湖南炎陵Yanling, Hunan 192 631 广西钦州Qinzhou, Guangxi 250 1053 重庆Chongqing 183 311 湖南龙山Longshan, Hunan 779 632 广西梧州Wuzhou, Guangxi 31 1154 安徽黄山Huangshan, Anhui 306 412 广东仁化Renhua, Guangdong 196 633 广西凭祥Pingxiang, Guangxi 255 1155 安徽利辛Lixin, Anhui 38 413 广东郁南Yunan, Guangdong 22 634 广西靖西Jingxi, Guangxi 764 1256 江苏泰兴Taixing, Jiangsu 7 414 广东博罗Boluo, Guangdong 13 735 云南富宁Funing, Yunnan 732 1257 江苏南京Nanjing, Jiangsu 22 415 广东开平Kaiping, Guangdong 8 736 云南广南Guangnan, Yunnan 1 079 1258 江苏姜堰Jiangyan, Jiangsu 8 416 广东清远Qingyuan, Guangdong 73 737 云南开远Kaiyuan, Yunnan 1 122 1359 河南内乡Neixiang, Henan 178 417 广东恩平Enping, Guangdong 17 739 云南勐腊Mengla, Yunnan 1 005 1461 陕西安康Ankang, Shaanxi 422 418 广东饶平Raoping, Guangdong 20 740 云南罗平Luoping, Yunnan 1 415 1462 陕西蒲城Pucheng, Shaanxi 488 419 广东五华Wuhua, Guangdong 123 741 云南麻栗坡Malipo, Yunnan 1 113 1463 陕西南郑Nanzheng, Shaanxi 716 420 广东东源Dongyuan, Guangdong 57 842 贵州兴义Xingyi, Guizhou 1 217 1564 甘肃舟曲Zhouqu, Gansu 1 354 421 广东茂名Maoming, Guangdong 162 843 贵州册亨Ceheng, Guizhou 993 1.1.2 试验设计
2013年3月在华南农业大学增城试验基地育苗,同年6月20日在四会试验点定植,该试验点设在广东省四会市迳口镇下寮村温氏新围猪场(112°45′02″ E、23°30′06″ N),属亚热带季风气候区,年均气温为21.3 ℃,极端最高温为38.5 ℃,极端最低温为-1.2 ℃,年均降雨量为1 803.6 mm,年均日照为1 702.3 h。随机区组设计,单株小区,45次重复。株行距为2 m×3 m,栽植穴规格为40 cm×40 cm,每穴施基肥3 kg。
1.2 试验林调查
于2016年11月进行外业调查,观测树高、胸径、枝下高、干型和冠幅5个性状,并统计保存率。其中干型分为4个等级,主干明显且通直为4级,主干明显但稍弯曲为3级,主干弯曲程度大为2级,主干不明显且多分枝为1级。
1.3 统计分析
气象因子数据为各采种点或相近气象台站点在中国气象数据网中国气象资料年值数据集中,1951—2012年的数据均值。地理因子数据为采种点GPS记录。
采用EXCEL 2011进行数据的初步整理与趋势面方程拟合;采用SPSS 20.0进行Pearson相关分析;采用SAS 9.0进行描述性统计、方差分析、典型相关分析及聚类分析;采用Surfer 13.0软件进行趋势面作图。
方差分析模型采用:
Yjk=μ+Bj+Pk+Rij 式中:Yij为j重复k种源某性状的测量值;μ为整体均值;Bj为j区组的效应值;Pk为k种源效应值;Rij为残差效应值。
趋势面分析是采用多元回归的方法建立某一性状与地理因子的曲面方差。若试验包含n个种源,测定各种源均值记为Zi,当x与y的最高系数为2时,公式如下:
Zi=a1+a2x+a3y+a4x2+a5y2+a6xy+εi 式中:a1~a6为回归系数,x为纬度,y表示经度,εi为随机误差;拟合精度公式为C=U/S×100,表示回归平方和U占总平方和S的比值百分数[27]。
2. 结果与分析
2.1 种源性状变异分析
对62个种源的树高、胸径、枝下高、干型、冠幅及保存率进行描述性统计与方差分析,结果见表 2。从各性状均值来看,平均树高为9.24 m,平均胸径为11.83 cm,说明各种源在四会试验点的生长较为迅速;枝下高、干型与冠幅的均值分别为3.12、2.41、3.14 m,说明大部分种源在四会试验点的干型表现较好;保存率为0.65,较高存活率保证有大量的遗传数据可用于分析。从变异幅度来看,各性状均有较大的变异幅度,树高与胸径的变异幅度分别为2.00~15.90 m与2.30~28.20 m,说明树高最高的单株较最矮的单株高87.42%,胸径最大的单株较最小的单株大91.84%;其中冠幅的变异幅度最大,为0.40~9.20 m,相差近22倍。从变异系数来看,干型与枝下高的变异系数较大,分别为48.25%与46.98%;其次是胸径与冠幅,分别为33.92%与33.54%;树高与保存率的变异系数则较小,分别为22.46%与21.05%。
表 2 苦楝生长性状种源变异分析Table 2. Provenance variances of growth traits of Melia azedarach性状Trait 均值Average 变幅Range 变异系数Coefficient of variation/% 标准差Standard deviation 变异来源Variation source 种源Provenance 区组Block 树高Tree height/m 9.24 2.00~15.90 22.46 2.07 5.87** 10.91** 胸径DBH/cm 11.83 2.30~28.20 33.92 4.01 8.52** 3.43** 枝下高Clear bole height/m 3.12 0.30~8.90 46.98 4.01 1.41* 7.04** 干型Stem form 2.41 1.00~4.00 48.25 1.16 2.58** 2.82** 冠幅Crown width/m 3.14 0.40~9.20 33.54 1.05 2.48** 5.09** 保存率Survival rate 0.65 0.38~0.90 21.05 0.14 3.62** 4.26** 注:*表示P < 0.05水平上差异显著,**表示P < 0.01水平上差异极显著。下同。Notes: * represents significant differences at P < 0.05 level,** represents extremely significant differences at P < 0.01 level. Same as below. 经方差分析,除枝下高在种源间的差异为显著水平(P < 0.05)外,其余各性状在种源、重复效应上均达到了极显著水平(P < 0.01),说明不同种源对苦楝生长性状有着较大的影响,种源间变异明显。
2.2 种源性状与采种点地理因子的相关性分析
与采种点地理因子的Pearson相关分析(表 3)表明,除保存率这一性状外,采种点经纬度与种源其它性状均呈极显著负相关,其中,枝下高与经度、树高与纬度的相关性更强;采种点海拔与枝下高呈极显著正相关,与干型、冠幅呈显著正相关,与保存率呈显著负相关,而与树高、胸径的相关性不显著;枝下高、保存率2个性状与纬度的相关系数绝对值较其与经度的相关系数绝对值小,其余各性状则呈相反趋势。综上所述,苦楝的生长性状受经度、纬度及海拔的多重控制,以纬向变异为主,保存率这一性状的地理变异则主要由海拔控制。这说明来自南部及西部的种源生长更快,干型更优,高海拔种源生长较低海拔更快,但保存率下降。
表 3 生长性状与地理因子的Pearson相关分析Table 3. Pearson correlation analysis of growth traits and geographic factors性状Trait 树高Tree height 胸径DBH 枝下高Clear bole height 干型Stem form 冠幅Crown width 保存率Survival rate 经度Longitude -0.387** -0.403** -0.501** -0.441** -0.399** 0.241 纬度Latitude -0.631** -0.598** -0.379** -0.497** -0.520** 0.081 海拔Elevation 0.118 0.208 0.354** 0.257* 0.264* -0.252* 树高、胸径为苦楝生长的主要调查性状。为能更清晰地描述树高、胸径的地理变异趋势,利用趋势面技术,研究在经纬度影响下的苦楝生长变异模式。二元二次趋势面回归方程见表 4,其拟合精度和显著性均符合统计要求。
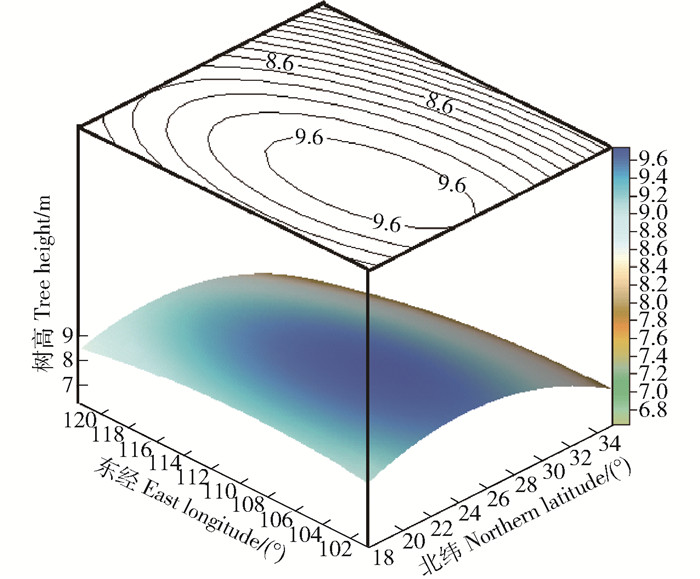
表 4 树高与胸径的趋势面分析回归方程Table 4. Regression equation of binary quadratic trend surface analysis for tree height and DBH性状Trait 趋势面回归方程Regression equation of trend surface analysis 拟合系数Fitting coefficient P 树高Tree height Z=1.179 5x+0.938 0y-0.018 0x2-0.004 1y2-0.003 0xy-54.497 0 0.628 < 0.01 胸径DBH Z=1.792 0x+2.016 6y-0.049 5x2-0.010 3y2+0.005 5xy-113.761 0 0.641 < 0.01 树高(图 2)、地径(图 3)的变异模式均为双向渐变。于树高这一性状,经度方向为负向变异,即西部种源的生长优于东部种源,变幅较小;纬度方向同为负向变异,即南部种源的生长优于北部种源,并有较大的变幅;由西南向东北方向,呈先上升后下降的趋势。胸径与树高有着同样的变化规律,但经向、纬向的变异幅度均较大。综上所述,树高、地径均受到经纬度的双重控制,但由纬度引起的差异更为显著,且偏西南的种源生长更好,北部种源的生长普遍偏差,这与Pearson相关分析一致。
![]() 图 2 树高与经纬度趋势面图Figure 2. Trend surface diagram of tree height and longitude as well as latitude
图 2 树高与经纬度趋势面图Figure 2. Trend surface diagram of tree height and longitude as well as latitude![]() 图 3 胸径与经纬度趋势面图Figure 3. Trend surface diagram of DBH and longitude as well as latitude
图 3 胸径与经纬度趋势面图Figure 3. Trend surface diagram of DBH and longitude as well as latitude2.3 种源性状与采种点气候因子的相关性分析
由表 5可知,气温相关因子对苦楝生长影响显著,采种点极端最低气温与枝下高呈显著正相关,与树高、胸径、干型、冠幅均呈极显著正相关;极端最高气温与保存率呈显著正相关,与干型、冠幅呈显著负相关,与枝下高呈极显著负相关;平均气温与冠幅呈显著正相关,与树高、胸径呈极显著正相关;平均最低气温、平均最高气温与干型、冠幅均呈显著正相关,与树高、胸径均呈极显著正相关。同时日照百分率、降水、极端最高气压等光照、降水、气压相关因子亦能显著影响苦楝的生长,极端最高本站气压与保存率呈极显著正相关,与枝下高呈极显著负相关;日降水量大于0.1 mm日数与树高、胸径、干型、冠幅均呈极显著正相关;日照百分率与胸径呈显著负相关,与保存率呈极显著负相关。由此可知,来自气温越高,降水量越多地区的种源在四会试验点生长更好,来自气压高、气温高、日照少地区的种源在四会试验点保存率更高。
表 5 生长性状与气候因子的Pearson相关分析Table 5. Pearson correlation analysis of growth traits and climatic factors气候因子
Climate factor树高
Tree height胸径
DBH枝下高
Clear bole height干型
Stem form冠幅
Crown width保存率
Survival rate极端最低气温Extreme minimum air temperature 0.461** 0.435** 0.254* 0.325** 0.387** 0.028 极端最高气压Extreme maximum air pressure -0.149 -0.184 -0.362** -0.174 -0.22 0.327** 极端最高气温Extreme maximum air temperature -0.205 -0.173 -0.412** -0.279* -0.257* 0.307* 平均气温Mean air temperature 0.403** 0.373** 0.107 0.245 0.304* 0.08 平均最低气温Mean minimum air temperature 0.404** 0.375** 0.117 0.257* 0.308* 0.125 平均最高气温Mean maximum air temperature 0.420** 0.396** 0.135 0.251* 0.322* -0.015 日降水量>0.1 mm日数Daily precipitation>0.1 mm days 0.434** 0.418** 0.166 0.327** 0.328** 0.165 日照百分率Sunshine percentage -0.227 -0.251* -0.051 -0.234 -0.181 -0.389** 典型相关分析可进一步揭示苦楝种源地理变异的气候生态学基础[28],将生长性状与8个主要气候因子进行典型相关分析(表 6)。典型相关系数为0.802,为极显著水平,说明气候因子对苦楝生长的地理变异有显著作用。在气候因子中,极端最低气温、平均最高气温的载荷较大,在0.5以上,其次是极端最高气温,其载荷为负值,绝对值为0.470,再其次是日降水量大于0.1 mm日数,载荷为0.459,极端最高本站气压、平均气温、平均最低气温的载荷绝对值均大于0.4。在性状中树高与胸径等生长性状的载荷较大,达0.8以上,其次是冠幅、干型、枝下高等形质性状,其载荷分别为0.762、0.678、0.582,性状中仅保存率的载荷为负值,其绝对值也较小,为0.222。这表明气温对树高、胸径的生长地理变异影响最大,来自气温高、降水多、气压低地区的种源,生长更为迅速,干型更好,这与Pearson相关分析一致。
表 6 生长性状与气候因子的典型相关Table 6. Canonical correlation analysis of growth traits and climate factors性状Trait 第一典型变量The first canonical variable(r=0.802,P < 0.001) 气候因子Climate factor 第一典型变量The first canonical variable(r=0.802,P < 0.001) 树高Tree height 0.857 极端最低气温Extreme minimum air temperature 0.555 0 胸径DBH 0.827 极端最高气压Extreme maximum air pressure -0.427 6 枝下高Clear bole height 0.582 极端最高气温Extreme maximum air temperature -0.469 5 干型Stem form 0.678 平均气温Mean air temperature 0.426 1 冠幅Crown width 0.762 平均最低气温Mean minimum air temperature 0.406 9 保存率Survival rate -0.222 平均最高气温Mean maximum air temperature 0.501 9 日降水量>0.1 mm日数Daily precipitation>0.1 mm days 0.458 9 日照百分率Sunshine percentage 0.015 7 2.4 种源聚类分析
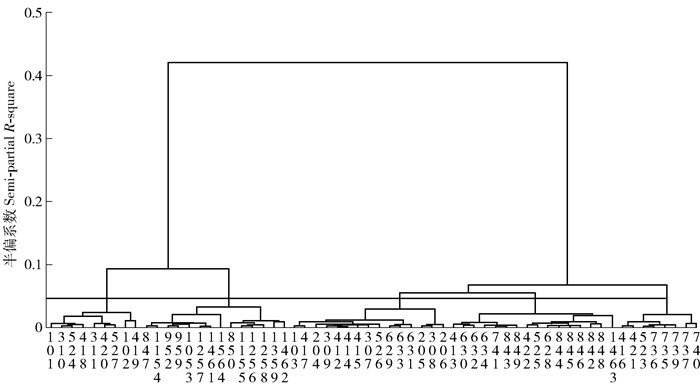
将种源树高、地径、枝下高、干型、冠幅及保存率等6个性状作为变量进行聚类分析,采用离差平方和法(Ward),将变量标准化为均值0,标准差为1,对62个种源进行聚类分析,见图 3。
以欧式距离为0.04,62个种源可被聚为5类。各类种源分别统计树高、胸径、干型及保存率等重要性状的均值(表 7),可以看出第4类生长最快,保存率最高,其次是第3类,生长最慢的是第2类,保存率最低的是第5类。第4类为来自贵州及广西东北部的种源,其表现最佳,树高均值为9.894 m,胸径均值为13.494 m,干型均值为2.718,保存率均值为0.764。第2类为来自江苏、浙江、安徽等华东地区及甘肃、陕西等西北地区的种源,其表现最差,树高均值为8.033 m,胸径均值为8.934 m,干型均值为2.007,保存率均值为0.633。
表 7 聚类分析结果Table 7. Result of cluster analysis类群Cluster 种源Provenance 地区Area 平均树高Mean treeheight/m 平均胸径Mean DBH/cm 平均干型Mean stem form 平均保存率Mean survival rate 1 101、102、310、311、418、419、420、524、527 闽西、粤东及海南等地区Northern Fujian, eastern Guangdong and Hainan, etc 8.838 10.715 2.118 0.579 2 847、850、952、959、1053、1154、1256、1257、1258、1359、1461、1462、1564 苏中南、浙北、安徽等华东地区,陇南、陕东等西北地区,渝及豫南等中部地区Central and southern of Jiangsu, northern Zhejiang, eastern Shaanxi, southern area of Henan and Gansu, Chongqing and Anhui, etc 8.033 8.934 2.007 0.633 3 103、204、205、206、307、308、309、412、414、415、417、526、629、631、633 粤桂湘赣的南部等地区Southern area of Guangdong, Guangxi, Hunan and Jiangxi, etc 9.671 12.725 2.526 0.661 4 413、422、525、628、630、632、634、741、842、843、844、845、846、848、849、1463 黔南及桂北等地区Southern Guizhou and northern Guangxi, etc 9.894 13.494 2.718 0.764 5 416、421、523、735、736、737、739、740 滇东南Eastern and southern Yunnan 9.411 12.48 2.51 0.507 3. 结论与讨论
苦楝于我国广泛分布,在长期的进化过程中,为适应复杂多变的地理生态环境而产生与之相应的遗传变异,并反映在生长性状中。因虫媒传粉植物的花粉扩散距离会被传粉昆虫的行为影响,能通过鸟类传播种子的植物的传播距离会被鸟类影响,而苦楝的野生群体多为小群体或单株散生,又是虫媒传粉植物,亦能被鸟类传播种子,这都促进了苦楝种源内的变异及种源间的分化[30-31]。本研究根据苦楝区划不同,采种进行种源试验,调查获得生长性状数据,分析其地理变异规律及气候生态学基础,为良种选育及种子调拨提供依据。参试种源数量达62个,涵盖全分布区,经纬度分布均匀。遗传分析方法多样,包括Pearson相关分析、趋势面分析、典型相关分析、聚类分析等,其中趋势面是通过对群体数据进行趋势拟合,表明各性状在地理因子影响下的具体变异趋势[2],多种分析方法所得结论也应更为科学更为全面。林木地理变异模式研究群体的树龄越大,研究的性状越多,越能反映林木真实的地理变异模式[29],本数据分析对象为4年生苦楝的6个性状,树龄较大,性状较多,研究结果有较高准确性,主要结论有以下4点:
(1) 研究结果表明,苦楝平均树高为9.24 m,变幅为2.00~15.90 m,平均胸径为11.83 m,变幅为2.30~28.20 m,性状变异系数区间为21.05%~48.25%,这6个性状在种源间、种源内均存在显著或极显著差异,说明四会试验点各种源长势较佳,种源间遗传分化显著,能初步反映出地理变异趋势。
(2) 除保存率这一统计性状外,其余性状与经纬度均呈极显著负相关,而保存率与海拔呈显著负相关,其中树高、胸径与纬度的相关系数绝对值较高,为0.631、0.598。树高、胸径是速生用材早期研究的重要指标,结合这2个性状的趋势面可知,树高、胸径的变异趋势为西南往东北方向,呈先上升后下降的趋势,结论表明随着采种点经纬度的递减,树木生长速度变快;随着采种点海拔的下降,保存率有所提高;来自偏西南地区的种源具有生长优势。结合Pearson与典型相关分析,发现来自极端气温气压温和、温度高、降水多地区的种源,生长优势更大。气候因子有较强的区域特征性,特别是温度相关因子,随着纬度的变化而产生显著差异,但在在经度上表现不明显,由典型相关分析可知极端最低气温、平均最高气温的载荷较大,分别为0.555、0.502,性状中树高与胸径的载荷较大,分别为0.857、0.827,这能说明温度是影响苦楝生长的主导因子,亦能说明温度是造成苦楝纬向变异的主要因素。
(3) 何霞等[19]对广州增城53个苦楝种源苗期地理变异研究表明采种点由南到北,由西到东,苦楝生长变慢,但树高以经度变异为主,这表明4年生苦楝的地理变异趋势与苗期保持一致,但随着树龄增加,树高的变异也将趋于纬度变异。陈羡德等[32]采集5个种源于福建省漳平市进行种源试验,发现来自低纬度地区的种源生长较佳,气温、降水量则是制约苦楝生长的主导因子,这与本研究结论一致。大多数树种的地理变异模式为经纬度双向渐变模式[33],苦楝也属其中。
(4) 聚类分析将62个种源聚为5类,来自闽西、粤东及海南等地区的种源可聚为一类;来自苏中南、浙北、安徽等华东地区,陇南、陕东等西北地区,渝及豫南可聚为一类;来自粤南、桂南、湘西南、赣南部地区的种源可聚为一类,来自黔南及桂北的种源可聚为一类,来自滇东南的种源可聚为一类,各类别之间差异明显。与程诗明等[3]报道的苦楝中国分布区的物候区划对比,结论基本一致,第1类来源于Ⅰ、Ⅳ物候区;第2类来源于Ⅵ、Ⅶ、Ⅹ物候区;第3类来源于Ⅱ、Ⅴ、Ⅵ物候区;第4类来源于Ⅵ物候区;第5类来源于Ⅲ物候区。
4. 展望
本研究是以4年生苦楝作为分析对象,可能此时各遗传性状并未得到充分表达或表达不稳定,且性状的早晚相关仍需进一步观测,性状与地理气候因子的相关性也需长期跟踪分析。本研究结论则可为试验的后续进行提供参考依据。根据表型性状可对苦楝种源进行早期区划,对地理变异模式进行初步的判断,但表型性状为本身遗传变异与环境因子互作的结果,为验证变异模式的准确性,应进行长期的多点种源试验,并加以分子遗传学的相关分析。
-
![]()
图 2 树高与经纬度趋势面图
Figure 2. Trend surface diagram of tree height and longitude as well as latitude
![]()
图 3 胸径与经纬度趋势面图
Figure 3. Trend surface diagram of DBH and longitude as well as latitude
表 1 苦楝各种源编号及海拔
Table 1 No. and elevation of Melia azedarach provenances
编号No. 种源地Provenance 海拔Elevation/m 编号No. 种源地Provenance 海拔Elevation/m 编号No. 种源地Provenance 海拔Elevation/m 101 福建连城Liancheng, Fujian 351 422 广东阳山Yangshan, Guangdong 93 844 贵州黎平Liping, Guizhou 650 102 福建永安Yongan, Fujian 373 523 海南海口Haikou, Hainan 129 845 贵州罗甸Luodian, Guizhou 386 103 福建漳平Zhangping, Fujian 219 524 海南屯昌Tunchang, Hainan 160 846 贵州荔波Libo, Guizhou 467 204 江西赣县Ganxian, Jiangxi 115 525 海南五指山Wuzhishan, Hainan 341 847 贵州德江Dejiang, Guizhou 851 205 江西于都Yudu, Jiangxi 145 526 海南陵水Lingshui, Hainan 79 848 贵州麻江Majiang, Guizhou 1 048 206 江西遂川Suichuan, Jiangxi 103 527 海南琼海Qionghai, Hainan 32 849 贵州贵阳Guiyang, Guizhou 1 205 307 湖南东安Dongan, Hunan 252 628 广西桂林Guilin, Guangxi 166 850 贵州石阡Shiqian, Guizhou 496 308 湖南浏阳Liuyang, Hunan 137 629 广西融安Rongan, Guangxi 226 952 浙江绍兴Shaoxing, Zhejiang 181 309 湖南通道Tongdao, Hunan 442 630 广西三江Sanjiang, Guangxi 240 959 浙江临安Lin’an, Zhejiang 47 310 湖南炎陵Yanling, Hunan 192 631 广西钦州Qinzhou, Guangxi 250 1053 重庆Chongqing 183 311 湖南龙山Longshan, Hunan 779 632 广西梧州Wuzhou, Guangxi 31 1154 安徽黄山Huangshan, Anhui 306 412 广东仁化Renhua, Guangdong 196 633 广西凭祥Pingxiang, Guangxi 255 1155 安徽利辛Lixin, Anhui 38 413 广东郁南Yunan, Guangdong 22 634 广西靖西Jingxi, Guangxi 764 1256 江苏泰兴Taixing, Jiangsu 7 414 广东博罗Boluo, Guangdong 13 735 云南富宁Funing, Yunnan 732 1257 江苏南京Nanjing, Jiangsu 22 415 广东开平Kaiping, Guangdong 8 736 云南广南Guangnan, Yunnan 1 079 1258 江苏姜堰Jiangyan, Jiangsu 8 416 广东清远Qingyuan, Guangdong 73 737 云南开远Kaiyuan, Yunnan 1 122 1359 河南内乡Neixiang, Henan 178 417 广东恩平Enping, Guangdong 17 739 云南勐腊Mengla, Yunnan 1 005 1461 陕西安康Ankang, Shaanxi 422 418 广东饶平Raoping, Guangdong 20 740 云南罗平Luoping, Yunnan 1 415 1462 陕西蒲城Pucheng, Shaanxi 488 419 广东五华Wuhua, Guangdong 123 741 云南麻栗坡Malipo, Yunnan 1 113 1463 陕西南郑Nanzheng, Shaanxi 716 420 广东东源Dongyuan, Guangdong 57 842 贵州兴义Xingyi, Guizhou 1 217 1564 甘肃舟曲Zhouqu, Gansu 1 354 421 广东茂名Maoming, Guangdong 162 843 贵州册亨Ceheng, Guizhou 993  下载: 导出CSV
下载: 导出CSV
表 2 苦楝生长性状种源变异分析
Table 2 Provenance variances of growth traits of Melia azedarach
性状Trait 均值Average 变幅Range 变异系数Coefficient of variation/% 标准差Standard deviation 变异来源Variation source 种源Provenance 区组Block 树高Tree height/m 9.24 2.00~15.90 22.46 2.07 5.87** 10.91** 胸径DBH/cm 11.83 2.30~28.20 33.92 4.01 8.52** 3.43** 枝下高Clear bole height/m 3.12 0.30~8.90 46.98 4.01 1.41* 7.04** 干型Stem form 2.41 1.00~4.00 48.25 1.16 2.58** 2.82** 冠幅Crown width/m 3.14 0.40~9.20 33.54 1.05 2.48** 5.09** 保存率Survival rate 0.65 0.38~0.90 21.05 0.14 3.62** 4.26** 注:*表示P < 0.05水平上差异显著,**表示P < 0.01水平上差异极显著。下同。Notes: * represents significant differences at P < 0.05 level,** represents extremely significant differences at P < 0.01 level. Same as below.
下载: 导出CSV
表 3 生长性状与地理因子的Pearson相关分析
Table 3 Pearson correlation analysis of growth traits and geographic factors
性状Trait 树高Tree height 胸径DBH 枝下高Clear bole height 干型Stem form 冠幅Crown width 保存率Survival rate 经度Longitude -0.387** -0.403** -0.501** -0.441** -0.399** 0.241 纬度Latitude -0.631** -0.598** -0.379** -0.497** -0.520** 0.081 海拔Elevation 0.118 0.208 0.354** 0.257* 0.264* -0.252*
下载: 导出CSV
表 4 树高与胸径的趋势面分析回归方程
Table 4 Regression equation of binary quadratic trend surface analysis for tree height and DBH
性状Trait 趋势面回归方程Regression equation of trend surface analysis 拟合系数Fitting coefficient P 树高Tree height Z=1.179 5x+0.938 0y-0.018 0x2-0.004 1y2-0.003 0xy-54.497 0 0.628 < 0.01 胸径DBH Z=1.792 0x+2.016 6y-0.049 5x2-0.010 3y2+0.005 5xy-113.761 0 0.641 < 0.01
下载: 导出CSV
表 5 生长性状与气候因子的Pearson相关分析
Table 5 Pearson correlation analysis of growth traits and climatic factors
气候因子
Climate factor树高
Tree height胸径
DBH枝下高
Clear bole height干型
Stem form冠幅
Crown width保存率
Survival rate极端最低气温Extreme minimum air temperature 0.461** 0.435** 0.254* 0.325** 0.387** 0.028 极端最高气压Extreme maximum air pressure -0.149 -0.184 -0.362** -0.174 -0.22 0.327** 极端最高气温Extreme maximum air temperature -0.205 -0.173 -0.412** -0.279* -0.257* 0.307* 平均气温Mean air temperature 0.403** 0.373** 0.107 0.245 0.304* 0.08 平均最低气温Mean minimum air temperature 0.404** 0.375** 0.117 0.257* 0.308* 0.125 平均最高气温Mean maximum air temperature 0.420** 0.396** 0.135 0.251* 0.322* -0.015 日降水量>0.1 mm日数Daily precipitation>0.1 mm days 0.434** 0.418** 0.166 0.327** 0.328** 0.165 日照百分率Sunshine percentage -0.227 -0.251* -0.051 -0.234 -0.181 -0.389**
下载: 导出CSV
表 6 生长性状与气候因子的典型相关
Table 6 Canonical correlation analysis of growth traits and climate factors
性状Trait 第一典型变量The first canonical variable(r=0.802,P < 0.001) 气候因子Climate factor 第一典型变量The first canonical variable(r=0.802,P < 0.001) 树高Tree height 0.857 极端最低气温Extreme minimum air temperature 0.555 0 胸径DBH 0.827 极端最高气压Extreme maximum air pressure -0.427 6 枝下高Clear bole height 0.582 极端最高气温Extreme maximum air temperature -0.469 5 干型Stem form 0.678 平均气温Mean air temperature 0.426 1 冠幅Crown width 0.762 平均最低气温Mean minimum air temperature 0.406 9 保存率Survival rate -0.222 平均最高气温Mean maximum air temperature 0.501 9 日降水量>0.1 mm日数Daily precipitation>0.1 mm days 0.458 9 日照百分率Sunshine percentage 0.015 7
下载: 导出CSV
表 7 聚类分析结果
Table 7 Result of cluster analysis
类群Cluster 种源Provenance 地区Area 平均树高Mean treeheight/m 平均胸径Mean DBH/cm 平均干型Mean stem form 平均保存率Mean survival rate 1 101、102、310、311、418、419、420、524、527 闽西、粤东及海南等地区Northern Fujian, eastern Guangdong and Hainan, etc 8.838 10.715 2.118 0.579 2 847、850、952、959、1053、1154、1256、1257、1258、1359、1461、1462、1564 苏中南、浙北、安徽等华东地区,陇南、陕东等西北地区,渝及豫南等中部地区Central and southern of Jiangsu, northern Zhejiang, eastern Shaanxi, southern area of Henan and Gansu, Chongqing and Anhui, etc 8.033 8.934 2.007 0.633 3 103、204、205、206、307、308、309、412、414、415、417、526、629、631、633 粤桂湘赣的南部等地区Southern area of Guangdong, Guangxi, Hunan and Jiangxi, etc 9.671 12.725 2.526 0.661 4 413、422、525、628、630、632、634、741、842、843、844、845、846、848、849、1463 黔南及桂北等地区Southern Guizhou and northern Guangxi, etc 9.894 13.494 2.718 0.764 5 416、421、523、735、736、737、739、740 滇东南Eastern and southern Yunnan 9.411 12.48 2.51 0.507
下载: 导出CSV
-
[1] 陈丽君, 邓小梅, 丁美美, 等.苦楝种源果核及种子性状地理变异的研究[J].北京林业大学学报, 2014, 36(1): 15-20. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=8cbe223d-0465-4045-9a5a-34346f8c6715 Chen L J, Deng X M, Ding M M, et al. Geographic variation in traits of fruit stones and seeds of Melia azedarach[J]. Journal of Beijing Forestry University, 2014, 36(1): 15-20. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=8cbe223d-0465-4045-9a5a-34346f8c6715
[2] 廖柏勇, 陈丽君, 王芳, 等.苦楝种源果核和种子性状地理变异的趋势面分析[J].林业科学研究, 2016, 29(5): 784-792. doi: 10.3969/j.issn.1001-1498.2016.05.023 Liao B Y, Chen L J, Wang F, et al. Trendhe surface analysis of provenances geographic variation of Melia azedarach stone and seed[J]. Forest Research, 2016, 29(5): 784-792. doi: 10.3969/j.issn.1001-1498.2016.05.023
[3] 程诗明, 顾万春.苦楝中国分布区的物候区划[J].林业科学, 2005, 41(3): 186-191. doi: 10.3321/j.issn:1001-7488.2005.03.032 Cheng S M, Gu W C. The phenological division of distribution area in China for Melia azedarach[J]. Scientia Silvae Sinicae, 2005, 41(3): 186-191. doi: 10.3321/j.issn:1001-7488.2005.03.032
[4] 程诗明, 顾万春.苦楝表型性状梯度变异的研究[J].林业科学, 2006, 42(5): 29-35. http://d.old.wanfangdata.com.cn/Periodical/lykx200605006 Chen S M, Gu W C. Studies on phenotypical characteristics gradient variation of Melia azedarach[J]. Scientia Silvae Sinicae, 2006, 42(5): 29-35. http://d.old.wanfangdata.com.cn/Periodical/lykx200605006
[5] 郭尧邦. 3年生苦楝生长量与坡向、坡位关系的研究[J].安徽农学通报, 2012, 18(6): 90-92, 133. doi: 10.3969/j.issn.1007-7731.2012.06.048 Guo Y B. Research on the relationship of Melia azedarach growth and slope direction, slope position[J]. Anhui Agricutural Science Bulletin, 2012, 18(6): 90-92, 133. doi: 10.3969/j.issn.1007-7731.2012.06.048
[6] 陈丽君, 刘明骞, 廖柏勇, 等.苦楝不同种源叶片性状变异研究[J].西南林业大学学报, 2016, 36(5): 16-20. http://d.old.wanfangdata.com.cn/Periodical/xnlxyxb201605003 Chen L J, Liu M Q, Liao B Y, et al. Variation of leaf morphological characters of Melia azedarach from different provenances[J]. Journal of Southwest Forestry University, 2016, 36(5): 16-20. http://d.old.wanfangdata.com.cn/Periodical/xnlxyxb201605003
[7] 教忠意, 徐长柏, 丁学农.苦楝优良家系初步选择[J].浙江林业科技, 2011, 31(3): 48-50. doi: 10.3969/j.issn.1001-3776.2011.03.012 Jiao Z Y, Xu C B, Ding X N. Preliminary selection of optimal family on Melia azedarach[J]. Journal of Zhejiang Forestry Science & Technology, 2011, 31(3): 48-50. doi: 10.3969/j.issn.1001-3776.2011.03.012
[8] 李桂英, 支国.苦楝皮提取物的抗肿瘤活性研究[J].安徽农业科学, 2012, 40(11): 6433-6434. doi: 10.3969/j.issn.0517-6611.2012.11.032 Li G Y, Zhi G. Anti-tumor activity of cortex Melia extract[J]. Journal of Anhui Agricultural Sciences, 2012, 40(11): 6433-6434. doi: 10.3969/j.issn.0517-6611.2012.11.032
[9] 王家源, 郭杰, 喻方圆.不同种源苦楝种子生物学特性差异[J].南京林业大学学报(自然科学版), 2013, 37(1): 49-54. http://cdmd.cnki.com.cn/Article/CDMD-10298-2008112957.htm Wang J Y, Guo J, Yu F Y. The difference of seed biological characters in Melia azedarach L. from different provenances[J].Journal of Nanjing Forestry University(Natural Science Edition), 2013, 37(1): 49-54. http://cdmd.cnki.com.cn/Article/CDMD-10298-2008112957.htm
[10] Akacha M, Lahbib K, Daami-Remadi M, et al. Antibacterial, antifungal and anti-inflammatory activities of Melia azedarach ethanolic leaf extract[J]. Bangladesh Journal of Pharmacology, 2016, 11(3): 666-674. doi: 10.3329/bjp.v11i3.27000
[11] Muhammad M T, Lubna, Fayyaz N, et al. Antibacterial activity of flower of Melia azedarach Linn. and identification of its metabolites[J]. Journal of the Korean Society for Applied Biological Chemistry, 2015, 58(2):219-227. doi: 10.1007/s13765-015-0029-7
[12] 王荣国.乡土树种苦楝的栽培与应用[J].安徽农学通报, 2011, 17(16): 80-81, 211. doi: 10.3969/j.issn.1007-7731.2011.16.045 Wang R G. Cultivation and application of native tree species of Mlie azedarach L.[J]. Anhui Agricutural Science Bulletin, 2011, 17(16): 80-81, 211. doi: 10.3969/j.issn.1007-7731.2011.16.045
[13] 廖源林, 蔡仕珍, 叶充, 等.模拟酸雨对苦楝生理生态特性的影响[J].生态学杂志, 2015, 34(10): 2764-2770. http://d.old.wanfangdata.com.cn/Periodical/stxzz201510011 Liao Y L, Cai S Z, Ye C, et al. Effects of simulated acid rain on ecophysiological characteristics of Melia azedarach[J].Chinese Journal of Ecology, 2015, 34(10): 2764-2770. http://d.old.wanfangdata.com.cn/Periodical/stxzz201510011
[14] 张子晗, 王家源, 喻方圆.盐胁迫对两种源苦楝种子萌发特性的影响[J].南京林业大学学报(自然科学版), 2014, 38(5): 107-112. http://d.old.wanfangdata.com.cn/Periodical/njlydxxb201405021 Zhang Z H, Wang J Y, Yu F Y. Effects of NaCl stress on germination characteristics of Melai azedarach L. seeds from two provenances[J].Journal of Nanjing Forestry University(Natural Science Edition), 2014, 38(5): 107-112. http://d.old.wanfangdata.com.cn/Periodical/njlydxxb201405021
[15] 曲良谱, 李霞.容器苗密度对苦楝苗木质量的影响[J].安徽农业科学, 2017, 45(3): 23-24. doi: 10.3969/j.issn.0517-6611.2017.03.008 Qu L P, Li X. Effect of container seedling densities on the quality of Melia azedarach seedlings[J]. Journal of Anhui Agricultural Sciences, 2017, 45(3): 23-24. doi: 10.3969/j.issn.0517-6611.2017.03.008
[16] 苏明洁, 廖源林, 叶充, 等.镉胁迫下苦楝(Melia azedarach L.)幼苗的生长及生理响应[J].农业环境科学学报, 2016, 35(11): 2086-2093. doi: 10.11654/jaes.2016-0929 Su M J, Liao Y L, Ye C, et al. The growth and physiological responses of Melia azedarach L. seedlings to cadmium stress[J]. Journal of Agro-environment Science, 2016, 35(11): 2086-2093. doi: 10.11654/jaes.2016-0929
[17] 廖柏勇, 陈晓阳, 陈丽君, 等.苦楝种源间种子发芽变异的观测[J].广东农业科学, 2014, 41(11): 43-47. doi: 10.3969/j.issn.1004-874X.2014.11.010 Liao B Y, Chen X Y, Chen L J, et al. Observation of Melia azedarach seed germination variation among provenances[J].Guangdong Agricultural Sciences, 2014, 41(11): 43-47. doi: 10.3969/j.issn.1004-874X.2014.11.010
[18] 陈丽君, 刘明骞, 廖柏勇, 等.苦楝不同种源苗期生长性状和生长节律研究[J].西南林业大学学报, 2014, 34(4): 1-7. doi: 10.3969/j.issn.2095-1914.2014.04.001 Chen L J, Liu M Q, Liao B Y, et al. Growth traits and growth rhythm of Melia azedarach seedlings from different provenances[J]. Journal of Southwest Forestry University, 2014, 34(4): 1-7. doi: 10.3969/j.issn.2095-1914.2014.04.001
[19] 何霞, 廖柏勇, 王芳, 等.苦楝种源幼林期生长性状地理变异的研究[J].华南农业大学学报, 2016, 37(4): 75-81. http://d.old.wanfangdata.com.cn/Periodical/hnnydxxb201604013 He X, Liao B Y, Wang F, et al. Geographic variations in growth traits of one year test forest with different Melia azedarach provenances[J]. Journal of South China Agricultural University, 2016, 37(4): 75-81. http://d.old.wanfangdata.com.cn/Periodical/hnnydxxb201604013
[20] 李世全, 喻才员.不同种源苦楝种子形态特征和播种品质差异研究[J].江西林业科技, 2009(3): 9-12. doi: 10.3969/j.issn.1006-2505.2009.03.003 Li S Q, Yu C Y. Studies on seed morphological characteristics and seeding quality of Melia azedarach seeds among provenances[J].Jiangxi Forestry Science and Technology, 2009(3): 9-12. doi: 10.3969/j.issn.1006-2505.2009.03.003
[21] 教忠意, 唐凌凌, 王保松, 等.东部沿海地区苦楝果实的性状变异[J].浙江林学院学报, 2009, 26(6): 792-796. doi: 10.3969/j.issn.2095-0756.2009.06.005 Jiao Z Y, Tang L L, Wang B S, et al. Fruits characteristics of Melia azedarach in eastern coastal areas of China[J]. Journal of Zhejiang Forestry College, 2009, 26(6): 792-796. doi: 10.3969/j.issn.2095-0756.2009.06.005
[22] 董玉峰, 姜岳忠, 马玲, 等.苦楝遗传改良研究进展[J].山东林业科技, 2010, 40(4): 104-107. doi: 10.3969/j.issn.1002-2724.2010.04.031 Dong Y F, Jiang Y Z, Ma L, et al. Genetic improvement of Melia azedarach[J].Shandong Forestry Science and Technology, 2010, 40(4): 104-107. doi: 10.3969/j.issn.1002-2724.2010.04.031
[23] 魏胜利, 王文全, 秦淑英, 等.甘草种源种子形态与萌发特性的地理变异研究[J].中国中药杂志, 2008, 33(8):869-873. doi: 10.3321/j.issn:1001-5302.2008.08.001 Wei S L, Wang W Q, Qin S Y, et al. Study on geographical variation of morphologic and germination characteristic of different Glycyrrhiza uralensis provenance seeds[J]. China Journal of Chinese Material Medica, 2008, 33(8):869-873. doi: 10.3321/j.issn:1001-5302.2008.08.001
[24] 惠文凯, 陈晓阳, 刘明骞, 等.麻风树种源间种实性状变异研究[J].北京林业大学学报, 2014, 36(3):110-114. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=b08f480f-c9eb-47f2-83fd-8e72817ada45 Hui W K, Chen X Y, Liu M Q, et al. Variation of fruits and seed traits of Jatropha curcas L. among provenances[J].Journal of Beijing Forestry University, 2014, 36(3):110-114. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=b08f480f-c9eb-47f2-83fd-8e72817ada45
[25] 孙操稳, 贾黎明, 叶红莲, 等.无患子果实经济性状地理变异评价及与脂肪酸成分相关性[J].北京林业大学学报, 2016, 38(12):73-83. http://www.cnki.net/KCMS/detail/detail.aspx?filename=BJLY201612009&dbname=cjfdtotal&dbcode=CJFD&v=MjM3NTdMdk1KeWZIZDdHNEg5Zk5yWTlGYllSNkRnOC96aFlVN3pzT1QzaVFyUmN6RnJDVVJMT2ZZT1p0RnkvZ1U= Sun C W, Jia L M, Ye H L, et al. Geographic variation evaluating and correlation with fatty acid composition of economic characters of Sapindus spp. fruits[J].Journal of Beijing Forestry University, 2016, 38(12):73-83. http://www.cnki.net/KCMS/detail/detail.aspx?filename=BJLY201612009&dbname=cjfdtotal&dbcode=CJFD&v=MjM3NTdMdk1KeWZIZDdHNEg5Zk5yWTlGYllSNkRnOC96aFlVN3pzT1QzaVFyUmN6RnJDVVJMT2ZZT1p0RnkvZ1U=
[26] 张伟红, 王润辉, 郑会全, 等.乐昌含笑种源地理变异研究[J].西南林业大学学报, 2015, 35(2):24-29. http://d.old.wanfangdata.com.cn/Periodical/xnlxyxb201502004 Zhang W H, Wang R H, Zheng H Q, et al. Geographic variation pattern of provenances of Michelia chapensis[J].Journal of Southwest Forestry University, 2015, 35(2):24-29. http://d.old.wanfangdata.com.cn/Periodical/xnlxyxb201502004
[27] 刘志龙, 虞木奎, 马跃, 等.不同种源麻栎种子和苗木性状地理变异趋势面分析[J].生态学报, 2011, 31(22): 6796-6804. http://d.old.wanfangdata.com.cn/Periodical/stxb201122016 Liu Z L, Yu M K, Ma Y, et al. A trend surface analysis of geographic variation in the traits of seeds and seedlings from different Quercus acutissima provenances[J]. Acta Ecologica Sinica, 2011, 31(22): 6796-6804. http://d.old.wanfangdata.com.cn/Periodical/stxb201122016
[28] 陈晓阳.典型相关分析在树种地理变异研究中的应用[J].北京林业大学学报, 1990, 12(3): 53-60. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=F48E6680-503D-4A34-B9A8-723B4C4AA455 Chen X Y. Canonical correlation analysis and its application in geographic variation of trees[J]. Journal of Beijing Forestry University, 1990, 12(3): 53-60. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=F48E6680-503D-4A34-B9A8-723B4C4AA455
[29] 刘宇, 徐焕文, 尚福强, 等. 16年生白桦种源变异及区划[J].林业科学, 2016, 52(9): 48-56. http://d.old.wanfangdata.com.cn/Periodical/lykx201609006 Liu Y, Xu H W, Shang F Q, et al. Variation and zoning of 16-year-old Betula platyphylla provenance[J].Scientia Silvae Sinicae, 2016, 52(9): 48-56. http://d.old.wanfangdata.com.cn/Periodical/lykx201609006
[30] 郑健, 胡增辉, 郑勇奇, 等.花楸树种源间表型性状的地理变异分析[J].植物资源与环境学报, 2012, 21(3): 50-56. doi: 10.3969/j.issn.1674-7895.2012.03.008 Zheng J, Hu Z H, Zheng Y Q, et al. Analysis on geographic variation of phenotypic traits of Sorbus pohuashanensis among different provenances[J].Journal of Plant Resources and Environment, 2012, 21(3): 50-56. doi: 10.3969/j.issn.1674-7895.2012.03.008
[31] 廖柏勇.苦楝地理变异和遗传多样性研究[D].广州: 华南农业大学, 2016. Liao B Y. Studies on geographic variation and genetic diversity ofChinese populations of Melia azedarach L. germplasm resources[D]. Guangzhou: South China Agriculture University, 2016.
[32] 陈羡德, 陈礼光, 阙茂文, 等.不同种源苦楝幼林试验初步研究[J].福建林学院学报, 2008, 28(1): 23-26. doi: 10.3969/j.issn.1001-389X.2008.01.006 Chen X D, Chen L G, Que M W, et al. Preliminary study on the provenance test of Melia azedarach[J].Journal of Fujian College of Forestry, 2008, 28(1): 23-26. doi: 10.3969/j.issn.1001-389X.2008.01.006
[33] 朱翔, 杨传平, 李忠, 等. 2年生白桦种源的地理变异[J].东北林业大学学报, 2001, 29(6): 7-10. doi: 10.3969/j.issn.1000-5382.2001.06.003 Zhu X, Yang C P, Li Z, et al. The Geographic variance of provenance for Betula platyphylla in two years old[J].Journal of Northeast Forestry University, 2001, 29(6): 7-10. doi: 10.3969/j.issn.1000-5382.2001.06.003
-
期刊类型引用(10)
1. 赵海文,周长品,刘一贞,樊小丽,汤道平,李发根. 粗皮桉早期生长模型拟合以及生长节律研究. 华南农业大学学报. 2025(02): 212-221 .  百度学术
百度学术
2. 吴云燕,张露,刘远生,程强强,苏仁峰,潘俊彬,刘军,刘玮,马际凯. 不同种源/家系毛红椿连年生长性状变异及早期选择. 江西农业大学学报. 2024(01): 106-117 . 百度学术
3. 崔琳,王力刚,张玉柱,韩勤,司思. 不同种源平榛生长性状变异特征及优良种源筛选. 中国果树. 2023(07): 52-59 . 百度学术
4. 王淼,吴国亮,张蕊,王家燚,王云鹏,黄大庄,周志春. 木荷18年生种源生长、材性的地理种源变异. 应用生态学报. 2023(09): 2337-2344 . 百度学术
5. 欧斌,朱江华,韩璐,李畅,李远章,徐晨,郭庭铣,崔子佳. 苦楝不同种源生长分析及优良种源选择研究. 南方林业科学. 2021(02): 26-31+62 . 百度学术
6. 尧云萍,赵兰,何万存,胡百强,晏雨鸿,陈梦义,丁菲,吴南生. 南酸枣果实表型性状及发芽率地理变异研究. 江西农业大学学报. 2021(05): 996-1006 . 百度学术
7. 于增金,郑风英,任可,吴佳木,荣俊冬,陈礼光,郑郁善. 凹叶厚朴种源苗期性状比较及地理变异. 福建农林大学学报(自然科学版). 2020(02): 185-191 . 百度学术
8. 符真珠,袁欣,王爱科,董晓宇,王利民,张和臣. 彩叶楝树组织培养技术. 北方园艺. 2020(13): 71-76 . 百度学术
9. 林春光,王伟,许天委,李国寅. 不同纬度大叶榄仁种实性状差异分析. 热带农业科学. 2020(12): 10-15 . 百度学术
10. 刘德浩,张卫华,潘文,徐斌,朱报著. 尾叶桉种源间数量性状的地理变异研究. 惠州学院学报. 2019(03): 59-65 . 百度学术
其他类型引用(5)

计量
- 文章访问数: 1928
- HTML全文浏览量: 362
- PDF下载量: 103
- 被引次数: 15
