
Optimization on the obstacle crossing ability and position of a forestry chassis with double-cylinder wheeled-legs
-
摘要:目的底盘是林业装备的关键部件,底盘技术性能中的越障性能及地形适应性能尤为关键,直接决定了林业装备是否能够上山入林。我国典型人工林具有多矮小障碍、多沟壑等立地条件特点,传统林业装备底盘在复杂立地条件下作业的能力就凸显不足,因此本研究旨在提高林业装备底盘越障性能。方法针对一种林用新型步态六轮变幅轮腿底盘,运用D-H参数法对底盘轮腿机构建立运动学模型。通过空间几何位置坐标变换方法对变幅轮腿底盘单缸控制轮腿与双缸控制轮腿越障能力进行理论计算和分析比较。运用D-H运动学正解方法计算得出双缸的最佳布置位置参数。通过仿真测试、样机测试与理论分析进行对比研究。结果双缸控制轮腿机构的越障高度始终大于单缸控制轮腿机构。理论分析、仿真分析与样机测试在最大越障高度数值的切合度分别为99.7%和97.8%。底盘轮腿越障高度可达285.9mm, 具有较强的越障性能和地形适应性,其越障能力完全满足我国林业生产对林业装备底盘越障性能的要求,充分验证了理论研究方法的正确性。结论为解决我国林业装备底盘越障性能较差这一难题提供了新的思路,为双缸控制轮腿机构运动特性、变幅轮腿底盘越障性能及大规模林业机械智能化采伐底盘的研究和发展应用提供了理论依据。Abstract:ObjectiveThe obstacle crossing performance and adaptability to different terrain are most important factors for the technical characteristics of the chassis that is the key part of forestry equipment, also they are the key reasons to decide whether the forestry equipment could work in the forest. Since the typical plantations in our country have many short obstacles and gullies, the obstacle crossing performance and adaptability of traditional chassis are not good enough to work in the complex terrain.MethodFor improving the obstacle crossing performance, a method based on D-H theory was used to establish a kinematics model for a new type of six-wheeled-leg forestry chassis with levelling initiative gait. By space geometry coordinates transformation method, single-cylinder wheeled-legs mechanism was compared with double-cylinder wheeled-legs mechanism in obstacle crossing ability.ResultIt was concluded that the obstacle crossing ability of double-cylinder wheeled-legs mechanism was always greater than single-cylinder wheeled-legs mechanism. The optimum arrangement of double-cylinder position was achieved with the D-H kinematics solution method. In contrast to prototype test, the highest obstacle crossing numerical similarities of theory analysis and simulation were 99.7% and 97.8%, respectively. The obstacle crossing height of wheeled-legs of chassis was much better than before, and could reach 285.9mm. So it is completely suitable for the application in the field of forestry and a typical unknown terrain. The correctness of the theoretical research method was validated.ConclusionThis paper provides theory basis for the research on the double-cylinder wheeled-legs mechanism motion characteristics, the obstacle crossing ability of chassis, and the large-scale development and application of intelligent forestry mechanical chassis.
-
丛枝菌根真菌(arbuscular mycorrhiza fungi,AMF),作为一种与植物关系最为密切的微生物,广泛分布于地球,能与陆地上大约90%的植物产生共生关系[1]。AMF能够为植物提供一个良好的根际环境,并通过增进营养吸收、提高光合效率等多种途径,改善植物的生长发育,促进植物与生态系统的C、N、P循环,提高植物生长量。Wu等[2]研究表明,AMF定殖欧美杨107(Populus × euramericana ‘Neva’)可以显著提高其氮素吸收及光合作用。
植物可通过提高次生代谢产物含量、防御蛋白活性或者改变体内营养成分含量等一系列生理生化反应提高其化学防御能力,抵御植食性昆虫的危害[3]。植物次生代谢产物是非营养物质,能够增加昆虫的代谢负担、降低昆虫对食物的利用效率,进而影响其正常生长发育[4-5]。防御蛋白包括防御酶和蛋白酶抑制剂。防御酶是植物由初生代谢转为次生代谢途径中的关键酶,对降低植物的营养价值和毒害植食昆虫方面具有重要意义[6],主要包括苯丙氨酸解氨酶(PAL)和多酚氧化酶(PPO);蛋白酶抑制剂能削弱昆虫肠道内蛋白酶活性或阻断消化酶对食物中蛋白质的消化利用,还能诱导昆虫消化酶的过度分泌,造成昆虫营养不良,进而影响昆虫的生长[7]。尽管,植物体内营养物质的含量是衡量植物抗虫性的另一种生理指标[8],但是,营养物质与次生代谢产物和防御蛋白对抗虫性的调控机制并不一致,因为营养物质含量与抗虫性并不一定呈线性相关,主要取决于含碳、氮营养物质的比例[9-10]。大量研究表明,AMF作为一种生物制剂在与植物形成共生体后,会与植物进行复杂的信号分子交换并产生诱导因子,诱导激发植物的化学防御[11],在一定程度上诱导激发植物的抗病性、抗虫性或抗应激能力,间接提高植物对不利环境条件的适应性[12]。例如:异形根孢囊霉(Rhizophagus irregularis)定殖车前草(Plantago lanceolata)后,能显著提高其叶片中次生代谢产物梓醇的含量,使甜菜夜蛾(Spodoptera exigua)的死亡率显著升高[13]。Nishida等[14]研究显示,将日本百脉根(Lotus japonicus)分别接种根内球囊霉(Glomus intraradices,GI)、幼套近明球囊霉(Glomus etunicatum,GE)、聚丛球囊霉(Glomus aggregatum,GA)、稍长无梗囊霉(Acaulospora longula)、球状巨孢囊霉(Gigaspora margarita)或玫瑰红巨孢囊霉(Gigaspora rosea)6种AMF可以显著提高其叶片内酚类的含量,使二斑叶螨(Tetranychus urticae)的取食和产卵受到显著抑制。GI能通过茉莉酸途径诱导黄瓜(Cucumis sativus)产生防御性物质葫芦素C,使甜菜夜蛾的取食量显著低于对照[15]。
目前,关于AMF的研究,多以豆科植物或农业经济型植物的抗逆性为主,其对林木抗虫性影响的研究较少。本研究以银中杨(Populus alba × P. berolinensis)为对象,对其接种摩西球囊霉(Glomus mosseae,GM)或GI,在侵染率达到最高时,测定银中杨叶片N和P元素、营养物质、次生代谢产物含量和防御蛋白活性,分析AMF对银中杨物质代谢及化学防御的影响,为利用AMF诱导增强林木化学防御及提高林木抗虫性提供理论依据。
1. 材料与方法
1.1 试验材料
供试植物:1年生银中杨扦插苗。扦插前一周对插穗进行沙埋,并定期浇水。将插穗取出后,用500 mg/L生根粉溶液浸泡12 h后扦插。
供试菌剂:摩西球囊霉(GM)和根内球囊霉(GI)由甘肃农科院提供,通过宿主玉米和三叶草扩繁完成,菌剂中包含孢子、菌丝、根段和沙子,其中孢子含量15 个/g。
土壤:草炭土∶蛭石∶沙子 = 3∶1∶1,混合后在121 ℃下高压灭菌2 h。
1.2 试验设计
试验前,将高11.5 cm的32孔塑料穴盘和容量为1 L的花盆用0.3%KMnO4溶液浸泡2 h进行消毒处理。在穴盘内装入灭菌后的土壤。将浸泡后的插穗扦插于穴盘,每穴1株,定期浇水,一个月后移栽至花盆。将花盆分为3组,在AMF处理组的每个花盆中,分别装入1.3 kg灭菌土壤与20 g GM或GI的均匀混合物,对照组只装入1.3 kg灭菌土壤,不加菌剂。每盆移栽1株扦插苗,每组150株,3组共450株。移栽后,在东北林业大学林木遗传育种苗圃温室进行培养,并进行定期浇水除草。
1.3 指标测定
1.3.1 侵染率测定
根据Phillips等[16]方法修改。在移植后30、60、90和100 d,从每组随机各选取5株样树,检测AMF侵染率。将样树根部须根剪下,洗净,剪成1 ~ 2 cm长的小段,先加入10%KOH,放入90 ℃水浴锅加热30 min使根透明(溶液变黄要及时更换,直至溶液不再变黄)。根透明后用蒸馏水清洗3 ~ 5次,再加入20%过氧化氢使根软化,静置10 ~ 15 min,取出根用蒸馏水洗净;然后加入2%稀盐酸酸化5 min,用蒸馏水洗净,接着加入曲利苯蓝溶液(乳酸(mL)∶甘油(mL)∶曲利苯蓝(g) = 100∶100∶0.2)进行染色30 min。染色完成后用蒸馏水清洗两次,最后放入乳酸−甘油(1∶1)中脱色12 h,取出后置于乳酸甘油溶液(乳酸∶甘油∶水 = 1∶1∶1)中,4 ℃保存,蒸馏水清洗后取样镜检。通过网格线相交法[17]计算AMF侵染率,以根段上出现孢子或丛枝为标准。在每次取样时,每个样树至少观察50小段根样,每个处理共观察250小段根样。在侵染率达到最高时,取样测定叶片内营养元素、营养物质和次生代谢产物含量,以及防御蛋白活性。
1.3.2 N、P元素测定
每个处理组随机选取9株样树,每3株一个重复,测3个重复。每株选取第3、第4片子叶进行测定,下同。N元素采用奈氏比色法,测定P元素采用钼锑抗比色法[18]。
1.3.3 营养物质测定
可溶性蛋白质含量采用考马斯亮蓝法[19]测定,可溶性糖和淀粉含量采用蒽酮比色法[20]测定。
1.3.4 防御蛋白活性及次生代谢物质含量测定
PAL采用苯丙氨酸比色法[20],PPO采用邻苯二酚比色法[21]。TI和CI依据孟昭军[20]和段文昌等[22]方法测定。总酚采用福林酚比色法[23]测定,单宁依据范旭东[24]的方法测定,黄酮含量依据Jia等[25]的方法测定,木质素含量依据任琴等[26]的方法测定。
1.4 数据处理与分析
使用Excel2010统计数据的平均值和标准误差。使用SPSS22.0 one-way ANOVA进行差异显著性方差分析,多重比较采用LSD法在0.05水平下检验各组之间的差异显著性。
2. 结果与分析
2.1 GM和GI侵染率随时间的变化趋势
在处理30、60、90、100 d后GM和GI侵染率随时间变化如图1所示。随着处理时间的延长,侵染率呈显著升高,且GM和GI处理组之间差异显著(P < 0.05)。在30 d时,GM侵染率为10.8%,GI为15.6%;60 d时,GM为20.1%,GI为30.5%;在90 d时侵染率达到最大值,GM为45.6%,GI为61.7%;在100 d时侵染率降低,GM为43.7%,GI为60.8%。CK组没有检测到侵染。
![]() 图 1 GM、GI处理组侵染率随时间变化的趋势CK.对照;GM.摩西球囊霉;GI.根内球囊霉。数据均为平均值 ± 标准差(n = 3);不同小写字母表示处理组与对照组之间差异显著(P < 0.05)。下同。CK, control; GM, Glomus mosseae; GI, Glomus intraradices. The data annotation in the picture is average value ± SD (n = 3); different lowercase letters mean significant differences between treatment group and control group (P < 0.05). The same below.Figure 1. Trend of infection rate of GM and GI with time
图 1 GM、GI处理组侵染率随时间变化的趋势CK.对照;GM.摩西球囊霉;GI.根内球囊霉。数据均为平均值 ± 标准差(n = 3);不同小写字母表示处理组与对照组之间差异显著(P < 0.05)。下同。CK, control; GM, Glomus mosseae; GI, Glomus intraradices. The data annotation in the picture is average value ± SD (n = 3); different lowercase letters mean significant differences between treatment group and control group (P < 0.05). The same below.Figure 1. Trend of infection rate of GM and GI with time2.2 GM和GI处理叶片中N、P 及营养物质的含量
银中杨接种GM、GI 90 d后叶片内N、P及营养物质含量如图2所示。GM和GI处理组叶片N、P元素含量显著高于CK组(P < 0.05)。P元素含量,GI组显著高于GM组(P < 0.05);N元素含量,GM与GI组差异不显著(如图2A)。可溶性蛋白质含量,GM和GI处理组显著高于CK组(P < 0.05),GM和GI组间差异不显著;可溶性糖含量,GM和GI处理组显著低于CK组(P < 0.05),GM与GI处理组差异不显著;淀粉含量3组间差异均不显著(如图2B)。
![]() 图 2 GM、GI处理组叶片中营养元素(N、P)和营养物质的含量Figure 2. Contents of nutrient elements (N, P) and nutrients in leaves with GM or GI treatment
图 2 GM、GI处理组叶片中营养元素(N、P)和营养物质的含量Figure 2. Contents of nutrient elements (N, P) and nutrients in leaves with GM or GI treatment2.3 GM和GI处理叶片中次生代谢产物的含量
接种GM、GI 90 d后,银中杨叶片内次生代谢产物含量如图3所示。总酚、黄酮、木质素和单宁含量,GM和GI处理组均显著高于CK组(P < 0.05)。在2个处理组间,木质素和单宁差异显著(P < 0.05),总酚和黄酮差异不显著(P > 0.05)。
![]() 图 3 GM和GI处理组叶片中总酚、黄酮、木质素和单宁的含量Figure 3. Contents of total phenols, flavonoids, lignin and tannins in leaves of GM or GI treatments
图 3 GM和GI处理组叶片中总酚、黄酮、木质素和单宁的含量Figure 3. Contents of total phenols, flavonoids, lignin and tannins in leaves of GM or GI treatments2.4 GM和GI处理叶片中防御蛋白的活性
接种GM、GI 90 d后,银中杨叶片内防御蛋白苯丙氨酸解氨酶(PAL)、多酚氧化酶(PPO)、胰凝乳蛋白酶抑制剂(CI)和胰蛋白酶抑制剂(TI)的活性均显著大于对照(见图4)。在2个处理组间,PAL活性差异显著(见图4A),PPO、CI和TI活性差异不显著(见图4B、C、D)。
![]() 图 4 GM和GI处理组叶片中PAL、PPO、CI和TI的活性Figure 4. Activities of PAL, PPO, CI and TI in leaves of GM or GI treatments
图 4 GM和GI处理组叶片中PAL、PPO、CI和TI的活性Figure 4. Activities of PAL, PPO, CI and TI in leaves of GM or GI treatments3. 结论与讨论
本研究表明,外源接种AMF(GM或GI)可以显著提高银中杨的物质代谢和化学防御,具体表现为叶片内N、P、可溶性蛋白质含量增加,可溶性糖含量减少,次生代谢产物含量增加,防御蛋白活性增强。
研究报道,GI可以改善欧美杨107对N、P的吸收和利用,其中N吸收与蛋白质的含量变化有关[2]。Tao等[27]研究显示,用不同浓度的GM、GI和GE混合菌种处理马利筋属(Asclepias)6种植株,能显著提高其根系和叶片内的N、P含量;随着P浓度的增加,马利筋生长速率显著加快,因而对黑脉金斑蝶(Danaus plexippus)危害的耐受性显著提高;同时N含量的增加使马利筋叶片的乳胶(主要防御物质)渗出量显著提高。用GM、GI和地表球囊霉(Glomus versiforme,GV)等3种AMF分别处理黄瓜,均可以在不同程度上显著提高其对土壤基质中N、P元素的吸收能力[28]。本研究结果进一步证实了这一点。N、P在植物生长过程中起到重要的生理作用,不仅可以提高植物叶片的营养水平,而且参与氮代谢和磷代谢等生理进程,进而提高植物的耐受性[29]。本试验显示,AMF处理组可溶性糖含量显著低于对照,淀粉含量差异不显著,其原因可能是植物碳素营养物质与AMF消耗植物糖类的水平有关[30]。Bonfante和Genre研究报道,在植物生长早期AMF会起到抑制作用,可能是由于真菌的生长需要寄主植物提供糖类[31]。相关研究[32]表明,接种GE和GM促进了柑橘幼苗叶片可溶性蛋白等的积累,提高了防御蛋白的活性,进而增强植物对干旱的适应能力。在我们的研究中,可溶性糖含量降低即碳素含量降低,氮素含量升高,碳氮比降低。据报道,植物体内的碳氮比是研究抗虫性的重要指标,碳氮比的高低对不同植物的抗虫性影响不同[33-34]。Gherlenda等[35]研究发现,高浓度的CO2条件下,细叶桉(Eucalyptus tereticornis)的C/N升高,对桉树龟金花虫(Paropsis atomaria)幼虫体重显著降低,血淋巴蛋白含量显著降低。马艳等[36]研究表明,在棉铃虫(Helicoverpa armigera)幼虫取食C/N低的棉花(Gossypium spp.)后,成虫产卵量增加,雄性成虫寿命显著增加。因此,本研究中银中杨碳氮比与抗虫性的关系有待我们进一步生物测定。
次生代谢产物含量和防御蛋白活性是衡量植物抗虫性的生理指标。分月扇舟蛾(Clostera anastomosis)危害69杨(P. deltoides)和895杨(P. delotides × P.euramericana cv.‘Nanlin895’)后,诱导杨树叶片内单宁、黄酮和总酚含量显著升高,延长分月扇舟蛾的幼虫期和蛹期,显著降低其化蛹率、蛹重、羽化率和产卵量 [37]。Jiang等[38]在研究茉莉酸诱导下长白落叶松(Larix olgensis)的抗虫机制时发现,针叶内防御蛋白酶和蛋白酶抑制剂活性的增加是抑制舞毒蛾(Lymantria dispar)生长发育的主要原因之一。本研究中,在接种AMF的银中杨叶片内,次生代谢产物总酚、黄酮、木质素和单宁的含量及防御蛋白PAL、PPO、CI和TI活性均显著高于对照,进一步说明AMF诱导可以提高银中杨的化学防御。相似的研究发现,接种GI的黑吉豆(Vigna mungo)叶片中积累大量的防御代谢产物,如酚类,木质素以及防御蛋白,大大降低了斜纹夜蛾(Spodoptera litura)的取食率[39]。接种不同浓度的GM、GI和GE混合菌种能够显著提高马利筋叶片内的次生代谢产物卡烯内酯含量,使黑脉金斑蝶的生长发育和繁殖(体重、取食量和产卵量)受到抑制[27]。本试验中AMF诱导银中杨化学防御的改变是否会增强银中杨的抗虫性,有待进一步生物测定来验证。
不同AMF对寄主植物的适应性不同[40],即不同的AMF和植物共生对植食昆虫的抗性有不同的影响。在本研究中,GI对银中杨的侵染率显著高于GM,说明GI对银中杨的适应性更强。接种GI的银中杨生长发育显著高于接种GM(另文发表),GI组P含量显著高于GM组,但GM组木质素、单宁含量和PAL活性显著高于GI组。因此,在实践中,可根据生产目的选取AMF菌种。
本试验结果说明AMF能够促进银中杨的物质代谢,增强树势,改善其耐受性;提高银中杨的化学防御,改变其抗虫性。但对抗虫性是否具有积极地意义,有待我们进一步的生物测定结果来验证,从而为之后关于AMF的研究奠定理论基础。
-
![]()
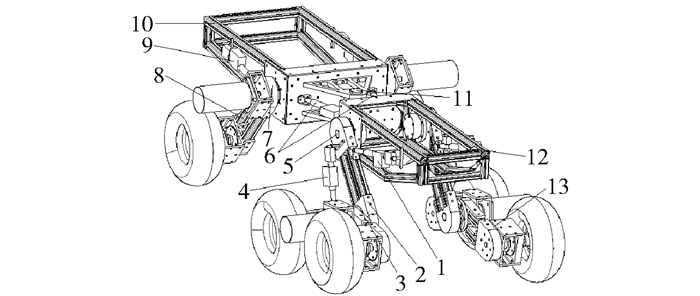
图 1 六轮变幅轮腿底盘结构示意图
1.控制缸1;2.右前轮腿; 3.右前轮“人”字形轮腿; 4.控制缸2;5.右前轮腿车体铰接点; 6.前后车架联接折腰控制缸; 7.右后轮腿车体铰接点; 8.右后轮腿; 9.右后轮腿控制缸; 10.后车架; 11.前后车架铰接点; 12.前车架; 13.左前轮腿“人”字形支腿。
Figure 1. Schematic diagram of six-wheeled-leg chassis
1, control cylinder 1; 2, right front wheeled-leg; 3, right front "人" shape wheeled-leg; 4, control cylinder 2; 5, hinged point of right front wheeled-leg; 6, control cylinder of connection point between front frame and back frame; 7, hinged point of right back wheeled-leg; 8, right back wheeled-leg; 9, control cylinder of right back wheeled-leg; 10, back frame; 11, hinged point of front frame and back frame; 12, front frame; 13, left front "人" shape wheeled-leg.
![]()
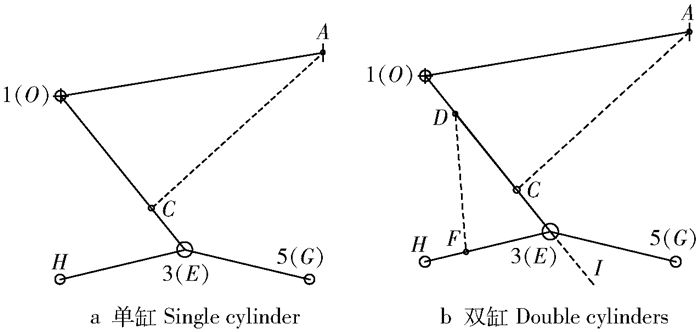
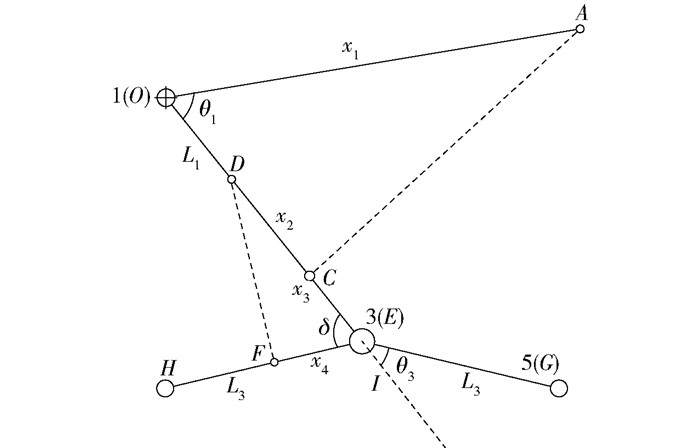
图 3 前轮腿单、双缸控制系统结构简图
A点为控制缸1车架安装点,O点为前轮腿车架铰接点,C点为控制缸1前轮腿安装点,D点为控制缸2前轮腿安装点,F点为控制缸2“人”字形轮腿安装点,E点为“人”字形轮腿铰接点,G点为“人”字形轮腿前轮轮心,H点为“人”字形轮腿后轮轮心,I点为前轮支腿延长线上点,O为基坐标系,1为前轮腿坐标系,3为“人”字形轮腿坐标系,5为“人”字形轮腿前轮轮心坐标系。
Figure 3. Structure diagram of the front wheeled-legs with single cylinder and double cylinders
A is the frame installation point of control cylinder 1, O is the frame hinged point of front wheeled-legs, C is the front wheeled-legs installation point of control cylinder 1, D is the front wheeled-legs installation point of control cylinder 2, F is the "人" shape wheeled-legs installation point of control cylinder 2, E is the hinged point of "人" shape wheeled-legs, G is the front wheel center point of "人" shape wheeled-legs, H is the back wheel center point of "人" shape wheeled-legs, I is the piont of front wheeled-legs extended line, O is the basic joint coordinate system, 1 is the joint coordinate system of front wheeled-legs, 3 is the joint coordinate system of "人" shape wheeled-legs, 5 is the front wheel center joint coordinate system of "人" shape wheeled-legs.
![]()
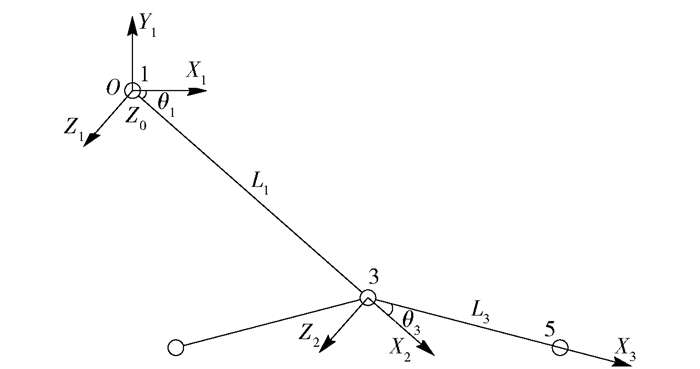
图 4 基于D-H法前轮腿坐标系
θ1为绕Z0轴的旋转角;θ3为绕Z1轴的旋转角;L1为沿X2轴平移距离;L3为沿X3轴平移距离。
Figure 4. Front wheeled-legs coordinate system based on D-H method
θ1 is the rotation angle around Z axis, θ3 is the rotation angle around Z1 axis, L1 is the translation length along the X2 axis, L3 is the translation length along the X3 axis.
![]()
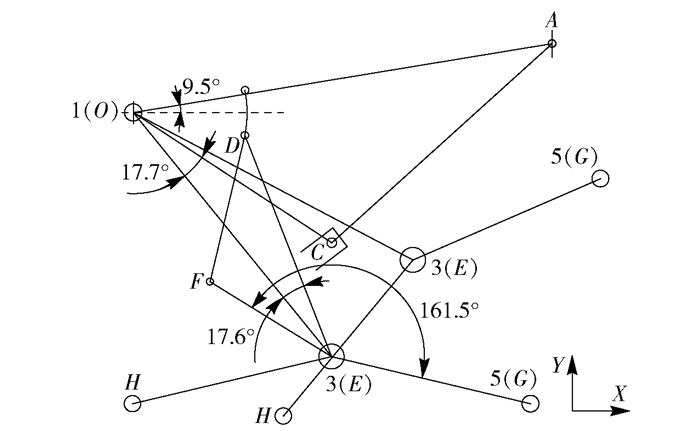
图 5 双缸控制轮腿越障过程分析示意图
Figure 5. Analysis schematic for obstacle crossing process of double-cylinder wheeled-legs
![]()
图 6 单缸控制轮腿越障过程分析示意图
Figure 6. Analysis schematic for obstacle crossing process of single-cylinder wheeled-legs
![]()
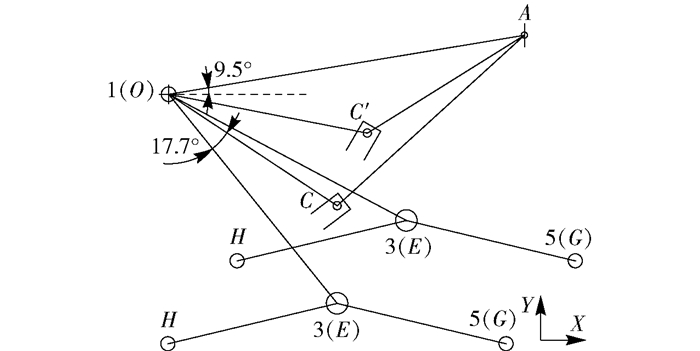
图 7 双缸最佳布置形式示意图
x1为AO的长度,x2为OC的长度,x3为DE的长度,x4为EF长度,∠AOC为θ1,∠GEI为θ3, ∠DEF为δ。
Figure 7. Best double-cylinder arrangement diagram
x1 is the length of AO, x2 is the length of OC, x3 is the length of DE, x4 is the length of EF, ∠AOC is θ1, ∠GEI is θ3, ∠DEF is δ.
![]()
图 8 当x1、x2、x3、x4变化时轮腿变幅机构实现最大越障高度时的仿真情景
Figure 8. Simulation scenarios of the mechanism achieving the maximum obstacle crossing height with x1, x2, x3, x4 changing
![]()
图 10 理论计算、仿真实验及实测实验对比
Figure 10. Comparison among theoretical calculations, simulation experiment and test experiment
表 1 D-H参数表
Table 1 D-H parameter list
连杆Link θ/(°) d/mm a/mm α/(°) 1 0 0 0 0 3 θ1 0 L1 0 5 θ3 0 L3 0 注:θ为两连杆夹角,d为两连杆距离,a为连杆长度,α为连杆扭角。Notes: θ is the angle between two connecting rods,d is the distance between two connecting rods,a is the length of connecting rod,and α is the twist angle of connecting rod.  下载: 导出CSV
下载: 导出CSV
表 2 理论数据
Table 2 Theoretical data
mm Py x1 x2 x3 x4 -285.9 550 300 406 190 -285.9 560 370 406 190 -285.9 570 390 406 190 -285.9 610 400 406 190 -285.9 650 410 406 190 -285.9 670 420 406 190 注:Py为越障高度,x1为控制缸1在车架上安装点A与前轮腿车架铰接点O之间的距离, x2为控制缸1在前轮腿上的安装点C与前轮腿车架铰接点O之间的距离, x3为控制缸2在前轮腿上的安装点D与控制缸2在“人”字形轮腿铰接点E之间的距离, x4为控制缸2在“人”字形轮腿上的安装点F与“人”字形轮腿铰接点E之间的距离。Notes: Py is the obstacle crossing height, x1 is the distance between installation point A on the body and hinged point O of front wheeled-legs on the body, x2 is the distance between installation point C of control cylinder 1 on the front wheeled-legs and hinged point O of front wheeled-legs on the body, x3 is the distance between installation point D of control cylinder 2 on the front wheeled-legs and hinged point E of control cylinder 2 on the “人” shaped wheeled-legs, x4 is the distance between installation point F on the “人” shaped wheeled-legs of control cylinder 2 and hinged point E of “人” shaped wheeled-legs.
下载: 导出CSV
表 3 仿真数据
Table 3 Simulation data
mm Py x1 x2 x3 x4 -782.1 350 190 480 270 -611.6 420 230 470 260 -598.3 480 260 465 243 -492.1 490 260 425 217 -371.2 500 270 421 200 -312.9 520 270 413 190 -283.6 550 280 409 195 -288.3 560 370 409 190 -281.7 570 390 409 190 -286.1 610 400 405 190 -285.7 650 410 405 190 -289.0 670 420 400 190 -315.5 680 430 400 190 -350.1 680 430 390 170 -562.3 690 440 350 150 -671.6 690 460 290 120
下载: 导出CSV
表 4 实测数据
Table 4 Actual measurement data
mm Py x1 x2 x3 x4 -791.5 300 300 500 270 -772.9 350 400 490 260 -621.3 390 426 470 233 -588.6 430 426 420 226 -369.5 500 431 410 203 -312.8 550 300 409 192 -285.3 560 370 409 185 -294.5 570 390 407 195 -286.2 610 400 409 193 -287.0 650 410 406 188 -283.9 670 420 405 190 -315.8 680 435 405 190 -566.1 690 433 350 155 -701.9 700 435 290 126 -730.6 700 430 290 120
下载: 导出CSV
-
[1] 迟宝山, 权清达, 刘西侠, 等.轮腿式行走系统研究综述[J].农业装备与车辆工程, 2013, 51(6): 46-49. doi: 10.3969/j.issn.1673-3142.2013.06.014 Chi B S, Quan Q D, Liu X X, et al. Research on wheel-legged walking system[J]. Agricultural Equipment & Vehicle Engineering, 2013, 51(6): 46-49. doi: 10.3969/j.issn.1673-3142.2013.06.014
[2] 谢霞, 康少华, 侍才洪.复杂地形移动底盘技术研究[J].工程机械, 2015, 46(3): 45-52. doi: 10.3969/j.issn.1000-1212.2015.03.010 Xie X, Kang S H, Shi C H. Study on mobile chassis traveling in complex terrains[J]. Construction Machinery and Equipment, 2015, 46(3): 45-52. doi: 10.3969/j.issn.1000-1212.2015.03.010
[3] 刘凯.仿人机器人运动控制系统和步态规划算法的研究及实现[D].上海: 上海交通大学, 2010. Liu K. Motion control and gait planning for humanoid robot[D]. Shanghai: Shanghai Jiao Tong University, 2010.
[4] 李涛.机器人规划与智能控制的研究及其仿真[D].淮南: 安徽理工大学, 2009. Li T. Research and simulation about robot planning and intelligent control[D]. Huainan: Anhui University of Science and Technology, 2009.
[5] 赵之光.基于SVM的双足机器人步态自学习控制方法[D].广州: 广东工业大学, 2012. Zhao Z G. Research of biped robot gait self-learning control algorithms based on SVM[D]. Guangzhou: Guangdong University of Technology, 2012.
[6] 孟廷豪.基于模糊控制的机器人避障研究[D].太原: 中北大学, 2013. Meng T H. The study of obstacle avoidance system based on fuzzy control for mobile robot[D]. Taiyuan: North University of China, 2013.
[7] 江道根.六自由度并联机器人RBF神经网络PID控制研究[D].镇江: 江苏大学, 2010. Jiang D G. Study on RBFNN PID control for a 6-DOF parallel robot[D]. Zhenjiang: Jiangsu University, 2010.
[8] Han D T, Liu J H, Kan J M, et al. Analysis of a kinematic model for a forestry six-wheeled luffing articulatedvehicle chassis[J]. Open Mechanical Engineering Journal, 2015, 9(1): 670-677. doi: 10.2174/1874155X01509010670
[9] 蔡自兴.机器人学基础[M].北京:机械工业出版社, 2009: 6. Cai Z X. Fundamentals of robotics[M]. Beijing:Engineering Industry Press, 2009: 6.
[10] 于靖军, 刘辛军, 丁希仑, 等.机器人机构学的数学基础[M].北京:机械工业出版社, 2008: 7. Yu J J, liu X J, Ding X L, et al. Mathematical foundations of robotic mechanisms[M]. Beijing: Engineering Industry Press, 2008: 7.
[11] Craig J J.机器人学导论[M].北京:机械工业出版社, 2006. Craig J J. Introduction to robotics mechanics and control[M]. Beijing: Engineering Industry Press, 2006.
[12] 宋小康, 谈大龙, 吴镇炜, 等.全地形轮式移动机器人运动学建模与分析[J].机械工程学报, 2008, 44(6): 148-154. doi: 10.3321/j.issn:0577-6686.2008.06.023 Song X K, Tan D L, Wu Z W, et al. Kinematics modeling and analyses of all-terrain wheeled mobile robots[J]. Chinese Journal of Mechanical Engineering, 2008, 44(6): 148-154. doi: 10.3321/j.issn:0577-6686.2008.06.023
[13] 于涌川, 原魁, 邹伟.全驱动轮式机器人越障过程模型及影响因素分析[J].机器人, 2008, 30(1): 1-6. doi: 10.3321/j.issn:1002-0446.2008.01.001 Yu Y C, Yuan K, Zou W. Dynamic model of all-wheel-drive mobile robot climbing over obstacles and analysis on its influential factors[J]. Robot, 2008, 30(1): 1-6. doi: 10.3321/j.issn:1002-0446.2008.01.001
[14] 魏巍.基于区间分析的轮式摆动车身车辆越野性能研究[D].长春: 吉林大学, 2012. Wei W. Research on off-road performance of wheeled vehicle with oscillating body based on interval analysis[D]. Changchun: Jilin University, 2012.
[15] 韩军, 陈高杰, 王红坚, 等.步行式挖掘机越障能力分析[J].中国工程机械学报, 2005, 3(1): 25-28. doi: 10.3969/j.issn.1672-5581.2005.01.006 Han J, Chen G J, Wang H J, et al. Climbing ability analysis for walking mobile excavator[J].Chinese Journal of Construction Machinery, 2005, 3(1): 25-28. doi: 10.3969/j.issn.1672-5581.2005.01.006
-
期刊类型引用(2)
1. 孙榕泽,宋焕芝,蒋珈琦,于晓南. 三倍体芍药品种染色体制片优化和核型分析. 河北农业大学学报. 2024(03): 46-55 .  百度学术
百度学术
2. 段英姿,客绍英,王晓英,张胜珍,马艳芝. 15个油用紫斑牡丹品种核型及亲缘关系分析. 种子. 2023(07): 96-104 . 百度学术
其他类型引用(0)








计量
- 文章访问数: 1918
- HTML全文浏览量: 472
- PDF下载量: 24
- 被引次数: 2
