
Production of hybrid triploids via inducing chromosome doubling of megaspore with high temperature treatment in Leuce poplar
-
摘要:目的鉴于杨树三倍体具有突出的营养生长优势,针对毛白杨良种选育中母本效应难以充分利用问题,选择毛白杨育性好的雌株诱导大孢子染色体加倍选育白杨杂种三倍体,为毛白杨三倍体育种提供技术支撑。方法本研究以毛白杨无性系MC1和MC2为母本,银腺杨YX1为父本。根据白杨大小孢子发生发育存在相关性的原理,以雄株小孢子发育进程为即时判别大孢子发生进程的参照,当小孢子发育至单核早期、单核靠边期和双核期时,分别施加38、40和42 ℃高温持续处理雌花序4 h。结果两个杂交组合的处理组共得到189株毛白杨杂种三倍体,其中MC1×YX1组合中共获得70株杂种三倍体,平均诱导率为56.45%;MC2×YX1组合中共获得119株杂种三倍体,平均诱导率为47.04%,对照组未获得杂种三倍体。当小孢子发育至单核靠边期,对应大孢子母细胞减数分裂至粗线期至中期Ⅰ,此时施加38和40 ℃高温处理4 h是毛白杨大孢子染色体加倍的最佳处理时期。两个杂交组合的杂种三倍体在平均苗高和地径生长方面均优于二倍体,但组合间存在一定差异,表明三倍体育种同样需要重视亲本选配。结论研究进一步证明高温诱导大孢子染色体加倍是一种高效获得毛白杨杂种三倍体的技术途径,相关技术进步对于推进毛白杨良种选育进程具有重要的意义。Abstract:ObjectiveTriploid poplars have been proved to be characterized with outstanding advantages in vegetative growth over diploids. However, it still has some difficulties to take full advantage of maternal effect in present P. tomentosa breeding work. By careful selection of female parents with good fertility, this study aims to produce hybrid triploids by megaspore chromosome doubling in P. tomentosa, which would provide technical support for triploid breeding in further research.MethodThe P. tomontosa clones MC1 and MC2 were served as the female parents, and the hybrid clone YX1 of P. alba×P. glandulosa was selected to be the male parent. According to the correlation between megasporocytes and microsporocytes during meiosis, the meiotic stage of megaspore in female flowers could be determined by the meiotic stage of microspore in male flowers. When the microspore developed into early stage of single nucleus, late uninucleate stage and binucleate stage, the female catkins were treated for 4 hours with high temperatures of 38, 40 and 42 ℃ to induce chromosome doubling of megaspore, respectively.ResultA total of 189 triploids were obtained in the two hybrid combinations. Among all the 189 triploids, 70 triploids were obtained from the cross combination MC1×YX1, and the average induction rate was 56.45%. 119 triploids were obtained from the combination MC2×YX1, and the mean induction rate was 47.04%. No triploids were achieved in the control groups. When the microspores developed into the uninucleate eccentric stage, the corresponding meiotic stage of macrospore mother cell was at the stages from pachytene to metaphase I, which could be the optimal treating period for inducing megaspore chromosome doubling by 38 and 40 ℃ high temperature treatment. The average seedling height and the ground diameter of triploid hybrids derived from two cross combinations were higher than that of the diploids, amid there were some differences between the two combinations, which suggested that some attentions should be paid to the parental selection in triploid breeding of P. tomentosa.ConclusionOur findings reveal that induction of megaspore chromosome doubling by high temperature treatment is an effective way to produce triploid hybrids in P. tomentosa, and the progress of breeding technologies plays an important role in genetic improvement of P. tomentosa.
-
Keywords:
- Populus tomentosa /
- megaspore /
- chromosome doubling /
- triploid /
- induction rate
-
物种保护不仅需要维持种群数量,还需保护种群的生活环境,而了解种群的分布范围是保护其生活环境的关键前提和基础[1]。生境适宜性评价是以野生动物栖息地环境为基础,分析研究动物分布区域内各种环境因子的状态,评估与筛选影响野生动物个体生存和种群繁衍的关键环境因子,明确研究范围内适宜动物生存、分布的空间,有助于提升保护管理策略。因此,开展野生动物生境适宜性科学评价,对物种保护和栖息地管理具有重要意义[2]。
中华斑羚(Naemorhedus griseus)简称斑羚,隶属于偶蹄目(Artiodactyla)、牛科(Bovidae)、斑羚属(Naemorhedus),主要分布在我国华北、华南、西南地区和印度、缅甸、泰国、越南[3]。斑羚被列入《濒危野生动植物国际贸易公约(CITES)》附录Ⅰ中,各地区种群均受到栖息地破碎化分割,数量持续下降,在世界自然保护联盟(IUCN)红色名录和《中国脊椎动物红色名录》中被评估为易危(VU)物种[4-5]。内蒙古赛罕乌拉国家级自然保护区(以下简称赛罕乌拉)是被社区包围的生物多样性孤岛,随着人口不断增加,人类活动逐渐加剧,斑羚栖息地破碎化风险日趋严重。因此,对赛罕乌拉斑羚生境适宜性评价有助于了解其栖息地的质量和空间分布,为斑羚种群的保护、管理和恢复提供科学依据。
近年来,随着3S(GIS、RS、GPS)技术的发展,在大尺度下对野生动物生境适应性评价成为热点[6-7]。以GIS为核心的3S技术的结合与集成,实现了对栖息地空间变量数据的实时采集、处理和分析,为野生动物栖息地规划、自然保护区地理分布提供空间信息支持和管理决策服务。该技术综合野生动物栖息地环境变量、出现点等信息,不仅能绘制物种分布图、模拟物种的空间分布格局,还可以研究物种分布动态变化,为野生动物管理者制定保护方案提供重要科学参考。目前有5种使用比较广泛的预测物种分布的生态位模型,包括箱盒环境包络(boxcar environmental envelope,BIOCLIM)、CLIMEX、Gower距离(Gower’s distance,DOMAIN)、基于遗传算法的规则组合模型(genetic algorithm for ruleset production,GARP)和最大熵模型(maximum entropy model,MaxEnt)[8-9]。王运生等通过接收者操作特征曲线(receiver operator characteristic curve,ROC)对这些模型的预测结果分析发现,以MaxEnt的ROC曲线下面积值(Areas under curve,AUC)最大,表明其预测效果最好[10-12]。同时,国内外研究学者已利用该模型对大熊猫(Ailuropoda melanoleuca)[13-14]、雪豹(Panthera uncia)[15]、白唇鹿(Przewalskium albirostris)[16]、朱鹮(Nipponia nippon)[17]等濒危珍稀野生动物的生境栖息地进行评价分析,并提出了相应的保护管理措施。
本研究应用3S技术和MaxEnt模型对赛罕乌拉孤立的斑羚小种群生境适宜性进行评价,分析影响斑羚生存的重要生境因子,制作栖息地分布图,同时探讨干扰因子(护林站、居民点、公路、铁路)对斑羚生境适宜性的影响,以期为赛罕乌拉斑羚种群及其栖息地的保护与恢复提供参考。
1. 研究区概况与研究方法
1.1 研究地概况
赛罕乌拉国家级自然保护区位于内蒙古自治区赤峰市巴林右旗北部,东与巴林左旗的石棚沟林场相连,西与林西县交界,南靠巴林右旗幸福之路苏木,北与锡林郭勒盟接壤,主要以多样性的珍稀濒危野生动植物及其栖息地,丰富的湿地、草原、森林等生态系统为保护对象,总面积约10万hm2[18](图 1)。该保护区地处大兴安岭南麓,是草原向森林、大兴安岭山地针叶林向阔叶林过渡的交替地带,还是中国动物地理区划中东北区、华北区、蒙新区动物类型的重要交汇区域,典型的地理位置和自然条件丰富了赛罕乌拉保护区的动植物区系。全区记录哺乳类物种6目16科44种,其中包括猞猁(Lynx lynx)、兔狲(Felis manul)、马鹿(Cervus elaphus)、中华斑羚4种国家Ⅱ级重点保护野生动物,鸟类235种,分属19目48科,有国家Ⅰ级重点保护种类3种:黑鹳(Ciconia nigra)、金雕(Aquila chrysaetos)、大鸨(Otis tarda)[19-21]。
1.2 模型分析方法
MaxEnt是一种基于生态位原理,由Phillips等开发,用于评价、预测动物栖息地分布的模型[22]。模型利用物种“出现点”的GPS数据,分析得出研究区域内物种分布位点的各种环境变量因子的约束条件,探寻与约束条件相似的环境像元,以此得到目标物种在研究区域内的栖息地分布[23-25]。模型具有良好的预测能力,广泛应用于目标物种在研究区域的生境评价、预测其适宜和潜在分布空间、气候变化对物种分布的影响和入侵物种监测等多方面内容[26]。模型采用受试者工作特征曲线(ROC)及其曲线下面积(AUC)对预测结果的精度进行验证,在排除空间自相关的情况下,数值越大,体现环境因子变量与模型的吻合度越高,显示模型分析预测结果越好。同时,通过创建响应曲线来确定各个环境因子对模型预测的作用范围,曲线横坐标表示环境因子的变化区间,纵坐标表示环境因子对生境适宜性的贡献率,其数值越大,表明栖息地适宜性越高[14]。SDM(species distribution model)是基于一系列的python语言脚本开发的物种分布模型,利用SDM toolbox工具中的环境变量空间自相关分析功能,可以剔除高度自相关的环境变量和位点,提高MaxEnt模型的应用效率[27-28]。
1.3 数据来源及预处理
斑羚分布点数据源于本研究野外样线调查以及红外相机监测(图 1)。调查范围覆盖斑羚栖息地全部生境类型,保证了分布点的代表性和准确性。同时,使用“Spatially Rarely Occurrence Data for SDMs”工具剔除重复位点、排除数据间聚集性后,共筛选出42个位点(图 1)。环境因子包括:地形变量数据(包括海拔高差、坡度和坡向,由DEM数据获取(本研究区域30m分辨率),来自:http://www.gscloud.cn/); 归一化植被指数(normalized difference vegetation index,NDVI)为2016年MYD13Q1的16d合成产品(空间分辨率为250m,来自:同上); 气候数据来自世界气候数据库WorldClim网站(http://worldclim.org/)中的19个气候因子Bio1~Bio19,分辨率为1km; 植被类型数据来自Landsat8遥感影像解译(https://glovis.usgs.gov/),可分为乔木林、灌木、草地、裸地4类,分辨率为30m;保护区内及周边的道路和居民点等相关图层数据来源于内蒙古赛罕乌拉自然保护区管理局,以欧式距离分析得到研究区域各个像元距公路、铁路、居民点及护林站距离等干扰因子[17]。
环境变量均以ArcGIS10.2为平台,将所有环境变量的图层统一边界,去除边界外的GPS坐标点,坐标系统一为WGS-1984-UTM-Zone-50N坐标,栅格统一为30m×30m,转化为MaxEnt识别的ASCⅡ文件格式。
1.4 分析过程
利用SDM工具包中的Raster Correlations and Summary Statistics工具,剔除相关性过高(|r|≥0.75)的环境变量[16, 29]。最终得到11个环境变量(坡向、坡度、海拔、归一化植被指数NDVI、植被类型、距公路距离、距铁路距离、距居民点距离、距护林站距离、月平均昼夜温差、最干月降水量)。将上述环境变量和斑羚的分布位点代入MaxEnt模型,验证模型时随机选择25%的分布位点,构建模型时使用其余的75%分布位点,创建每个环境因子的响应曲线,以此评价每个环境因子对模型的贡献率。设置模型运行次数为10次,其他参数的选定均设为模型自动默认值,计算结果以Logistic格式显示输出。利用AUC检验模型分析预测效果,其值在0~1,其中0.5~0.6为不合格; 0.6~0.7为差; 0.7~0.8为普通; 0.8~0.9为良; 0.9~1为优秀。将模型输出结果导入ArcGIS中进一步分析,采用灵敏度和特异度之和最大值(maximized training sensitivity plus specificity)作为阈值,对模型预测的赛罕乌拉斑羚分布图进行重分类,将大于阈值的像元划分为斑羚的适宜生境[30]。
2. 结果与分析
2.1 MaxEnt预测结果
ROC曲线评价结果AUC值平均值为0.967±0.019(均值±标准差),表明MaxEnt模型的预测结果达到优秀水平(图 2)。
2.2 斑羚分布与环境变量间的关系
各环境变量对模型的综合贡献率结果表明,月平均昼夜温差对模型的贡献率最高,贡献率为23.2%,最干月降水量次之,贡献率为22.8%,海拔、距护林站距离、距居民点距离、归一化植被指数的贡献率分别为17.8%、11.8%、8.4%和7.4%;前6个环境变量的累计贡献率达91.4%,表明这6个环境变量基本解释了斑羚生境适宜性。距公路距离、距铁路距离、坡度、坡向以及植被类型对斑羚栖息地选择影响较小,贡献率分别为2.5%、2.1%、2.0%、1.3%、0.7%。在干扰因子变量中,距护林站、居民点距离变量的贡献率明显高于距公路、铁路距离这两个环境变量,表明护林站和居民点对斑羚生境选择的影响高于公路和铁路。
从前6个斑羚生境选择的主要环境因子响应曲线可以看出(图 3),随月平均昼夜温差的增加,斑羚的出现概率逐渐降低; 斑羚生存受最干月降水量影响明显,最干月降水量上升到1.5mm时,斑羚的出现概率迅速增大,达到峰值1.5mm之后逐渐减小; 适宜斑羚生存的海拔范围在1300~1700m,海拔过高或者过低都不利于斑羚生存; 干扰因子方面,距护林站、居民点的距离对斑羚出现概率的影响呈单峰型,表明斑羚的适宜生境与人类活动保持着一定距离; 归一化植被指数达到0.15时,植被覆盖度的贡献率增加缓慢(图 3)。
![]() 图 3 环境变量对模型预测的响应曲线10次运行结果均值(红色)标准差(蓝色)The mean result with 10 replicate runs(red) and the standard deviation (blue)Figure 3. Response curves of environmental factors to model prediction
图 3 环境变量对模型预测的响应曲线10次运行结果均值(红色)标准差(蓝色)The mean result with 10 replicate runs(red) and the standard deviation (blue)Figure 3. Response curves of environmental factors to model prediction2.3 斑羚适宜生境范围
在MaxEnt模型运算结果中,选择灵敏度和特异度之和的最大值0.276为阈值,对斑羚分布预测图进行重分类,最终得到赛罕乌拉保护区斑羚生境适宜性分布图。结果表明,斑羚的有效生境适宜面积为4891.23hm2,不适宜生境的面积为101718.09hm2,分别占保护区总面积的4.59%和95.41%(图 4a)。不考虑公路、铁路、居民、护林站等干扰因子情况下,灵敏度和特异度之和的最大值为0.12,以此为阈值进行重分类,得到潜在生境适宜面积为17576.37hm2,不适宜生境的面积为89032.95hm2,分别占保护区总面积的16.49%和83.51%(图 4b)。从图 4a可以看出,存在干扰因子情况下,斑羚主要适宜区分布在保护区的中南部,这与现实已观测到的结果相符。
![]() 图 4 赛罕乌拉国家级自然保护区斑羚适宜生境分布Figure 4. Suitable habitat distribution of N. griseus in Saihanwula National Nature Reserve
图 4 赛罕乌拉国家级自然保护区斑羚适宜生境分布Figure 4. Suitable habitat distribution of N. griseus in Saihanwula National Nature Reserve3. 讨论
物种栖息地中包含诸多环境变量,但只有少数因子起着重要作用,若简单地将全部因子同等对待,容易弱化关键因子的生态作用[17, 31]。因此,本研究使用SDM工具对环境变量进行自相关分析,最终从25个环境变量中筛选出11个变量用于建模,其中月平均昼夜温差、最干月降水量、海拔、距护林站距离、距居民点距离、归一化植被指数对斑羚的分布有重要影响。两个气候因子的贡献率位居前列,这可能是由北方地区寒冷、干旱等严苛的自然条件造成的。
野外实地调查与模型预测海拔因子的响应曲线结果相符(图 3),海拔过高或者过低都不适合斑羚生存,适宜其生存的栖息地不是保护区内海拔最高之处。这些地区多为更新世冰川侵蚀形成的U型谷、山体裸岩等冰川遗迹,地势陡峭,植被覆盖度低。斑羚常生活在这样的山体裸岩周围,利用陡峭的地势来躲避捕食者。归一化植被指数响应曲线表明,NDVI达到0.15(低植被覆盖度)后,斑羚出现概率增加缓慢,与中等覆盖度(NDVI=0.3~0.5)区域斑羚出现概率相当。出现该现象的可能原因在于U型谷两壁和裸岩根部具有遮风、挡雨,躲避恶劣气候的作用,实际观察也发现斑羚有经常利用这些地方卧息的现象,同时这些区域靠近裸岩,便于其利用险峻的地势逃生,此外,斑羚有在裸岩上晒太阳以获取能量的行为,而裸岩山体为斑羚提供了开阔的视野,更容易及时发现敌害,由此可见,斑羚善于利用低植被覆盖的裸岩区域躲避捕食者。NDVI指数间接表征了食物资源的丰富程度[16],斑羚选择利用中等植被覆盖度区域觅食,以至在不同等级的植被覆盖度中斑羚的出现率相当(图 3)。由此分析,斑羚对适宜生境的选择可能与觅食利益和捕食风险的权衡有关。
野生动物对干扰因子具有回避效应,不同类型的道路干扰(公路、铁路)存在差异[31-32]。本研究结果表明斑羚对干扰因子也具有回避效应,但对不同类型的道路干扰差异不明显,可能原因在于铁路靠近部分公路。栖息地距护林站、居民点距离的贡献率明显高于距公路、铁路的距离,即护林站和居民点对斑羚栖息地的影响高于公路和铁路。出现该现象的原因在于本研究的道路数据只包括乡道及以上级别公路、铁路,缺乏如防火道、便道等低等级道路(泥路)数据,而保护区内的部分护林站深入林区、居民点靠近缓冲区,远离主干道路(乡道及以上等级道路),距离斑羚栖息地相对更近,因此使得护林站和居民点对斑羚的影响较大。同时,本研究表明人为干扰因子对斑羚栖息地影响较大,无干扰因子情况下斑羚适宜生境面积将在现有基础上扩大2.59倍,由此分析,如果增加林区内防火道路及山间小道等干扰变量,斑羚生境适宜面积可能会进一步缩小。因此,建议管控无关人员进入保护区,以降低人为干扰的负面影响。
模型预测结果与野外调查基本相符,除保护区中南部外,其他地方鲜有适宜斑羚生存的栖息地(图 4a)。当不考虑干扰因子时(图 4b),适宜区范围在原有基础增加有所扩大,同时在保护区南部区域新增两个孤立的适宜区,据保护区管理人员介绍,曾经在该地区救助过斑羚。由此可见,该区域应该是潜在适宜生境,但随人类活动的加剧,斑羚也远离了该区域。此外,保护区西部与北部高海拔区域也出现斑羚潜在适宜区,但该区域面积小且破碎化严重、受人为干扰强烈,现阶段不利于斑羚生存。因此,建议在该区域建立生物廊道,扩大斑羚的适宜栖息地。
本研究存在一些不足之处,如使用了分辨率相对粗糙的气候数据,难免导致预测结果会产生一定误差。但模型模拟结果与野外调查大致相符,因此模型预测结果具有参考价值,能体现斑羚在保护区内的实际分布状况,对保护区制定斑羚栖息地保护措施具有指导作用。
-
![]()
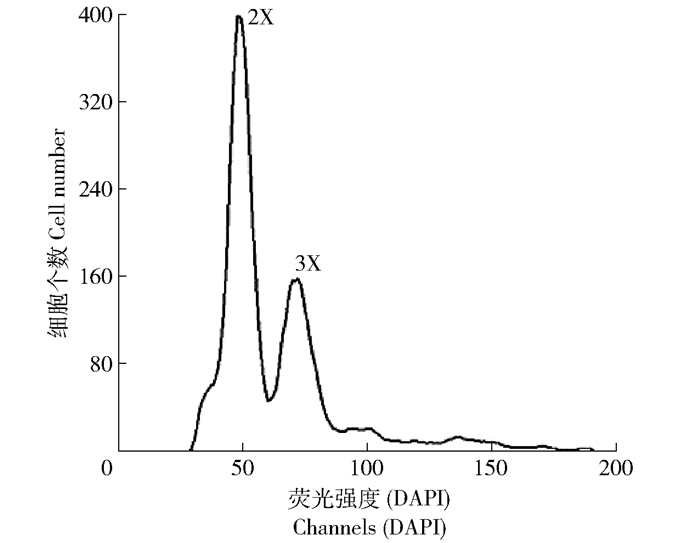
图 1 毛白杨×银腺杨杂种三倍体倍性鉴定
三倍体与二倍体植株叶片细胞混合液流式细胞仪检测结果,其中2X表示二倍体,3X表示三倍体。
Figure 1. Ploidy identification of P. tomentosa×(P. alba × P. glandulosa) hybrid triploids
Flow cytometric detection results of mixed foliage cell liquor of diploid and triploid plants, in the picture, 2X represents diploid and 3X represents triploid.
![]()
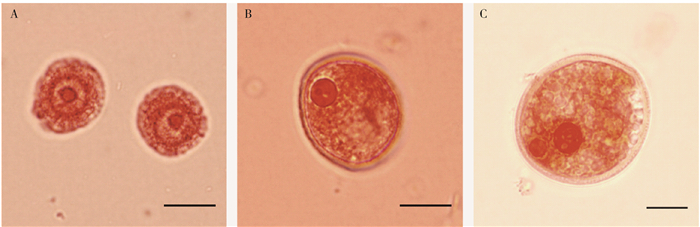
图 2 毛白杨参照雄株MX1、MX2小孢子发育阶段显微观察
A.单核小孢子早期;B.单核靠边期;C.双核期。比例尺=5 μm。
Figure 2. Microscopical observation on the microspore development stage of reference male MX1 and MX2 of P. tomentosa
A, early stage of single nucleus; B, late uninucleate stage; C, binucleate stage. Bar=5 μm.
![]()
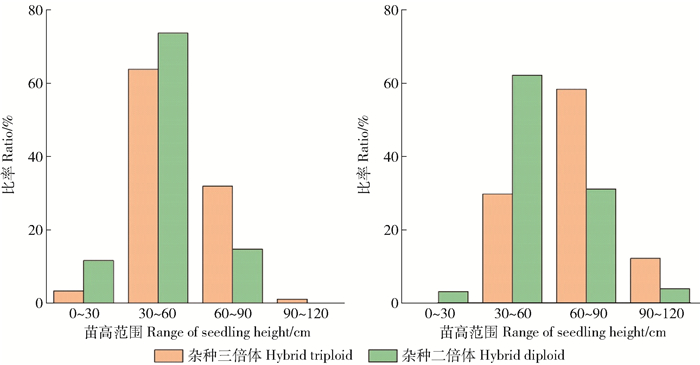
图 3 毛白杨MC2×YX1一年生苗木苗高地径对比
Figure 3. Comparison diagram of height and diameter of 1-year-old P. tomentosa×(P. alba × P. glandulosa) seedlings
表 1 高温诱导大孢子染色体加倍获得毛白杨MC1×YX1三倍体
Table 1 Triploid P. tomentosa MC1× (P. alba×P. glandulosa) YX1 produced by inducing the chromosome doubling of megaspore with high temperature treatment
小孢子发育状态
Microspore development status处理温度Treatment temperature/℃ 处理时长Treatment duration/h 种子粒数Seed amount 成苗株数Seedling amount 三倍体株数Triploid amount 三倍体得率Triploid yieldrate/% 小孢子单核早期
Early stage of single nucleus38 4 81 9 7 78.78 40 38 1 0 0 42 143 18 15 83.33 单核靠边期
Late uninucleate stage38 4 659 2 2 100 40 758 86 45 52.33 42 0 0 0 0 双核期
Binucleate stage38 4 96 0 0 0 40 281 4 0 0 42 93 4 1 25.00 对照Control 823 67 0 0 合计Total 2 972 191 70  下载: 导出CSV
下载: 导出CSV
表 2 高温诱导大孢子染色体加倍获得MC2×YX1三倍体
Table 2 Triploid P. tomentosa MC2×(P. alba × P. glandulosa) YX1 produced by inducing the chromosome doubling of megaspore with high temperature treatment
小孢子发育状态Microspore development status 处理温度Treatment temperature/℃ 处理时长Treatment duration/h 种子粒数Seed amount 成苗株数Seedling amount 三倍体株数Triploid amount 三倍体得率Triploid yieldrate/% 小孢子单核早期Early stage of single nucleus 38 4 52 12 8 66.67 40 58 3 3 100 42 50 11 11 100 单核靠边期Late uninucleate stage 38 4 573 60 44 73.33 40 571 73 40 54.79 42 57 14 11 78.57 双核期Binucleate stage 38 4 456 61 2 3.28 40 203 19 0 0 42 102 0 0 0 对照Control 744 11 0 0 合计Total 2 866 264 119
下载: 导出CSV
表 3 毛白杨×银腺杨杂种二三倍体一年生苗木生长状况分析
Table 3 Analysis on growth status of 1-year P. tomentosa × (P. alba × P. glandulosa) diploid and triploid seedlings
项目Item MC1×YX1 MC2×YX1 苗高Seedling height/cm 相对值Relative value/% 地径Ground diameter/cm 相对值Relative value/% 苗高Seedling height/cm 相对值Relative value/% 地径Ground diameter/cm 相对值Relative value/% 三倍体平均值Triploid average 69.17±23.43 124.86 8.86±2.52 143.13 55.24±16.17 115.78 6.99±1.71 126.17 二倍体平均值Diploid average 55.40±17.73 100.00 6.19±1.56 100.00 47.71±14.34 100.00 5.54±1.60 100.00 三倍体变异系数Triploid coefficient of variation/% 33.87 28.44 29.27 24.46 二倍体变异系数Diploid coefficient of variation/% 32.00 25.20 30.06 29.63 t检验t test t=2.942** t=6.822** t=3.637** t=6.398** 注:**表示差异极显著(P<0.01)。Note: ** means extremely significant difference (P<0.01).
下载: 导出CSV
-
[1] 康向阳. 毛白杨细胞遗传与三倍体选育[D]. 北京: 北京林业大学, 1996. Kang X Y. Study on cytogenetic and allotriploid breeding of Populus tomentosa[D]. Beijing: Beijing Forestry University, 1996.
[2] 叶培忠.白杨繁殖育种[J].林业科学, 1955, 1(1): 37-46. http://www.cnki.com.cn/Article/CJFDTOTAL-BJLY201607001.htm Ye P Z. Poplar breeding[J]. Scientia Silvae Sinicae, 1995, 1(1): 37-46. http://www.cnki.com.cn/Article/CJFDTOTAL-BJLY201607001.htm
[3] 朱之悌.全国毛白杨优树资源收集、保存和利用的研究[J].北京林业大学学报, 1992, 14(增刊3): 1-25. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=e121682d-3db3-4dc8-a997-bc9deb141e1c Zhu Z T. Collection, conservation and utilization of plus tree resources of Populus tomentosa in China[J]. Journal of Beijing Forestry University, 1992, 14(Suppl. 3): 1-25. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=e121682d-3db3-4dc8-a997-bc9deb141e1c
[4] 林惠斌, 朱之悌.毛白杨杂交育种战略的研究[J].北京林业大学学报, 1988, 10(3): 97-101. http://www.cnki.com.cn/Article/CJFDTOTAL-BJLY198803014.htm Lin H B, Zhu Z T. Studies on breeding strategies of Populus tomentosa[J]. Journal of Beijing Forestry University, 1988, 10(3): 97-101. http://www.cnki.com.cn/Article/CJFDTOTAL-BJLY198803014.htm
[5] 朱之悌, 林惠斌, 康向阳.毛白杨异源三倍体B301等无性系选育的研究[J].林业科学, 1995, 31(6): 499-505. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK199500347545 Zhu Z T, Lin H B, Kang X Y. Studies on allotriploid breeding of Populus tomentosa B301 clones[J]. Scientia Silvae Sinicae, 1995, 31(6): 499-505. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK199500347545
[6] 张志毅, 李凤兰.白杨染色体加倍技术研究及三倍体育种(Ⅰ):花粉染色体加倍技术[J].北京林业大学学报, 1992, 14(增刊3): 52-58. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=ae4e0519-53d0-46c4-902c-618e5b3bd30a Zhang Z Y, Li F L. Studies on chromosome doubling and triploid breeding of white poplar(Ⅰ): the techniques of the pollen chromosome doubling[J]. Journal of Beijing Forestry University, 1992, 14(Suppl.3): 52-58. http://bjly.chinajournal.net.cn/WKC/WebPublication/paperDigest.aspx?paperID=ae4e0519-53d0-46c4-902c-618e5b3bd30a
[7] 康向阳, 朱之悌, 林惠斌.杨树花粉染色体加倍有效处理时期的研究[J].林业科学, 1999, 35(4): 21-24. doi: 10.3321/j.issn:1001-7488.1999.04.004 Kang X Y, Zhu Z T, Lin H B. Study on the effective treating period for pollen chromosome doubling of Populus tomentosa×P. bolleana[J]. Scientia Silvae Sinicae, 1999, 35(4): 21-24. doi: 10.3321/j.issn:1001-7488.1999.04.004
[8] 康向阳, 朱之悌, 张志毅.高温诱导白杨2n花粉有效处理时期的研究[J].北京林业大学学报, 2000, 22(3): 1-4. doi: 10.3321/j.issn:1000-1522.2000.03.001 Kang X Y, Zhu Z T, Zhang Z Y. Suitable period of high temperature treatment for 2n pollen of Populus tomentosa×P. bolleana[J]. Journal of Beijing Forestry University, 2000, 22(3): 1-4. doi: 10.3321/j.issn:1000-1522.2000.03.001
[9] 李云, 朱之悌, 田砚亭, 等.极端温度处理白杨雌花芽培育三倍体植株的研究[J].北京林业大学学报, 2000, 22(5): 7-12. doi: 10.3321/j.issn:1000-1522.2000.05.002 Li Y, Zhu Z T, Tian Y T, et al. Obtaining triploids by high and low temperature treating female flower buds of white poplar[J]. Journal of Beijing Forestry University, 2000, 22(5): 7-12. doi: 10.3321/j.issn:1000-1522.2000.05.002
[10] 康向阳, 朱之梯, 张志毅.银腺杨与毛新杨正反交三倍体选育[J].北京林业大学学报, 2000, 22(6): 8-11. doi: 10.3321/j.issn:1000-1522.2000.06.003 Kang X Y, Zhu Z T, Zhang Z Y. Breeding of triploids by the reciprocal crossing of Populus alba × P. glandulosa and P. tomentosa×P. bolleana[J]. Journal of Beijing Forestry University, 2000, 22(6): 8-11. doi: 10.3321/j.issn:1000-1522.2000.06.003
[11] 康向阳, 张平冬, 高鹏, 等.秋水仙碱诱导白杨三倍体新途径的发现[J].北京林业大学学报, 2004, 26(1): 1-4. http://d.old.wanfangdata.com.cn/Periodical/bjlydxxb200401001 Kang X Y, Zhang P D, Gao P, et al. Discovery of a new way of poplar triploids induced with colchicine after pollination[J]. Journal of Beijing Forestry University, 2004, 26(1) : 1-4. http://d.old.wanfangdata.com.cn/Periodical/bjlydxxb200401001
[12] 李艳华, 康向阳.诱导大孢子染色体加倍选育白杨杂种三倍体[J].北京林业大学学报, 2007, 29(5): 22-25. doi: 10.3321/j.issn:1000-1522.2007.05.007 Li Y H, Kang X Y. Triploid induction in white poplar by chromosome doubling of megaspore[J]. Journal of Beijing Forestry University, 2007, 29(5): 22-25. doi: 10.3321/j.issn:1000-1522.2007.05.007
[13] 康宁, 白凤莹, 张平冬, 等.高温诱导胚囊染色体加倍获得毛白杨杂种三倍体[J].北京林业大学学报, 2015, 37(2): 79-86. doi: 10.13332/j.cnki.jbfu.2015.02.021 Kang N, Bai F Y, Zhang P D, et al. Inducing chromosome doubling of embryo sac in Populus tomentosa with high temperature exposure for hybrid triploids[J]. Journal of Beijing Forestry University, 2015, 37(2): 79-86. doi: 10.13332/j.cnki.jbfu.2015.02.021
[14] 康向阳, 王君.杨树多倍体诱导技术研究[M].北京:科学出版社, 2010. Kang X Y, Wang J. Poplar polyploid induction technology research[M]. Beijing: Science Press, 2010.
[15] 张妍, 崔彬彬, 魏俊杰, 等.毛白杨大、小孢子母细胞减数分裂进程的相关性[J].西北农林科技大学学报(自然科学版), 2015, 43(11): 63-70. http://d.old.wanfangdata.com.cn/Periodical/xbnydxxb201511010 Zhang Y, Cui B B, Wei J J, et al. Correlation between megesporecytes and microsporecytes meiosis stage of Populus tomentosa Carr.[J]. Journal of Northwest A & F University (Nat. Sci. Ed.), 2015, 43(11): 63-70. http://d.old.wanfangdata.com.cn/Periodical/xbnydxxb201511010
[16] Bai F Y, Kang N, Zhang P D, et al. Selection of female parents with high fertility and high combining abilities for cross-breeding Populus tomentosa[J/OL]. Journal of Forest Research, 2018[2018-07-05]. https://link.springer.com/content/pdf/10.1007%2Fs11676-018-0684-8.pdf.
[17] Galbraith D W, Harkins K R, Maddox J M, et al. Rapid flow cytometric analysis of the cell cycle in intact plant tissues[J]. Science, 1983, 220: 1049-1051. doi: 10.1126/science.220.4601.1049
[18] Wang J, Li D L, Kang X Y. Induction of unreduced megaspores with high temperature during megasporogenesis in Populus[J]. Annals of Forest Science, 2011, 69(1): 59-67. http://d.old.wanfangdata.com.cn/Conference/8447539
[19] Lu M, Zhang P D, Kang X Y. Induction of 2n female gametes in Populus adenopoda Maxim by high temperature exposure during female gametophyte development[J]. Breed Science, 2013, 63(1): 96-103. doi: 10.1270/jsbbs.63.96
[20] 康向阳.关于杨树多倍体育种的几点认识[J].北京林业大学学报, 2010, 32(5): 149-153. http://d.old.wanfangdata.com.cn/Periodical/bjlydxxb201005026 Kang X Y. Some understanding on polyploid breeding of poplars[J]. Journal of Beijing Forestry University, 2010, 32(5): 149-153. http://d.old.wanfangdata.com.cn/Periodical/bjlydxxb201005026
[21] Ramanna M S, Jacobsen E. Relevance of sexual polyploidization for crop improvement: a review[J]. Euphytica, 2003, 133(1): 3-8. doi: 10.1023/A:1025600824483
[22] 王君. 青杨派树种多倍体诱导技术研究[D]. 北京: 北京林业大学, 2009. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1489837 Wang J. Techniques of polyploid induction in Populus spp. (Section Tacamahaca)[D]. Beijing: Beijing Forestry University, 2009. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1489837
[23] Wahid A, Gelani S, Ashraf M, et al. Heat tolerance in plants: an overview[J]. Environmental & Experimental Botany, 2007, 61(3): 199-223. doi: 10.1016-j.envexpbot.2007.05.011/
[24] Einspahr D W. Production and utilization of triploid hybrid aspen[J]. Iowa State J Res, 1984, 58(4): 401-409.
[25] Weisgerber H, Rau H M, Gartner E J, et al. 25 years of forest tree breeding in Hesse[J]. Allge Forstze, 1980, 26: 665-712.
[26] Liao T, Cheng S, Zhu X, et al. Effects of triploid status on growth, photosynthesis, and leaf area in Populus[J]. Trees, 2016, 30(4):1137-1147. doi: 10.1007/s00468-016-1352-2




计量
- 文章访问数: 1463
- HTML全文浏览量: 348
- PDF下载量: 32
