
Population structure and spatial distribution point patterns of Tsuga chinensis var. tchekiangensis in Wuyishan Mountain, Jiangxi of eastern China
-
摘要:目的 了解武夷山自然保护区内南方铁杉的种群结构及空间分布情况,对揭示南方铁杉的演替更新具有重要意义。方法 对南方铁杉种群进行样地调查,通过种群年龄结构分析、静态生命表编制、生存分析、时间序列模型预测和点格局分析探索了南方铁杉种群结构和空间分布格局。结果 (1)南方铁杉种群年龄结构呈“金字塔”型,属增长型种群,存活曲线趋于Deevey-Ⅰ型,存活曲线、死亡率和消失率曲线以及生存函数分析表明种群具有前期稳定、中期减少和后期衰退的特点。(2)时间序列预测未来3、6和9个龄级时间后南方铁杉种群各龄级个体数均逐渐增加,南方铁杉幼年个体数相对丰富,老龄级树有较充足的后备资源得以补充,种群能自然更新。(3)南方铁杉种群整体在小尺度(r < 20 m)呈聚集分布,随尺度的增加呈随机分布或均匀分布;种群不同发育阶段的空间分布格局与研究尺度紧密相关,幼树明显呈聚集分布,中树随研究尺度增大由聚集分布向随机分布转变,老树基本呈随机分布。结论 南方铁杉种群天然更新能力较强,保护区维持现有的生境条件,南方铁杉种群将继续呈增长状态。Abstract:Objective Understanding the population structure and spatial distribution pattern of Tsuga chinensis var. tchekiangensis in Wuyishan Nature Reserve, Jiangxi of eastern China, is of great significance to reveal the succession and regeneration of Tsuga chinensis var. tchekiangensis.Method Based on plot investigation of Tsuga chinensis var. tchekiangensis population, we analyzed the population structure and spatial distribution point pattern of Tsuga chinensis var. tchekiangensis by studying many parameters, such as age structure, static life table, survival curves, time sequence model and point pattern analysis.Result (1) The population presented an increasing population with an age structure of ‘pyramid’ type, and the survival curves suggested that population of Tsuga chinensis var. tchekiangensis tended to be the type of Deevey-Ⅰ. The survival curve, mortality curve and vanish curve and survival function analysis of population showed that the characteristics of the population were stable in early age stage, decreased in middle age period, then declined in old age period. (2) Time sequence model predicted that the number of each age class individuals of the Tsuga chinensis var. tchekiangensis population would increase in the age classes of 3, 6 and 9 in the future. The number of Tsuga chinensis var. tchekiangensis was relatively abundant at a young stage, so that sufficient reserve resources can replenish the old trees, and the population can naturally renewed. (3) Point pattern analysis indicated that the individuals were significantly clustered at the small scale (r < 20 m), and the distribution became more random and regular at larger scales. The spatial distribution pattern of Tsuga chinensis var. tchekiangensis population at different developmental periods was closely related to the study scale. The spatial distribution point pattern of young individuals was of obviously aggregated distribution. The spatial distribution point pattern of middle individuals was aggregated to random as the study scale increased, and the old individuals showed randomly distribution.Conclusion The population can regenerate naturally and the population will tend to increase under the maintaining of the current habitat.
-
南方铁杉(Tsuga chinensis var. tchekiangensis)为松科(Pinaceae)铁杉属下一变种,是我国特有的珍稀裸子植物,广泛分布于浙江、安徽黄山、武夷山、江西武功山、湖南莽山、广东乳源、广西兴安及云南麻栗坡等地,生于海拔600 ~ 2 100 m处的针阔叶混交林中[1]。南方铁杉分布虽广,地跨中亚热带至北亚热带,但长期以来,其生境不断遭受破坏,导致南方铁杉生存竞争能力降低、种群数量不断减少,适应生存面积缩小、空间分布斑块化[2],且目前尚无人工造林,故南方铁杉已被列为国家三级重点保护植物[3]。南方铁杉属热带暖性针叶树种,喜温凉湿润的气候及深厚肥沃的酸性土壤,其材质坚实,纹理细致,耐阴湿,适于作建筑、家具等用材,不仅是珍贵的用材树种和良好的观赏树种,亦是南方山地优良的绿化树种。南方铁杉是第三纪孑遗植物,被称为植物“活化石”[4],具有很高的科研价值和潜在的经济价值[1]。
目前,国内外学者对南方铁杉已开展大量研究,包括群落结构及数量动态[2,5-6]、空间分布格局及更新状况[7-9]、种间关系[10-12]、生长规律[8,13],越冬策略[14]、金属含量与土壤养分的关系[15]、生态位[16]、菌根生态学[17-18]、林隙干扰规律[19]、栽培育苗[20]等方面,为南方铁杉的保护管理与发展利用提供参考。然而有关于南方铁杉的更新能力和空间分布格局的研究存在较大争议,冯祥麟等[5]、丁巧玲等[6]、祁红艳等[8]认为贵阳高坡、湖南桃源、江西武夷山南方铁杉自我更新能力存在一定挑战,种群更新比较困难;而袁荣斌等[21]、杨清培等[2]认为江西武夷山南方铁杉结构虽存在一定波动,但能进行自我更新。另外,王大来[22]利用方差/均值比率法分析种群的分布格局,认为南方铁杉呈聚集分布;何建源等[9]利用几项聚集度指标测定南方铁杉的空间分布格局,认为南方铁杉呈随机分布。杨培清等[7]用点格局分析发现,南方铁杉幼苗分布随研究尺度增长由聚集分布向随机分布演变。
江西武夷山自然保护区内保存有全国少见的较大区域的南方铁杉天然林,多分布于海拔1 200 ~ 2 000 m的阴坡或谷地,其分布区原生性植被较多。本文旨在通过对保护区南方铁杉分布核心区内该物种的更新情况及分布格局展开探索,利用时间序列模型预测未来南方铁杉种群的变化趋势,采用点格局分析从不同尺度揭示其空间分布类型,以期揭示该地区南方铁杉种群结构和自我更新能力,阐明其空间分布规律以及影响其分布的因子,并且提出些高效实际的管理措施来促进南方铁杉种群的发展,为加强南方铁杉的保护和科学管理提供理论依据。
1. 研究区概况
江西武夷山自然保护区位于江西省铅山县,地处武夷山脉北段的西北坡,地理范围为117°39′30″ ~ 117°55′47″E、27°48′11″ ~ 28°00′35″N,属于中亚热带常绿阔叶林带。该地具有亚热带东部季风气候特征,年均气温为14.2 ℃,年均降水量2 583 mm,最高达3 544 mm,年平均相对湿度84%。保护区总面积16 007 hm2,南方铁杉天然分布区1 560 hm2,自然环境独特,地质构造复杂,地貌类型多变,土壤多为山地棕壤,植被茂盛且垂直分布明显。南方铁杉常与阔叶树组成混交林,其伴生树种有多脉青冈(Cyclobalanopsis multinervis)、闽皖八角(Illicium minwanense)、鹿角杜鹃(Rhododendron latoucheae)、白檀(Symplocos paniculata)、灯笼树(Enkianthus chinensis)、岩柃(Eurya saxicola)等,是中山温性针叶林的典型代表[21]。
2. 研究方法
2.1 样地设置及实地调查
在前期查阅文献及实地踏查基础上,在保护区黄冈山脉北坡的南方铁杉分布核心区内设置面积为160 m × 400 m的研究样地1个(海拔1 700 ~ 1 800 m,西北坡向,坡度30° ~ 70°),并将其划分为640个10 m × 10 m小样方,并依次编号。以小样方为调查单元,对样地内的所有胸径(DBH) ≥ 1.0 cm的乔木进行每木检尺,记录样方编号、植物中文名及其主干编号、胸径(cm)、树高(m)、枝下高(m)、冠幅x(m)、冠幅y(m)、坐标(x, y)等指标,并记录整个样方的生境条件及植被状况。每木检尺时,以基株为单位进行统计,若一个树桩上生长出多个无性系分株,则记录龄级最大的一株。同时,以小样方的左下方为坐标原点(0, 0),测量的乔木个体空间位置,坐标值用样方的投影距离(m)表示。
2.2 数据处理
2.2.1 种群龄级结构
树木年轮是木本植物年龄的直接标志,南方铁杉作为国家三级重点保护植物,难以获得树干解析木测定每一个体的确切年龄,本文采用空间代替时间的方法,根据野外调查数据特征并结合南方铁杉的生活史特点可知[23-24],南方铁杉树形高大,主干通直,可将其胸径(DBH)视作与龄级一一对应,以此分析种群的年龄结构。将南方铁杉胸径大小划分为10个龄级,胸径DBH ≤ 10 cm为第1龄级,10 cm < DBH ≤ 20 cm为第2龄级,20 cm < DBH ≤ 30 cm为第3龄级,以此类推,将南方铁杉种群共划分为10个龄级,统计各龄级个体数。并以龄级为横坐标,各龄级株数为纵坐标,绘制样地南方铁杉种群年龄结构图。把树木从小到大的龄级大小看作是时间先后顺序,将南方铁杉种群1 ~ 3级视为幼龄级,4 ~ 7级为中龄级,8 ~ 10级为老龄级。
2.2.2 静态生命表及存活曲线
静态生命表的编制及各参数具体算法参考相关文献[6,25],利用树木胸径代表龄级,制作南方铁杉静态生命表,并以龄级为横坐标,标准化存活数、死亡率和消失率为纵坐标,分别绘制南方铁杉种群的存活曲线、死亡率曲线和消失率曲线图。
2.2.3 种群生存分析
为揭示南方铁杉种群的动态变化规律,引入生存分析中生存率函数S(i)、累计死亡率函数F(i)、死亡密度函数f (ti)和危险率函数λ(ti),计算方法参考文献[26]。并根据上述4个生存函数估值绘制生存曲线、累计死亡率曲线、死亡密度曲线和危险率曲线。
2.2.4 种群数量动态的时间序列预测模型
本文采用时间序列分析的移动平均法[27-28]对南方铁杉种群的年龄结构进行预测,其模型如下:
M(1)t=1nt∑k=t−n+1Xk 式中:n表示需要预测的未来时间年限;t表示龄级,Xk表示k龄级内的个体数,
Mt 表示经过n时间后未来t龄级的种群大小。本文对未来经历3、6、9个龄级时间后的种群各龄级的个体数量进行预测。2.2.5 点格局分析
基于南方铁杉个体分布空间坐标信息,运用点格局分析中单变量O-ring统计方法,分析南方铁杉种群个体在多尺度上的空间分布格局。本文计算的空间尺度为0 ~ 80 m,选择步长(ring width)为1 m,空间分辨率为1 m,通过99次Monte Carlo随机模拟计算95%的置信区间,得出南方铁杉种群的空间分布情况,具体分析过程利用Programita软件[29]完成。
通过O11(r)分析单一种群的格局特征,分析过程中,单变量O-ring统计函数采用Monte Carlo检验拟合包迹线,结果用空间尺度r作为横坐标,O-ring函数值为纵坐标绘图。用南方铁杉种群实际分布数据计算得到的不同尺度下的O11(r)值,在某一尺度处,若函数值在包迹线以内,则符合随机分布;若在包迹线以上,则属于聚集分布;若在包迹线以下,则属于均匀分布[30]。
3. 结果与分析
3.1 年龄结构特征
南方铁杉各龄级个体数量分布如图1,由图可知,幼龄级个体比例较大,老龄级比例较小,种群年龄结构呈“金字塔”型,属增长型种群。南方铁杉种群个体数存在一定波动性,幼龄级个体(1 ~ 3)占总数的47.8%,中龄级(4 ~ 7)个体占总数的46.5%,二者共占总个体数的94.5%,说明幼龄树和中龄树的数量较为丰富,而老龄级个体比例仅为5.5%,说明南方铁杉种群偏向于增长型种群,能自然更新。
![]() 图 1 南方铁杉种群年龄结构Figure 1. Age structure of Tsuga chinensis var. tchekiangensis population
图 1 南方铁杉种群年龄结构Figure 1. Age structure of Tsuga chinensis var. tchekiangensis population3.2 南方铁杉种群静态生命表及存活曲线
南方铁杉种群静态生命表如表1所示。以龄级为x轴,以lnlx、qx、Kx为y轴绘制各年龄组的存活曲线、死亡率(qx)和消失率(Kx)曲线图(图2)。由表1可知,南方铁杉种群幼龄的生命期望(ex)最大,说明幼龄个体的生存能力较好,但随着龄级的增长而呈现不断下降的趋势,老龄种群生命期望(ex)最低,表明南方铁杉的生存能力随着年龄的增加而下降。
表 1 南方铁杉种群静态生命表Table 1. Static life table of Tsuga chinensis var. tchekiangensis population龄级
Age classDBH/cm 组中值
Group median/cmax lx lnlx dx qx Lx Tx ex Kx Sx 1 0 ~ 10 5 146 1 000 6.908 14 0.014 0 993 5 629 5.629 0.014 0.986 2 10 ~ 20 15 144 986 6.894 41 0.041 6 965.5 4 636 4.702 0.043 0.958 3 20 ~ 30 25 138 945 6.851 41 0.043 4 924.5 3 670.5 3.884 0.045 0.957 4 30 ~ 40 35 132 904 6.806 41 0.045 4 883.5 2 746 3.038 0.046 0.955 5 40 ~ 50 45 126 863 6.760 233 0.270 0 746.5 1 862.5 2.158 0.314 0.730 6 50 ~ 60 55 92 630 6.446 164 0.260 3 548 1 116 1.771 0.322 0.740 7 60 ~ 70 65 68 466 6.144 261 0.560 1 335.5 568 1.219 0.821 0.440 8 70 ~ 80 75 30 205 5.323 123 0.600 0 143.5 232.5 1.134 0.916 0.400 9 80 ~ 90 85 12 82 4.407 34 0.414 6 65 89 1.085 0.536 0.585 10 90 ~ 100 95 7 48 3.871 − − 24 24 0.5 − − 注:ax存活数;lx存活量;dx死亡数;qx死亡率;Lx区间寿命;Tx总寿命;ex期望寿命;Kx消失率;Sx存活率。Notes: ax means survival; lx means survival quantity; dx means death number; qx means mortality rate; Lx means span life; Tx means total life; ex means life expectancy; Kx means vanish rate; Sx means survival rate. ![]() 图 2 南方铁杉种群存活曲线、死亡率(qx)、消失率(Kx)曲线Figure 2. Survival curve, mortality rate (qx) and vanish rate (Kx) curves of Tsuga chinensis var. tchekiangensis population
图 2 南方铁杉种群存活曲线、死亡率(qx)、消失率(Kx)曲线Figure 2. Survival curve, mortality rate (qx) and vanish rate (Kx) curves of Tsuga chinensis var. tchekiangensis population由图2可见,南方铁杉种群存活曲线为凸曲线,表现为Deevey-Ⅰ型。幼龄级的南方铁杉种群存活率较高且变化趋势幅度较小,随着龄级增长,存活率明显呈现波动性下降,第10龄级的存活率达到最低,说明幼龄级南方铁杉生长稳定,随龄级的增加,种群存活率明显减少,而死亡率及消失率逐渐升高。南方铁杉种群死亡率曲线qx和消失率曲线Kx变化大致相似,随龄级增加逐渐增大,且在第8龄级达到峰值,最大死亡率为0.6,最大消失率为0.916,这可能与南方铁杉的生理衰老有关。
3.3 生存分析
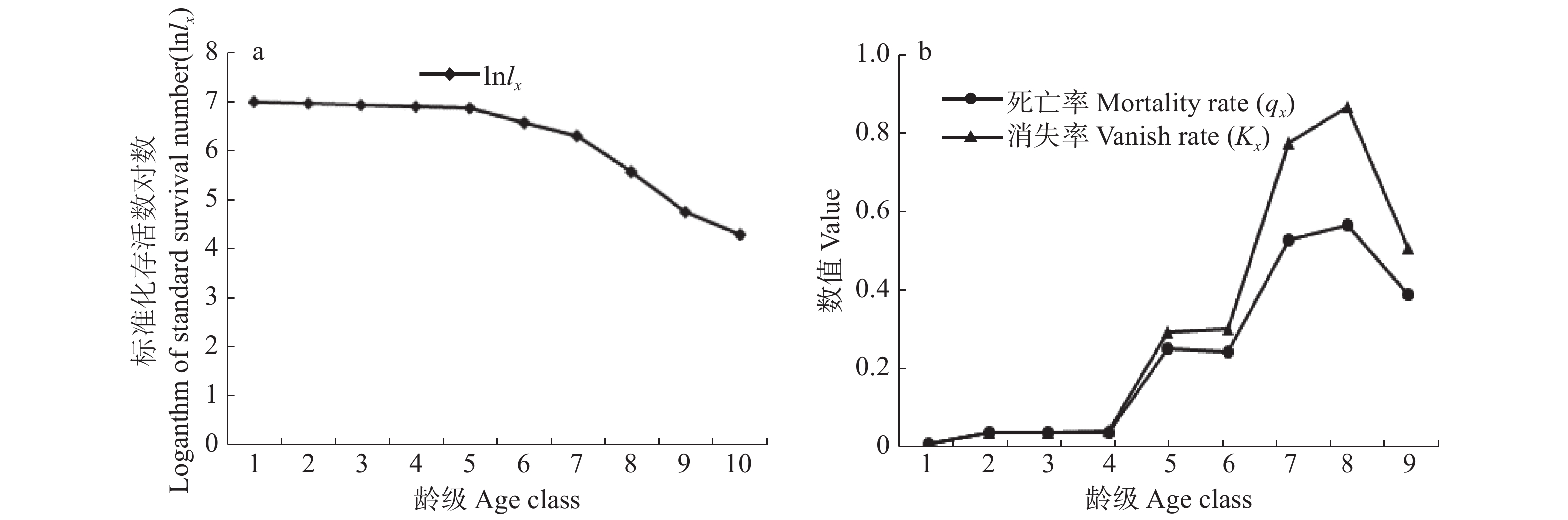
南方铁杉种群生存分析结果(图3)显示,南方铁杉种群的生存率函数曲线随着龄级的增加下降,累积死亡率函数曲线则与之相反。1 ~ 3龄级内,南方铁杉种群生存率较高,死亡率较低,说明幼树及小树生长情况良好;生存率曲线在3 ~ 8龄级内锐减而累积死亡率函数骤增,在第8龄级之后二者的变化幅度趋于平缓。南方铁杉种群的死亡密度曲线和危险率曲线变化趋势前期基本相似且较低,第3龄级后存在一定波动,死亡密度曲线在第6龄级出现峰值,且危险率较高,说明南方铁杉生长到中龄级阶段,个体对空间和自然条件需求不断加大,导致种群内资源竞争加剧,种群遭受强烈的环境筛。危险率曲线在第9龄级达到峰值,但死亡密度很小,说明南方铁杉种群在老龄级时逐渐进入生理衰退状态。4个生存函数变化曲线表明,南方铁杉种群具有前期稳定、中期减少、后期衰退的特点,这与种群存活曲线分析的结果一致。
![]() 图 3 南方铁杉种群生存分析曲线Figure 3. Survival analysis curves of Tsuga chinensis var. tchekiangensis population
图 3 南方铁杉种群生存分析曲线Figure 3. Survival analysis curves of Tsuga chinensis var. tchekiangensis population3.4 时间序列预测
基于南方铁杉种群各龄级原始数据,利用时间序列模型按照一次移动平均法对未来3、6、9个龄级时间后南方铁杉个体数进行预测(表2)。由表2可知未来3、6和9个龄级时间后南方铁杉各龄级的种群个体数量呈现不同程度增加的趋势。经过未来3、6个龄级时间后,第6龄级个体数分别增加了27.2%、41.3%,且随着时间的推移,龄级越大,其内的种群个体数量增加的幅度越大。经过未来3、6、9个龄级时间后,第9龄级个体数分别增加了192%、541.6%、725%,这表明南方铁杉种群生长中后期阶段的个体数量得到一定补充。表明该种群仍处在增长状态。综合整个种群的发展趋势,虽然老龄级个体数明显增多,但其幼龄级个体储备丰富,随时间不断生长有充足的中龄级个体补充以维持后期龄级的更新,总体来说南方铁杉种群在未来一段演替时间内呈增长趋势,仍是优势种群。
表 2 南方铁杉种群龄级结构的时间预测Table 2. Time sequence prediction in the quantitative dynamics of Tsuga chinensis var. tchekiangensis population龄级
Age class原始数据
Primary dataM3(1) M6(1) M9(1) 1 146 − − − 2 144 − − − 3 138 143 − − 4 132 138 − − 5 126 132 − − 6 92 117 130 − 7 68 95 117 − 8 30 63 98 − 9 12 35 77 99 10 7 16 56 83 3.5 南方铁杉种群空间分布格局
南方铁杉种群不同生长阶段的点格局分析结果如图4,南方铁杉幼树在0 ~ 80 m空间尺度上的O11(r)值大多高于置信区间,基本呈聚集分布;中树在0 ~ 68 m的空间尺度上的O11(r)值高于置信区间,呈聚集分布,随着尺度增加,在69 ~ 80 m的空间尺度O11(r)值基本位于置信区间内,呈随机分布;老树在O11(r)值基本位于置信区间内,呈随机分布。南方铁杉种群整体空间分布表明,个体在0 ~ 20 m空间尺度上呈显著的聚集分布,在24 ~ 26 m、58 ~ 62 m空间尺度上,空间分布表现为随机分布。在28 ~ 57 m、63 ~ 69 m、76 ~ 80 m空间尺度上O11(r)值低于置信区间,南方铁杉种群呈现明显的均匀分布。南方铁杉种群所有个体在空间尺度较小时呈明显的聚集分布,随着空间尺度范围变大,呈随机分布或均匀分布。
![]() 图 4 南方铁杉种群空间分布Figure 4. Point pattern analysis of Tsuga chinensis var. tchekiangensis population
图 4 南方铁杉种群空间分布Figure 4. Point pattern analysis of Tsuga chinensis var. tchekiangensis population4. 讨 论
4.1 南方铁杉种群结构及更新情况
南方铁杉种群结构受其生物学特征与环境因素综合影响,不同学者对南方铁杉种群结构及更新能力的研究结果多有差异。祁红艳等[8]对江西武夷山南方铁杉成熟林中1 800 m2的样方进行调查,结果发现南方铁杉种群结构呈“倒金字塔”型,林下幼苗幼树较少,其种群数量动态呈衰退趋势,自然更新困难。但杨培清等[7]对武夷山典型南方铁杉分布核心区中3 000 m2的样地进行调查,结果显示南方铁杉种群呈“金字塔”型,属增长型种群,南方铁杉完全能自然更新。这表明南方铁杉种群结构与其生境条件密切相关,前者所选样地是人为干扰较少的成熟林,调查面积较小,后者研究地包括部分塌方干扰形成的林窗及部分成熟林,均难以客观精准地反映南方铁杉种群结构及更新状况。
本研究选择的样地是南方铁杉核心分布区内具有代表性的天然林,调查区域范围较大,地形多变、地貌复杂,涵盖不同生境条件,且人为干扰较少,群落原生性植被较多。结果表明,样地内南方铁杉种群年龄结构呈“金字塔”型,存活曲线趋于Deevey-Ⅰ型,说明南方铁杉属增长型种群,有能力进行自然更新。死亡率曲线、消失率曲线和生存分析曲线分析表明,南方铁杉种群具有前期稳定,中期波动幅度较大,后期衰退的特点,这是因为南方铁杉幼苗能耐一定的蔽荫,且气候温暖湿润,故幼龄树存活率较高,长势良好。随龄级的增长,南方铁杉死亡率逐渐升高,这是因为南方铁杉与常绿阔叶树(鹿角杜鹃、多脉青冈、闽皖八角等)组成了多层混交林,南方铁杉性喜阳光但生长缓慢,处于生长盛期的个体对光照、养分等资源的需求量加大,而群落的郁闭度和种间竞争便成为限制其生长的主要环境因素,并随着南方铁杉的生长更新,环境筛强度增大,导致竞争力弱的中龄级个体逐渐衰亡。老龄级南方铁杉多处于林冠层,冠幅大且根系深,对土壤、空间等生境资源利用率较高,但大多已进入生理衰老阶段,个体竞争能力下降便逐渐走向衰亡,故种群后期死亡率较高。时间序列预测分析表明,由于幼龄个体的储备,南方铁杉种群经过3、6和9个龄级时间后仍呈小幅度增长状态,说明该地生境条件优良且营养资源丰富,南方铁杉更新能力较强,与年龄结构分析结果一致。
4.2 南方铁杉种群空间分布格局
植物种群分布格局是物种自身生物学特性、种内种间关系及生境条件共同作用的结果,反映出种群个体在水平空间上彼此的关系及其在群落中的作用和地位[31-32]。种群空间分布格局的研究方法较多,点格局分析法在1977年由Ripley提出[33],其优越性在于能分析任意尺度下的空间分布格局,目前已广泛应用于植物种群分布格局的研究中。不同学者研究结果中,南方铁杉种群在不同生长阶段、不同研究尺度下呈现不同的分布状态[7,9,22],为避免研究方法和研究尺度对分析结果的影响,本研究基于160 m × 400 m的大样地,结合较先进的点格局法,客观地反映江西武夷山南方铁杉种群的空间分布格局。结果表明,南方铁杉种群整体上在小尺度范围内(r < 20 m)呈明显的聚集分布,随尺度增加表现为随机分布和均匀分布,这符合自然群落种群分布的一般规律 [34-35]。不同发育阶段的南方铁杉种群呈现不同类型的分布格局,这与南方铁杉生物学特性(种子扩散机制、繁育特征、种内种间关系等)及生境条件(地貌、土壤、水分、光照等)密切相关:南方铁杉性喜阳光,花粉无气囊,种子散布距离较近[21],幼苗通常出现在母树个体周围,故种群幼树多呈聚集分布,这有利于幼树发挥群体效应,增强适应环境能力,促进个体生长。随着种群龄级的增大,种群空间分布在大尺度上呈随机分布,这是由于林窗大小及幼树的密度机制影响其存活,南方铁杉种群个体生长至阔叶混交林中的更新层,盛长的个体对土壤养分、水分、光照等营养资源的需求增大,种内、种间竞争增强,经过自疏、他疏的作用,导致种群聚集度降低,逐渐呈随机分布,也是南方铁杉种群趋于稳定的表现。老龄级的南方铁杉个体多为高大植株,根系发达且处于林冠层,经过复杂的环境筛,存活的植株可获得充分的光照及土壤养分,空间竞争趋于缓和,故种群格局呈随机分布。
5. 结 论
南方铁杉作为我国特有珍稀裸子植物,亦是国家三级重点保护植物,研究其种群结构变化及分布格局,可揭示其种群演替趋势及分布成因,为珍稀濒危植物保护提供参考。本研究表明,南方铁杉种群属增长型种群,能进行自然更新,种群结构具有前期稳定、中期减少和后期衰退的特点;时间序列预测未来3、6和9个龄级时间后南方铁杉种群各龄级个体数均逐渐增加,说明样地生境条件优良,适合南方铁杉生长更新。南方铁杉种群空间分布格局与研究尺度紧密相关,在小尺度上呈聚集分布,随尺度的增加呈随机分布或均匀分布;种群不同龄级个体的空间分布格局,幼树明显呈聚集分布,中树随研究尺度增大由聚集分布向随机分布转变,老树基本呈随机分布。以上结果表明,保护区维持现有的生境条件,继续封山育林,并根据南方铁杉的生长特性,适当的进行人为干扰。间伐以降低群落郁闭度,减少种内竞争;及时清理枯病、倒伐木;加强对林窗幼苗的抚育措施,对促进南方铁杉种群生长更新及保护管理具有重要意义。
-
![]()
图 1 南方铁杉种群年龄结构
Figure 1. Age structure of Tsuga chinensis var. tchekiangensis population
![]()
图 2 南方铁杉种群存活曲线、死亡率(qx)、消失率(Kx)曲线
Figure 2. Survival curve, mortality rate (qx) and vanish rate (Kx) curves of Tsuga chinensis var. tchekiangensis population
![]()
图 3 南方铁杉种群生存分析曲线
Figure 3. Survival analysis curves of Tsuga chinensis var. tchekiangensis population
![]()
图 4 南方铁杉种群空间分布
Figure 4. Point pattern analysis of Tsuga chinensis var. tchekiangensis population
表 1 南方铁杉种群静态生命表
Table 1 Static life table of Tsuga chinensis var. tchekiangensis population
龄级
Age classDBH/cm 组中值
Group median/cmax lx lnlx dx qx Lx Tx ex Kx Sx 1 0 ~ 10 5 146 1 000 6.908 14 0.014 0 993 5 629 5.629 0.014 0.986 2 10 ~ 20 15 144 986 6.894 41 0.041 6 965.5 4 636 4.702 0.043 0.958 3 20 ~ 30 25 138 945 6.851 41 0.043 4 924.5 3 670.5 3.884 0.045 0.957 4 30 ~ 40 35 132 904 6.806 41 0.045 4 883.5 2 746 3.038 0.046 0.955 5 40 ~ 50 45 126 863 6.760 233 0.270 0 746.5 1 862.5 2.158 0.314 0.730 6 50 ~ 60 55 92 630 6.446 164 0.260 3 548 1 116 1.771 0.322 0.740 7 60 ~ 70 65 68 466 6.144 261 0.560 1 335.5 568 1.219 0.821 0.440 8 70 ~ 80 75 30 205 5.323 123 0.600 0 143.5 232.5 1.134 0.916 0.400 9 80 ~ 90 85 12 82 4.407 34 0.414 6 65 89 1.085 0.536 0.585 10 90 ~ 100 95 7 48 3.871 − − 24 24 0.5 − − 注:ax存活数;lx存活量;dx死亡数;qx死亡率;Lx区间寿命;Tx总寿命;ex期望寿命;Kx消失率;Sx存活率。Notes: ax means survival; lx means survival quantity; dx means death number; qx means mortality rate; Lx means span life; Tx means total life; ex means life expectancy; Kx means vanish rate; Sx means survival rate.  下载: 导出CSV
下载: 导出CSV
表 2 南方铁杉种群龄级结构的时间预测
Table 2 Time sequence prediction in the quantitative dynamics of Tsuga chinensis var. tchekiangensis population
龄级
Age class原始数据
Primary dataM3(1) M6(1) M9(1) 1 146 − − − 2 144 − − − 3 138 143 − − 4 132 138 − − 5 126 132 − − 6 92 117 130 − 7 68 95 117 − 8 30 63 98 − 9 12 35 77 99 10 7 16 56 83
下载: 导出CSV
-
[1] 张志祥. 珍稀濒危植物南方铁杉研究进展[J]. 生物学教学, 2011, 36(6):3−5. doi: 10.3969/j.issn.1004-7549.2011.06.002 Zhang Z X. Research advance of Tsuga chinensis var. tchekiangensis[J]. Biological Teaching, 2011, 36(6): 3−5. doi: 10.3969/j.issn.1004-7549.2011.06.002
[2] 杨清培, 钟安建, 金志农, 等. 江西武夷山南方铁杉群落分类及更新能力评价[J]. 江西农业大学学报, 2014, 36(6):1275−1283. Yang Q P, Zhong A J, Jin Z N, et al. A community classification of natural forest of Tsuga chinensis var. tchekiangensis and its regeneration capacity[J]. Acta Agriculturae Universitatis Jiangxiensis, 2014, 36(6): 1275−1283.
[3] 陈林, 龚粤宁, 谢国光, 等. 广东南岭国家级自然保护区珍稀濒危植物及其保护[J]. 植物科学学报, 2012, 30(3):277−284. Chen L, Gong Y N, Xie G G, et al. Rare and endangered plants and conservation in Guangdong Nanling National Nature Reserve[J]. Plant Science Journal, 2012, 30(3): 277−284.
[4] 李林, 魏识广, 黄忠良, 等. 猫儿山两种孑遗植物的更新状况和空间分布格局分析[J]. 植物生态学报, 2012, 36(2):144−150. Li L, Wei S G, Huang Z L, et al. Regenerative condition and analysis of spatial distribution pattern of two relic plants in Maoershan Mountain, China[J]. Chinese Journal of Plant Ecology, 2012, 36(2): 144−150.
[5] 冯祥麟, 胡刚, 刘正华. 贵阳高坡南方铁杉群落特征及种群结构调查研究[J]. 贵州林业科技, 2011, 39(2):26−29. Feng X L, Hu G, Liu Z H. Research on the community characteristics and population dynamics of Tsuga chinensis var. tchekiangensis on Gaopo in Guiyang City[J]. Guizhou Forestry Science and Technology, 2011, 39(2): 26−29.
[6] 丁巧玲, 刘忠成, 王蕾, 等. 湖南桃源洞国家级自然保护区南方铁杉种群结构与生存分析[J]. 西北植物学报, 2016, 36(6):1233−1244. Ding Q L, Liu Z C, Wang L, et al. Structure and survival analysis of Tsuga chinensis var. tchekiangensis populations in Taoyuandong National Nature Reserve, Hunan Province[J]. Acta Botanica Boreali-Occidentalia Sinica, 2016, 36(6): 1233−1244.
[7] 杨清培, 金志农, 裘利洪, 等. 江西武夷山南方铁杉更新格局及代际关联性分析[J]. 生态学杂志, 2014, 33(4):939−945. Yang Q P, Jin Z N, Qiu L H, et al. Regeneration pattern and intergeneration association of Tsuga chinensis var. tchekiangensis in Jiangxi Wuyishan National Nature Reserve[J]. Chinese Journal of Ecology, 2014, 33(4): 939−945.
[8] 祁红艳, 金志农, 杨清培, 等. 江西武夷山南方铁杉生长规律及更新困难的原因解释[J]. 江西农业大学学报, 2014, 36(1):137−143. doi: 10.3969/j.issn.1000-2286.2014.01.021 Qi H Y, Jin Z N, Yang Q P, et al. Growing law and cause of poor regeneration of Tsuga chinensis var. tchekiangensis in Jiangxi Wuyishan National Nature Reserve[J]. Acta Agriculturae Universitatis Jiangxiensis, 2014, 36(1): 137−143. doi: 10.3969/j.issn.1000-2286.2014.01.021
[9] 何建源, 卞羽, 吴焰玉, 等. 不同坡向濒危植物南方铁杉的分布格局[J]. 中国农学通报, 2010, 26(13):122−125. He J Y, Bian Y, Wu Y Y, et al. Spatial distribution pattern of the endangered plant Tsuga chinensis var. tchekiangensis in different aspect[J]. Chinese Agricultural Science Bulletin, 2010, 26(13): 122−125.
[10] 罗金旺. 福建光泽天然林中南方铁杉的种内与种间竞争[J]. 林业科技开发, 2011, 25(4):71−74. doi: 10.3969/j.issn.1000-8101.2011.04.018 Luo J W. Intra and interspecific competition in Tsuga chinensis var. tchekiangensis in a natural forest in Guangze, Fujian, China[J]. China Forestry Science & Technology, 2011, 25(4): 71−74. doi: 10.3969/j.issn.1000-8101.2011.04.018
[11] 赵峰. 莽山南方铁杉群落种间关系研究[J]. 中国农学通报, 2011, 27(31):68−72. Zhao F. The relationship of species in Tsuga chinensis var. tchekiangensis community in Mangshan Mountain Natural Reserve[J]. Chinese Agricultural Science Bulletin, 2011, 27(31): 68−72.
[12] 何建源, 荣海, 吴焰玉, 等. 武夷山南方铁杉群落乔木层种间联结研究[J]. 福建林学院学报, 2010, 30(2):169−173. doi: 10.3969/j.issn.1001-389X.2010.02.018 He J Y, Rong H, Wu Y Y, et al. Study on interspecific association of main populations in arborous layer of a Tsuga chinensis var. tchekiangensis community in Wuyishan National Nature Reserve of Fujian Province[J]. Journal of Fujian College of Forestry, 2010, 30(2): 169−173. doi: 10.3969/j.issn.1001-389X.2010.02.018
[13] 罗金旺. 福建光泽南方铁杉天然林的生长规律与生物量[J]. 福建林学院学报, 2011, 31(2):156−160. doi: 10.3969/j.issn.1001-389X.2011.02.013 Luo J W. Growth and biomass of Tsuga chinensis var. tchekiangensis in a natural forest in Guangze, Fujian, China[J]. Journal of Fujian College of Forestry, 2011, 31(2): 156−160. doi: 10.3969/j.issn.1001-389X.2011.02.013
[14] 张强, 郭传友, 张兴旺, 等. 基于光合作用和抗氧化机制的南方铁杉和褐叶青冈越冬策略研究[J]. 植物研究, 2015, 35(2):200−207. Zhang Q, Guo C Y, Zhang X W, et al. Photosynthesis and antioxidant defense strategies in overwintering plants of Tsuga chinensis var. tchekiangensis and Cyclobalanopsis stewardiana[J]. Bulletin of Botanical Research, 2015, 35(2): 200−207.
[15] 张志祥, 刘鹏, 徐根娣, 等. 不同群落类型下南方铁杉金属元素含量差异及其与土壤养分因子的关系[J]. 植物生态学报, 2010, 34(5):505−516. doi: 10.3773/j.issn.1005-264x.2010.05.004 Zhang Z X, Liu P, Xu G D, et al. Metal element contents of Tsuga chinensis var. tchekiangensis in different community types and its relationship with soil nutrient factors in eastern China[J]. Chinese Journal of plant Ecology, 2010, 34(5): 505−516. doi: 10.3773/j.issn.1005-264x.2010.05.004
[16] 谢琼中. 南方铁杉群落物种多样性及乔木优势种生态位初步研究[J]. 天津农业科学, 2011, 17(2):133−136. doi: 10.3969/j.issn.1006-6500.2011.02.039 Xie Q Z. Primary study on species diversity and niche characteristics of dominant tree species in Tsuga chinensis var. tchekiangensis community[J]. Tianjin Agricultural Science, 2011, 17(2): 133−136. doi: 10.3969/j.issn.1006-6500.2011.02.039
[17] 吴九玲, 钱晓鸣, 刘燕. 南方铁杉外生菌根的扫描电镜观察[J]. 厦门大学学报(自然版), 2001, 40(6):1337−1341. Wu J L, Qian X M, Liu Y. SEM-observation of ectomycorrhizal outer surface of Tsuga chinensis var. tchekiangensis (Flous) Cheng[J]. Journal of Xiamen University (Natural Science Edition), 2001, 40(6): 1337−1341.
[18] 钱晓鸣, 黄耀坚, 张艳辉, 等. 武夷山自然保护区南方铁杉外生菌根生物多样性[J]. 福建农林大学学报(自然版), 2007, 36(2):180−185. Qian X M, Huang Y J, Zhang Y H, et al. Biodiversity of the ectomycorrhizal on the rare and endangered tree species Tsuga chinensis var. tchekiangensis (Flous) Cheng in Wuyishan Nature ReServe[J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2007, 36(2): 180−185.
[19] 何建源, 卞羽, 吴焰玉, 等. 南方铁杉林林隙自然干扰规律[J]. 西南林业大学学报, 2009, 29(6):7−10. doi: 10.3969/j.issn.2095-1914.2009.06.002 He J Y, Bian Y, Wu Y Y, et al. Study on the law of natural disturbance in the gaps of Tsuga chinensis var. tchekiangensis fores[J]. Journal of Southwest Forestry University, 2009, 29(6): 7−10. doi: 10.3969/j.issn.2095-1914.2009.06.002
[20] 李晓铁, 玉伟朝, 罗远周, 等. 南方铁杉扦插繁殖技术[J]. 林业实用技术, 2008(6):21−22. Li X T, Yu W C, Luo Y Z, et al. Cutting propagation techniques of Tsuga chinensis var. tchekiangensis[J]. Practical Forestry Technology, 2008(6): 21−22.
[21] 袁荣斌, 邹思成, 兰文军, 等. 江西武夷山国家级自然保护区南方铁杉资源调查初报[J]. 南方林业科学, 2012(4):37−39. Yuan R B, Zou S C, Lan W J, et al. Report of resources investigation of Tsuga chinensis var. tchekiangensis in Jiangxi Wuyishan National Nature Reserve[J]. South China Forestry Science, 2012(4): 37−39.
[22] 王大来. 莽山南方铁杉种群格局分布格局研究[J]. 中国农学通报, 2010, 26(1):74−77. Wang D L. Distribution pattern of Tsuga chinensis var. tchekiangensis populations in Mang Mountain[J]. Chinses Agricultural Science Bulletin, 2010, 26(1): 74−77.
[23] 郑万钧. 中国树种分类分布的研究[J]. 林业科学, 1981, 17(4):453−455. Zheng W J. Note on the scientific names and geographical distribution of some Chinese trees[J]. Scientia Silvae Sinicae, 1981, 17(4): 453−455.
[24] 陈易之. 武夷山自然保护区动、植物资源[J]. 资源科学, 1994, 16(3):65−67. Chen Y Z. Plant and animal resources in natural reserve of Wuyishan Mountain[J]. Resources Science, 1994, 16(3): 65−67.
[25] 袁在翔, 金雪梅, 马婷瑶, 等. 南京灵谷寺栓皮栎种群结构与动态[J]. 生态学杂志, 2017, 36(6):1488−1494. Yuan Z X, Jin X M, Ma T Y, et al. The population structure and dynamics of Quercus variadilis in Nanjing Spirit Valley[J]. Chinese Journal of Ecology, 2017, 36(6): 1488−1494.
[26] 杨凤翔, 王顺庆, 徐海根, 等. 生存分析理论及其在研究生命表中的应用[J]. 生态学报, 1991, 11(2):153−158. doi: 10.3321/j.issn:1001-9332.1991.02.013 Yang F X, Wang S Q, Xu H G, et al. The theory of survival analysis and its application to life table[J]. Acta Ecologica Sinica, 1991, 11(2): 153−158. doi: 10.3321/j.issn:1001-9332.1991.02.013
[27] 李妍, 李登武, 韩东辰. 贺兰山东坡青海云杉种群结构[J]. 浙江农林大学学报, 2014, 31(1):50−56. Li Y, Li D W, Han D C. Population dynamics of Picea crassifolia on the eastern slope of Helan Mountain[J]. Journal of Zhejiang A & F University, 2014, 31(1): 50−56.
[28] 刘丹, 刘士玲, 郭忠玲, 等. 水冬瓜赤杨种群生命表与时间序列分析[J]. 北京林业大学学报, 2017, 39(10):62−69. Liu D, Liu S L, Guo Z L, et al. Population life table and time sequence prediction of Alnus sibirica[J]. Journal of Beijing Forestry University, 2017, 39(10): 62−69.
[29] Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology[J]. Oikos, 2004, 104(2): 209−229. doi: 10.1111/oik.2004.104.issue-2
[30] 王鑫厅, 张维华, 姜超, 等. 重复取样条件下的点格局分析[J]. 植物生态学报, 2017, 41(5):577−584. Wang X T, Zhang W H, Jiang C, et al. Point pattern analysis under conditions of replicated sampling[J]. Chinese Journal of Plant Ecology, 2017, 41(5): 577−584.
[31] 张金屯. 植被数量生态学方法[M]. 北京: 中国科学技术出版社, 1995: 79−86. Zhang J T. Methods of exposure vegetation ecology [M]. Beijing: China Science and Technology Press, 1995: 79−86.
[32] He F, Legendre P, Lafrankie J V. Distribution patterns of tree species in a Malaysian tropical rain forest[J]. Journal of Vegetation Science, 2010, 8(1): 105−114.
[33] Ripley B D. Modelling spatial patterns[J]. Journal of the Royal Statistical Society, 1977, 39(2): 172−212.
[34] 宗国, 白雪娇, 张淑媛, 等. 辽东山区次生林乔木幼苗分布格局与种间空间关联性[J]. 应用生态学报, 2018, 29(1):18−24. Zong G, Bai X J, Zhang S Y, et al. Spatial pattern and interspecific spatial association of tree seedlings in a secondary forest in montane region of eastern Liaoning Province, China[J]. Chinese Journal of Applied Ecology, 2018, 29(1): 18−24.
[35] 樊登星, 余新晓. 北京山区栓皮栎林优势种群点格局分析[J]. 生态学报, 2016, 36(2):318−325. Fan D X, Yu X X. Spatial point pattern analysis of Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing[J]. Acta Ecologica Sinica, 2016, 36(2): 318−325.

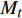


计量
- 文章访问数: 1121
- HTML全文浏览量: 153
- PDF下载量: 25
