
Overexpression mechanism of PeREM1.3 from Populus euphratica enhancing salt tolerance in transgenic tobacco
-
摘要:目的盐害作为一类非生物胁迫严重危害了农作物的生存以及产量。Remorin作为一类植物特有的蛋白质在植物适应环境过程中具有重要功能。本研究克隆了胡杨remorin蛋白PeREM1.3的编码基因PeREM1.3,并研究PeREM1.3基因在植物耐盐性中的作用。方法笔者将基因构件35S::PeREM1.3转入模式植物烟草中, 在盐胁迫条件下,通过生理生化的方法对表达PeREM1.3的转基因烟草进行基因功能的分析。结果研究显示PeREM1.3蛋白定位于细胞质膜上,其编码基因PeREM1.3的开放阅读框(ORF)长600 bp,编码199个氨基酸。胡杨中PeREM1.3能够响应盐胁迫和渗透胁迫表达上调。结果表明,在烟草中过表达PeREM1.3明显地提高了耐盐性。过表达PeREM1.3的烟草转基因株系中抗氧化物酶如SOD、POD和CAT活性显著提高,降低了活性氧水平,调控活性氧平衡。另外植物抗逆相关基因SOS1、HAK、NHA1、VAG1和PMA4的转录水平显著增高,调控K+/Na+平衡。结论这些结果说明PeREM1.3蛋白通过维持植物的活性氧平衡和K+/Na+平衡来提高植物的耐盐性。Abstract:ObjectiveThe tolerance to salt stress, which is a major abiotic stress, is critical to plant survival and productivity. Remorins are plant-specific proteins and play an important role in plant adaptation to adverse environments. In this study, the gene PeREM1.3 coding for a remorin protein was cloned from Populus euphratica, and the role of PeREM1.3 in plant salt tolerance was investigated.MethodThe gene constructs 35S::PeREM1.3 was transferred to model plant tobacco to investigate the function of PeREM1.3 in salt tolerance by physiological and biochemical methods.ResultStudies showed that PeREM1.3 protein localized on the plasma membrane. The 600 bp full-length of open reading frame (ORF) of PeREM1.3 encoded a putative protein of 199 amino acids. The expression of PeREM1.3 was up-regulated under salt and osmotic stress in Populus euphratica. The results showed that the expression of PeREM1.3 increased the salt tolerance of tobacco. The activities of antioxidant enzymes such as SOD, POD and CAT significantly increased and decreased the ROS level to maintain ROS homeostasis in transgenic lines expressing PeREM1.3 under salt stress. On the other hand, the transcriptional levels of plant stress-resistance related genes, such as SOS1, HAK, NHA1, VAG1 and PMA4, were significantly increased and then maintained the K+/Na+ homeostasis in transgenic plants.ConclusionThe above experimental data indicate that remorin protein could maintain ROS and K+/Na+ homeostasis and then enhance plant salt tolerance.
-
Keywords:
- Populus euphratica /
- remorin protein /
- salt tolerance /
- reactive oxygen /
- transgenic tobacco
-
绿色空间是各类自然、人工绿地及水域覆盖的地表空间,良好的绿色空间体系是协调城市发展和自然保护的重要保障[1-2]。在我国城镇化到绿色城镇化的发展历程中,学者不断梳理绿色空间与生态环境的关系,将绿色空间特征与调节城市热环境[3]、评估生态系统服务[4]、促进公众健康等目标有机融合[5],在城市群[6]、省市县[7]、村镇[8]等多尺度研究中广泛运用。其中,土地利用的快速转变影响了绿色空间的景观结构和生态功能,在城市区域尺度的响应尤为突出[9]。
自19世纪初期景观生态学萌芽,到分别以自然景观和人类活动为主要研究对象的路径分野,再到基于地理信息系统的空间定量计算 [10] ,区域绿色空间研究经历了静态向过程、客体向主体、现象向规律等扩展。明确绿色空间时空演变的规律和机制是区域绿色空间体系优化的依据,近年在学界形成广泛共识。其中,演变的研究主要涉及结构指标的计算,基于多时段遥感影像解译,通过 Fragstats软件计算绿色空间景观格局指数(landscape pattern indices,LPI)或景观形态空间格局分析(morphological spatial pattern analysis,MSPA)分析绿色空间格局演化[11],也有学者关注了各类绿色空间面积比例的城乡梯度特征 [12]。既有研究多直接采用土地利用一级分类划分区域绿色空间类型,一般分为林地、草地、耕地、水域和未利用地,忽略了植被覆盖密度及其生态价值的差异[12];而土地利用二级分类数量繁多,不便于以区域绿地规划为目标的研究采用。归一化差分植被指数(normalized difference vegetation index,NDVI)的遥感估算能够弥合这一缺陷,以植被覆盖密度为依据合并土地利用二级分类,进而划分绿色空间类型[13]。演变机制的研究则重点涉及空间效应的测度[14]。在空间效应特征识别方面,主要途径是通过莫兰指数(Moran’s I)、热点分析(Getis-Ord Gi*)、地理加权回归(geographically weighted regression,GWR)等空间统计方法测度绿色空间类型在研究区内的相关或异质性[15];在空间效应的驱动判断方面,多基于地理探测器等工具测度潜在影响因子的显著性和影响力 [16]。不局限于理论研究,国土空间、公园城市规划等实际操作中逐渐关注可选择的规划模式以确保生态系统服务价值等目标的最大实现[17],识别绿色空间时空演变特征是模拟预测的前置环节。
黄河下游是我国工农业生产核心区之一[18],沿岸城市发展是中国城镇化的典型缩影[19]。历史以来,河流与滩区为下游城市创造了丰厚独特的农业条件和农耕文化基础,也参与构建区域绿色空间,影响城市生态安全[20]。《黄河流域生态保护和高质量发展规划纲要》提出,有效推进下游绿色生态走廊建设和滩区生态综合整治。目前,黄河下游城市区域绿色空间时空演变研究数量有限,既有研究中尚缺对于滩区空间的考虑。基于上述认识,本文以10年为间隔,将1990、2000、2010、2020年作为时间节点,基于土地利用二级分类和植被覆盖密度划分黄河下游4个典型城市区域绿色空间类型,识别绿色空间类型转移、景观格局与城乡梯度等时空变化规律,并以地理探测方法揭示绿色空间演变的驱动机制,旨在丰富和深化黄河下游地区绿色空间发展的研究成果,为优化国土空间格局、协调生态保护与经济发展提供参考。
1. 研究区概况与数据
1.1 研究区概况
根据黄河下游自然环境的异质性和城市发展的代表性,选取郑州、新乡、济南、菏泽4城市市域范围为研究区(图1)。从自然环境看,黄河主河道流经郑州、新乡、菏泽边缘,贯穿济南市域空间;郑州市、新乡市、菏泽市境内为黄河宽滩区,其中新乡市封丘县境内还有一定面积的倒灌区,济南市境内则均为窄滩区[21]。此外,济南、郑州、新乡境内有山地分布,菏泽全境地形均为平原。从城市发展看,济南和郑州为区域性中心城市,40多年间经历了剧烈的城市化进程,新乡、菏泽则在近10年间经济增速较快。各城市基本情况见表1。
表 1 研究城市基本情况Table 1. Summary of selected cities城市 City 常住人口数量
Permanent resident population2020年国民生产总值/亿元
GDP in 2020/108 CNY行政区面积
Administrative area/km2济南 Jinan 920.2 × 104 10 140.9 10 220.0 菏泽 Heze 879.6 × 104 3 483.1 12 142.8 郑州 Zhengzhou 1 260.1 × 104 12 691.0 7 515.5 新乡 Xinxiang 617.1 × 104 3 232.5 8 271.0 1.2 数据来源
研究主要数据来源如下:
(1)土地利用/覆盖数据(https://www.resdc.cn/)。来源于由中科院地理所构建的中国多时期土地利用/土地覆盖遥感监测数据库(CNLUCC)。1990—2010年、2020年数据分别以Landsat-TM和Landsat-8遥感图像为信息源,采用5 月上旬至10 月中旬无云图,经波段提取、色彩合成、图像纠正与解译处理等过程生成,数据库Kappa系数为0.889,空间分辨率为30 m × 30 m。
(2)水系数据(https://www.webmap.cn/)。来源于国家基础地理信息中心发布的全国1∶100万公众版地理信息数据库。
(3)滩区类型数据。来源于既有文献中的判断和划定,将研究区划分宽滩区、倒灌区、窄滩区、涉滩区和滩外区[21-23]。
(4)土壤类型数据(https://www.resdc.cn/)。来源于中科院地理所发布的中国土壤类型空间分布数据库。
(5)高程数据(https://search.asf.alaska.edu/#/)。来源于NASA官方网站发布的ALOS卫星12.5 m DEM数据库。
(6)重点城镇中心点空间数据。包括4个城市主城区与周边的一、二级城镇[24],结合中国县级行政边界数据与百度地图识别获得政府所在地的空间点位。
(7)各区县经济、人口和产业数据。整理自各市统计局发布的历年统计年鉴及当年国民经济和社会发展统计公报。
2. 研究框架与方法
2.1 研究框架
研究以类型转移、景观格局和城乡梯度变化作为基本表征,识别绿色空间演变规律;进而进行自然环境和社会经济因素的地理探测,分析绿色空间演变机制(图2)。
2.2 研究方法
2.2.1 空间分类
根据既有研究对绿色空间的界定[4,25],结合CNLUCC 数据分类体系说明书,将研究区域划分为裸露地表和3类绿色空间分别为低覆盖密度绿色空间、中高覆盖密度绿色空间和高覆盖密度绿色空间(以下称为低、中、高密度绿色空间)。裸露地表包括建设用地及植被覆盖度在5%以下的空间。低密度绿色空间包括耕地及覆盖度为5% ~ 50%的草地。中密度绿色空间包括郁闭度 > 40%的矮林地与灌丛林地、郁闭度为10% ~ 30%的林地和苗圃、覆盖度 > 50%的草地及坑塘等小面积水体。高密度绿色空间包括郁闭度 > 30%的林地及河渠、湖泊、水库等大面积水体[25]。
2.2.2 空间转移矩阵
通过计算研究期初各类型面积转出和研究期末各类型空间面积转入的数值构建各类空间转移矩阵,以反映4个城市30年间各类空间相互转化的动态过程信息。转移矩阵的通用形式如下:
{{\boldsymbol{S}}_{ ij}} = \left[ {\begin{array}{*{20}{c}} {{s_{11}}}&{{s_{12}}}& \cdots &{{s_{1n}}} \\ {{s_{21}}}&{{s_{22}}}& \cdots &{{s_{2n}}} \\ \cdots & \cdots & \cdots &{{s_{3n}}} \\ {{s_{n1}}}&{{s_{n2}}}& \cdots &{{s_{nn}}} \end{array}} \right] (1) 式中:Sij为研究初期到末期时间段内i类向j类空间转移的面积矩阵;s为空间转移面积;n为空间总类型,本研究中n = 4。空间转移方向与转移量在ArcMap 10.5软件中计算,后在Origin 2021平台上绘制桑基图。
2.2.3 景观格局指数计算
选取表征空间斑块面积、密度、形状和聚散性等维度特征的6个景观格局指数,分别为斑块面积比(percentage of landscape,PLAND)、斑块数量(number of patches,NP)、最大斑块面积比(largest patch index,LPI)、斑块平均面积(mean patch size,MPS)、面积加权平均形状指数(area-weighted mean shape index,AWMSI)、功能连通度(functional connectivity,FC)[10]。所有景观格局指数均在Fragstats 4.2软件中计算。
2.2.4 距离指数与城乡梯度计算
参照中外学者对于“城市中心”的普遍定义,以各空间斑块与研究区4城市CBD的欧氏距离表征所处的城乡梯度[26]。考虑城市之间的横向比较,引入归一化距离指数(normalized distance index,NDI),其公式如下:
{\rm{NDI}} = \displaystyle \sum\nolimits_{i=1}^n {\frac{{\sqrt {{{\left( {{x_i} - X} \right)}^2} + {{\left( {{y_i} - Y} \right)}^2}} }}{{{d_{\max }}}}} (2) 式中:X、Y表示城市中心的横纵坐标,xi和yi表示绿色空间斑块中心的横纵坐标,分子即为斑块i中心到城市中心的欧氏距离,dmax为所有斑块中心到城市中心欧式距离中的最大值。欧氏距离在ArcMap 10.5软件中计算,以0.01个距离为单位计算各类空间面积占比,后在RStudio 3.3.1 平台上调用ggplot2 软件包,插入“Loess”统计曲线模型,基于局部加权回归原理形成城乡梯度轨迹曲线[27]。
2.2.5 影响因素的地理探测
使用地理探测器工具中的因子探测计算影响因子对绿色空间地理分布的影响,使用交互探测计算两两因子的共同作用。地理探测器是一种探测自变量对因变量空间异质性解释力的统计学方法,基本假设为:如果某种自变量能够影响因变量的地理空间分布,那么自变量与因变量应具有相似的地理空间特征[28]。
{q} = 1 - \frac{1}{{n{\sigma ^2}}}\sum\nolimits_{h = 1}^L {{n_h}} \sigma _h^2 (3) 式中:q为影响因子对绿色空间地理分布的解释力;n、
{\sigma ^2} 分别为整个研究区域中绿色空间地理分布的样本量和方差,{n_h}、\sigma _h^2 分别为次级区域h中绿色空间地理分布的样本量和方差。q取值范围为[0,1],数值越接近1表示解释力越强。在地理探测步骤,对所有数值型数据进行离散化处理得到类型栅格数据集,建立渔网并作采样。3. 结果与分析
3.1 绿色空间转移面积
由表2计算,1990—2020年研究区各类空间转移面积为5 139 km2,占总面积的13.47%。绿色空间总计向裸露地表转出3 223 km2;裸露地表向各类绿色空间转出1 181 km2;绿色空间内部互相转换735 km2,其中较高密度向较低密度绿色空间转出466 km2,较低密度向较高密度绿色空间转出269 km2。4个城市转移量按比例大小排顺序为济南、菏泽、郑州、新乡。山东省两城市绿色空间呈现一增三减的变化,裸露地表面积增加,高、中、低密度绿色空间面积减少。河南省两城市为二增二减,裸露地表与高密度绿色空间面积增加,中低密度绿色空间面积减少。各级城镇边缘、宽滩区沿线绿色空间转移明显(图3)。
表 2 1990—2020年绿色空间转移矩阵Table 2. Matrix of greenspace transfer from 1990 to 2020km2 年份 Year 城市 City 指标 Index 2020 裸露地表
Bare surface低密度
Low density中密度
Medium density高密度
High density1990 济南 Jinan 裸露地表 Bare surface 904 296 1 36 低密度 Low density 1 137 5 319 6 85 中密度 Medium density 16 19 349 3 高密度 High density 115 139 1 1 792 菏泽 Heze 裸露地表 Bare surface 1 462 532 1 25 低密度 Low density 1 005 8 727 0 66 中密度 Medium density 9 35 6 0 高密度 High density 26 106 0 139 郑州 Zhengzhou 裸露地表 Bare surface 1 453 54 0 54 低密度 Low density 583 4 098 1 65 中密度 Medium density 24 56 141 21 高密度 High density 37 57 6 864 新乡 Xinxiang 裸露地表 Bare surface 1 166 127 0 55 低密度 Low density 253 5 473 0 20 中密度 Medium density 0 4 43 2 高密度 High density 18 42 1 1 062 ![]() 图 3 1990—2020年绿色空间转移的空间分布Figure 3. Distribution of greenspace transfer from 1990 to 2020
图 3 1990—2020年绿色空间转移的空间分布Figure 3. Distribution of greenspace transfer from 1990 to 202030年间,低密度绿色空间一直是裸露地表面积的主要转入来源(图4)。2000—2010年间,济南、菏泽两市低密度绿色空间与裸露地表大量相互转化,主要是受2006年山东省出台的《城镇建设用地增加与农村建设用地减少相挂钩管理办法》等相关政策影响[29],大量零星分散的农村居民点发生搬迁撤并或土地流转。新乡、郑州则主要是沿黄滩涂向耕地的转移。
![]() 图 4 1990—2020年绿色空间转移桑基图Figure 4. Sankey diagram of greenspace transfer from 1990 to 2020
图 4 1990—2020年绿色空间转移桑基图Figure 4. Sankey diagram of greenspace transfer from 1990 to 20203.2 绿色空间景观指数变化
3.2.1 景观指数总体变化
由1990—2020年景观指数变化图(图5)可知:在斑块面积比(PLAND)方面,郑州和济南低密度绿色空间数值减少较多,分别为6.41%和6.50%,裸露地表分别增加7.13%、9.14%,均远高于平均值。中、高密度绿色空间变化较小。
![]() 图 5 1990—2020年绿色空间景观指数总体变化PLAND.斑块面积比;NP.斑块数量;LPI.最大斑块面积比;MPS.斑块平均面积;AWMSI.面积加权平均形状指数;FC.功能连通度。下同。PLAND, percentage of landscape; NP, number of patch; LPI, largest patch index; MPS, mean patch size; AWMSI, area-weighted mean shape index; FC, functional connectivity. The same below.Figure 5. Overall changes of landscape pattern indices from 1990 to 2020
图 5 1990—2020年绿色空间景观指数总体变化PLAND.斑块面积比;NP.斑块数量;LPI.最大斑块面积比;MPS.斑块平均面积;AWMSI.面积加权平均形状指数;FC.功能连通度。下同。PLAND, percentage of landscape; NP, number of patch; LPI, largest patch index; MPS, mean patch size; AWMSI, area-weighted mean shape index; FC, functional connectivity. The same below.Figure 5. Overall changes of landscape pattern indices from 1990 to 2020斑块数量(NP)方面,除济南低密度绿色空间与裸露地表数值增加外,其余各城市均有所减少,低、中、高密度绿色空间减少量均以郑州为最大,新乡次之,可见4个城市大多数绿色空间斑块均发生整合。
最大斑块面积比(LPI)方面,4个城市裸露地表数值均有所增加,低密度绿色空间数值均有减少,以郑州与济南的变化量最大,均达到平均水平的2倍以上。中、高密度绿色空间数值变化量相对较小,个体斑块面积增减量未影响整体优势性,表明大规模自然林地格局总体平稳。
面积加权平均形状指数(AWMSI)方面,济南裸露地表数值增加明显,低密度绿色空间数值减少。在NP增加的情况下,表明无植被覆盖的城乡建设用地向外延伸而降低了整体形态的规则程度,同时耕地、草地被人工方整切割。此外,郑州和新乡裸露地表的数值也有较大程度的减少,在PLAND增加、NP减少的情况下,能够反映出郑州城乡建设用地以集聚扩张的形式增长,具有向心性特征。
平均斑块面积(MPS)指数方面,变化主要发生于低密度绿色空间。除济南数值减少外,郑州、新乡、菏泽均有增加。结合NP值变化,可知济南的大面积的低密度绿色空间被切分独立;菏泽低密度绿色空间数值增加最大,表明大量小碎斑块转出。新乡的高密度绿色空间数值增量最大,河南城市增量明显大于山东城市。
功能连接度(FC)方面,郑州、济南的低密度绿色空间,郑州、济南、菏泽的中密度绿色空间,以及济南的高密度绿色空间数值有所增加,且区域性中心城市的改善更为全面。同时,4个城市的裸露地表的数值变化较小,在NP减少、PLAND增加的前提下,较小的FC变化表明斑块之间的最大可能连接数变小,即斑块面积的增加与数量的减少在空间中是相对均匀发生的,反映了城市与村镇建设用地共同发生扩张。
3.2.2 景观格局指数变化轨迹
由1990—2020 年景观格局指数变化折线图(图6)可知, 4个城市区域绿色空间景观格局指数在过去 30 年间的变化轨迹具有一定共性。低密度绿色空间的PLAND、AWMSI、FC,中密度绿色空间的PLAND、NP、LIP、AWMSI,高密度绿色空间的各项指数及裸露地表的PLAND、NP、LIP、MPS、FC变化轨迹均较为相似。比较特殊的轨迹表现于以下方面:郑州中密度绿色空间的MPS、AWMSI和FC的基数在30年内均为最大,其中MPS在1990—2010年间的上升趋势强于其他3市,2010年以来与其他城市呈现一致的平稳趋势。济南裸露地表的AWMSI在2000—2010年间发展迅速并超越郑州至首位,空间限制使建设用地形态不规则度上升。同一时期低密度绿色空间的LIP明显单调下降,耕地和草地面积有所收缩;NP单调上升,变化趋势与其他3市相反,并超越郑州发展至首位。上述变化在2010年后恢复平稳,严格的耕地保护和土地集约利用政策效果显现。菏泽低密度绿色空间的MPS、FC基数远大于其他城市,受黄淮海平原农业开发战略影响,相较于其他城市的平稳轨迹经历了先上升后下降的明显变化。与其他城市的平稳趋势相比,新乡高密度绿色空间的MPS在30年间保持速率均匀的单调上升趋势; FC以2000年前后为转折,呈现“∩”型发展趋势,但始终处于首位。
![]() 图 6 1990—2020年景观格局指数变化折线图Figure 6. Point-fold line chart of landscape pattern indices from 1990 to 2020
图 6 1990—2020年景观格局指数变化折线图Figure 6. Point-fold line chart of landscape pattern indices from 1990 to 2020此外,4个城市高密度绿色空间的LIP、AWMSI数值在30年间均无较大波动,由高到低顺序始终为新乡、郑州、济南、菏泽,河南两城市总体高于山东两城市。中密度绿色空间的LIP、MPS、AWMSI值也未发生排位顺序的变化,由高到低顺序为郑州、济南、新乡、菏泽,区域性中心城市总体高于一般城市。
3.3 绿色空间面积比例的城乡梯度变化
![]() 图 7 1990—2020年绿色空间面积占比的城乡梯度变化Figure 7. Urban-to-rural trajectory of the area percentage of greenspace from 1990 to 2020
图 7 1990—2020年绿色空间面积占比的城乡梯度变化Figure 7. Urban-to-rural trajectory of the area percentage of greenspace from 1990 to 2020低密度绿色空间的城乡梯度轨迹在时间序列中的变化较为复杂。1990和2000年,济南的轨迹由中心向外围先缓慢降低再升高,降低区间为山区地带;2010和2020年,由于城市建设用地扩张导致近郊低密度绿色空间比例锐减,而在NDI > 0.750的区间上保持平稳。2000年,郑州的轨迹在0.125 < NDI < 0.375的区间较前期上升,但2010年后又下降,到2020年,在近远郊地带均出现一定程度下降。新乡较早时期的城乡梯度变化轨迹是“W”形,波峰出现在NDI = 0.500的中间地带。2000和2020年,这一波峰逐渐削弱,轨迹逐步转变为“U”形。菏泽市的轨迹则在 30年间均可见波谷,其形态在30年间由“U”向“V”发展,波谷稳定存在于0.375 < NDI < 0.625的区间。
各城市中密度绿色空间的城乡梯度轨迹在时间序列上体现出一致性。济南中心到外围趋于水平,整体缓慢降低。菏泽的轨迹更加平直,且始终处于低位。郑州、新乡两城市的城乡梯度变化轨迹呈现较为相似的“U”形,分别在NDI = 0.50和NDI = 0.375的距离地带出现了小的峰值。新乡在1990—2010年间的轨迹为“U”形,中部地带为较宽波谷。
高密度绿色空间的梯度相对固定,并反映出城市自然本底、城镇布局方式等空间特征。如济南的峰值出现在NDI = 0.500的中段地带,郑州则在NDI > 0.750的区间上升,均对应于山区自然林地。平原城市菏泽城乡梯度轨迹在30年间没有明显差异,从中心向外保持平稳波动,整体比例处于与中密度绿色空间轨迹类似的低位,最大值出现在远郊水域地带。早期新乡的轨迹在市域中间地带存在小的峰值,后随着城市扩张而被削弱,但可见近郊高密度空间在近年出现增量,主要是依托引黄干渠等水系而发展的城市绿地。
裸露地表的城乡梯度轨迹反映了市域城、镇、村建设用地的发展格局。济南30年间的轨迹表现为单调递增的指数形态,反映城镇布局向外延伸,以及泰山山脉南侧莱芜、钢城两区的发育态势。郑州在1990前年有类似的情况,2000和2010年,在中心城区边缘地带内先后有中牟、荥阳和新郑等城镇发育扩张,使整体变化轨迹呈现出“U”形。新乡的轨迹则在1990和2000年从中心向外围缓慢升高,2010和2020年在NDI = 0.500的距离上形成波峰,对应于第一圈层城镇的扩张成熟。菏泽轨迹的波动幅度不大(图7)。
3.4 绿色空间演变的驱动机制
以绿色空间分布为地理探测中的因变量,以4个表征自然环境的因子(X1 ~ X4)与4个表征社会经济情况的因子(X5 ~ X8)为自变量,计算单一因子、因子间交互解释力。由于1990年的社会经济数据难以全面获取,以下仅探讨2000—2020年间各项因子对绿色空间演变的驱动机制。
根据因子探测的结果(表3),高程、与水系距离、与城镇距离、一产比例、GDP和城乡人口比例6项因子在20年间均对济南区域绿色空间分布有显著贡献,综合解释力由高至低,其中社会经济因素解释力的q累计值上升5.2%。与滩区关系在20年间始终对菏泽区域绿色空间分布显著贡献,除此之外有显著解释力的均为社会经济因素,q累计值上升3.1%。土壤类型外的其他7项因子均在20年间对郑州区域绿色空间分布有显著贡献,高程的解释力最大,社会经济因素解释力的q累积值上升3.9%。自然环境因素在新乡占据明显主导,社会经济因素解释力的q累计值上升8.4%。
表 3 因子探测结果Table 3. Factor detection results城市
City年份
Year与水系距离
Distance from waters (X1)土壤类型
Soil type (X2)与滩区关系
Relationship with floodplain (X3)高程
Altitude (X4)与城镇距离
Distance from town (X5)生产总值
GDP (X6)城乡人口比例
Proportion of
urban and rural
population (X7)一产比例
Proportion of
primary industry (X8)济南 Jinan 2000 0.304*** 0.001 0.003** 0.328*** 0.094*** 0.029*** 0.003*** 0.024*** 2010 0.244 0.001 0.001 0.283*** 0.092*** 0.018*** 0.013*** 0.028*** 2020 0.243*** 0.001 0.001 0.283*** 0.073*** 0.048*** 0.014*** 0.067*** 菏泽 Heze 2000 0.004 0.002 0.005* 0.002 0.004 0 0 0.005** 2010 1.000 0.872 0.011*** 0.002 0.032*** 0.003 0.004* 0.003* 2020 0 0 0.011*** 0.002 0.036*** 0.000 0.001 0.001 郑州 Zhengzhou 2000 0.039*** 0 0.012*** 0.005 0.027*** 0.014*** 0.031*** 0.022*** 2010 0.037*** 0.001 0.012*** 0.194 *** 0.034*** 0.026*** 0.077*** 0.073*** 2020 0.045*** 0.001 0.017*** 0.192*** 0.037*** 0.056*** 0.077*** 0.063*** 新乡 Xinxiang 2000 0.412*** 0.004 0.047*** 0.531*** 0.096*** 0.109*** 0.145*** 0.067*** 2010 0.401*** 0.004 0.043*** 0.526*** 0.103*** 0.166*** 0.086*** 0.082*** 2020 0.393*** 0.005 0.042*** 0.518*** 0.111*** 0.164*** 0.045*** 0.181*** 注:表中数字为解释力q值;*表示在P < 0.1水平上差异显著,**表示在P < 0.05水平上差异显著,***表示在P < 0.01水平上差异显著。Notes: the number in the table is the explanatory power q-value; * means significant differences at P < 0.1 level, ** means significant differences at P < 0.05 level, and *** means significant differences at P < 0.01 level. 根据交互探测结果,菏泽未见因子间的显著交互作用,而郑州、新乡、济南3城市均可见因子共同作用的效果大于独立效果之和的情况,即两两交互的类型为非线性增强。以郑州为例(表4),在2000年仅有土壤类型∩高程产生显著非线性增强效果;2010年增加两对,分别是与滩区关系∩高程、土壤类型∩一产比例;2020年,土壤类型分别与高程、GDP、城乡人口比例、一产比例产生非线性增强效果。表明社会经济因素与自然环境因素体现愈加明显的交互协同。
表 4 交互探测结果(以郑州为例)Table 4. Interactive detection results (taking Zhengzhou as an example)年份
YearX1 X2 X3 X4 X5 X6 X7 X8 2000 X1 0.039 X2 0.003 X3 0.012 X4 0.221 0.205 X5 0.027 X6 0.014 X7 0.031 X8 0.022 2010 X1 0.037 X2 0.001 X3 0.012 X4 0.209 0.209 0.194 X5 0.034 X6 0.026 X7 0.077 X8 0.088 0.073 2020 X1 0.045 X2 0.001 X3 0.017 X4 0.240 0.211 0.192 X5 0.037 X6 0.067 0.056 X7 0.065 0.057 X8 0.073 0.063 注:表中数字为交互作用解释力q值,此表仅显示具有显著性的q值。Notes: the number in the table is the explanatory power q-values of interaction. This table only shows the q values with significant interaction relationship. 4. 讨 论
4.1 区域绿色空间演变特征识别
(1)各类空间的面积增减。研究显示,30年间黄河下游4个城市裸露地表规模持续扩张,低密度绿色空间面积减量最大,中、高密度绿色空间面积增减相对平稳。研究区城乡建设用地共同经历扩张,但区域性中心城市与一般城市之间存在增量和形态上的差别,即新增城市建设用地更多直接接壤城市边缘,集中连片的增长现象更为显著。济南中心城区以山前平原为核心,北向黄河、南向浅山地带逐步延展,在空间的制约中侵占了一定面积中、高密度绿色空间。郑州中心城区的发展以近郊城镇集聚和自身边缘扩张为主,北部边缘向滩区与水域靠近的同时,也建设了有规模的高密度绿色空间,将滩地滩涂转为坑塘湿地和林地为主的生态公园。4个城市内,小而碎的低密度绿色空间均有转出,但城乡建设行为也挤压或分割了大量规模性的耕地、草地。近年来,在严格耕地保护政策出台和城市存量发展导向下,低密度绿色空间减量得到控制。
(2)同类空间的整合连通。30年间,破碎分散向整合连通是各城市区域绿色空间斑块的主要发展趋势。中、高密度绿色空间表现更为明显,其破碎度降低、连通度和集聚水平增加,能够对物种保护、能量交换等生态系统服务价值改善起到积极作用[8],同时可见区域性中心城市的改善尤为全面、明显。此外,济南中心城区低密度绿色空间在2010年以前的形态破碎而不规则,但这一现象在近10年有所改观,反映出山东省土地综合整治等政策的效果。
(3)面积占比的时空分异。30年间,城乡轨迹变化最明显的是区域性中心城市的低密度绿色空间,而各城市中、高密度绿色空间30年间的轨迹具有相对一致性。研究范围内的黄淮海冲击平原区和鲁西平原区是典型的农业景观,耕地为主的低密度绿色空间是最主要的景观类型,外围太行、嵩山、泰山山区内则以中、高密度绿色空间为主,构成了市域重要的生态屏障。在此自然本底上,绿色空间面积比例的城乡梯度本身具有一定特殊性,剧烈的城市化进程中使之发生了不同幅度的变化,与“从城区到郊区,人造绿地数量不断减少,自然绿色空间逐渐增加”的西方经验有所区别[30]。
(4)“自然−社会”的互构作用。研究呈现了“水”“山”“滩”对黄河下游城市区域绿色空间长期性、大尺度的影响,探测了“自然−社会”交互作用的增长,与黄河下游城市“经济系统和谐发展指数最高,环境系统次之”的研究结果相互印证[31]。从同一时期的城乡梯度上看,菏泽、新乡的低密度绿色空间在城市近郊和远郊占比最大,这一现象与两个城市圈层式的布局有关。高密度植被面积比例的城乡梯度轨迹则主要体现了市域范围内山地、滩区等地理单元的空间特征,同时反映出区域政策的引导下城市边缘区生态质量的改变。
4.2 区域绿色空间优化建议
区域绿色空间规划需充分考虑其演变的规律和机制,“因城制宜”。如与其他城市相比,菏泽市域范围自然基底更均质,绿色空间演变多以社会经济发展为动力,需加强规划决策对区域绿色空间建设的助推作用。现状低密度绿色空间在市域范围内均占据主导,应科学规划林地−树团−点状树木等防护绿地结构,丰富农田为主的大尺度国土景观,保障林地、水网为非农生境提供稳定的生物多样性及生态效应[32];中心城区近郊地带中高密度绿色空间面积比例偏低但有较好的连通性基础,水系与林网构成的生态廊道应是优化重点[33]。自然环境因素对新乡区域绿地格局影响力的主导地位最为突出,未来仍需围绕自然要素优化绿色空间格局,将高密度绿色空间面积平稳上升的趋势由山区延伸至近郊与中心城区,进一步发挥滩区与水系的诱导效应,提升滩区沿线、引黄干渠沿线的生境质量。在区域性中心城市郑州、济南,区域绿色空间与社会经济因素关联性日益增强,中心城区周边生态改善趋势显现。未来建设中,一方面需重点考虑城−滩关系,通过资源的保护利用引导现状滩区沿线中密度绿色空间向高密度过度,提升城市边缘区高密度绿色空间的连通性和植被多样性,以公园、绿道建设为引力增进绿色空间建设与经济发展之间的良性互动;另一方面要规划生态分区,防止中、高密度绿色空间规模损失,防范城市建设行为对滩区和山区的重要生境造成扰动。值得注意的是,土壤类型因子在4个城市中往往不具备独立影响力,但与各项表征社会经济的因子交互协同,表明人工绿地建设受到土壤条件的制约。黄河下游尤其是滩区周边城市在进行区域绿色空间规划时,需根据立地条件合理配置不同生长习性的植被,保障区域绿地的可持续发展。
5. 结 论
研究定量测度了1990—2020年间黄河下游4城市区域绿色空间的类型转移、景观格局、城乡梯度的时空变化规律,并揭示了绿色空间演变的驱动机制,主要结论如下:(1)30年间,区域绿色空间总量损失,绿色空间内部转移以中、高密度向低密度的转出为主,各级城镇建成区边缘、宽滩区沿线转移较多;(2)景观格局的总体变化趋势为由明显波动到趋近平稳、由破碎分散向整合连通,中、高密度绿色空间指标的改善态势在区域性中心城市更为明显;(3)城乡梯度特征在城市间、绿色空间类型间存在较大差异;(4)影响因子间对绿色空间地理分布的协同作用渐增,绿色空间逐渐成为自然−社会互构的结果。伴随黄河下游簇群城镇形态的成熟与黄河流域高质量发展战略的深入实施,未来的绿色空间应由“屏障”转为“枢纽”,在区域生态保护和经济社会发展中发挥更多助益。
受限于数据的可获取性,本研究仅对近20年的绿色空间演变机制进行研究,且基于时间节点数据的分析结果相对局限。另一方面,不同遥感数据的差异可能会对比较研究造成干扰。未来将加强遥感、经济、人口等面板数据的追踪分析,构建黄河下游城市多精度LUCC、社会经济、政策发展综合数据库,并深入生态系统服务、市域生态网络构建等多目标研究。
-
![]()
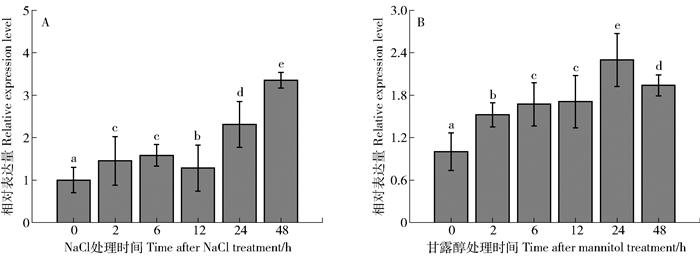
图 1 不同胁迫处理下胡杨PeREM1.3基因在胡杨叶片中的表达量变化
A.NaCl处理下PeREM1.3的表达量;B.甘露醇处理下PeREM1.3的表达量;不同字母表示在P<0.05水平上差异显著。
Figure 1. Expression changes of PeREM1.3 in leaf under different stress treatments in Populus euphratica
A.Expression of PeREM1.3 under NaCl treatment; B. Expression of PeREM1.3 under mannitol treatment; different letters mean significant difference at P < 0.05 level.
![]()
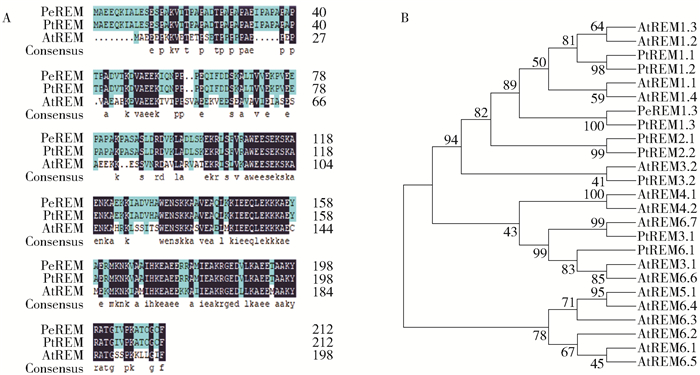
图 2 胡杨PeREM1.3序列分析
A.胡杨PeREM1.3氨基酸序列与其他物种REM多重系列比对; B.胡杨PeREM1.3的系统进化树分析。Pe.胡杨; Pt.毛果杨; At.拟南芥。
Figure 2. Analysis of the amino acid sequence encoded by PeREM1.3
A, multiple amino acid sequence alignment of PeREM1.3 with other REM from different plant species; B, phylogenetic tree analysis of PeREM proteins; Pe, Populus euphratica; Pt, Populus trichocarpa; At, Arabidopsis thaliana.
![]()
图 3 PeREM1.3的亚细胞定位
PeREM1.3-GFP.GFP荧光场;FM4-64.膜染料;Merged.叠加场。
Figure 3. Subcellular location of PeREM1.3
PeREM1.3-GFP, GFP field; FM4-64, plasma membrane dye; Merged, merged field.
![]()
图 4 转基因烟草各株系荧光定量和半定量RT-PCR检测PeREM1.3
PeREM1.3基因在野生型(WT)、转空载体(VC)和转基因烟草(OE1、OE6、OE7、OE9、OE11、OE16、OE18、OE22)中的表达量,EF1α为内参基因。A为荧光定量PCR分析;B为半定量PCR分析。不同字母表示在P<0. 05水平上差异显著。下同。
Figure 4. Quantitative reverse transcription PCR and semi-quantitative reverse transcription PCR analysis of each line in tobacco plants
Expression level of PeREM1.3 of wild-type (WT), vector control (VC) and PeREM1.3-transgenic tobacco (OE1, OE6, OE7, OE9, OE11, OE16, OE18, OE22) plants, EF1α was used as the internal control. A, real-time PCR analysis. B, semi-quantitative sRT-PCR analysis. Different letters mean significant difference at P < 0.05 level. The same below.
![]()
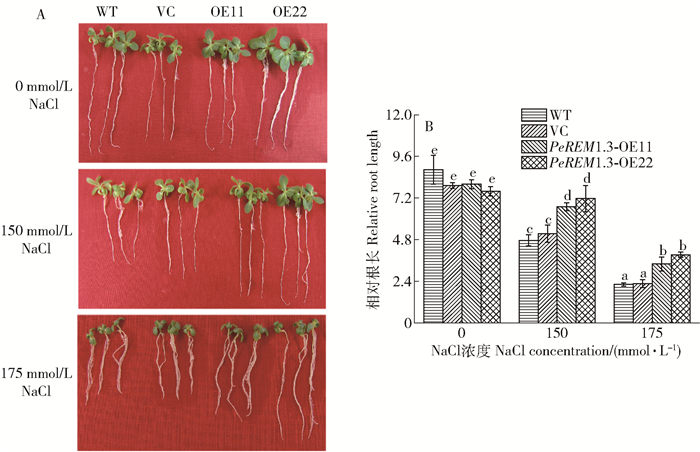
图 5 烟草在含有不同浓度NaCl培养基上的根长生长情况
A. NaCl对根长生长的影响; B.根系生长的定量分析;烟草播种在普通1/2MS培养基上,垂直生长7 d后,移植到含有0、150、175 mmol/L NaCl培养基上,生长5 d后统计根长生长情况并拍照,不同字母表示在P<0.05水平上差异显著。
Figure 5. Root length of tobacco on MS medium supplemented with different NaCl concentrations
A, effect of NaCl on root length; B, quantitative analysis of root growth; seeds of each line were sowed on 1/2 MS medium for 7 days, then transferred to new MS medium with 0, 150, 175 mmol/L NaCl, after 5 days, root length was measured and photographed. Different letters mean significant difference at P < 0.05 level.
![]()
图 6 盐处理后烟草中SOD酶活性(A)、POD酶活性(B)和CAT酶活性(C)变化
A.盐处理下SOD酶活性;B.盐处理下POD酶活性;C.盐处理下CAT酶活性。不同字母表示在P<0.05水平上差异显著。
Figure 6. Activity of SOD (A), POD (B) and CAT (C) in tobacco plants after salt treatment
A, activity of SOD under NaCl treatment; B, activity of POD under NaCl treatment; C, activity of CAT under NaCl treatment; different letters mean significant difference at P < 0.05 level.
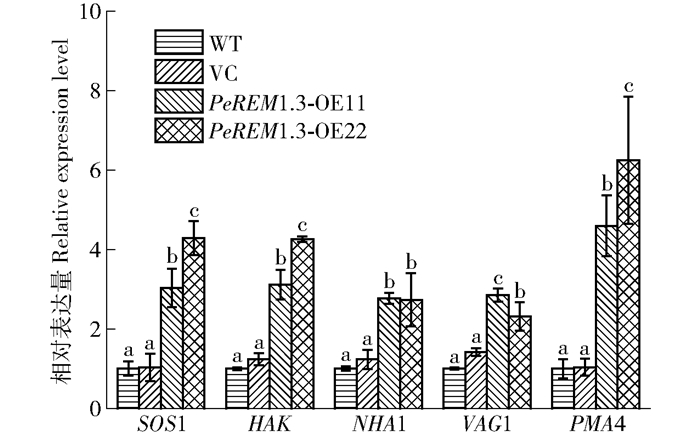
![]()
图 7 盐胁迫对PeREM1.3转基因烟草抗逆相关基因SOS1、HAK、NHA1、VAG1和PMA4表达的影响
内参基因:EF1α,n=3。不同字母表示在P<0.05水平上差异显著。
Figure 7. Effects of NaCl on relative expression of stress- resistance related genes in transgenic tobacco plants
Reference gene: EF1α, n=3. Different letters mean significant difference at P < 0. 05 level.
表 1 本研究中使用的引物序列以及名字
Table 1 Gene-specific primer sequences and names used in this work
引物名称
Primer name上游引物
Forward primer(5′-3′)下游引物
Reverse primer(5′- 3′)HAK ATCCACACCGAGCTTGTTTCAGGA TGGGTCCAATTCTTCCCACCAAGA SOS1 GCGTGCTTATTTCCACCTTTTG TTTGATGACGGCTCCCCAGT PMA4 TTTCCCGAGCACAAGTATGA GGTAACCTCCAAGAACAACAC NHA1 CCTTATGCTTGTCGGTGCTTTC GCCCATTGTGCTTCCCTTTC VAG1 GGCACGTAACCACAGTGAAG AGAAGCAGCCATGCCTAGTC EF1α GCTGTGAGGGACATGCGTCAAA GTAGTAGATATCGCGAGTACCACCA PeActin ATTGGCCTTGGGGTTAAGAG CACACTGGAGTGATGGTTGG PeREM1.3-0029GFP GCTGCAGCATGGCAGAGGAGGAGCCAAAG CCCGGGTAAAAAAATTCCAAGAAGCTTC PeREM1.3-2300 CGGGATCCATGGCAGAGGAGGAGCCAAAG GCGTCGACCTATAAAAAAATTCCAAGAAGCTT PeREM1.3 GTCCAAAGCAGAAAACAAAGCTCA CACGTTTAGCTTCGATAATCGCC PeREM1.3-Fulllength ATGGCAGAGGAGGAGCCAAAG CTATAAAAAAATTCCAAGAAGCTT  下载: 导出CSV
下载: 导出CSV
-
[1] Jacinto T, Farmer E E, Ryan C A. Purification of potato leaf plasm a membrane protein pp34, a protein phosphorylated in response to oligogalacturonide signals for defense and development[J]. Plant Physiology, 1993, 103: 1393-1397. doi: 10.1104/pp.103.4.1393
[2] Peskan T, Westermann M, Oelmuller R. Identification of low-density triton X-100-insoluble plasma membrane microdomains in higher plants[J]. European Journal of Biochemistry, 2000, 267(24): 6989-6995. doi: 10.1046/j.1432-1327.2000.01776.x
[3] Mongrand S, More L J, Laroche J, et al. Lipid rafts in higher plant cells: purification and characterization of Triton X-100-insoluble microdomains from tobacco plasm a membrane[J]. Journal of Biological Chemistry, 2004, 279(35): 36277-36286. doi: 10.1074/jbc.M403440200
[4] Kreps J A, Wu Y, Chang H S, et al. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress[J]. Plant Physiology, 2002, 130(4): 2129-2141. doi: 10.1104/pp.008532
[5] Bray E A. Abscisic acid regulation of gene expression during water-deficit stress in the era of the Arabidopsis genome[J]. Plant, Cell and Environment, 2002, 25: 153-161. doi: 10.1046/j.1365-3040.2002.00746.x
[6] Reddy A R, Ramakrishna W, Sekhar A, et al. Novel genes are enriched in normalized cDNA libraries from drought-stressed seedlings of rice (Oryza sativa L. subsp. indica cv. Nagina 22)[J]. Genome, 2002, 45: 204-211. doi: 10.1139/g01-114
[7] Lin F, Xu S L, Ni W M, et al. Identification of ABA-responsive genes in rice shoots via cDNA microarray[J]. Cell Research, 2003, 13(1): 59-68. doi: 10.1038/sj.cr.7290151
[8] Malakshah S N, Rezaei M H, Heidary M, et al. Proteomics reveals new salt responsive proteins associated with rice plasma membrane[J]. Bioscience, Biotechnology, and Biochemistry, 2007, 71(9): 2144-2154. doi: 10.1271/bbb.70027
[9] Gui J, Zheng S, Liu C, et al. OsREM4.1 interacts with OsSERK1 to coordinate the interlinking between abscisic acid and brassinosteroid signaling in rice[J]. Developmental Cell, 2016, 38: 201-213. doi: 10.1016/j.devcel.2016.06.011
[10] 张一南, 王洋, 张会龙, 等.过表达胡杨PeRIN4基因拟南芥提高质膜H+-ATPase活性和耐盐性[J].北京林业大学学报, 2017, 39(11) : 1-8. doi: 10.13332/j.1000-1522.20170124 Zhang Y N, Wang Y, Zhang H L, et al. Overexpression of PeRIN4 enhanced salinity tolerance through up regulation of PM H+ -ATPase in Arabidopsis thaliana[J]. Journal of Beijing Forestry University, 2017, 39 (11) :1-8. doi: 10.13332/j.1000-1522.20170124
[11] Sun J, Wang M J, Ding M Q, et al. H2O2 and cytosolic Ca2+ signals triggered by the PM H+-coupled transport system mediate K+/Na+ homeostasis in NaCl-stressed Populus euphratica cells[J]. Plant, Cell and Environment, 2010, 33: 943-958. doi: 10.1111/pce.2010.33.issue-6
[12] Sun J, Li L S, Liu M Q, et al. Hydrogen peroxide and nitric oxide mediate K+/Na+ homeostasis and antioxidant defense in NaCl-stressed callus cells of two contrasting poplars[J]. Plant Cell Tissue and Organ Culture, 2010, 103(2): 205-215. doi: 10.1007/s11240-010-9768-7
[13] Ding M Q, Hou P C, Shen X, et al. Salt-induced expression of genes related to Na+/K+ and ROS homeostasis in leaves of salt-resistant and salt-sensitive poplar species[J]. Plant Molecular Biology, 2010, 73: 251-269. doi: 10.1007/s11103-010-9612-9
[14] Chen S L, Polle A. Salinity tolerance of Populus[J]. Plant Biology, 2010, 12 : 317-333. http://d.old.wanfangdata.com.cn/Periodical/bjlydxxb201711001
[15] Sun J, Chen S L, Dai S X, et al. NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species[J]. Plant Physiology, 2009, 149: 1141-1153. http://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper_PM19028881
[16] Wang R G, Chen S L, Ma H, et al. Genotypic differences in antioxidative stress and salt tolerance of three poplars under salt stress[J]. Frontiers of Forestry in China, 2006, 1: 82-88. doi: 10.1007/s11461-005-0019-8
[17] Kraus T E, Fletcher R A. Paclobutrazol protects wheat seedlings from heat and paraquat injury is detoxification of active oxygen involved[J]. Plant and Cell Physiology, 1994, 35: 45-52. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=HighWire000002843271
[18] Abbott A G, Ainsworth C C, Flavell R B. Characterization of anther differentiation in cytoplasmic male sterile maize using a specific isozyme system (esterase)[J]. Theoretical and Applied Genetics, 1984, 67: 469-473. doi: 10.1007/BF00263415
[19] Shen Z D, Yao J, Sun J, et al. Populus euphratica HSF binds the promoter of WRKY1 to enhance salt tolerance[J]. Plant Science, 2015, 235: 89-100. doi: 10.1016/j.plantsci.2015.03.006
[20] Sylvain R, Emmanuelle B, David L, et al. Remorin, a solanaceae protein resident in membrane rafts and plasmodesmata, impairs potato virus X movement[J]. The Plant Cell, 2009, 21:1541-1555. doi: 10.1105/tpc.108.064279
[21] Wang R G, Chen S L, Zhou X Y, et al. Ionic homeostasis and reactive oxygen species control in leaves and xylem sap of two poplars subjected to NaCl stress[J]. Tree Physiology, 2008, 28: 947-957. doi: 10.1093/treephys/28.6.947
[22] Mittler R, Vanderauwera S, Gollery M, et al. Reactive oxygen gene network of plants[J]. Trends in Plant Science, 2004, 9: 490-498. doi: 10.1016/j.tplants.2004.08.009
[23] Zhu M, Shabala L, Cuin T A, et al. Nax loci affect SOS1-like Na+/H+ exchanger expression and activity in wheat[J]. Journal of Experimental Botany, 2016, 67: 835-844. doi: 10.1093/jxb/erv493
[24] Zhu J K. Regulation of ion homeostasis under salt stress[J]. Current Opinion in Plant Biology, 2003, 6: 441-445. doi: 10.1016/S1369-5266(03)00085-2
[25] Blumwald E. Sodium transport and salt tolerance in plants[J]. Current Opinion in Cell Biology, 2000, 12(4): 431-434. doi: 10.1016/S0955-0674(00)00112-5
[26] Wu C A, Yang G O, Meng Q W, et al. The cotton GhNHX1 gene encoding a novel putative tonoplast Na+/H+ antiporter plays an important role in salt stress[J]. Plant Cell Physiology, 2004, 45(5):600-607. doi: 10.1093/pcp/pch071
[27] Fukuda A, Nakamura A, Tagiri A, et al. Function, intracellular localization and the importance in salt tolerance of a vacuolar Na+ /H+ antiporter from rice[J]. Plant Cell Physiology, 2004, 45(2): 149-159. http://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper_PM14988485
[28] Takahashi R, Nishio T, Ichizen N, et al. Cloning and functional analysis of the K+ transporter, PhaHAK2, from salt-sensitive and salt-tolerant reed plants[J]. Biotechnology Letters, 2007, 9(3): 501-506. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=4682b0e4dbd595aaf9c414773231e657
[29] 马挺军, 向远寅, 王沙生.盐胁迫对胡杨液泡膜H+-ATPase水解活性的影响[J].新疆农业大学学报, 2003, 26(2):43-48. doi: 10.3969/j.issn.1007-8614.2003.02.011 Ma T J, Xiang Y Y, Wang S S. Effects of salt stress on the hydrolytic activity of H+ ATPase from Populus euphratica[J]. Journal of Xinjiang Agricultural University, 2003, 26(2):43-48. doi: 10.3969/j.issn.1007-8614.2003.02.011
[30] Moriau L, Bogaerts P, Jonniaux J L, et al. Identification and characterization of a second plasma membrane H+-ATPase gene subfamily in Nicotiana plumbaginifolia[J]. Plant Molecular Biology, 1993, 21(6):955-963. doi: 10.1007/BF00023594
[31] Wang M J, Wang Y, Sun J, et al. Overexpression of PeHA1 enhances hydrogen peroxide signaling in salt-stressed Arabidopsis[J]. Plant Physiology and Biochemistry, 2013, 71(2):37-48. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=585b971772f2d855c1bfb01b4960a634
[32] Shabala S, Cuin T A. Potassium transport and plant salt tolerance[J]. Physiologia Plantarum, 2008, 133(4): 651-669. doi: 10.1111/ppl.2008.133.issue-4
-
期刊类型引用(4)
1. 牟韵洁,高德民,龙腾腾,郭在军,牛海峰. 张家口崇礼区主要植物叶片理化性质及燃烧性分析. 南京林业大学学报(自然科学版). 2024(06): 239-244 .  百度学术
百度学术
2. 王博,韩树文,顾泽,陈锋,白夜,刘晓东. 不同烈度林火对油松林潜在地表火行为的影响. 生态学报. 2023(05): 1812-1821 . 百度学术
3. 韩懂懂,杨光,邸雪颖,李兆国. 兴安落叶松林火烧迹地土壤理化性质驱动因子. 生态学报. 2023(21): 8727-8738 . 百度学术
4. 王博,韩树文,武英达,牛树奎,刘晓东. 辽河源自然保护区油松林火烧迹地林木更新研究. 北京林业大学学报. 2020(04): 41-50 . 本站查看
其他类型引用(6)















计量
- 文章访问数: 2122
- HTML全文浏览量: 565
- PDF下载量: 66
- 被引次数: 10
