
Delayed effects of reproductive costs in dioecious species Acer barbinerve
-
摘要:目的植物将大部分资源用于繁殖活动时必将导致其对生长发育等其他功能的资源投入下降,生活史理论把这种现象称为繁殖代价。植物繁殖代价主要体现为当年繁殖资源投入与当年营养生长资源投入之间的权衡,以及当年繁殖资源投入与未来生长发育资源投入之间的权衡,这意味着植物的繁殖代价可能存在着一定的延迟效应。本文利用雌雄异株植物雌雄功能分离的属性,验证吉林蛟河针阔混交林中簇毛槭是否存在繁殖代价的延迟效应。方法2012年选取簇毛槭雌雄各90株,将不同性别植株分成3个处理组(A为对照组,B为摘除花朵即开花不结实组,C为摘除花蕾组)进行试验操作。2013年和2017年跟踪调查不同处理组中簇毛槭繁殖及生长状况。利用双因素方差分析检验不同处理方式下雌雄植株的1年生枝条长度和叶片生物量的差异显著性。结果2013年花期,在1年生枝条水平和2级侧枝水平上1年生枝条长度顺序为C组 > B组 > A组。其中雌株的B组和C组显著大于A组,雄株的处理组间差异显著。2013年花期,在1年生枝条水平和2级侧枝水平上雌株的叶片生物量顺序为C组 > B组 > A组;其中B组和C组显著大于A组。2017年花期,在1年生枝条水平雄株的叶片生物量顺序为C组 > B组 > A组,其中C组显著大于A组。结论构件水平影响着繁殖代价延迟效应的检测结果;雌雄异株树种簇毛槭的繁殖代价的延迟效应在营养器官上的表现具有性别和养分分配差异;繁殖代价延迟效应持续表现时间也具有性别差异。Abstract:ObjectivePlant resource allocation is limited such that an increase reproduction incurs a reduction in other functions, quantified as reduced current growth, development or survival. This phenomenon is called reproductive cost based on the theory of life history evolution. Sometimes the cost of reproduction can be shown in the trade-off between the cost of reproduction in the current year and the growth in the future, suggesting that the reproductive cost has a delayed effect. Dioecious plant Acer barbinerve, in which reproductive functions are separated into two distinct sexual forms, is especially suitable for studying the delayed effect of reproductive cost in a coniferous and broadleaved mixed forest in Jiaohe, Jilin Province of northeastern China.MethodTo explore the delayed effect of reproductive cost, we selected 90 female and 90 male individuals with no significant difference in size of dioecious species Acer barbinerve in 2012. The individuals of each sex were divided into three treatments (A, control; B, removing flower called flowers only without fruits; C, removing flower bud). We investigated the reproduction and growth of Acer barbinerve in different treatments in 2013 and 2017. Two-way ANOVAs was performed to test the difference of length of new shoot and foliage biomass for each sex under different treatments.ResultIn flowering time in 2013, the sequence of length of new shoot was group C > group B > group A at shoot and branch level, in which the difference of group B and group C with group A was significantly separated. The differences under different treatments of male were significant. In flowering times in 2013, the foliage biomass of female was group C > group B > group A at shoot and branch level. The leaf biomass of male plants in the flowering times at shoot level of 2017 was C > B > A, in which group C was significantly larger than group A.ConclusionThe delayed effect of reproductive cost of dioecy Acer barbinerve was influenced by module level and showed sexual difference in different vegetative organs and nutrient distribution, also had gender differences in the duration.
-
Keywords:
- reproductive cost /
- dioecy /
- Acer barbinerve /
- delayed effect
-
磨削是木材加工中必不可少的重要工序,所用磨具大多为砂带,主要用于工件定厚和表面抛光。砂带磨削具有“冷态磨削”和“弹性磨削”的特点,是一种高精度、高效率、低成本的加工技术。材料去除率和表面粗糙度作为最常用的砂带磨削性能指标,是衡量砂带磨削效率、砂带寿命,以及评定加工质量的重要指标。材料去除率是指磨削过程中砂带在单位时间内去除被磨削材料的质量百分比,该指标直接影响到工件的定厚效果和表面粗糙度。影响砂带材料去除率的主要因素有砂带制造工艺、砂带目数、磨削压力、磨削速度和磨削延续长度。任青剑等[1]进行了300M超高强度钢的切入式磨削实验,发现在微观形貌方面,磨粒间距较大的砂带多是因磨粒磨损而失效,这是磨削效率降低的一个原因,但针对磨削效率降低的其他原因及其影响因素的探究仍然不够深入。Torrance[2]建立了磨料磨损模型,并将该方法应用于磨削过程中磨削力和金属材料去除率之间关系的预测以及磨料劣化的预测中。由于木材种类繁多,各项性质差异较大,此模型在木材磨削领域的应用存在局限性,仍需进一步研究探索。表面粗糙度的影响因素主要是关于木材特性的,包括纹理、密度、含水率等。刘博等[3]研究发现磨削表面质量会受到木材材性和加工工艺等因素的影响。Tian等[4]使用磨削效率测试系统和表面粗糙度测量仪进行磨削实验,分析比较了3种试材磨削效率和表面粗糙度的变化规律以及相关因素对磨削效率和表面粗糙度的影响,结果表明:磨削效率与表面粗糙度的变化趋势是随磨削次数的增加而逐渐降低,且同一树种、相同的磨削次数下,横向磨削时的磨削效率高于纵向磨削时的磨削效率。该实验在较为宏观的层面下进行探讨,缺乏对于砂带与试件表面微观形貌的进一步研究与分析。
砂带在磨削材料过程中将不可避免地产生磨损,且砂带的磨损是多个因素共同作用的结果,如磨削压力、砂带速度、工件材料等,在磨削不同材料或不同的磨削条件下的砂带磨损形式也会有差异。Ferguson等[5]在进行磨料磨损实验时,通过改变磨料磨粒尺寸、磨削压力、滑动速度和滑动距离来评估复合材料的磨损率。Malinov等[6]发现随着磨削压力的增加,Fe-B合金的耐磨性降低,磨损机制从微切割变为微切割和微耕的混合模式。在实际生产中,砂带生产厂家无法提供准确的砂带使用寿命,多依赖熟练工人的经验来判别砂带的使用寿命,这在一定程度上影响企业的生产效率和经济效益。随着人工智能和传感技术的发展,木材加工智能化已成为重要的发展方向,智能化控制一方面是提高机械设备的智能化控制程度,更重要的一方面是使切削(包括磨削)过程和结果可控,也就是根据要求达到相应的材料去除效率和表面加工质量[7]。比如美国将砂带磨削应用于汽车制造业的FMS生产线上,从磨削过程和设备控制两方面共同调控以达到最终的生产过程智能化。美国3M公司发明的微晶复制砂带使磨粒形状细致均匀且统一,使得磨削过程变得更加精密和高效[8]。
有关木材磨削方面的研究基础较为薄弱,而金属磨削理论并不完全适用于木材,对于木材砂带磨损和磨削效率方面的研究则是少之又少,砂带磨损机理也尚不明晰,这严重阻碍了木材加工技术和砂带制造技术向高效智能方向的发展。针对以上问题,本研究对水曲柳试件开展砂带磨削实验,分析砂带磨损机理及磨粒磨损对材料去除率和表面粗糙度的影响,进一步完善木材磨削理论,并为发展适用于木材的砂带磨削技术提供理论支持。
1. 材料和方法
1.1 材 料
水曲柳(Fraxinus mandshurica)材质坚韧致密,富弹性,纹理通直,刨面光滑,胶接、油漆性能较好,具有良好的装饰性能,在建筑、飞机、造船、仪器、运动器材、家具等行业应用广泛,因此本研究选用水曲柳作为实验材料,其气干密度为0.76 g/cm3,含水率为5.41%。制备的试件尺寸为46 mm(长) × 46 mm(宽) × 24 mm(厚);所有试件取自边材,表面平整,无明显缺陷。本研究所选用砂带为60目普通布基疏植砂型砂带(JW342,DEER,韩国)。
1.2 研究方法
1.2.1 砂带磨削实验装置
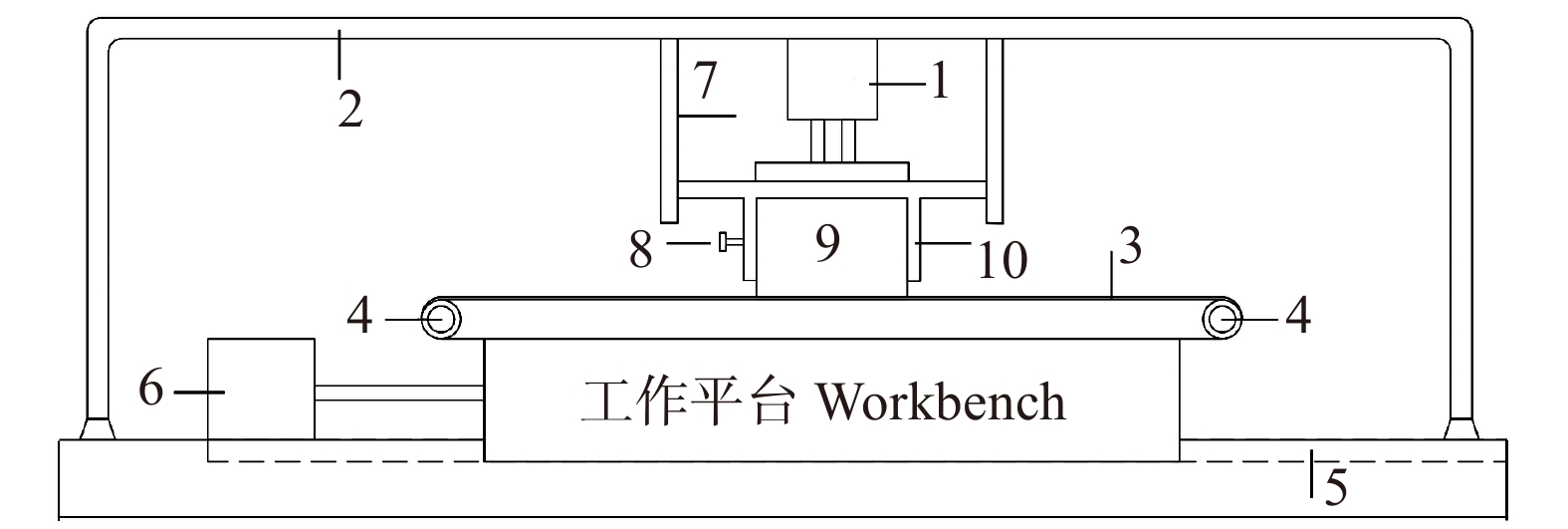
本研究所使用的实验装置如图1所示。该装置主要由使工作平台作直线往复运动的驱动系统和提供恒定磨削压力的气动装置组成。将砂带展平放置在工作平台上,调节两端的张紧轮,确保砂带张紧固定;将木制试件放置于夹具内,调节夹紧螺母完成试件的装夹;通过立式布置的带直线导轨的气缸(ADNGF-50-40-A,FESTO,德国),为试件提供恒定的磨削压力(约为100 N);随着试件材料不断去除,其厚度也逐渐减小,试件夹具可沿立式滑轨在气缸导杆行程内下移,使试件与砂带表面始终相接触;驱动系统带动工作平台沿水平滑轨作往复直线运动(平均速度约为0.3 m/s),同时智能计数装置完成磨削次数记录。
![]() 图 1 砂带磨削实验装置示意图1. 气缸Cylinder;2. 机架Framework;3. 砂带Abrasive belt;4. 张紧轮Tensile wheel;5. 水平滑轨Horizontal slideway; 6. 驱动系统Driver system;7. 竖直滑轨Vertical slideway;8. 夹紧装置Fasten device;9. 试件Sample;10. 夹具Fixture.Figure 1. Schematic of abrasive belt sanding apparatus
图 1 砂带磨削实验装置示意图1. 气缸Cylinder;2. 机架Framework;3. 砂带Abrasive belt;4. 张紧轮Tensile wheel;5. 水平滑轨Horizontal slideway; 6. 驱动系统Driver system;7. 竖直滑轨Vertical slideway;8. 夹紧装置Fasten device;9. 试件Sample;10. 夹具Fixture.Figure 1. Schematic of abrasive belt sanding apparatus1.2.2 材料去除率表征及表面形貌分析
在本研究中,材料去除率定义为每磨削1 000次后试件的质量变化率。每磨削一次则磨削长度为50 mm,可将磨削次数累计与磨削长度进行换算。实验过程中,采用精密分析天平(BSA4235,Sartorious,德国)对试件和砂带进行称重,且每次称重前使用高压气枪(额定压力为3.0 MPa)去除试件和砂带表面的磨屑,进而计算相应的材料去除率和砂带的质量变化情况,直至材料去除率降至3%以内,认为此时砂带寿命已达极限,砂带上的磨粒已无法完成对试件材料的有效切除。采用3D轮廓仪(VR5000,KEYENCE,日本)对磨削过程中试件磨削表面和砂带表面进行扫描,再通过专业分析软件(VR Series version 3.2.0.277)测定试件磨削表面以及砂带的表面粗糙度,并使用扫描电子显微(日立S-3400N Ⅱ)对试件的磨削表面形貌进行分析。实验中对水曲柳分别进行顺纹磨削(磨削方向平行于木材纹理方向)和横纹磨削(磨削方向垂直于木材纹理方向)。
2. 结果与讨论
2.1 横纹磨削
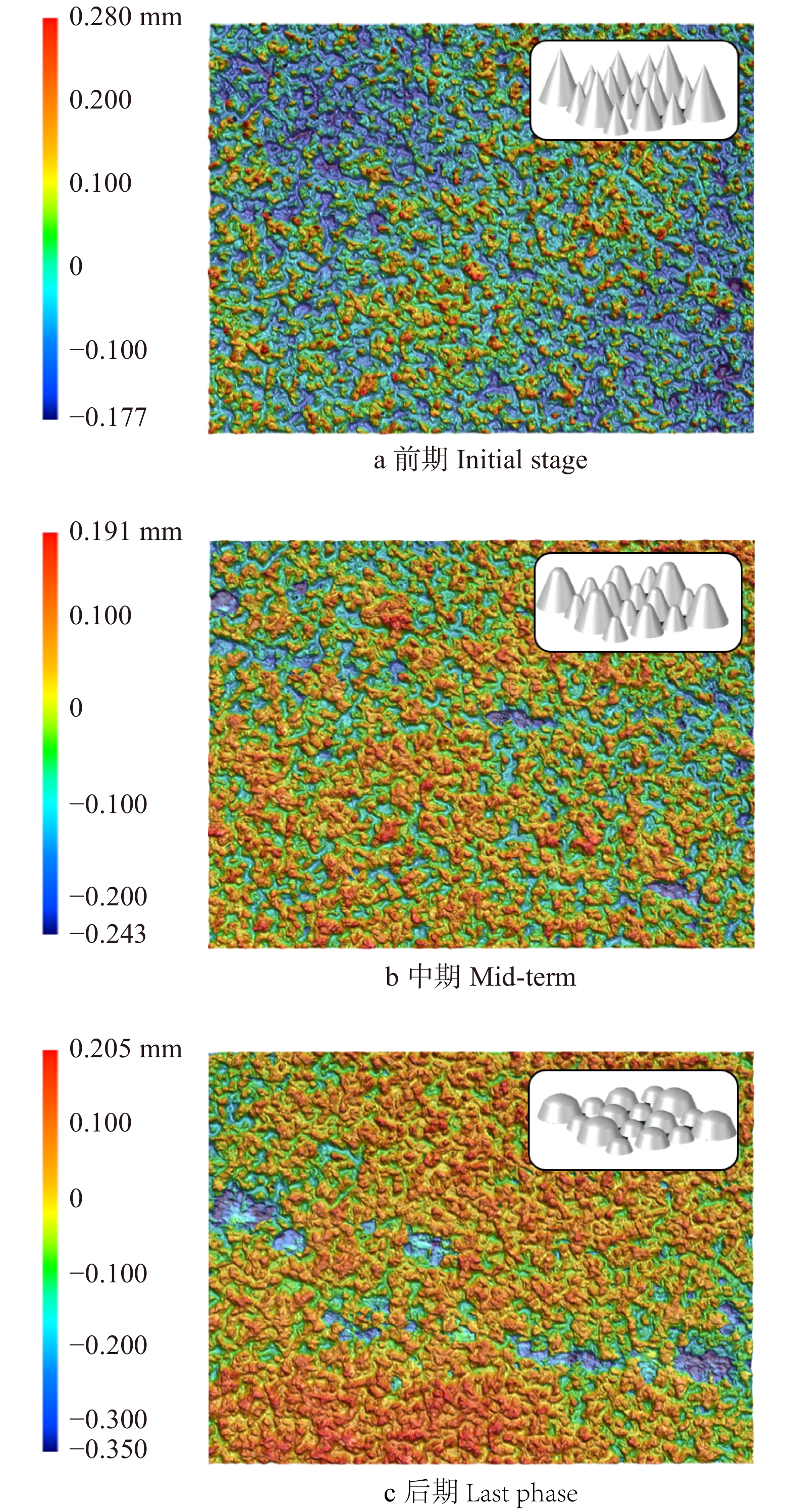
横纹磨削时,磨粒的切削方向与木材纤维方向基本垂直,磨粒多呈负前角或小后角,锋利的刃口将木材纤维拦腰截断,木材纤维基本上以剥离的形式去除[9-10]。观察发现图2a中有较多细窄且清晰的单个磨粒磨削痕迹,而图2b、2c中这种磨削痕迹则逐渐减少,直至较宽的磨削痕迹占绝大多数。由于现有的磨粒制备技术及植砂工艺,新砂带上的磨粒等高性不好,如图3a所示,切削刃曲率半径很小且参差不齐,所以磨削初期会在试件表面上留下细窄、清晰的磨削痕迹。
![]() 图 2 横纹磨削时水曲柳表面形貌与磨粒磨损示意图Figure 2. Surface topography and grit wear diagram during transverse sanding
图 2 横纹磨削时水曲柳表面形貌与磨粒磨损示意图Figure 2. Surface topography and grit wear diagram during transverse sanding![]() 图 3 横纹磨削时砂带表面三维形貌图Figure 3. Three-dimensional topography of the belt surface during transverse sanding
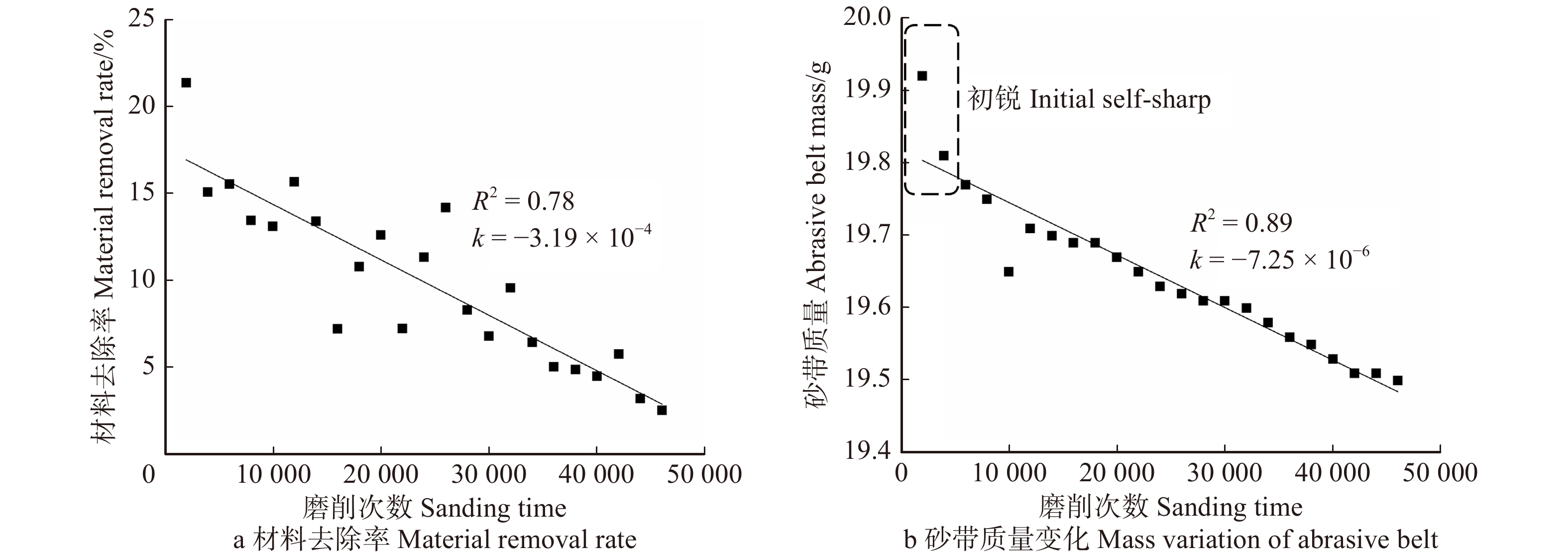
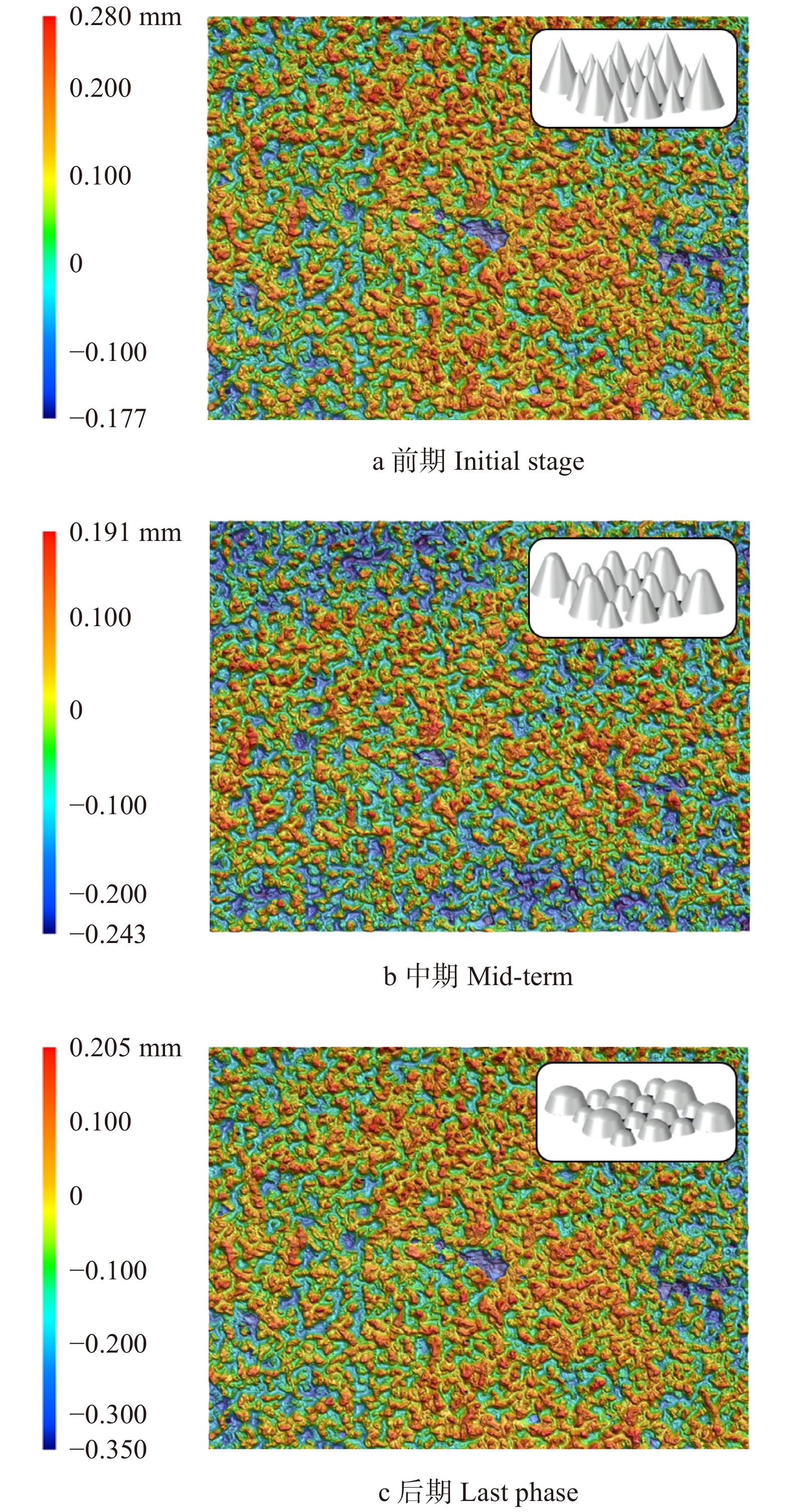
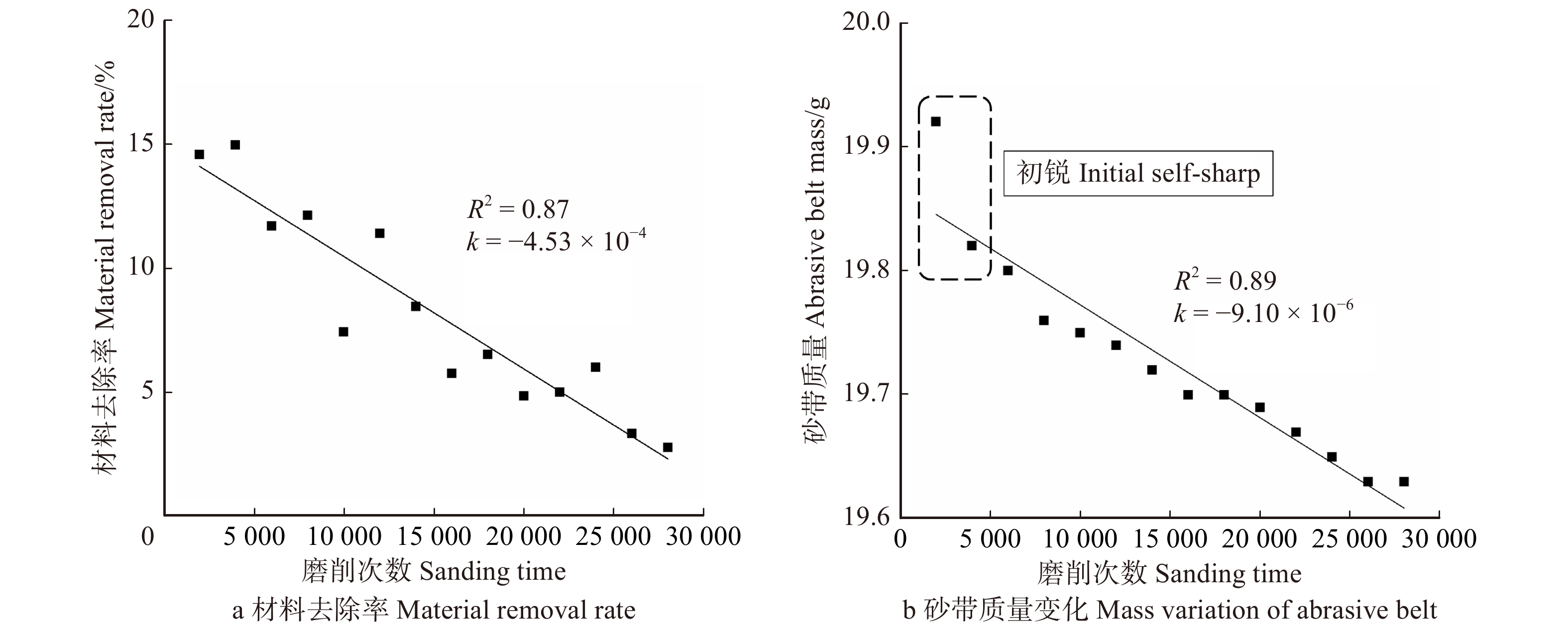
图 3 横纹磨削时砂带表面三维形貌图Figure 3. Three-dimensional topography of the belt surface during transverse sanding随着磨削次数增加,高度较高的磨粒由于与试件接触深度加大,会首先发生破碎和脱落,这也就是所谓的“初锐阶段”[11]。从图4b中明显看出,初锐阶段大约在磨削次数5 000次以内,砂带上的磨粒在发生脱落和破碎之外,磨粒的磨损也同时发生,此时砂带质量减小的速率较快,只是砂带脱落、破碎和磨损所占比例不同。图5b中对应的砂带表面粗糙度较高,即磨粒高度之间差异较大,所以更易压入试件进行磨削,故此时对应图4a处材料去除率极高。
![]() 图 4 横纹磨削水曲柳时材料去除率与砂带质量变化Figure 4. Material removal rate of workpiece and the mass variation of abrasive belt during transverse sanding
图 4 横纹磨削水曲柳时材料去除率与砂带质量变化Figure 4. Material removal rate of workpiece and the mass variation of abrasive belt during transverse sanding![]() 图 5 横纹磨削时试件、砂带的表面粗糙度Figure 5. Surface roughness of workpiece and abrasive belt during transverse sanding
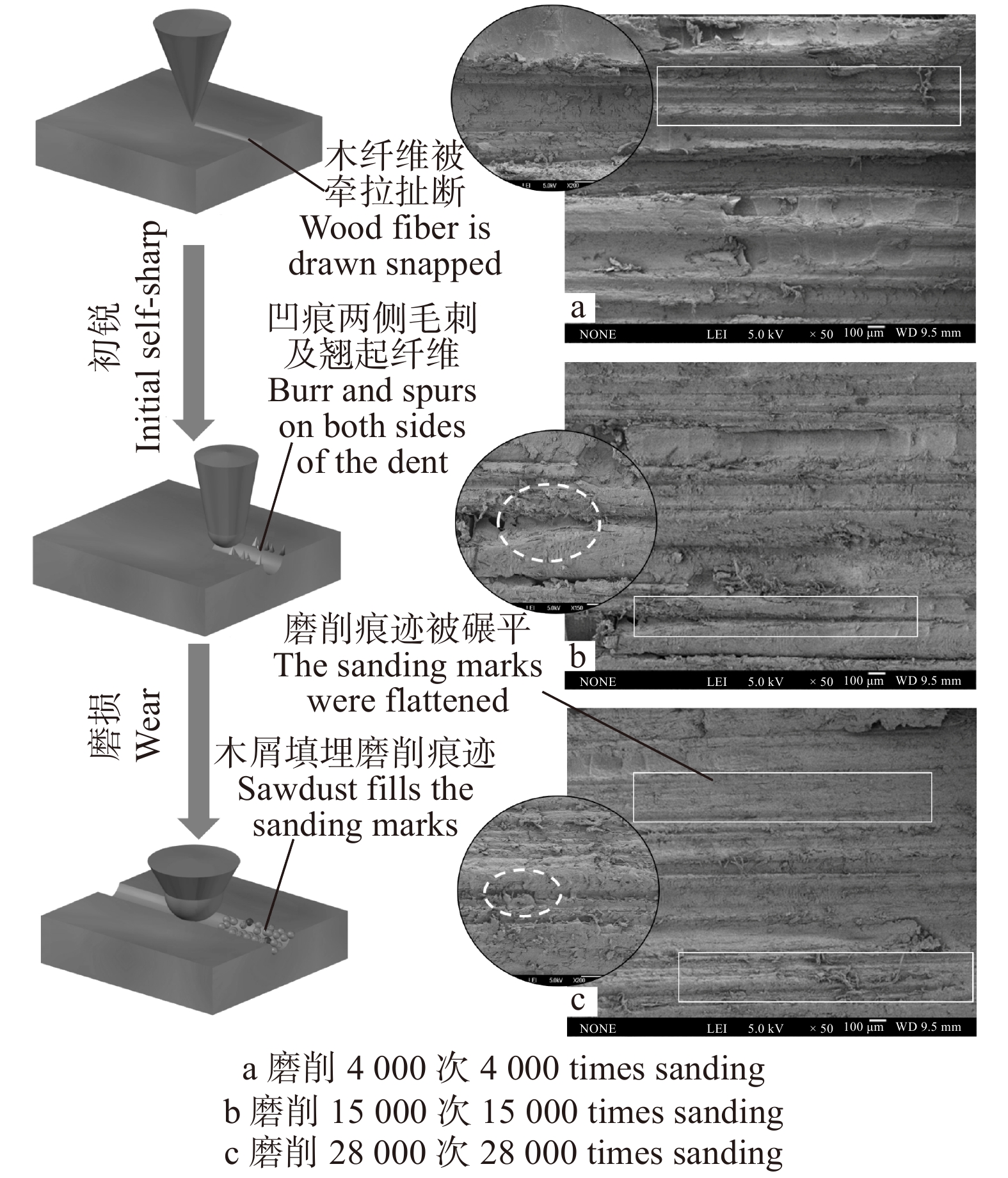
图 5 横纹磨削时试件、砂带的表面粗糙度Figure 5. Surface roughness of workpiece and abrasive belt during transverse sanding初锐阶段结束后,在图3b中可观察到砂带表面相同高度磨粒的个数明显增加,主要由于试件与磨粒之间的相互作用使得磨粒尖端钝化为小平面,其等高性变好且高度变矮,材料去除率会随之下降。此时磨粒协同干涉作用逐渐显著,从图2b中可以看出在一条较宽的磨削痕迹中有许多条细小的磨削痕迹,但是并没有非常明显且完整的单个磨粒的磨削痕迹,这是由于前一个磨粒所产生的磨削痕迹会被后续的磨粒进行进一步“加工”,而最终体现出来的磨削痕迹是众多磨粒共同作用的效果,消除了之前单个磨粒的磨削痕迹,使磨削痕迹的界限变得模糊,这种多磨粒干涉效应[12]也是使工件表面变光滑的最关键因素。
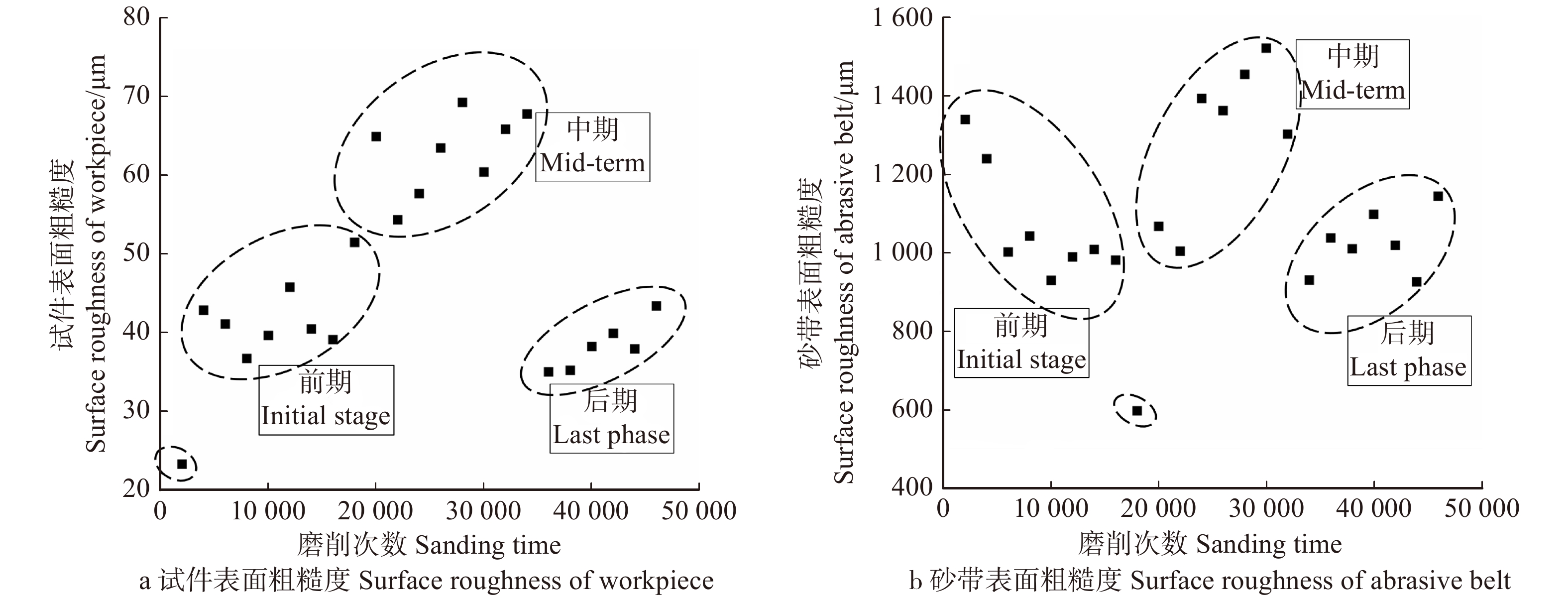
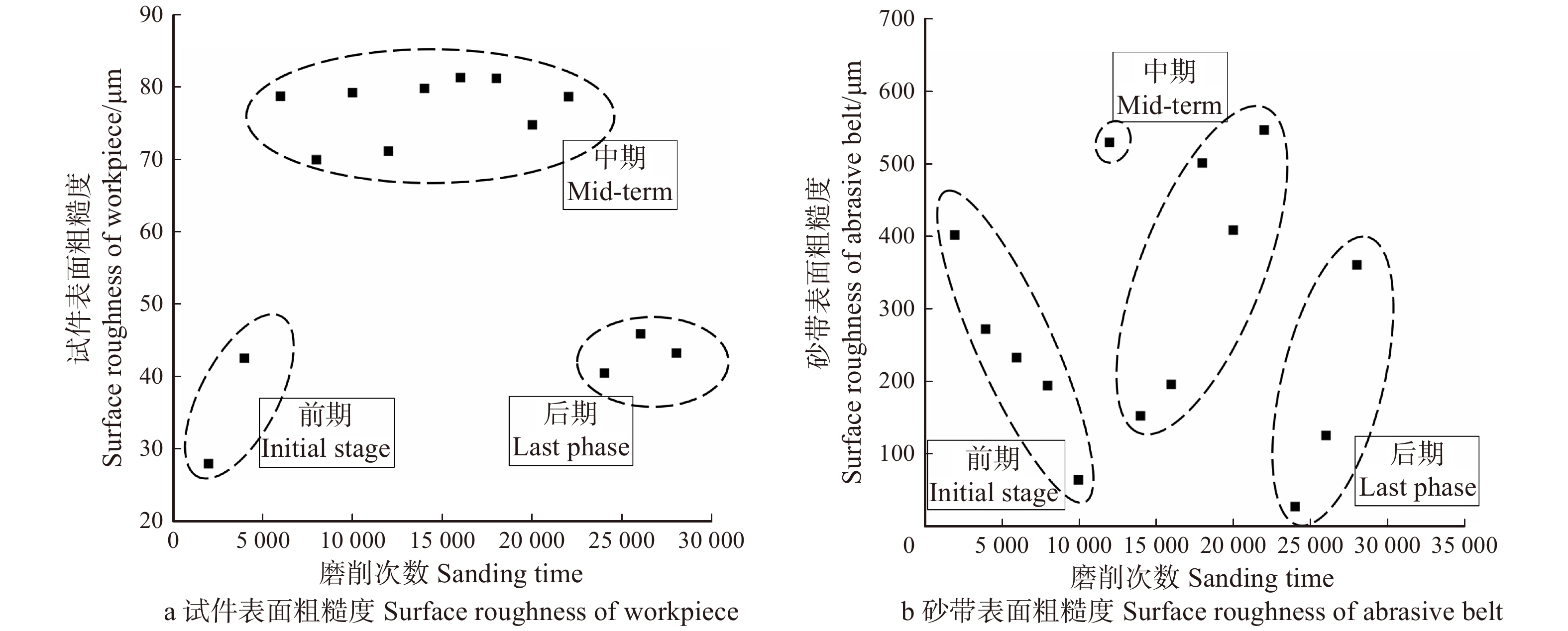
随着磨削阶段到达中、后期,从图2c中可以看出磨粒将木纤维从两侧挤出翘起,在试件表面犁出凹痕,只能切除少量材料,故材料去除率呈下降趋势并在一定程度上影响了试件表面的粗糙度。从图5可以看出,磨削中期试件表面的粗糙度是比较高的,但磨削后期由于磨粒钝化更加明显且高度更低,如图3c,砂带表面等高性进一步增加,表面的粗糙度迅速降低,磨粒与试件表面接触深度变浅,试件表面发生弹塑性变形,导致几乎无法去除材料,表面凹痕更浅,故可见图5a在中、后期表面粗糙度明显下降。观察图4b可得磨削中期到后期砂带质量基本是线性减小的,也就说明砂带的磨损、脱落、破碎最终达到一个较为平衡的阶段。
2.2 顺纹磨削
顺纹磨削时,切削方向与木材纤维方向基本平行,磨粒是从木材纤维的端向切入木材[13]。与横纹磨削类似,在初期阶段磨粒顶端的曲率半径较小,切削刃较为锋利,如图6a,砂带上磨粒的等高性较差,易压入木材做有效切削,从图7a中可观察到初期阶段试件表面有多条清晰且完整的磨粒磨削痕迹。
![]() 图 6 顺纹磨削时的砂带表面三维形貌图Figure 6. Three-dimensional topography of the belt surface during longitudinal sanding
图 6 顺纹磨削时的砂带表面三维形貌图Figure 6. Three-dimensional topography of the belt surface during longitudinal sanding![]() 图 7 顺纹磨削时水曲柳表面形貌和磨削过程示意图Figure 7. Surface topography and the schematic diagram for workpiece during longitudinal sanding process
图 7 顺纹磨削时水曲柳表面形貌和磨削过程示意图Figure 7. Surface topography and the schematic diagram for workpiece during longitudinal sanding process对比图8a与图4a发现,水曲柳横纹磨削时材料去除率整体大于顺纹磨削的材料去除率,且顺纹磨削时材料去除率的下降速率大于横纹的下降速率。这说明砂带在顺纹磨削时更难去除材料,主要由于顺纹磨削过程中,磨粒切入木材时是从木纤维的端头切入,加之磨粒一般为负前角,很难依靠锋利的刃口将木纤维切断,更多的是将其牵扯拉断[14]。从图8b中也可发现此时砂带的磨损速率更快,导致砂带使用寿命也更短。
![]() 图 8 顺纹磨削水曲柳时材料去除率、砂带质量变化Figure 8. Material removal rate of workpiece and the mass variation of abrasive belt during longitudinal sanding
图 8 顺纹磨削水曲柳时材料去除率、砂带质量变化Figure 8. Material removal rate of workpiece and the mass variation of abrasive belt during longitudinal sanding随磨削次数增加,砂带磨粒高度降低,顶端刃口曲率半径不断增大,在图7b中可以看到许多较宽的磨削痕迹且在凹痕两侧有较多毛刺及翘起的木纤维,故在图9a中可明显观察到试件表面粗糙度增高。在磨削中期,从图6b可看出,砂带上磨粒脱落,破碎所占比例较大,等高性增大,磨粒与试件的接触深度降低且切割纤维能力减弱,在试件表面发生更多的是耕犁现象,木纤维在多次牵拉下被扯断,如图7b左侧示意图。
![]() 图 9 顺纹磨削时试件、砂带的表面粗糙度Figure 9. Surface roughness of workpiece and abrasive belts during longitudinal sanding
图 9 顺纹磨削时试件、砂带的表面粗糙度Figure 9. Surface roughness of workpiece and abrasive belts during longitudinal sanding至磨削后期,观察图7c发现部分磨削痕迹被木屑填埋,翘起的毛刺被抹平,且凹痕大多较浅,故可见图9a中试件表面粗糙度明显下降。如图6c,此时磨粒整体高度下降,磨粒磨损情况严重,大多仅摩擦试件表面,几乎无法切除材料,同时将折断、翘起的木纤维碾平并填充空隙,使得表面粗糙度降低。但是不难从图9b中发现,顺纹磨削时后期砂带的表面粗糙度更大,这也和切削方式和磨粒破碎形式有关,因为在顺纹切削时纤维大多被拉断,所以磨粒更易受拉力造成破碎和脱落,这也是砂带在顺纹和横纹磨削时的一个主要差异[15]。
2.3 材料去除率、表面粗糙度灰色预测模型
在砂带寿命范围内,累计磨削长度与磨削过程中材料去除率、表面粗糙度间有着密切联系,但由于样本量少[16-17],且存在如木材非均质、砂带植砂磨粒分布偶然性等较多未知因素,较难建立磨削长度与材料去除率、表面粗糙的回归模型。20世纪80年代邓聚龙教授创立了一种就数找数的方法,即灰色系统生成法。其研究对象是“部分信息已知,部分信息未知”的“小样本”、“贫信息”不确定性系统,用于解决“小样本”、“贫信息”的不确定性问题,这种方法容错率较高,也易于实现模糊控制[18]。因此本研究采用均值GM(1,1)预测模型建立磨削长度与材料去除率、表面粗糙度间的灰色预测模型,如图10所示。
2.3.1 模型预测
在本次模型中,系列特征序列为材料去除率和表面粗糙度,相关因素序列为磨削长度,模型原始数据及建模步骤如下:
第一步,根据表1、2中数据,建立初始材料去除率序列
X(0)1 、表面粗糙度序列X(0)2 。表 1 横纹磨削模型数据Table 1. Model data of transverse sanding序号
No.磨削长度
Sanding length/m表面粗糙度
Surface roughness/μm材料去除率
Material removal rate/%1 200 41.178 15.11 2 400 39.723 13.48 3 600 40.546 15.71 4 800 51.551 7.27 5 1 000 54.409 12.66 6 1 200 63.536 11.39 7 1 400 60.472 8.35 8 1 600 67.846 9.61 注:为提高所建灰色模型的预测精度,选取数据均在砂带最佳使用寿命范围内。下同。Notes: to improve the precision of the established gray model, the selected data is located in the range of optimal abrasive belt life. Same as below. 表 2 顺纹磨削模型数据Table 2. Model data of longitudinal sanding序号
No.磨削长度
Sanding length/m表面粗糙度
Surface roughness /μm材料去除率
Material removal rate/%1 100 42.572 14.57 2 200 78.737 14.96 3 300 69.976 11.70 4 400 79.229 12.13 5 500 71.151 7.43 6 600 79.812 11.40 7 700 81.305 8.46 8 800 81.221 5.77 9 900 74.79 6.53 10 1 000 78.662 4.86 X(0)1=(X(0)1(1),X(0)1(2),X(0)1(3),⋯,X(0)1(n−1),X(0)1(n))X(0)2=(X(0)2(1),X(0)2(2),X(0)2(3),⋯,X(0)2(n−1),X(0)2(n)) 式中:
X(0)1(n) 代表材料去除率第n个数据的原始数值;X(0)2(n) 代表表面粗糙度第n个数据的原始数值;n表示原始数列的个数,本文横纹磨削时n = 8,顺纹磨削时n = 10。第二步,均值GM(1,1)建模。
(1)对原始数据做一次累加得
X(1)1 和X(1)2 ,原始序列的1-AGO生成;(2)
X(1)(n) 表示原始序列第n个数的一阶累加数值,X(0)(n) 表示原始序列第n个数值,构造数据矩阵B和数据向量Y;B=[−12(X(1)(1)+X(1)(2)1−12(X(1)(2)+X(1)(3)1⋮⋮−12(X(1)(n−1)+X(1)(n)1],Y=[X(0)(2)X(0)(3)⋮X(0)(n)] GM(1,1)模型可表示为
Y=Bu ,即[X(0)(2)X(0)(3)⋮X(0)(n)]=[−12(X(1)(1)+X(1)(2)1−12(X(1)(2)+X(1)(3)1⋮⋮−12(X(1)(n−1)+X(1)(n)1][ˆaˆb] (3)计算待估向量
ˆu ;ˆu=(ˆa,ˆb)T=(BT⋅B)−1BTY 式中:a为发展灰数,表示序列的发展趋势;b为内生控制灰数,表示数据间的变化关系。
(4)建立模型。
dX(1)1dt+a1X(1)=b1 dX(1)2dt+a2X(1)=b2 式中:t表示序号1,2,······,n−1,n的连续变量;a1、b1分别为计算材料去除率预估向量的发展灰数和内生控制灰数;a2、b2分别为计算表面粗糙度预估向量的发展灰数和内生控制灰数。
第三步,模型检验。
从图11a中数据计算得到:横纹磨削过程中,材料去除率平均模拟相对误差为18.362%,表面粗糙度的平均模拟相对误差为5.170%。从图11b中数据计算可得:顺纹磨削过程中,材料去除率平均模拟相对误差为13.491%,表面粗糙度的平均模拟相对误差为4.603%。
2.3.2 结果分析
由图11可知:采用灰色模型均值GM(1,1)的预测平均模拟相对误差都是在20%以内。考虑预测系统本身局限性,以及木质材料的各向异性导致原始数据的随机性较高,本身规律性不是很明显的情况下,这些因素都加大了预测难度,一定程度上影响了模拟预测的准确性。故本模型适用于预测水曲柳磨削过程中,磨削长度分别与材料去除率、表面粗糙度之间关系的预测,对实际生产具有借鉴意义。
3. 结 论
木材磨削过程中材料去除行为会影响木材加工表面质量,其影响因素主要有磨削方向、材料种类以及磨削过程中砂带的磨损情况。本文主要研究了水曲柳在不同磨削方向时的材料去除率和试件、砂带表面粗糙度变化情况,探究了磨粒磨损过程及其与材料去除率之间的关系,得出以下结论:
(1)磨削过程中磨削方向对砂带磨损的影响不同,顺纹磨削对砂带上磨粒的磨损大于横纹磨削。磨粒的磨损对材料去除率有较大的影响,磨粒磨损程度越大,材料去除率越小。当材料去除率降低至3%时,可认为达到砂带使用寿命,应及时更换砂带。
(2)磨削过程中,砂带上磨粒的等高性对材料去除率也有较大影响。等高性越差,材料去除率越高。随磨削次数增加,砂带材料去除能力不断下降,试件表面粗糙度则呈现先增大后减小的趋势。
(3)采用灰色模型均值GM(1,1)的预测平均模拟相对误差都是在20%以内。适用于水曲柳磨削过程中磨削长度分别与材料去除率、表面粗糙度之间关系的预测。
-
![]()
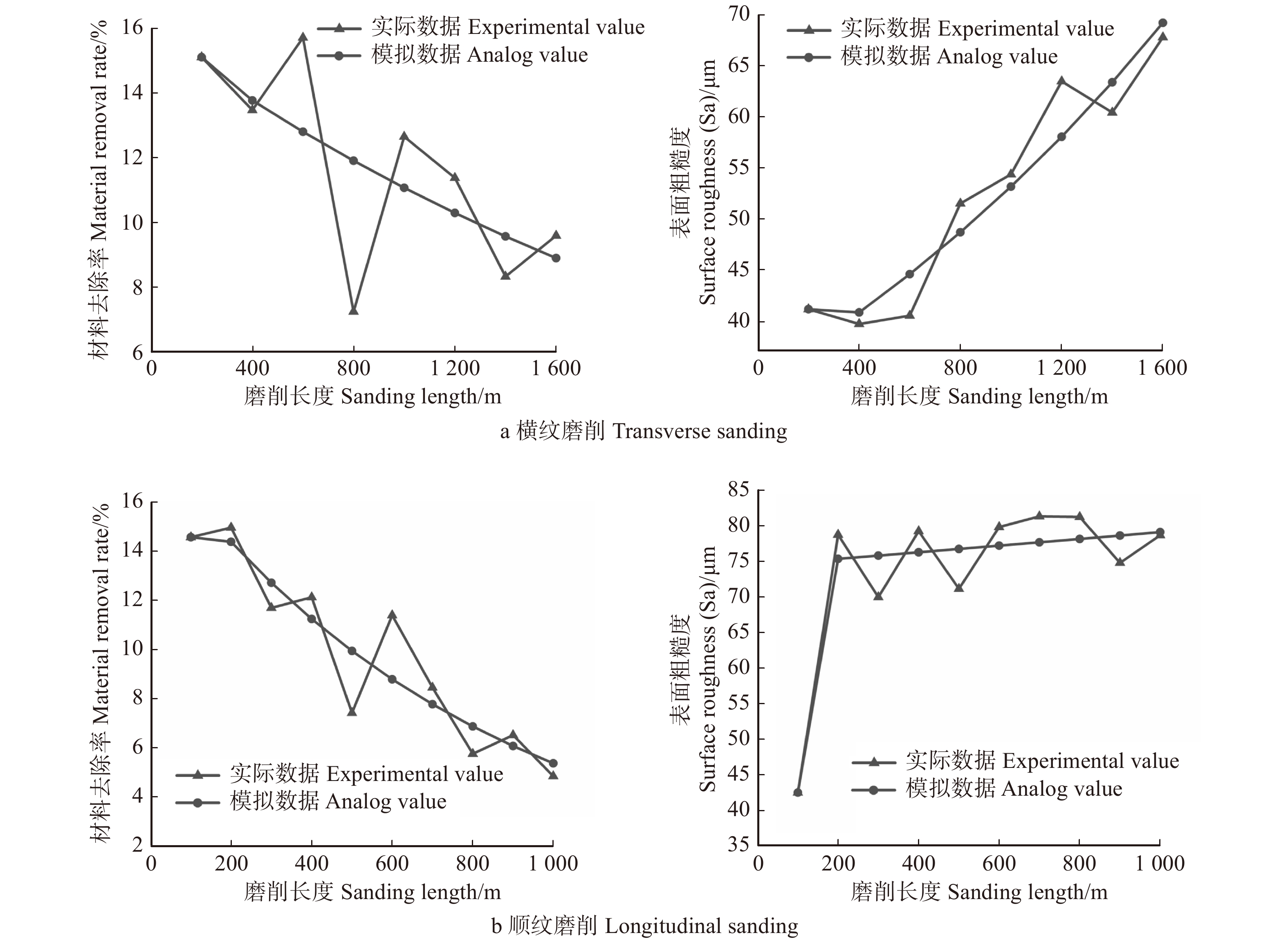
图 1 1年生枝条水平不同处理方式下簇毛槭1年生枝条长度对比
实心点为雌性,空心点为雄性。 图中不同字母表示在P < 0.05水平上差异显著。下同。Solid point: female; hollow point: male. Varied letters denote significant differences at P < 0.05 level. Same as below.
Figure 1. Comparing the length of 1 year old new shoot under different treatments at shoot level for Acer barbinerve
![]()
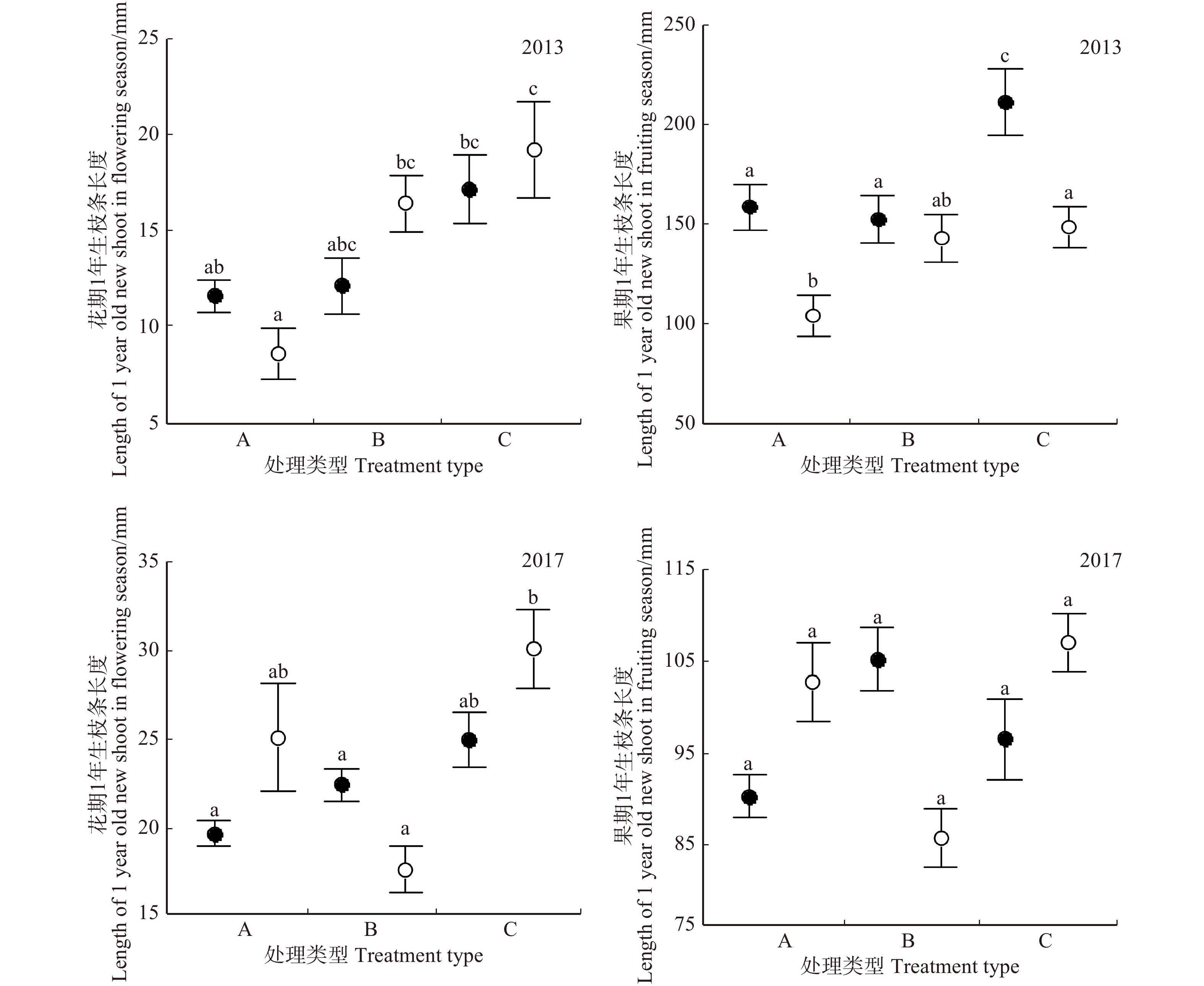
图 2 1年生枝条水平不同处理方式下簇毛槭叶片生物量对比
Figure 2. Comparing foliage biomass under different treatments at 1 year old new shoot level for Acer barbinerve
![]()
图 3 2级侧枝水平不同处理方式下簇毛槭1年生枝条长度对比
实心点为雌性,空心点为雄性. 图中不同字母表示在P < 0.05水平上差异。Solid point: female; hollow point: male. Varied letters denote significant differences at P < 0.05 level.
Figure 3. Comparing the length of 1 year old new shoot under different treatments at grade 2 lateral branch level for Acer barbinerve
![]()
图 4 2级侧枝水平不同处理方式下簇毛槭叶片生物量对比
Figure 4. Comparing foliage biomass under different treatments at grade 2 lateral branch level for Acer barbinerve
表 1 试验选取的簇毛槭样株情况
Table 1 Sample individual conditions of Acer barbinerve
年份
Year株数
Tree number胸径
DBH/mm树高
Tree height/m冠幅东西
Crown diameter of east-west/m冠幅南北
Crown diameter of north-south/m雌株
Female雄株
Male雌株
Female雄株
Male雌株
Female雄株
Male雌株
Female雄株
Male2013 180 20.367a 19.176a 2.816a 2.543a 2.233a 2.051a 2.098a 2.085a 2017 145 21.511a 20.282a 2.720a 2.444a 2.553a 2.114b 2.224a 1.920a 注:不同字母表示在P < 0.05水平上差异显著。Note: different letters indicate significant difference at P < 0.05 level.  下载: 导出CSV
下载: 导出CSV
表 2 1年生枝条水平簇毛槭1年生枝条长度影响要素分析
Table 2 Analysis of influencing factors of the length of 1 year old new shoot at 1 year old shoot level for Acer barbinerve
变异来源 Variance source 2013年花期
Flowering season in 20132013年果期
Fruiting season in 20132017年花期
Flowering season in 20172017年果期
Fruiting season in 2017F P F P F P F P 性别 Gender 0.01 0.908 35.81 0.001 1.84 0.175 15.36 0.001 处理 Treatment 122.68 0.001 0.75 0.475 9.70 0.001 4.25 0.014 性别 × 处理 Gender × treatment 12.93 0.001 7.62 0.001 17.33 0.001 6.26 0.002
下载: 导出CSV
表 3 1年生枝条水平簇毛槭叶片生物量影响要素分析
Table 3 Influencing factors of biomass of foliage at 1 year old new shoot level for Acer barbinerve
变异来源 Variation source 2013年花期
Flowering season in 20132013年果期
Fruiting season in 20132017年花期
Flowering season in 20172017年果期
Fruiting season in 2017F P F P F P F P 性别 Gender 58.31 0.001 114.9 0.001 6.55 0.011 221.91 0.001 处理 Treatment 22.82 0.001 107.85 0.001 218.36 0.001 140.45 0.001 性别 × 处理 Gender × treatment 96.50 0.001 11.54 0.001 88.01 0.001 60.53 0.001
下载: 导出CSV
表 4 2级侧枝水平簇毛槭1年生枝条长度影响要素分析
Table 4 Influencing factors of the length of 1 year old new shoot at grade 2 lateral branch level for Acer barbinerve
变异来源 Variation source 2013年花期
Flowering season in 20132013年果期
Fruiting season in 20132017年花期
Flowering season in 20172017年果期
Fruiting season in 2017F P F P F P F P 性别 Gender 0.81 0.369 25.01 0.001 2.85 0.092 0.20 0.656 处理 Treatment 13.07 0.001 11.31 0.001 6.40 0.002 0.68 0.507 性别 × 处理 Gender × treatment 2.85 0.058 3.35 0.036 4.39 0.013 5.04 0.007
下载: 导出CSV
表 5 2级侧枝水平簇毛槭叶片生物量影响要素分析
Table 5 Influencing factors of the biomass of foliage at grade 2 lateral branch level for Acer barbinerve
变异来源 Variation source 2013年花期
Flowering season in 20132013年果期
Fruiting season in 20132017年花期
Flowering season in 20172017年果期
Fruiting season in 2017F P F P F P F P 性别 Gender 15.50 0.001 11.85 0.001 22.31 0.001 3.35 0.068 处理 Treatment 19.14 0.001 1.94 0.145 17.95 0.001 16.60 0.001 性别 × 处理 Gender × treatment 23.27 0.001 1.88 0.153 21.93 0.001 7.86 0.001
下载: 导出CSV
-
[1] Karlsson P S, Andersson M, Svensson B M. Relationships between fruit production and branching in monocarpic shoot modules of Rhododendron lapponicum[J]. Ecoscience, 2006, 13(3): 396−403. doi: 10.2980/i1195-6860-13-3-396.1
[2] Berger J D, Shrestha D, Ludwig C. Reproductive strategies in Mediterranean Legumes: trade-offs between phenology, seed size and vigor within and between wild and domesticated Lupinus Species collected along aridity gradients [J/OL]. Frontiers in Plant Science, 2017, (2017−04−13)[2018−01−20]. https://doi.org/10.3389/fpls.2017.00548.
[3] Obeso J R. Costs of reproduction in Ilex aquifolium: effects at tree, branch and leaf levels[J]. Journal of Ecology, 1997, 85(2): 159−166. doi: 10.2307/2960648
[4] Nicotra A B. Reproductive allocation and the long-term costs of reproduction in Siparuna grandiflora, a dioecious neo-tropical shrub[J]. Journal of Ecology, 1999, 87(1): 138−149. doi: 10.1046/j.1365-2745.1999.00337.x
[5] Miyazaki Y. Dynamics of internal carbon resources during masting behavior in trees[J]. Ecological Research, 2013, 28(2): 143−150. doi: 10.1007/s11284-011-0892-6
[6] Thorén L M, Karlsson P S, Tuomi J. Somatic cost of reproduction in three carnivorous Pinguicula species[J]. Oikos, 1996, 76(3): 427−434. doi: 10.2307/3546336
[7] Newell E A. Direct and delayed costs of reproduction in Aesculus californica[J]. Journal of Ecology, 1991, 79(2): 365−378. doi: 10.2307/2260719
[8] Ehrlén J, Van Groenendael J. Storage and the delayed costs of reproduction in the understorey perennial Lathyrus vernus[J]. Journal of Ecology, 2001, 89(2): 237−246. doi: 10.1046/j.1365-2745.2001.00546.x
[9] Horvitz C C, Schemske D W. Demographic cost of reproduction in a neotropical herb: an experimental field study[J]. Ecology, 1988, 69(6): 1741−1745. doi: 10.2307/1941152
[10] Horibata S, Hasegawa S F, Kudo G. Cost of reproduction in a spring ephemeral species, Adonis ramosa (Ranunculaceae): carbon budget for seed production[J]. Annals of Botany, 2007, 100(3): 565−571. doi: 10.1093/aob/mcm131
[11] Matsuyama S, Sakimoto M. Sexual dimorphism of reproductive allocation at shoot and tree levels in Zanthoxylum ailanthoides, a pioneer dioecious tree[J]. Botanique, 2010, 88(10): 867−874. doi: 10.1139/B10-058
[12] Cipollini M L, Whigham D F. Sexual dimorphism and cost of reproduction in the dioecious shrub Lindera benzoin (Lauraceae)[J]. American Journal of Botany, 1994, 81(1): 65−75. doi: 10.1002/j.1537-2197.1994.tb15410.x
[13] Bañuelos M J, Obeso J R. Resource allocation in the dioecious shrub Rhamnus alpinus: the hidden costs of reproduction[J]. Evolutionary Ecology Research, 2004, 6(3): 397−413.
[14] Mge M, Antos J A, Allen G A. Modules of reproduction in females of the dioecious shrub Oemleria cerasiformis[J]. Revue Canadienne De Botanique, 2004, 82(3): 393−400.
[15] Matsuyama S, Sakimoto M. Allocation to reproduction and relative reproductive costs in two species of dioecious Anacardiaceae with contrasting phenology[J]. Annals of Botany, 2008, 101(9): 1391−1400. doi: 10.1093/aob/mcn048
[16] Geber M A, Dawson T E, Delph L F. Gender and sexual dimorphism in flowering plants [M]. Berlin: Springer, 1999: 33−60.
[17] Delph L F, Knapczyk F N, Taylor D R. Among-population variation and correlations in sexually dimorphic traits of Silene latifolia[J]. Journal of Evolutionary Biology, 2010, 15(6): 1011−1020.
[18] Chen J, Niu Y, Yang Y, et al. Sexual allocation in the gynodioecious species Cyananthus macrocalyx (Campanulaceae) at high elevations in the Sino-Himalaya Mountains[J]. Alpine Botany, 2016, 126(1): 49−57. doi: 10.1007/s00035-015-0154-2
[19] Huan H, Wang B, Liu G, et al. Sexual differences in morphology and aboveground biomass allocation in relation to branch number in Morus alba saplings[J]. Australian Journal of Botany, 2016, 64(3): 269−275. doi: 10.1071/BT15189
[20] Fonseca D C, Oliveira M L R, Pereira I M, et al. Phenological strategies of dioecious species in response to the environmental variations of rupestrian grasslands[J]. Cerne, 2018, 23(4): 517−527.
[21] Juvany M, Munnébosch S. Sex-related differences in stress tolerance in dioecious plants: a critical appraisal in a physiological context[J]. Journal of Experimental Botany, 2015, 66(20): 6083−6092. doi: 10.1093/jxb/erv343
[22] Chen J, Duan B, Wang M, et al. Intra-and inter-sexual competition of Populus cathayana under different watering regimes[J]. Functional Ecology, 2014, 28(1): 124−136. doi: 10.1111/fec.2014.28.issue-1
[23] Wang J, Zhang C, Zhao X, et al. Reproductive allocation of two dioecious Rhamnus species in temperate forests of northeast China[J]. Iforest Biogeosciences & Forestry, 2013, 7(1): 25−32.
[24] Zang R G, Xu H C. Canopy disturbance regimes and gap regeneration in a Korena pine-broadleaved forest in Jiaohe, northeast China[J]. Bulletin of Botanical Research, 1999, 19(2): 112−120.
[25] 中国科学院中国植物志委员会. 中国植物志: 1卷[M]. 北京: 科学出版社, 2004. The Chinese Flora of the Chinese Academy of Sciences. Flora of China: I [M]. Beijing: Science Press, 2004.
[26] Zajitschek F, Connallon T. Partitioning of resources: the evolutionary genetics of sexual conflict over resource acquisition and allocation[J]. Journal of Evolutionary Biology, 2017, 30(4): 826−838. doi: 10.1111/jeb.2017.30.issue-4
[27] Tonnabel J, David P, Pannell J R. Sex-specific strategies of resource allocation in response to competition for light in a dioecious plant[J]. Oecologia, 2017, 185(4): 675−686. doi: 10.1007/s00442-017-3966-5
[28] Sletvold N J Å. Climate-dependent costs of reproduction: survival and fecundity costs decline with length of the growing season and summer temperature[J]. Ecology Letters, 2015, 18(4): 357−364. doi: 10.1111/ele.2015.18.issue-4
[29] Hossain S M Y, Caspersen J P, Thomas S C. Reproductive costs in Acer saccharum: exploring size-dependent relations between seed production and branch extension[J]. Trees, 2017, 31(4): 1−10.
[30] Xiao Y A, Dong M, Wang N, et al. Effects of organ removal on trade-offs between sexual and clonal reproduction in the stoloniferous herb Duchesnea indica[J]. Plant Species Biology, 2016, 31(1): 50−54. doi: 10.1111/psbi.2016.31.issue-1
[31] Torimaru T, Tomaru N. Reproductive investment at stem and genet levels in male and female plants of the clonal dioecious shrub Ilex leucoclada (Aquifoliaceae)[J]. Botanique, 2012, 90(4): 301−310. doi: 10.1139/b2012-004
[32] Teitel Z, Pickup M, Field D L, et al. The dynamics of resource allocation and costs of reproduction in a sexually dimorphic, wind-pollinated dioecious plant[J]. Plant Biology, 2016, 18(1): 98−103. doi: 10.1111/plb.12336
[33] Leigh A, Cosgrove M, Nicotra A. Reproductive allocation in a gender dimorphic shrub: anomalous female investment in Gynatrix pulchella[J]. Journal of Ecology, 2006, 94(6): 1261−1271. doi: 10.1111/jec.2006.94.issue-6
[34] Hasegawa S, Takeda H. Functional specialization of current shoots as a reproductive strategy in Japanese alder (Alnus hirsuta var.sibirica)[J]. Canadian Journal of Botany, 2001, 79(1): 38−48.
[35] Díaz-Barradas M C, Zunzunegui M, Collantes M, et al. Gender-related traits in the dioecious shrub Empetrum rubrum in two plant communities in the Magellanic steppe[J]. Acta Oecologica, 2014, 60(10): 40−48.
[36] Xiao Y, Zhao H, Yang W, et al. Variations in growth, clonal and sexual reproduction of Spartina alterniflora responding to changes in clonal integration and sand burial[J]. CLEAN − Soil, Air, Water, 2015, 43(7): 1100−1106. doi: 10.1002/clen.v43.7
[37] Wang H, Matsushita M, Tomaru N, et al. Differences in female reproductive success between female and hermaphrodite individuals in the subdioecious shrub Eurya japonica (Theaceae)[J]. Plant Biology, 2015, 17(1): 194−200. doi: 10.1111/plb.12189
[38] Midgley J J. Causes of secondary sexual differences in plants : evidence from extreme leaf dimorphism in Leucadendron (Proteaceae)[J]. South African Journal of Botany, 2010, 76(3): 588−592. doi: 10.1016/j.sajb.2010.05.001
[39] Jaquish L L, Ewers F W. Seasonal conductivity and embolism in the roots and stems of two clonal ring: porous trees, Sassafras albidum (Lauraceae) and Rhus typhina (Anacardiaceae)[J]. American Journal of Botany, 2001, 88(2): 206−212. doi: 10.2307/2657011
[40] Zunzunegui M, Barradas M C D, Clavijo A, et al. Ecophysiology, growth timing and reproductive effort of three sexual foms of Corema album (Empetraceae)[J]. Plant Ecology, 2006, 183(1): 35−46. doi: 10.1007/s11258-005-9004-4
[41] Milla R, Castrodíez P, Maestromartínez M, et al. Costs of reproduction as related to the timing of phenological phases in the dioecious shrub Pistacia lentiscus L.[J]. Plant Biology, 2006, 8(1): 103−111. doi: 10.1055/s-2005-872890
[42] Nicotra A B. Sexually dimorphic growth in the dioecious tropical shrub, Siparuna grandiflora[J]. Functional Ecology, 2010, 13(3): 322−331.
[43] Sánchez V J, Pannell J R. Sexual dimorphism in resource acquisition and deployment: both size and timing matter[J]. Annals of Botany, 2011, 107(1): 119−126. doi: 10.1093/aob/mcq209
[44] Tozawa M, Ueno N, Seiwa K. Compensatory mechanisms for reproductive costs in the dioecious tree Salix integra[J]. Botany-botanique, 2009, 87(3): 315−323. doi: 10.1139/B08-125
[45] Álvarez-Cansino L, Zunzunegui M, Barradas M C D, et al. Gender-specific costs of reproduction on vegetative growth and physiological performance in the dioecious shrub Corema album[J]. Annals of Botany, 2010, 106(6):989−998.
[46] Fujii S, Toriyama K. Suppressed expression of retrogrand - regulated male sterility restores pollen fertility in cytoplasmic male sterile rice plants[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(23): 9513−9518. doi: 10.1073/pnas.0901860106
[47] Kumar N, Gupta S, Tripathi A N. Gender-specific responses of Piper betle L. to low temperature stress: changes in chlorophyllase activity[J]. Biologia Plantarum, 2006, 50(4): 705−708. doi: 10.1007/s10535-006-0111-4
[48] Aschan G, Pfanz H. Non-foliar photosynthesis: a strategy of additional carbon acquisition[J]. Flora, 2003, 198(2): 81−97. doi: 10.1078/0367-2530-00080
[49] Cai Z Q. Shade delayed flowering and decreased photosynthesis, growth and yield of Sacha Inchi (Plukenetia volubilis) plants[J]. Industrial Crops & Products, 2011, 34(1): 1235−1237.
-
期刊类型引用(5)
1. 罗斌,杜瑶,柳浩雨,王钦悦,李春瑜,李黎,刘红光. 定组态砂带粒度对刨花板磨削的影响. 北京林业大学学报. 2024(09): 141-150 .  本站查看
本站查看
2. 施晨阳,赖文峰,文国卫,蒋天雨,朱晓如,吕增伟,张国防. 基于Maxent模型预测水曲柳的潜在适生区. 西北林学院学报. 2022(02): 149-156 . 百度学术
3. 罗斌,张健,柳浩雨,李黎,刘红光,李昊. 木质材料磨削理论及技术研究进展. 木材科学与技术. 2022(04): 6-12 . 百度学术
4. 杜瑶,田彪,张健,刘红光,罗斌. 中密度纤维板磨削时的砂带磨损研究及寿命评判. 北京林业大学学报. 2021(10): 126-134 . 本站查看
5. 卢冬冬,杨铁牛,杨保健,叶榕伟,阳亚,杜华娜. 肉桂打磨方法优化. 农业工程. 2020(01): 73-78 . 百度学术
其他类型引用(5)


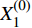
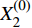
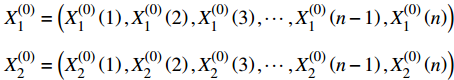


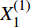
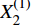
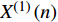

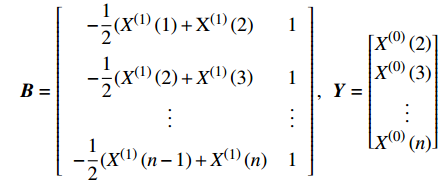
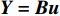
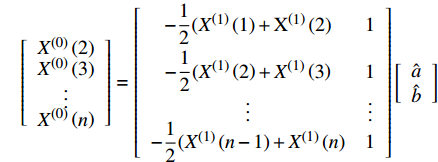
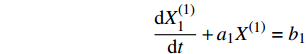
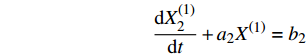


计量
- 文章访问数: 2486
- HTML全文浏览量: 890
- PDF下载量: 71
- 被引次数: 10
