
Karyotype analysis and meiotic chromosome behavior observation on Paeonia × lemoinei ‘High Noon’
-
摘要:目的芍药属牡丹组革质花盘亚组与肉质花盘亚组间的远缘杂交是现代牡丹育种的重要方向之一。然而,亚组间杂种普遍高度不育,很难继续用于杂交育种。‘正午’牡丹是一个观赏性好、适应性强的亚组间杂交品种,虽然其通常高度不育,但仍被用作亲本培育出一些优异的杂交后代,表现出一定的育性。研究其减数分裂的染色体行为,可为揭示其极低育性的形成机制提供重要的信息。方法本研究以‘正午’幼嫩花蕾中的雌蕊为材料,进行体细胞染色体核型分析;以花药为材料进行花粉母细胞减数分裂观察。结果‘正午’牡丹为二倍体,核型公式为2n = 2x = 10 = 7m + 1sm + 2st(1SAT)。其花粉母细胞可通过正常的减数分裂形成四分体及小孢子,减数第一次分裂中约有70%的花粉母细胞发生染色体行为异常,包括单价体及多价体、染色体桥、断片、落后染色体、不等分裂等,其中染色体桥出现频率最高;减数第二次分裂中,染色体行为异常率同样高达70%,常见的异常类型包括纺锤体定位异常、不同步分裂、染色体桥、断片、落后染色体等,最后形成二分体、三分体、微核或特小的额外小孢子。结论‘正午’减数分裂存在大量异常可能与其高度杂合的核型有关。普遍存在的单价体及多价体引起的不等分裂和染色体桥及断片造成的染色体片段缺失等染色体行为异常可能是导致‘正午’牡丹高度不育的重要原因。同时,仍有一小部分花粉母细胞能够顺利完成减数分裂,形成小孢子。其中一部分通过二分体或三分体形成未减数小孢子,表明其具有用于培育多倍体牡丹的潜力。Abstract:ObjectiveThe inter-subsectional distant hybridization between subsect. Delacayanae and subsect. Vaginatae of Paeonia sect. Moutan is a significant direction in modern tree peony breeding. Nevertheless, the inter-subsectional hybrids are generally sterile, therefore hard to be used as breeding parents subsequently. Paeonia × lemoinei ‘High Noon’ is an inter-subsectional cultivar with good ornamental and resistant traits. In spite of its sterility in most cases, ‘High Noon’ was still the parent of a few excellent hybrids, showing some degree of fertility. The research on the meiotic chromosome behavior of ‘High Noon’ could provide important messages to uncover the forming mechanisms of its low fertility.MethodWe employed the pistils from the young flower buds to detect the somatic karyotype, and the anthers to observe the meiotic chromosome behaviors during microsporogenesis in this study.Result‘High Noon’ was a diploid, whose karyotype formula was 2n = 2x = 10 = 7m + 1sm + 2st (1SAT). The pollen mother cells (PMCs) of ‘High Noon’ could generate tetrads and microspores through regular meiosis. The meiotic abnormalities in the first meiotic division took place in about 70% of PMCs, including univalent, multivalent, chromosome bridge, chromosome segmentation, lagging chromosome, unequal chromosome segregation, etc., among which, the chromosome bridge emerged most frequently. In the second meiotic division, meiotic abnormalities occurred in 70% of PMCs as well, involving abnormal spindle orientation, chromosome segregation asynchronicity, chromosome bridge, chromosome segmentation, lagging chromosome, etc., resulting in the formation of the dyad, triad, micronucleus, and extra microspores.ConclusionThe vast number of abnormality in meiosis of ‘High Noon’ might attributed to its heterozygous karyotype. The abnormal meiotic chromosome behaviors including the widespread unequal chromosome segregation caused by univalent or multivalent, and the chromosome segment missing due to the chromosome bridge and chromosome segmentation, might be the leading causes for the high sterility of ‘High Noon’. Meanwhile, a fraction of PMCs could accomplish the meiosis process successfully, forming the microspores. Therein some unreduced microspores were produced via dyad or triad formation, indicating its potential for polyploid tree peony breeding.
-
Keywords:
- Paeonia × lemoinei ‘High Noon’ /
- inter-subsectional hybrids /
- meiosis /
- chromosome
-
2017年9月23日中共中央国务院批复了《北京城市总体规划(2016年—2035年)》,要求“优化城市功能和空间布局,坚持疏解整治促提升,坚决拆除违法建筑,加强对疏解腾退空间的利用和引导,腾笼换鸟、留白增绿”。提出“疏解北京非首都功能,改善生态环境,建设和谐宜居之都”“健全市域绿色空间体系,建设森林城市,让森林进入城市的目标”。森林城市建设要求重视生物多样性保护,满足居民休闲游憩需求,传播生态文化知识。从20世纪60年代中期开始,国外一些林学家,提出在市区和郊区发展城市森林,把森林引入城市[1]的理念。结合国情,我国学者也从多学科多角度对城市森林进行概念定义、实践探究。关注的重点也从城市森林林木本身扩大到城市森林协调城市生态环境、维持生态系统良性循环等方面。作为城市生态系统的重要组成部分, 城市森林在维护碳氧平衡、净化环境、调节气候、涵养水源和保护生物多样性等方面具有重要作用[2-4]。大力发展城市森林,使城市与森林和谐共存,已经成为新世纪世界生态城市的发展方向[5]。
《北京城市总体规划(2016年—2035年)》通过后,北京市将拆违腾退土地用于“留白增绿”,作为绿色空间为城市提供生态服务,增加绿色游憩场所。计划拆除违法建设4 000万m2以上,腾退占地3 974 hm2,其中用于“留白增绿”1 986 hm2[6]。本研究定义“留白增绿”背景下的城市森林概念为:在城市地域范围内拆违腾退土地上,结合地块尺度、周边用地类型、总体规划定位三大要素所建立的能够针对性发挥城市森林生态、生活、生产功能,符合城市森林景观风貌的林木群落及其环境所构成的森林生态系统。如何在“留白增绿”的背景下充分利用拆违腾退土地,建设与城市发展背景相契合的城市森林成为新的议题。
1. 基于“留白增绿”的城市森林功能类型
本研究对2 039个,共计1 951.77 hm2的“留白增绿”地块进行梳理统计,地块面积从数平方米到数十公顷不等;空间区位遍布北京各个市辖区;地块类型涵盖公园绿地、防护绿地、生产绿地、农林用地、其他非建设用地等;部分地块位于总体规划中市域绿色空间的城市公园环、郊野公园环、环首都森林湿地公园环、历史文化名城保护区、文化保护带、市级通风廊道等;周边用地类型各异,涵盖居住用地、商业服务设施用地、物流仓储用地等。
城市森林的类型因划分标准的不同而呈现多样化,本文以城市森林主要功能为标准对城市森林类型进行探讨。以往的研究表明,从降低空气细菌含量方面考虑,绿地面积大于0.09 hm2才能形成“森林内环境”,为居民提供良好的游憩地[7];鸟类栖息地面积范围至少要大于1.5 hm2,且在1.5~30 hm2之间都具有较高的生态效益比[8],10~35 hm2的公园可为大部分鸟类提供家园[9]。李延明等[10]的研究表明,规模大于3 hm2且绿化覆盖率达到60%以上的集中绿地, 能够发挥较好的缓解城市热岛效应的功能;而任志彬[11]的研究表明城市森林斑块面积对热环境效应作用效率最大的区间为2.8~25 hm2。本文基于以上研究结论,对2 039个“留白增绿”地块尺度进行统计(图 1),其中0.09~1.5 hm2范围的地块比例高达49.5%,说明大部分“留白增绿”地块能够起到提供良好游憩服务的功能,1.5 hm2以上的地块占比15.8%,能够发挥居民游憩、鸟类栖息、缓解热岛效应等不同功能。
![]() 图 1 “留白增绿”不同尺度地块的数量Figure 1. Classification of different scales of "leave blank apace and increase green space"
图 1 “留白增绿”不同尺度地块的数量Figure 1. Classification of different scales of "leave blank apace and increase green space"结合不同尺度城市森林主导功能、场地周边用地类型、总体规划定位三大要素对城市森林功能类型进行探讨。在划分生态、生活、生产3个一级分类的基础上,提出栖息生境型、环境调节型、景观游憩型、科普教育型、康养服务型、设施防护型、经济生产型共7个二级分类(图 2)。
![]() 图 2 “留白增绿”背景下的城市森林功能类型划分Figure 2. Urban forest function type division under the background of "leave blank space and increase green space"
图 2 “留白增绿”背景下的城市森林功能类型划分Figure 2. Urban forest function type division under the background of "leave blank space and increase green space"城市化进程迅猛推进后,建设城市森林成为协调人与自然关系、发展生态城市的重要手段。然而,城市森林建设受到诸多因素影响,如何确定城市森林主导功能类型成为一大难点。因此对“留白增绿”背景下的城市森林功能类型进行探讨,以期为城市森林营建提供目标指引。
2. 栖息生境型城市森林营建策略
《北京城市总体规划(2016年—2035年)》提出建设森林环绕的生态城市,提升生态空间品质;要求加强城市生态建设,重视生物多样性保护。城市化进程造成了城市生物多样性锐减,重视和保护生物多样性成为城市生态化发展的重点。因此,本文主要对以提高物种多样性为主要目标的栖息生境型城市森林进行营建策略研究。
生境又称栖息地,是指生物的居住场所,即生物个体、种群或群落能在其中完成生命过程的空间[12]。本文所提到的栖息生境是指动物发生取食、繁殖、筑巢等行为的主要生存环境。多样化的生境包含更多适宜物种生存的条件[13],生境多样性的营建是实现城市生物多样性再生的关键技术之一[14]。
2.1 目标物种选择
为保证生态系统的良性循环,需要考虑构建完整的食物链。城市人工生境可分为人工陆地生境和人工水体生境两大类。人工生境营造中主要针对的生物类型有鱼类、底栖类、两栖类、爬行类、鸟类、哺乳类等。在栖息生境型城市森林营建中,应考虑在以上生物类型中选择目标类群,并针对其中的目标物种营建栖息生境。
鱼类、底栖类、两栖类、爬行类、哺乳类对生境的需求相对较为简单,在有一定绿化面积、一定浅水区域、植物群落结构丰富、土壤有机质充足的环境中能够普遍适应。鸟类处于城市生态系统的顶端[15],其种类及数量在一定程度上决定了其他等级生物的生存状态,在一定程度上代表了生态系统的物种多样性及生态系统的健康程度[16-17],同时鸟类可以作为环境监测的指标[18],是城市生态系统综合质量的指示生物类群[19]。因此,本研究以鸟类为主要目标物种,从林地生境、水体生境两种主要生境类型提出栖息生境型城市森林营建策略。
2.2 林地生境营建策略
2.2.1 增加乡土植物比例
植物多样性是影响鸟类多样性的重要生态因子[20],同时植被也是影响城市森林景观风貌的重要自然因素,乡土植物能够充分适应当地环境和气候条件,体现地区植物区系景观风貌,创造“景观文化”的本土化[21]。乡土植物的应用有助于奠定城市森林景观风貌的整体基调。《国家森林城市评价指标:LY/T2004—004林》中提出,乡土树种应占城市绿化树种使用数量的80%以上。北京五环内记录到的包含279种乡土植物在内的共536种植物[22]及北京市《乡土植物资源发展名录》推荐的82种乔灌草植被为提高城市森林乡土植物比例和奠定森林景观风貌的基础提供了参考。结合国家森林城市评价指标,本研究认为在北京市城市森林营造中应保持乡土植物比例在80%以上。
2.2.2 调整植物群落结构
空间层次越复杂、植物群落越丰富,提供给鸟类的栖息场所和食物越多[23]。通过改进植物群落的树种组成和结构可以有效提高城市绿地鸟类的多样性水平[24]。乔木层和灌木层的结构和组成对栖息地质量起到决定性作用[25]。大面积多树种的阔叶林,可提高鸟类繁殖群落的多样性[26]。植被结构的中、下层绿化,以丰富的垂直结构增加了环境次级结构的多样性[27],为鸟类提供了更多繁殖、摄食和隐蔽的空间[26]。陈自新等[28]对北京市绿地生态效益调研分析后指出,北京市绿地中所用乔木(株)、灌木(株)、草本(m2)、绿地(m2)最适合的种植比例不少于1:6:21:29。本研究认为在栖息生境型城市森林营造中,应在上述结论的基础上,适当增加阔叶林、灌草层植物的比例,控制每29 m2绿地上乔木(株)、灌木(株)、草本(m2)比例在1:6:21以上。
2.2.3 控制植物群落郁闭度
植被郁闭度可以作为衡量栖息地的重要指标[29]。不同郁闭度的乔、灌、草植物群落保证了植物在单元空间中的互相作用,更加充分的吸收利用自然资源。完全郁闭的树林,阻碍鸟类的飞行出入,因此成林时郁闭度应小于90%[30],纯针叶林郁闭度应小于85%[23]。本研究认为城市森林应控制整体林地植物群落郁闭度在70%~85%左右,为鸟类提供更适宜的生活空间。结合黄越[31]对北京鸟类生境类型的调查研究,本文提出10种不同郁闭度的二级生境类型(表 1),对应不同乔灌草搭配比例及高度。除草地外,其他9种二级生境可结合常绿、落叶、混交构成,形成27种三级生境类型。共形成28种不同郁闭度三级生境类型。
表 1 不同郁闭度二级生境类型Table 1. Secondary habitat types with different canopy closure一级类型
Primary classification二级类型
Secondary classification说明
Description乔木林
Tree forest疏林草地
Sparse forest-grass land无灌木或灌木覆盖度小于15%,乔木覆盖度在30%~70%之间
The shrub-free or shrub coverage is less than 15%, and the tree coverage is between 30% and 70%密林草地
Jungle forest-grass land无灌木或者灌木覆盖度小于15%,乔木覆盖度在70%~90%之间
The shrub-free or shrub coverage is less than 15%, and the tree coverage is between 70% and 90%疏林灌丛
Sparse forest-shrubbery灌木覆盖度在30%~70%之间,乔木覆盖度在30%~70%之间
Shrub coverage is between 30% and 70%, and the tree coverage is between 30% and 70%密林灌丛
Jungle forest-shrubbery灌木覆盖度在30%~70%之间,乔木覆盖度在70%~90%之间
Shrub coverage is between 30% and 70%, and the tree coverage is between 70% and 90%疏林灌草地
Sparse forest-shrubbery-grass land灌木覆盖度在15%~30%之间,乔木覆盖度在30%~70%之间
Shrub coverage is between 15% and 30%,and the tree coverage is between 30% and 70%密林灌草地
Jungle forest-shrubbery-grass land灌木覆盖度在15%~30%之间,乔木覆盖度在70%~90%之间
Shrub coverage is between 15% and 30%, and the tree coverage is between 70% and 90%复层林
Multiple layer forest灌木覆盖度大于15%,乔木覆盖度大于30%
Shrub coverage is more than 15% and tree coverage is more than 30%灌丛
Shrubbery矮灌丛
Short shrubbery灌丛高度小于1 m,灌木覆盖度大于30%,乔木覆盖度小于30%
Shrub height is less than 1 m, coverage is more than 30%, and tree coverage is less than 30%灌丛
Shrubbery灌丛高度大于1 m,独干灌木高度小于1.5 m,灌木覆盖度大于30%,乔木覆盖度小于30%
The shrub higher than 1 m, the height of branch independent plant is less than 1.5 m, the shrub coverage is more than 30%, and the tree coverage is less than 30%草地Lawn 草地
Lawn以草本植物为主,木本植物覆盖度小于15%
Mainly herbaceous, woody plant coverage is less than 15%2.2.4 增加食源树种比例
食源树种的种类和数量对鸟类物种多样性产生影响[32]。木本植物挂果期长短与鸟类多度存在显著相关性[20]。乡土树种果实成熟期与鸟类繁殖期或迁徙期基本一致,本地鸟类经过长期的自然选择,更倾向于在乡土树种上取食、栖息和停留[33]。在栖息生境型城市森林营造中应考虑增加能够为鸟类提供食物(如种子、花芽、果实和浆果等)的树种[34],并保证四季均衡供应[32]及食源树种在生境范围内的均匀分布。通过对前人研究的总结,发现因调查样地尺度各异、环境条件复杂多变、植物配置情况多样、鸟类取食喜好多样,难以对最优食源树种组团类型及比例进行量化。故本研究仅提出增加食源树种比例、兼顾食源树种季相搭配及均衡分布的策略,而不对具体比例进行限定。
2.2.5 增加常绿乔木比例
增加常绿乔木的比例能为鸟类提供良好的栖息场所,提高鸟类多样性[35],同时常绿针叶植物能够在冬季为鸟类提供一定的食源。根据李淑凤[36]对北京市公园绿地植物配置的研究,常绿树一般占落叶树的30%~40%即可,即常绿落叶比例约为23:77~29:71。在栖息生境型城市森林营造中,为保证冬季鸟类觅食及隐蔽需求,将栖息生境城市森林中常绿落叶植物比例提高到4:6,为鸟类创造更适宜的栖息条件。
2.3 水体生境营建策略
2.3.1 丰富水体形式
水域生态的复杂多样影响鸟类群落多样性[37]。为增加环境多样性,在保持原有水域的基础上适当增加各种形式的水体,以满足鸟类的生存需要[27]。通过曲折蜿蜒的岸线,提供港湾、浅滩、半岛等栖息条件,营造有近圆形核心、弯曲边界和边缘指状突起的水系形态[38](图 3)。创造丰富栖息生境的同时增加物种传播的可能。
在水域面积大于5 hm2时,建议在水中设置岛屿,供鸟类栖息[31]。研究表明,随着岛屿面积增大,单位面积内岸线周长迅速减小,当岛屿面积超过3 000 m2时,岸线变化趋于平缓[38](图 4)。创造多种生境类型,需控制岸线与岛屿面积比例。因此,在栖息生境型城市森林营造中应控制单个岛屿面积在3 000 m2以下,且有大小变化。控制岛屿与水体岸线距离在10 m以上[39]。
![]()
2.3.2 控制水体深度
不同种水鸟在觅食和营巢上对水深有着不同的需求,涉禽和游禽对水深条件要求相对较高。在选定以涉禽和游禽为水体生境主要目标物种的前提下,认为北京地区更适合通过低水位营造野生动物栖息地[40],水深通常应不大于1 m[41]。在水域范围内,分别营造0~10 cm、10~20 cm、20~30 cm的水深环境满足涉禽生境需求,营造30~100 cm、100 cm以上水深满足游禽生境需求,同时控制整体水深不超过6 m(图 5),并根据选定目标种中游禽、涉禽所占比例,分配不同水深的水体面积。
2.3.3 控制水生植物覆盖度
水生植物的多样性对鸟类群落的影响较大[37]。结合前人的研究,不同鸟类对水生植物覆盖度有不同要求,但普遍低于70%。在栖息生境型城市森林营造中,保留至少1/3的水面供鸟类起飞或降落[42],2/3的水面分别以0%~30%、30%~50%、50%~70%的覆盖度种植水生植物(图 6),并根据不同物种的需求,构建不同覆盖度水体环境,保证生境类型的多样性。
![]() 图 6 不同水生植物覆盖度示意图Figure 6. Schematic representation of aquatic plants with different coverage
图 6 不同水生植物覆盖度示意图Figure 6. Schematic representation of aquatic plants with different coverage2.3.4 改良驳岸类型
硬质驳岸及硬化池底极不利于水生生物的生存,影响部分肉食性水鸟的取食。在栖息生境型城市森林营造中,为便于鸟类觅食、隐藏等活动,驳岸坡度应尽量控制在10:1或更小,营造一定的裸露滩涂和砂石驳岸[41]。以透气性较好的煤渣垫层、有机介质等铺设池底,使土壤渗透率保持在0.025~0.35 cm/h[38]。在保证一定面积水域常水位水深的基础上,充分利用自然降水变化形成季节性水体环境(图 7)。
3. 横街子栖息生境型城市森林营建
本文以“留白增绿”地块中位于东南五环与京沪高速交汇处的横街子地块为研究对象(图 8)。横街子地块由3块“留白增绿”地块组成,总面积62.6 hm2,处于北京市第一道绿化隔离带范围内。场地内部有较好的水源条件,因拆迁所产生的约9.2 hm2低地可以作为水系营造使用。李湛东等[43]的研究结果表明,横街子地块及周边区域植物多样性较低。周边绿地数量较多、功能较为完善,能满足游人休闲游憩的需求。结合前文对城市森林功能类型的探讨,认为横街子地块适宜营建以提高生物多样性为主要目标的栖息生境型城市森林。
3.1 目标物种选择
以前文的营建策略为基础,在横街子栖息生境型城市森林营造中,以鸟类作为生境营建的主要目标类群。根据黄越[31]对北京市圆明园、中山公园等26个城市公园鸟类生境的调查研究,林鸟中红嘴蓝鹊(Urocissa erythroryncha)、金翅雀(Carduelis sinica)和斑鸫(Turdus eunomus)可以作为针叶林生境代表物种;白头鹎(Pycnonotus sinensis)、四声杜鹃(Cuculus micropterus)、大杜鹃(Cuculus canorus)、丝光椋鸟(Sturnus sericeus)、黄眉柳莺(Phylloscopus inornatus)、燕雀(Fringilla montifringilla)可以作为阔叶林生境代表种;红喉姬鹟(Ficedula parva)、褐柳莺(Phylloscopus fuscatus)和黄喉鹀(Emberiza elegans)可以作为灌丛生境的代表种;水鸟中苍鹭(Ardea cinerea)、草鹭(Ardea purpurea)、大白鹭(Ardea alba)可以作为不同营巢习性的代表种;黑水鸡(Gallinula chloropus)、鸳鸯(Aix galericulata)、小鸊鷉(Tachybaptus ruficollis)可以分别作为水岸、近水及深水的代表种;近水鸟普通翠鸟是溪流状近水生境的代表种。在横街子栖息生境型城市森林营造中以上述代表物种作为目标种营建栖息生境。
3.2 横街子栖息生境型城市森林营建的空间策略
横街子地块紧邻东南五环与京沪高速,有较大的噪声干扰。研究表明针阔混交林对实时交通噪声有较好的衰减作用[44],其中阔叶乔木降噪能力优于针叶乔木[45]。乔灌层宽度对噪音衰减值的影响最为显著,在30 m内随着林带宽度的增加, 降噪效果更加明显[46]。为避免快速交通对城市森林内部的干扰,在场地临近快速路一侧设置30 m针阔混交林并搭配灌草层作为隔离带。在场地内部营建生境核心斑块(图 9)。
3.3 横街子城市森林生境营造策略
3.3.1 林地生境营造
参照前文栖息生境型城市森林营建策略,在林地生境营造中,以北京市乡土植物为主要树种结合北京市《乡土植物资源发展名录》推荐的82种乔灌草植被作为横街子城市森林植物选择名录,搭配食源树种,控制不同类型植物群落郁闭度(图 10),控制常绿落叶比例约为4:6(图 11),形成栖息生境型城市森林基底。在此基础上依据林地生境目标鸟类的生活习性营建栖息生境(表 2)。
![]() 图 10 不同郁闭度群落分布示意图Figure 10. Schematic diagram of the distribution of plant communities with different canopy closure
图 10 不同郁闭度群落分布示意图Figure 10. Schematic diagram of the distribution of plant communities with different canopy closure![]() 图 11 常绿及落叶植物分布示意图Figure 11. Schematic representation of evergreen and deciduous plant distribution表 2 林地鸟类生境营造Table 2. Woodland bird habitat construction conditions
图 11 常绿及落叶植物分布示意图Figure 11. Schematic representation of evergreen and deciduous plant distribution表 2 林地鸟类生境营造Table 2. Woodland bird habitat construction conditions林地类型
Forest land type鸟的种类
Species of birds生境需求
Habitat requirement针叶林
Coniferous forest金翅雀Carduelis sinica、斑鸫Turdus eunomus 保证上层林木有一定盖度且生境下层较开敞, 草本植物丰富[49]
Ensure that the upper trees have a certain degree of coverage and the lower layer of the habitat is open with rich herbs[49]红嘴蓝鹊Urocissa erythroryncha 连续常绿林面积至少4 hm2[31]
Continuous evergreen forest area is of at least 4 ha[31]阔叶林
Broadleaved forest habitat燕雀Fringilla montifringilla 拾取草籽为食,喜上层乔木主干清晰,下层灌木稀疏的群落[47]
Picking up grass seeds for food, like the environment with clear branches of upper trees and lower sparse shrub community[47]白头鹎Pycnonotus sinensis、丝光椋鸟Sturnus sericeus 喜主干清晰下层开阔的阔叶乔木群落,灌木下层稀疏的植物或体量小的球形灌木[49]。距地面大多2~3 m筑巢,亦有筑在6~6.5 m高大乔木上[30]
Like the environment with clear branches and broadleaved trees, lower sparse shrub community or small spherical shrub[49]. Nesting on places 2-3 m higher than the ground, and nesting on 6-6.5 m tall trees[30]四声杜鹃Cuculus micropterus、大杜鹃Cuculus canorus 喜乔木主干通直,下层相对开阔,以阔叶林为主。搭配树龄较长的针叶树。喜既不影响飞行又能覆盖地表的丰富小灌木及草本空间[47]
Like trees with straight trunk, the lower layer is relatively open, mainly broadleaved forest. Pair with longer-aged conifers. Like rich shrubs and herbaceous spaces cover the surface but do not affect flight activities[47]黄眉柳莺Phylloscopus inornatus 喜株型挺拔整齐的高大乔木, 偏好落叶林和常绿林,如落叶密林草地和常绿密林灌丛草地[31]
Like straight and neat trees, preference deciduous forest and evergreen forest, such as deciduous jungle grassland and evergreen jungle shrub grassland[31]灌丛Shrub 黄喉鹀Emberiza elegans、红喉姬鹟Ficedula parva、褐柳莺Phylloscopus fuscatus 喜中等灌木和低矮的自然地被[30],常活动在林缘以及溪流沿岸的疏林与灌丛[48]
Like medium shrubs and low herbs[30], of ten active in forests and shrubs along the forest margins and streamlined[48]3.3.2 水体生境营造
根据场地基础条件,场地内部有约9.2 hm2水面能够营造水体生境。通过曲折蜿蜒的岸线,提供港湾、浅滩、半岛等栖息条件,并营造多个小型岛屿(图 12)。在此基础上依据水体生境目标鸟类的生活习性设计水体深度及水生植被覆盖率(表 3)。
![]() 图 12 水体生境示意图Figure 12. Schematic representation of evergreen and deciduous plants distribution表 3 水体鸟类生境营造Table 3. Aquatic habitat construction conditions
图 12 水体生境示意图Figure 12. Schematic representation of evergreen and deciduous plants distribution表 3 水体鸟类生境营造Table 3. Aquatic habitat construction conditions类别
Category科
Family种
Species生境需求
Habitat requirement水鸟
Water birds鹭科
Ardeidae大白鹭
Ardea alba喜在近水域或水中岛屿高树上营巢[49]。水域内需沉水、挺水植物覆盖率40%~60%之间,高于1 m的植物占60%左右,木本植物、挺水植物4 m以内越高越好。陆地植物7 m内越高越适宜[30]
Like to nest on high trees in the water or on the island[49].The coverage of aquatic plants in the waters is between 40% and 60%, plants above 1 m is about 60%. Woody plants and aquatic plants should be within 4 m and plants on land should within 7 m, the higher, the better [30]水鸟
Water birds鹭科
Ardeidae草鹭
Ardea purpurea喜栖息在水边灌丛或芦苇(Phragmites communis)沼泽[49]。水域内需沉水、挺水植物覆盖率40%~60%之间,高于1 m的植物占60%左右,木本植物、挺水植物4 m以内越高越好。陆地植物7 m内越高越适宜[30]
Like to inhabit in the shrubs or the reeds.The coverage of aquatic plants in the waters is between 40% and 60%, plants above 1m is about 60%. Woody plants and aquatic plants should be within 4 m and plants on land should within 7 m, the higher, the better [30]水鸟
Water birds鹭科
Ardeidae苍鹭
Ardea cinerea喜栖息于有大片芦苇和水生植物的浅水域[49]。水域内需沉水、挺水植物覆盖率40%~60%之间,高于1 m的植物占60%左右,木本植物、挺水植物4 m以内越高越好。陆地植物7 m内越高越适宜[30]
Like to inhabit in shallow waters with large reeds and aquatic plants.The coverage of aquatic plants in the waters is between 40% and 60%, plants above 1 m is about 60%. Woody plants and aquatic plants should be within 4m and plants on land should within 7 m, the higher, the better [30]水鸟
Water birds鸭科
Anatidae黑水鸡
Gallinula chloropus栖息于富有树木、芦苇和水生挺水植物遮蔽的淡水水域,不喜欢很开阔的场所[49]。栖息地水域植被覆盖率50%~75%为宜,其中30%~50%灌木,40%~70%挺水植物,0%~10%乔木,以及25%水面,水深0.9 m以内水域中宜有倒伏树干[30]
Inhabit in freshwater covered with trees, reeds and aquatic plants, do not like very open space[49]. The vegetation coverage 50%-75%, with 30%-50% shrubs, 40%-70% aquatic plants, 0%-10% trees, and 25% water surface. There should be fallen tree trunks in the water within 0.9 m[30]水鸟
Water birds鸭科
Anatidae鸳鸯
Aix galericulata喜在水深不超过2 m的浅水区觅食,水域植被覆盖率50%~75%为宜,近水区域结合少量乔灌,以木本上层结构为主[30]
Like to eat in shallow waters with a water depth no more than 2 m, vegetation coverage in the waters should is 50%-75%, near-water area combined with a small amount of trees and shrubs, mainly based on woody superstructure[30]水鸟
Water birds鸭科
Anatidae小鸊鷉Tachybaptus ruficollis 在芦苇、香蒲(Typha orientalis Presl)、灯芯草(Juncus effusus L.)等挺水植被区域活动,挺水植物占整个水面比例小于30%,栖息地生态岛上高度0.6~10 m的植被覆盖率50%以上为宜,距离人类活动不宜小于400 m[30]
Activities in the watery vegetation areas such as reeds, cattails, and rushes. Vegetation coverage is less than 30%. The coverage of trees with a height of 0.6-10 m on the habitat ecological island is more than 50%. Human activities should be outside the distance of 400 m[30]近水鸟
Near- water birds翠鸟科
Alcedinidae普通翠鸟Alcedo atthis 栖息于有灌丛或疏林、水清澈而缓流的小河、溪涧、湖泊以及灌溉渠等水域[49]。岸际15 m以内浮水植物、原木、岩石等阻碍越少越好[30]
Inhabited in rivers, streams, lakes and irrigation canals with shrubs or sparse forests, clear waters and slow streams[49]. The less obstructions such as floating plants, logs and rocks within 15 m of the coast, the better[30]4. 结论与启示
城市森林功能的多样性、城市环境条件的异质性决定了未来城市森林建设模式的多样性。本文将“留白增绿”背景下的城市森林定义为在城市地域范围内拆违腾退土地上,结合地块尺度、周边用地类型、总体规划定位三大要素所建立的能够针对性发挥城市森林生态、生活、生产功能,符合城市森林景观风貌的林木群落及其环境所构成的森林生态系统。根据“留白增绿”地块尺度、周边用地类型及总体规划定位要求对可能营造的城市森林功能类型进行探讨,在生态型、生活型、生产型3个一级分类的基础上,提出栖息生境型、环境调节型、景观游憩型、科普教育型、康养服务型、设施防护型、经济生产型共7个二级分类,以期为城市森林营建提供目标指引。多种功能类型的城市森林将成为城市生态环境的重要组成部分。
在此基础上,基于“留白增绿”的城市发展背景,选择目标物种,依据目标物种所需的主要生境类型,提出栖息生境型城市森林营建策略。主要分为林地生境营建策略、水体生境营建策略。林地生境营建策略包括增加乡土树种比例、调整植物群落结构、控制植物群落郁闭度、增加食源植物比例、增加常绿乔木比例;水体生境营建策略包括丰富水体形式、控制水体深度、控制水生植物覆盖度、改良驳岸类型。以横街子栖息生境型城市森林为例,根据场地内部条件,选择目标物种,结合上述林地生境、水体生境营建策略,为目标物种营造适宜栖息的生境条件。
但需要指出的是,本文是基于提高生物多样性为目标的栖息生境型城市森林所进行的营建策略研究,其建设策略更多地是以适宜生物栖息为原则提出的。营建城市森林是落实城市生态文明建设的重要途径,但城市森林功能并不唯一,其环境调节、生活游憩、生产防护等功能也是城市森林营建时需要考虑和重视的。因此,针对不同的场地条件,我们需要全方位考虑并择优选择和建立不同功能类型的城市森林。
-
![]()
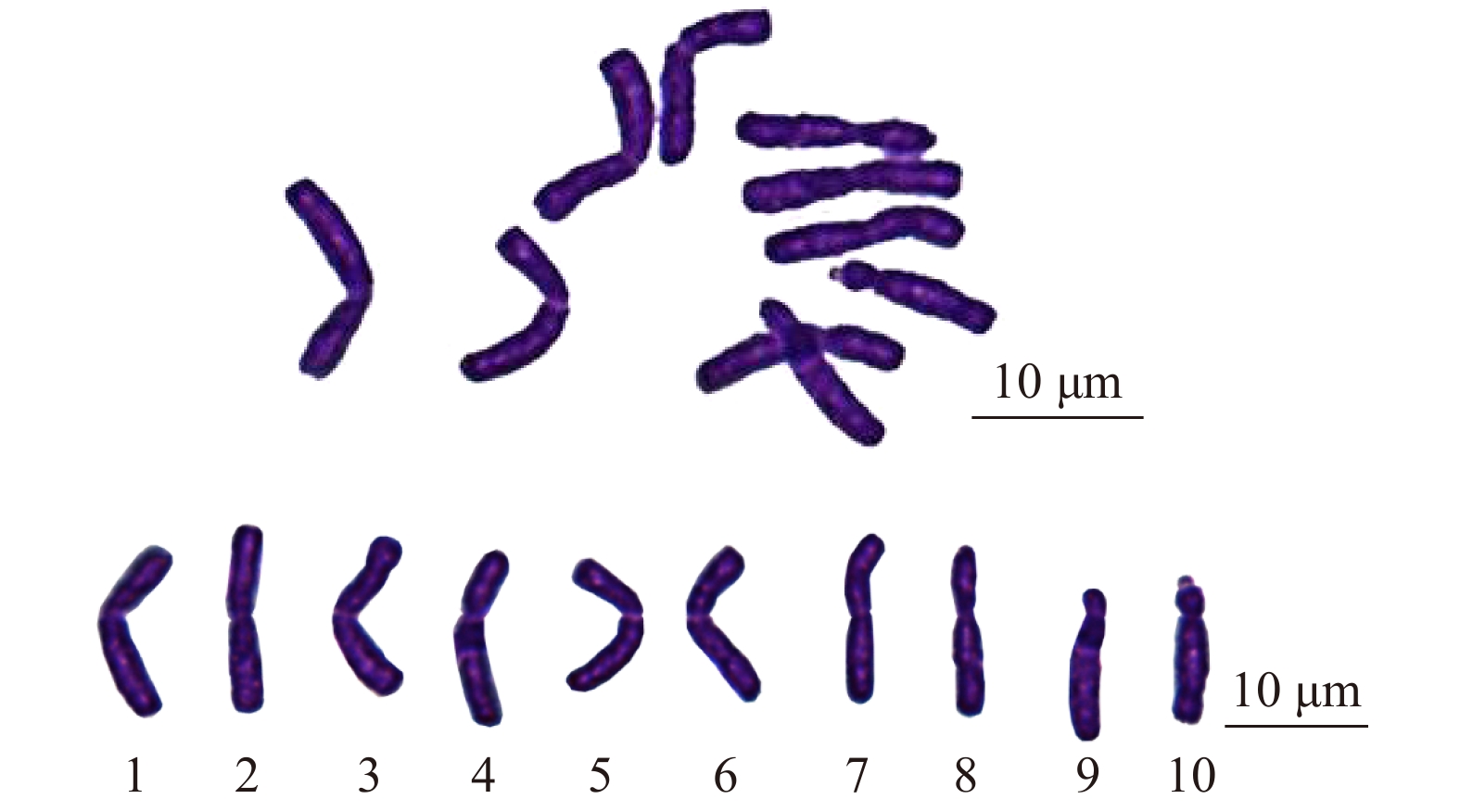
图 1 ‘正午’牡丹中期染色体及核型
Figure 1. Metaphase chromosomes and karyotype of Paeonia × lemoinei ‘High Noon’
![]()
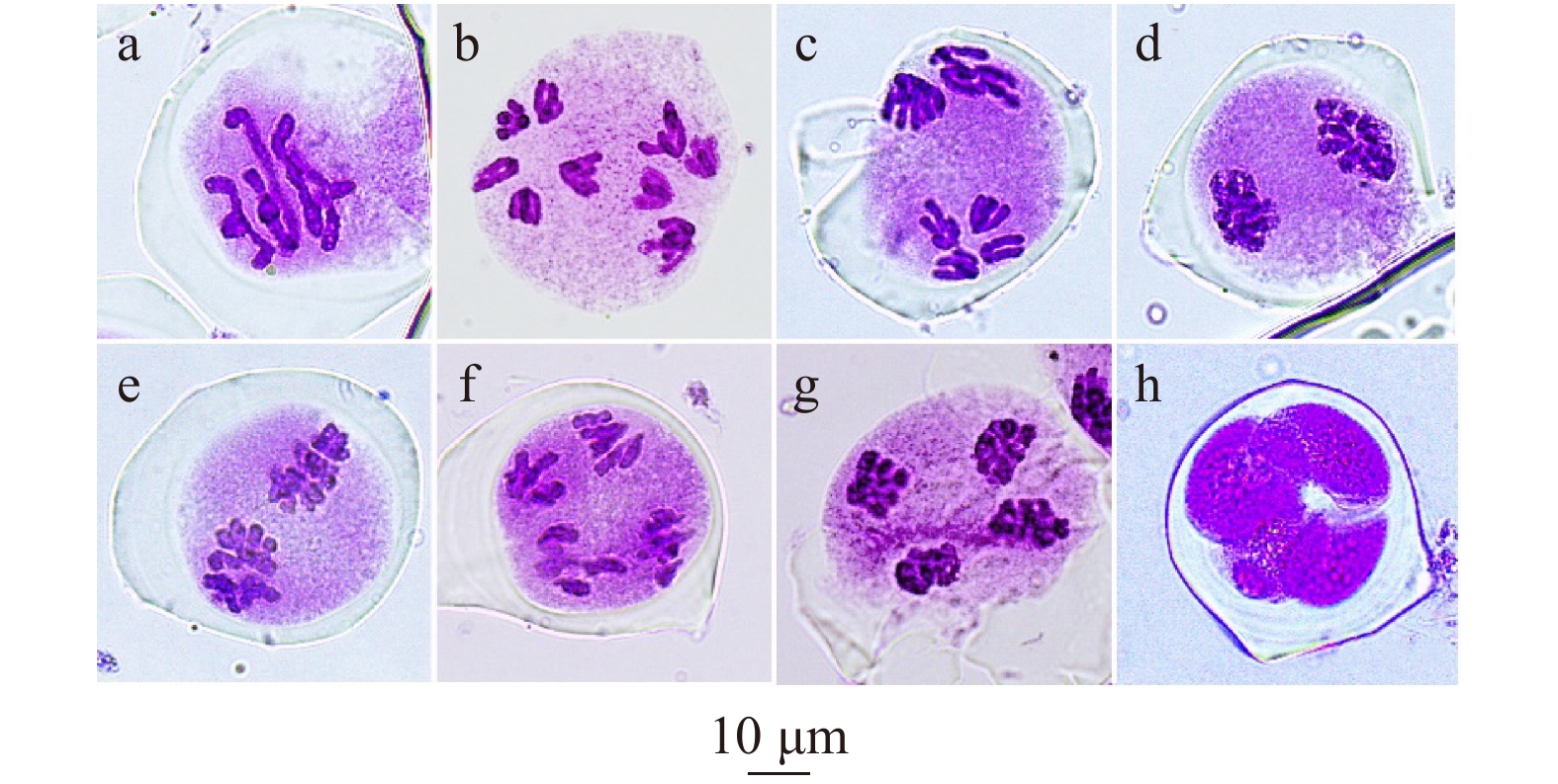
图 2 ‘正午’牡丹花粉母细胞的正常减数分裂过程
a:中期I,5个整齐排列的棒状二价体;b、c:后期I;d:末期I;e:中期II;f:后期II;g:末期II;h:四分体时期。 a, metaphase I with 5 rod bivalents orderly arranged; b,c, anaphase I; d, telophase I; e, metaphase II; f, anaphase II; g, telophase II; h, tetrad.
Figure 2. Regular meiosis of pollen mother cells in Paeonia × lemoinei ‘High Noon’
![]()
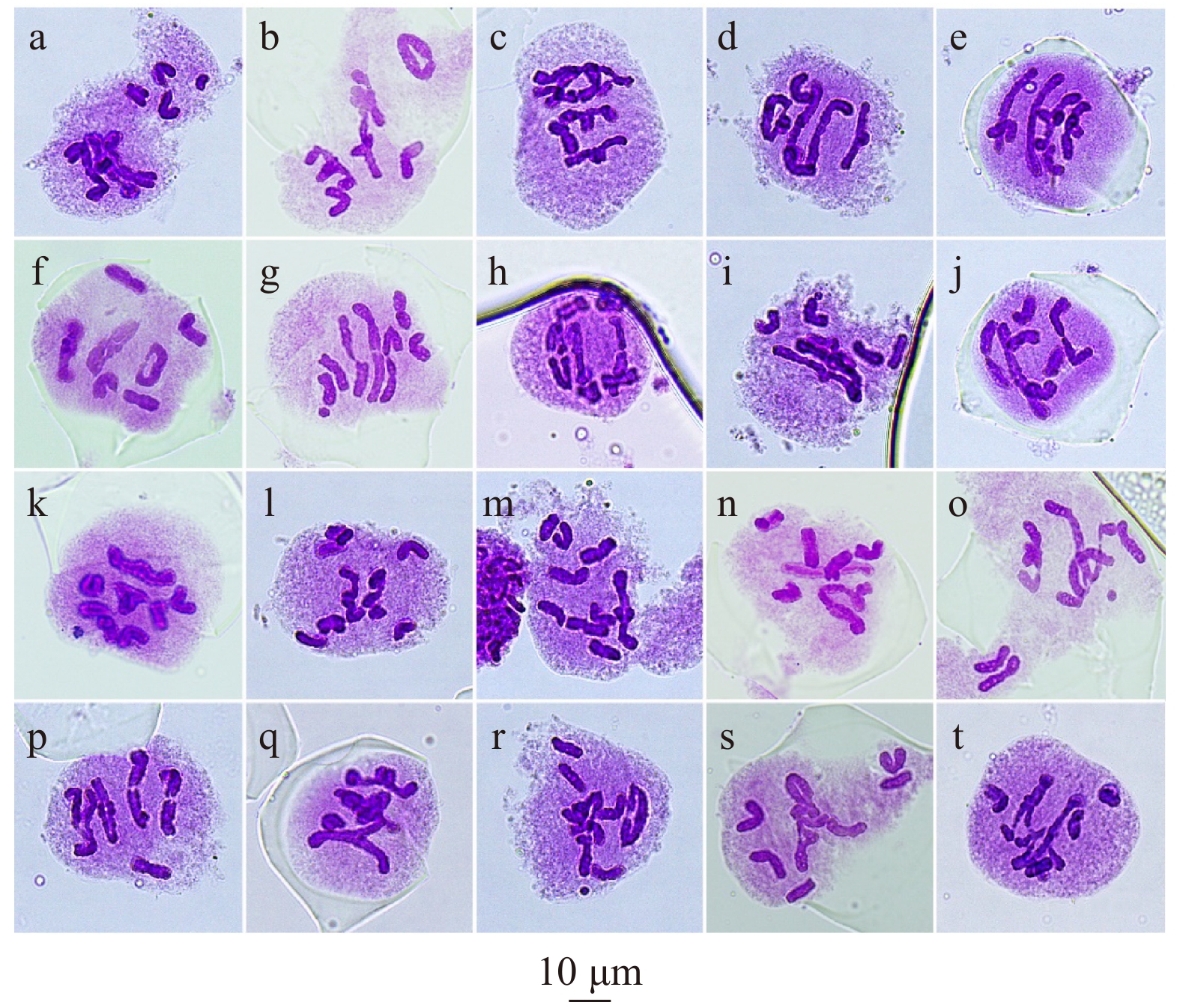
图 3 减数第一次分裂中期的异常
a ~ e:4个二价体 + 2个单价体(a、e:4个棒状二价体 + 2个单价体,b ~ d:3个棒状二价体 + 1个环形二价体 + 2个单价体);f ~ j:3个二价体 + 4个单价体;k、l:2个二价体 + 6个单价体(k:棒状二价体 + 环形二价体 + 6个单价体,l:2个棒状二价体 + 6个单价体);m:1个二价体 + 8个单价体;n:1个二价体 + 1个三价体 + 5个单价体;o:2个二价体 + 1个三价体 + 3个单价体;p:3个二价体 + 1个三价体 + 1个单价体;q:1个二价体 + 1个七价体 + 1个单价体;r:1个二价体 + 1个五价体 + 3个单价体 + 1个断片;s:1个五价体 + 5个单价体;t:1个六价体 + 4个单价体。a−e, 4 bivalents + 2 univalents (a, e, 4 rod bivalents + 2 univalents, b−d, 3 rod bivalents + 1 ring bivalent + 2 univalents); f−j, 3 bivalents + 4 univalents; k−l, 2 bivalents + 6 univalents (k, rod bivalent + ring bivalent + 6 univalents; l, 2 bivalents + 6 univalents); m, 1 bivalent + 8 univalents; n, 1 bivalent + 1 trivalent + 5 univalents; o, 2 bivalents + 1 trivalent + 3 univalents; p, 3 bivalents + 1 trivalent + 1 univalent; q, 1 bivalent + 1 heptavalent + 1 univalent; r, 1 bivalent + 1 quinquevalent + 3 univalents + 1 fragment; s, 1 quinquevalent + 5 univalents; t, 1 hexavalent + 4 univalents.
Figure 3. Meiotic abnormalities in metaphase I
![]()
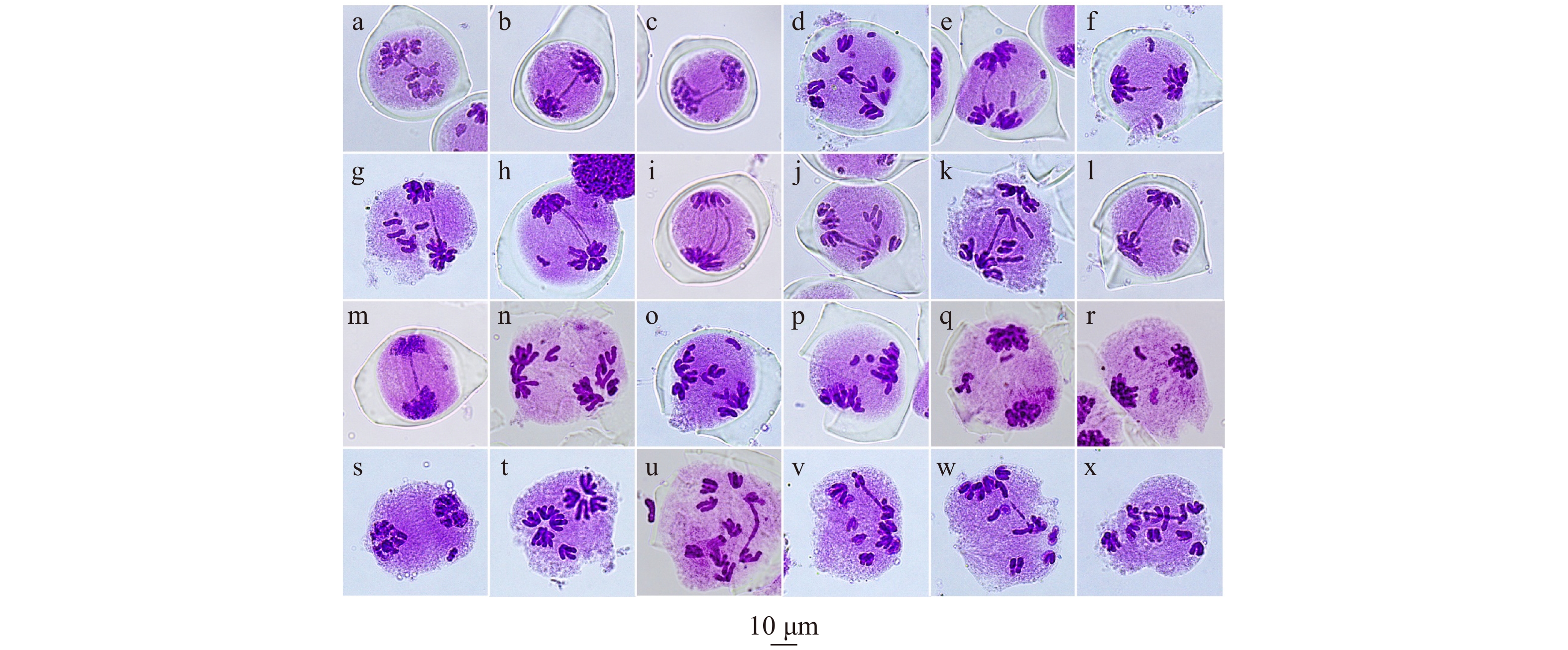
图 4 减数第一次分裂后期到末期的异常
a ~ c:有染色体桥,无断片;d ~ g:有染色体桥及1 ~ 6个断片;h、i:染色体双桥及断片;j ~ l:染色体桥和落后染色体;m:末期染色体桥及断片;n ~ s:无染色体桥,有落后染色体及断片;t:染色体不等分离;u ~ x:不等分离、染色体桥、落后染色体及断片同时存在。a−c, chromosome bridge with no fragment; d−g, chromosome bridge with 1−6 fragments; h−i, chromosome double bridge with fragments; j−l, chromosome bridge with lagging chromosome; m, chromosome bridge with fragment in telophase I; n−s, lagging chromosome and fragment with no chromosome bridge; t, unequal segregation of chromosomes; u−x, unequal segregation of chromosomes with chromosome bridge, lagging chromosome and fragment.
Figure 4. Meiotic abnormalities in anaphase I and telophase I
![]()
图 5 减数第二次分裂的异常
a ~ f:中期II的不同纺锤体定位(a:直线纺锤体,b:垂直纺锤体,c、d:三极纺锤体,e:融合纺锤体,f:平行纺锤体);g ~ n:减II后期的异常(g:染色体桥,h:对角线桥,i、j:三极纺锤体后期形成的2组减数染色体及1组未减数染色体,k:融合纺锤体后期形成的2组未减数染色体,l:4组染色体及额外染色体,m:后期II的不同步性,形成2组减数染色体及1组未减数染色体,n:后期II的对角线桥及染色体断片);o ~ t:末期II的异常(o、p、s:2个减数核和1个未减数核,p:3个核及染色体桥、断片,q:2个未减数核,r:4个核及2个微核,t:末期对角线桥);u:三分体;v:三分体和微核;w:二分体;x:2个正常小孢子、1个具微核的小孢子和1个特小的小孢子。a−f, aspects of meiotic spindles at metaphase II (a, linear spindle, b, perpendicular spindle, c−d, tripolar spindle, e, fused spindle, f, parallel spindle); g−n, meiotic abnormalities in anaphase II (g, chromosome bridge, h, chromosome diagonal bridge, i−j, 2 groups of reduced chromosomes and 1 group of unreduced chromosomes come from the tripolar spindles, k, 2 groups of unreduced chromosomes come from the fused spindles, l, 4 groups of chromosomes with extra chromosomes, m, 2 groups of reduced chromosomes and 1 group of unreduced chromosomes because of the asynchronized anaphase II, n, chromosome diagonal bridge and fragment in anaphase II); o−t, meiotic abnormalities in telophase II (o, p, s, 2 reduced nucleuses and 1unreduced nucleus, p, 3 nucleuses with chromosome bridge and fragment, q, 2 unreduced nucleuses, r, 4 nucleuses with 2 micronucleuses, t, diagonal bridge in telophase II); u, triad; v, triad with micronucleus; w, dyad; x, 2 regular microspores, 1 microspore with micronucleus and 1 extra small microspore.
Figure 5. Meiotic abnormalities in the second meiotic division
表 1 ‘正午’牡丹核型数据
Table 1 Karyotype parameters of Paeonia × lemoinei ‘High Noon’
染色体序号
Chromosome No.相对长度
Relative length/%臂比
Arm ratio染色体类型
Chromosome type1 11.77 ± 0.53 1.40 ± 0.05 m 2 11.48 ± 0.53 1.17 ± 0.09 m 3 10.94 ± 0.52 1.16 ± 0.08 m 4 10.65 ± 0.23 1.52 ± 0.13 m 5 10.25 ± 0.34 1.39 ± 0.10 m 6 10.09 ± 0.24 1.15 ± 0.11 m 7 9.59 ± 0.29 1.23 ± 0.21 m 8 9.37 ± 0.47 1.71 ± 0.24 sm 9 8.27 ± 0.59 4.39 ± 0.37 st 10 7.58 ± 0.42 3.53 ± 0.17 st* 注:*随体染色体,随体长度未计算在染色体长度内。Note: * represents sat-chromosome, the length of satellite was not included.  下载: 导出CSV
下载: 导出CSV
-
[1] 李嘉珏, 张西方, 赵孝庆, 等. 中国牡丹[M]. 北京: 中国大百科全书出版社, 2011. Li J J, Zhang X F, Zhao X Q, et al. Tree peony of China[M]. Beijing: Encyclopedia of China Publishing House, 2011.
[2] 王莲英. 中国牡丹品种图志[M]. 北京: 中国林业出版社, 1997. Wang L Y. Chinese tree peony[M]. Beijing: China Forestry Publishing House, 1997.
[3] 成仿云, 李嘉珏. 中国牡丹的输出及其在国外的发展Ⅰ: 栽培牡丹[J]. 西北师范大学学报(自然科学版), 1998, 34(1):112−119. Cheng F Y, Li J J. Exportation of Chinese tree peonies (Mudan) and their development in other countriesⅠ: cultivated[J]. Journal of Northwest Normal University (Natural Science), 1998, 34(1): 112−119.
[4] 成仿云, 李嘉珏, 于玲. 中国牡丹的输出及其在国外的发展Ⅱ: 野生牡丹[J]. 西北师范大学学报(自然科学版), 1998, 34(3):106−111. Cheng F Y, Li J J, Yu L. Exportation of Chinese tree peonies (Mudan) and their developments in other countries Ⅱ : wild species[J]. Journal of Northwest Normal University (Natural Science), 1998, 34(3): 106−111.
[5] Zhou S L, Zou X H, Zhou Z Q, et al. Multiple species of wild tree peonies gave rise to the ‘king of flowers’, Paeonia suffruticosa Andrews[J/OL]. Proceedings of the Royal Society B: Biological Sciences, 2014, 281: 1687 [2018−05−06]. https://royalsocietypublishing.org/doi/10.1098/rspb.2014.1687.
[6] Cheng F Y. Advances in the breeding of tree peonies and a cultivar system for the cultivar group[J]. International Journal of Plant Breeding, 2007, 1(2): 89−104. doi: 10.3923/ijpbg.2007.89.94
[7] Page M. The gardener’s peony: herbaceous and tree peonies[M]. Cambridge: Timber Press, 2005.
[8] Wister J C. The peonies[M]. Hopkins: The American Peony Society, 1995.
[9] 王莲英, 袁涛, 王福, 等. 中国芍药科野生种迁地保护与新品种培育[M]. 北京: 中国林业出版社, 2013: 37−38. Wang L Y, Yuan T, Wang F, et al. The ex-situ conservation centre of Chinese Paeoniceae wild species and cultivation of new cultivars[M]. Beijing: China Forestry Publishing House, 2013: 37−38.
[10] 何桂梅, 成仿云. ‘正午’牡丹有性生殖败育的形态学观察[J]. 园艺学报, 2006, 33(3):660−663. doi: 10.3321/j.issn:0513-353X.2006.03.044 He G M, Cheng F Y. Morphological observation of sexual reproduction abortion in ‘High Noon ’ tree peony[J]. Acta Horticulturae Sinica, 2006, 33(3): 660−663. doi: 10.3321/j.issn:0513-353X.2006.03.044
[11] Hao Q, Aoki N, Katayama J, et al. Crossability of American tree peony ‘High Noon ’ as seed parent with Japanese cultivars to breed superior cultivars[J]. Euphytica, 2013, 191: 35−44. doi: 10.1007/s10681-012-0853-3
[12] 吴静, 成仿云, 张栋. ‘正午’牡丹的杂交利用及部分杂种AFLP鉴定[J]. 西北植物学报, 2013, 33(8):1551−1557. Wu J, Cheng F Y, Zhang D. Utilizing ‘High Noon’ in the crossing breeding of tree peonies and early identification of some hybrids by AFLP markers[J]. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(8): 1551−1557.
[13] 李懋学, 陈瑞阳. 关于植物核型分析的标准化问题[J]. 武汉植物学研究, 1985, 3(4):297−302. Li M X, Chen R Y. A suggestion on the standardizaton of karyotype analysis in plants[J]. Journal of Wuhan Botanical Research, 1985, 3(4): 297−302.
[14] 侯小改, 郭大龙, 张亚冰, 等. 部分牡丹品种的染色体核型分析[J]. 河南科技大学学报(自然科学版), 2012, 33(5):83−87. doi: 10.3969/j.issn.1672-6871.2012.05.019 Hou X G, Guo D L, Zhang Y B, et al. Chromosome karyotype analysis of partial cultivars in tree peony[J]. Journal of Henan University of Science and Technology (Natural Science), 2012, 33(5): 83−87. doi: 10.3969/j.issn.1672-6871.2012.05.019
[15] 侯小改, 段春燕, 刘素云, 等. 中国牡丹染色体研究进展[J]. 中国农学通报, 2006, 22(2):307−309. doi: 10.3969/j.issn.1000-6850.2006.02.083 Hou X G, Duan C Y, Liu S Y, et al. Advances on chromosome study of Chinese tree peony[J]. Chinese Agricultural Science Bulletin, 2006, 22(2): 307−309. doi: 10.3969/j.issn.1000-6850.2006.02.083
[16] 龚洵, 顾志建, 武全安. 黄牡丹七个居群的细胞学研究[J]. 云南植物研究, 1991, 13(4):402−410. Gong X, Gu Z J, Wu Q A. A cytological study of seven populations in Paeonia delavayi var. lutea[J]. Acta Botanica Yunnanica, 1991, 13(4): 402−410.
[17] 舒巧利, 陈志, 党江波, 等. 染色体识别技术及其在园艺植物上的应用进展[J]. 园艺学报, 2017, 44(9):1772−1786. Shu Q L, Chen Z, Dang J B, et al. Progress of chromosome identification technology and application on horticultural plants[J]. Acta Horticulturae Sinica, 2017, 44(9): 1772−1786.
[18] Stebbins G L. Cytogenetic studies in Paeonia (Ⅱ): the cytology of the diploid species and hybrids[J]. Genetics, 1938, 23(1): 83−110.
[19] 何丽霞, 李睿, 张延东. 杨山牡丹×紫牡丹杂交后代“梦幻”的细胞遗传学[J]. 兰州大学学报(自然科学版), 2010, 46(4):70−75. He L X, Li R, Zhang Y D. Cytogenetics of the ‘Dream’ hybrid from Paeonia ostii × Paeonia delavayi[J]. Journal of Lanzhou University (Natural Sciences), 2010, 46(4): 70−75.
[20] 何丽霞. 黄牡丹花粉母细胞减数分裂过程的细胞遗传学观察[J]. 兰州大学学报, 2004, 40(6):78−82. doi: 10.3321/j.issn:0455-2059.2004.06.019 He L X. The cytoenetic observation on pollen mother cell of Paeonia lutea[J]. Journal of Lanzhou University (Natural Sciences), 2004, 40(6): 78−82. doi: 10.3321/j.issn:0455-2059.2004.06.019
[21] 李子峰, 王佳, 胡永红, 等. ‘凤丹白’牡丹核型分析与减数分裂的细胞遗传学观察[J]. 园艺学报, 2007, 34(2):411−416. doi: 10.3321/j.issn:0513-353X.2007.02.028 Li Z F, Wang J, Hu Y H, et al. Karyotype analysis and cytology observation on eiosis of Paeonia ostii ‘Phoenix White’[J]. Acta Horticulturae Sinica, 2007, 34(2): 411−416. doi: 10.3321/j.issn:0513-353X.2007.02.028
[22] 王士泉, 张大明. 四川牡丹和块根芍药第五号染色体异常的减数分裂证据[J]. 植物分类学报, 2008, 46(2):155−162. Wang S Q, Zhang D M. Meiotic abnormalities of the fifth pair of chromosomes in Paeonia decomposita and P. intermedia[J]. Journal of Systematics and Evolution, 2008, 46(2): 155−162.
[23] 王雁, 李艳华, 彭镇华. 黄牡丹的大小孢子发生及雌雄配子体发育[J]. 东北林业大学学报, 2010, 38(5):62−65. doi: 10.3969/j.issn.1000-5382.2010.05.018 Wang Y, Li Y H, Peng Z H. Megasporogenesis, microsporogenesis and development of female and male gametophytes of Paeonia delavayi var. lutea[J]. Journal of Northeast Forestry University, 2010, 38(5): 62−65. doi: 10.3969/j.issn.1000-5382.2010.05.018
[24] 李树贤. 植物染色体与遗传育种[M]. 北京: 科学出版社, 2008. Li S X. Plant chromosome and genetic breeding[M]. Beijing: Science Press, 2008.
[25] Zhang X, Cao Q, Zhou P, et al. Meiotic chromosome behavior of the male-fertile allotriploid lily cultivar ‘Cocossa’[J]. Plant Cell Reports, 2017, 36(10): 1641−1653. doi: 10.1007/s00299-017-2180-6
[26] Hong D Y. Peonies of the world: taxonomy and phytogeography[M]. London: Royal Botanic Gardens, Kew, 2010.
[27] 刘淑敏, 王莲英, 秦魁杰, 等. 牡丹三倍体品种: 首案红[J]. 北京林学院学报, 1983(4):62−67. Liu S M, Wang L Y, Qin K J, et al. ‘Shou An Hong’, a triploid cultivar of tree peony (Paeonia suffruticosa)[J]. Journal of Beijing Forestry College, 1983(4): 62−67.
[28] 李懋学, 张斅方. 三倍体牡丹的细胞遗传学观察[J]. 遗传, 1982, 4(5):19−21. doi: 10.3321/j.issn:0253-9772.1982.05.009 Li M X, Zhang X F. Cytogenetic observation of triploid tree peony[J]. Hereditas, 1982, 4(5): 19−21. doi: 10.3321/j.issn:0253-9772.1982.05.009
[29] 赵娜. 牡丹远缘杂交后代及亲本形态与核型研究[D]. 北京: 北京林业大学, 2014. Zhao N. Study on morphology and chromosome karyotype of distant hybridization progenies and their parents of tree peony[D]. Beijing: Beijing Forestry University, 2014.
[30] Sang T, Pan J, Zhang D M, et al. Origins of polyploids: an example from peonies (Paeonia) and a model for angiosperms[J]. Biological Journal of the Linnean Society, 2004, 82: 561−571. doi: 10.1111/j.1095-8312.2004.00341.x
-
期刊类型引用(0)
其他类型引用(3)













计量
- 文章访问数: 3041
- HTML全文浏览量: 905
- PDF下载量: 92
- 被引次数: 3
