
Forest biomass characteristics and influencing factors in different restoration stages in the Daxing’anling forest region of Inner Mongolia, northern China
-
摘要:目的针对内蒙古大兴安岭林区中火烧、采伐干扰后退化森林生态系统恢复演替进程,研究其林分生物量特征,确定林分、地形和气候因子对林分生物量的影响,建立林分生物量通用模型,进行森林碳汇功能研究。方法利用野外实测的胸径和树高推算生物量。采用方差分析确定林分、地形和气候因子以及恢复时间、干扰类型等对林分生物量有无影响,使用一般线性模型建立林分生物量的多元线性模型,比较退化森林生态系统在不同生境下林分生物量特征。结果(1)不同恢复阶段的林分生物量差异明显,森林各器官的生物量分配有所不同,生物量最大比重多出现在4 ~ 8 cm、14 cm和18 ~ 26 cm径阶。(2)方差分析结果表明,恢复时间、干扰类型、林分优势树种、海拔对林分生物量恢复有显著性影响,而气候因子或气候因子的2个主成分对林分生物量的影响都不显著,2个林分生物量模型的确定系数在0.85以上,可以用来预测退化森林生态系统中的森林生物量。结论不同恢复阶段森林生物量有较大差异,森林乔木各器官生物量分配、径阶分配受到干扰类型和恢复时间影响。林分生物量与时间和海拔成正比,干扰方式与林型会影响林分生物量,研究结果可为大兴安岭林区森林生物量预测以及森林碳库研究提供借鉴。Abstract:ObjectiveTo study the succession process of degraded forest ecosystems recovered from the interference of fire and logging in the Inner Mongolia Daxing’anling forest area of northern China, a general stand biomass model was established to analyze the forest biomass characteristics and figure out the effects of stand, topography and climate factors on stand biomass.MethodBiomass was calculated from the published biomass equations based on corresponding diameter at breast (DBH) and tree height (H) measured in the field. Variance analysis was used to determine the significance of stand, topographic and climatic factors, restoration time and interference type to the stand biomass. General linear model method was used to establish multivariate linear model of stand forest biomass and the forest biomass characteristics of degraded forest ecosystem were compared among different habitats.Result(1) The forest biomass in different recovery stages of forest ecosystems was significantly different. The forest biomass organic allocation was also significantly different in a way that the maximum proportion of biomass mostly occurred in 4−8 cm, 14 cm and 18−26 cm diameter class. (2) Variance analysis showed that restoration time, interference type, dominant tree species and altitude had significant effects on stand biomass, while the two main climatic factors and climatic factors had no effect on stand biomass. The coefficient of determination for two stand biomass models was above 0.85 and could be used to predict forest biomass in degraded forest ecosystem.ConclusionThere are large differences in forest biomass in varied degraded stages of forest ecosystems. The forest biomass organic and diameter allocation are affected by interference type and restoration time. The stand biomass is proportional to restoration time and altitude. The interference type and forest type will affect the stand biomass. The research results can provide reference for forest biomass prediction and forest carbon pool research in Daxing’anling forest area.
-
Keywords:
- burned land /
- cutover land /
- restoration time /
- biomass /
- influencing factor
-
森林生态系统是最大的陆地生态系统,具有复杂的结构和多种功能,在维持生物多样性和调节全球碳平衡等方面具有不可替代的作用[1]。随着经济快速发展和森林资源的高强度开发利用以及自然灾害频繁发生而导致森林退化,是当前世界范围内所面临的一个主要的环境问题[2-3]。林火是影响森林生态系统的一个重要的非生物生态因子[4]。一方面,森林火灾可快速烧毁大片的森林,破坏森林生态系统的结构与功能,威胁到人类的生命和财产安全;另一方面,适度的火烧可以消耗森林可燃物,降低森林可燃性,起到改变森林生态系统结构的作用[5-6]。森林采伐是获取木材资源的生产作业,是森林经营的重要内容,森林采取可持续经营措施,适时将森林转化成木质林产品,有助于生态系统固碳[7],但采伐作业对森林生产力和森林生态系统的结构与功能有重大影响[8]。不论是自然干扰(林火等)还是人为干扰(采伐等),都会改变森林的年龄结构、树种组成、森林更新,最后发生演替,而遭受干扰后的退化森林生态系统中植被恢复是森林生态功能恢复的基础[9-11],有研究表明,在火烧迹地中不同林型的乔木生物量约占该生态系统生物量的0.6 ~ 0.8之间且变化稳定[12],说明乔木的更新状况对森林群落的演替趋势具有一定的指示作用。干扰会使森林郁闭度降低,产生林间空隙或形成林窗,改变林木的生长环境,进而影响整个生态系统[13]。
兴安岭林区是我国重要的森林生态系统,对区域森林生态系统碳平衡有巨大影响,其作用和地位是其他地区无法替代的[14]。该区既是火灾多发林区(据1987 年火警火灾登记报表显示,大兴安岭地区 1987 年共发生火警火灾46起),又是主要的木材产出地区,形成了许多火烧迹地和采伐迹地,因此加强迹地生态系统物种多样性保护和生态功能恢复具有重要意义。综上所述,本研究以大兴安岭内蒙古林区乌尔旗汗林业局不同恢复阶段的退化森林生态系统为研究对象,主要研究目的有两点:(1) 基于野外实测数据和相关生物量模型得到不同恢复阶段的森林生物量,了解火烧和采伐后森林生物量特征(总量特征、各组分分配特征、径阶分配特征与树种分配特征等);(2) 以林分因子为基础,结合地形因子、气象因子、恢复时间和干扰类型等作为退化森林生态系统的非生物环境因子,分析林分生物量的影响因素,建立林分生物量的多元线性回归模型,为大兴安岭森林碳汇功能研究及林业碳汇评价指标体系的建立提供理论基础。
1. 研究区概况与研究方法
1.1 研究区概况
研究区位于内蒙古自治区牙克石市乌尔旗汗林业局的莫拐林场、克里河林场和库口林场。乌尔旗汗镇地处大兴安岭西坡,地理坐标121°23′E、49°34′N,该区属于寒温带大陆性季风半湿润森林气候,年平均气温2.6 ℃,全年最高气温34.5 ℃,最低气温− 47.6 ℃,年均降雨量442 mm。由于特殊的气候条件,乌尔旗汗的森林资源十分丰富,总面积59.36万hm2,森林覆盖率为78.97%,林木绿化率为83.0%,主要树种有落叶松(Larix gmelinii)、白桦(Betula platyphylla)和山杨(Populus davidiana)等[15],其主要林型为:兴安落叶松−白桦丛林、白桦−兴安落叶松林、白桦−山杨林。
1.2 数 据
1.2.1 调查数据
2018年7月,在火烧干扰后恢复12、22和31年以及采伐干扰后恢复14、20和29年的森林生态系统中随机布设大小为20 m × 20 m的样地,研究区内无人为影响,所有植被均为自然恢复演替。火烧迹地按照样地内胸径大于 > 5 cm且有明显火烧痕迹的枯立木株数来划分火烧程度(分为轻度、中度和重度),不计后期自然恢复的植被由于其他原因造成的死亡,采伐迹地的采伐方式为皆伐。干扰类型共有4种(轻度火烧迹地、中度火烧迹地、重度火烧迹地和采伐迹地),每种类型有3个重复,共计36块样地。记录各样地的GPS坐标、海拔、坡度和坡向,实测各样地内乔木胸径和树高,本次调查样木共计2 692株,其中活立木2 319株,枯立木373株,本研究针对活立木,数据处理时剔除了每个样地的枯立木,需要说明的是:下文中提到的恢复时间(10、20、30年组)是整化的时间,计算时为实际恢复时间。
1.2.2 林分因子
由调查数据得到各样地的林分因子,详细计算公式见《测树学》教材[16],树种组成是利用样地内某一树种的胸高断面积占样地总断面积的百分比得到,并由此方法确定各样地的优势树种。
1.2.3 气候数据
气候数据来自ClimateAP [17],本研究利用样地GPS坐标和海拔,逐年提取样地的气候数据:包括年平均温度、年平均降水、年平均温度与年平均降水的比值[18]、月平均最高温、月平均最低温、近30年极端最低气温等,样地的气候因子取干扰发生时间至2016年期间的平均值。1987—2016年样地的年平均温度呈现先上升后下降趋势,在− 2.3 ℃左右波动,年平均降水呈现先下降后上升的趋势,在430 mm左右波动,二者变化趋势都不明显(图1)。
![]() 图 1 1987—2016年的年平均气温与年平均降水量的变化趋势Figure 1. Trends of mean annual temperature and precipitation in 1987−2016
图 1 1987—2016年的年平均气温与年平均降水量的变化趋势Figure 1. Trends of mean annual temperature and precipitation in 1987−20161.2.4 地形数据
坡向和坡度按照国家森林资源连续清查技术规定(2014)[19]划分等级,坡向划分标准为阴坡181° ~ 360°和0° ~ 90°,阳坡90° ~ 270°;坡度划分为平坡 < 5°,缓坡5° ~ 14°,斜坡15° ~ 24°,陡坡25° ~ 34°,急坡35° ~ 44°,险坡≥ 45°。样地地的平均坡度为7.5°,其中仅5个样地为斜坡,其余大都为平坡或缓坡,海拔在782 ~ 990 m之间,地势较为平坦。
1.3 研究方法
1.3.1 生物量计算
此次调查所涉及的树种有落叶松、白桦、山杨、云杉(Picea asperata)、柳树(Salix matsudana)、稠李(Padus racemosa)等,使用相关二元单木生物量模型推算生物量,包括地上和地下生物量,其中树干、树皮、树枝、树叶之和为地上生物量,地上与地下生物量之和为总生物量。落叶松、白桦、云杉以及山杨(杨树即将出版)等树种采用林业行业标准《生物量模型及碳计量参数》[20-22]中的二元生物量模型;柳树和稠李等树种按软阔类,采用《中国森林植被生物量和碳储量评估》中的全国生物量模型[23]。林分生物量(t/hm2)用累加法获得。
1.3.2 影响林分生物量的因素及通用模型
地下生物量可以根据根茎比得到,因此本研究选择林分地上生物量为因变量,采用多因素方差分析判定林分因子、气候因子、地形因子、恢复时间以及干扰类型对林分生物量的影响,使用逐步剔除结合变量显著性消除共线性,进而建立回归模型来表示不同恢复阶段林分生物量与各因子间的定量关系,用决定系数R2判断模型解释效果[24-28]。具体过程如下:先使用方差分析法得到所有变量的影响,然后删除最不显著的变量,重新将剩下的变量与林分生物量做方差分析,直至所有影响因子的P值小于0.05。由于气象变量较多,对气象因子做主成分分析,提取主成分,然后重复上述步骤。最后用所有显著的因子与林分生物量建立多元线性回归模型,其中定性变量使用哑变量方法求解参数,并用F检验判断不同模型间的显著性差异。
2. 结果与分析
2.1 不同恢复阶段的林分生物量特征
不同回复阶段林分生物量见表1。从总量上看:10年组迹地总生物量为(54.91 ± 63.74) t/hm2;20年组迹地总生物量为(115.15 ± 12.54) t/hm2,30年组迹地总生物量为(74.53 ± 16.79) t/hm2,这是由于采用时空互代法的原因,不同林分的立地质量有所不同,不同生境条件下,不同森林类型的更新方式以及不同树种间的生长快慢不同,若对同一类型的退化森林生态系统采用连续观测,则生物量与时间之间的关系应该满足“S”型曲线这一特征。从干扰类型上看,轻度火烧迹地的总生物量远远大于重度火烧迹地,采伐迹地的生物量则与重度火烧迹地接近,因为重度火烧过后,林分内的多数林木被烧死,保留下来的活立木数量很少,与皆伐样地的情形相似,进而两者的生物量较为接近,各组分生物量总体趋势为树干 > 地下 > 树枝 > 树皮 > 树叶。
表 1 不同阶段的森林林分生物量 (t·hm− 2)Table 1. Stand biomass of different restoration stages(t·ha− 1) 恢复时间/年
Restoration
time/year干扰类型
Interference
type树干生物量
Stem
biomass树皮生物量
Bark
biomass树枝生物量
Branch
biomass树叶生物量
Leaf
biomass地上生物量
Aboveground
biomass地下生物量
Root
biomass总生物量
Total
biomass12 轻度火烧
Mild fire78.54 11.36 23.44 5.46 118.80 41.63 160.43 中度火烧
Moderate fire23.70 3.45 7.72 1.90 36.77 13.56 50.33 重度火烧
Severe fire1.39 0.38 0.74 0.26 2.77 1.09 3.86 22 轻度火烧
Mild fire68.92 9.39 18.87 4.19 101.37 34.93 136.30 中度火烧
Moderate fire57.45 7.60 15.06 3.31 83.42 28.81 112.23 重度火烧
Severe fire54.82 7.34 14.65 3.28 80.09 27.80 107.89 31 轻度火烧
Mild fire42.72 8.70 22.81 4.26 78.49 24.09 102.58 中度火烧
Moderate fire30.71 6.48 13.56 3.28 54.03 17.64 71.67 重度火烧
Severe fire28.69 6.00 10.77 3.08 48.54 16.17 64.71 14 采伐 Cutover 1.76 0.51 0.97 0.34 3.58 1.44 5.02 20 采伐 Cutover 52.56 10.23 15.42 4.69 82.90 21.27 104.17 29 采伐 Cutover 26.45 5.65 9.79 2.90 44.79 14.38 59.17 2.2 生物量分配格局
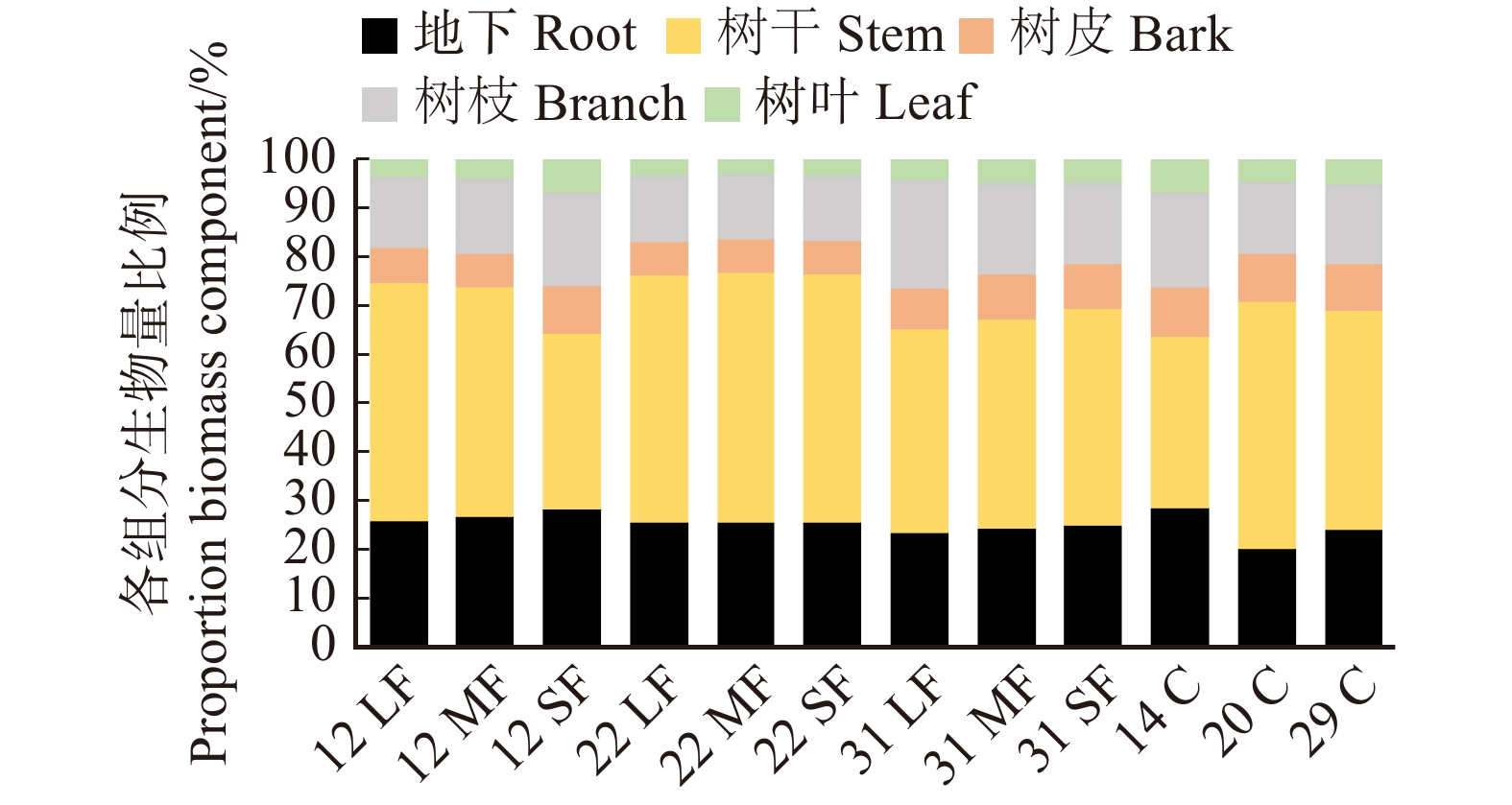
各器官的生物量及其所占比例见图2,树干和树根约占据了乔木层生物量的70%,树干的生物量最高,占45%以上,波动也最大,树根则占25%以上,其中14年采伐迹地的地下生物量占比最高(28.7%),树皮、树枝、树叶生物量的平均占比为8.4%、16.5%、4.4%,树叶生物量的占比最小。其中干扰类型对树皮(P < 0.01)和树叶(P < 0.01)的生物量分配有显著影响,而对树干(P = 0.35)、树枝(P = 0.87)、和地下(P = 0.31)生物量的分配没有显著影响。而恢复时间仅对地下(P < 0.01)生物量分配有显著影响,对于树干(P = 0.57)、树枝(P = 0.24)、树皮(P = 0.12)和树叶(P = 0.26)生物量的分配没有显著影响。可见,干扰类型和恢复时间影响各器官的生物量分配。
![]() 图 2 不同恢复阶段的森林林分生物量分配格局LF表示轻度火烧;MF表示中度火烧;SF表示重度火烧;C表示采伐。数字12、14、20、22、29、31表示恢复时间(年)。下同。LF, mild fire; MF, moderate fire; SF, severe fire; C, cutover. Number 12, 14, 20, 22, 29, 31 represent 12, 14, 20, 22, 29, 31 years restoration time, respectively. The same below.Figure 2. Stand biomass distribution pattern of different restoration stages
图 2 不同恢复阶段的森林林分生物量分配格局LF表示轻度火烧;MF表示中度火烧;SF表示重度火烧;C表示采伐。数字12、14、20、22、29、31表示恢复时间(年)。下同。LF, mild fire; MF, moderate fire; SF, severe fire; C, cutover. Number 12, 14, 20, 22, 29, 31 represent 12, 14, 20, 22, 29, 31 years restoration time, respectively. The same below.Figure 2. Stand biomass distribution pattern of different restoration stages2.3 生物量的径阶分配特征
按2 cm整化径阶,不同径阶的生物量占比见表2。生物量占比多在中间径阶达到峰值,而大径阶和小径阶的生物量占比较小。生物量占比较多的径阶有:4 ~ 8 cm、14 cm和18 ~ 26 cm,呈现中间径阶高,两头径阶低的特点。采伐迹地与重度火烧迹地的生物量径阶分配特征比较相似,轻度、中度火烧迹地的总生物量在径阶结构上的差异不大,不同阶段的森林总量的径阶分配有所不同。
表 2 不同阶段森林林分生物量的径阶分配Table 2. Forest biomass proportion in different diameter classes of varied restoration stages% 径阶
Diameter
class/cm12年
12 years22年
22 years31年
31 years14年
14 years20年
20 years29年
29 yearsLF MF SF LF MF SF LF MF SF C C C 2 — 0.48 6.1 — — — 0.09 0.52 0.75 9.04 0.36 1.58 4 0.14 0.48 21.77 0.08 — — 0.36 1.22 5.49 43.98 4.18 9.21 6 0.52 — 47.28 0.22 0.06 0.29 0.29 1.67 16.63 36.8 11.22 21.53 8 1.91 1.33 24.85 0.12 — 0.46 0.93 4.73 26.18 10.19 19.13 32.99 10 5.25 3.07 — 0.58 0.7 1.16 1.78 13.81 17.46 — 28.35 9.11 12 6.23 5 — 1.03 — 2.12 1.91 14.67 10.08 — 22.94 2.59 14 6.08 19.66 — 2.91 2.08 5.97 5.03 21.89 3.78 — 8.35 2.35 16 11.47 11.91 — 4.77 7.31 13.04 8.97 20.45 3.01 — 4.41 — 18 8.15 29.65 — 18.25 9.31 7.25 3.68 9.57 3.44 — 1.07 5.82 20 17.93 9.11 — 17.97 15.23 18.03 14.59 — 5.33 — — 5.09 22 10.38 19.31 — 16.07 10.86 8.77 11.79 8.75 3.34 — — 3.31 24 8.81 — — 22.55 16.43 12.86 12.62 2.72 — — — 6.42 26 8.76 — — 3.95 21.37 11.51 8.6 — 4.5 — — — 28 4.36 — — 9.28 11.94 — 7.12 — — — — — 30 5.27 — — 2.21 — 7.69 — — — — — — 32 — — — — — 3 4.53 — — — — — 34 — — — — — — — — — — — — 36 — — — — — 7.85 — — — — — — 38 4.69 — — — — — — — — — — — 40 — — — — — — 7.22 — — — — — 50 — — — — — — 10.5 — — — — — 2.4 生物量的树种分配特征
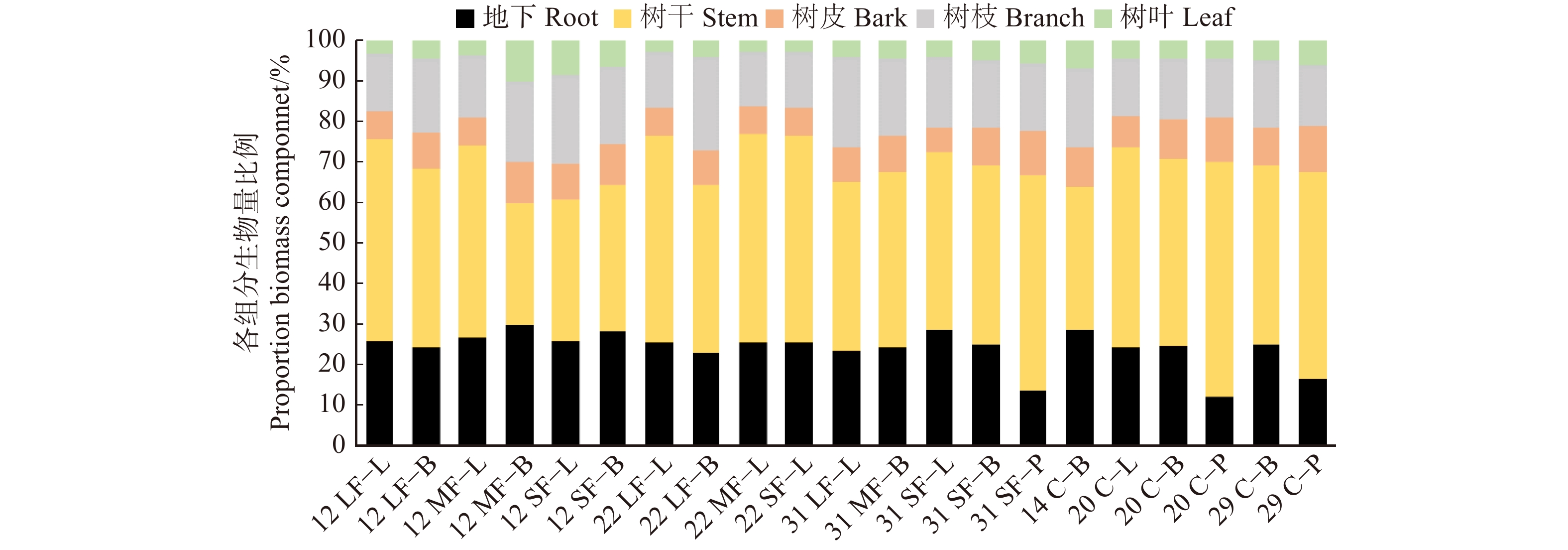
不同恢复阶段的迹地的林型不同,进而影响林分生物量的大小(图3),12年轻度和中度火烧迹地落叶松生物量分别占整个林分生物量的88%和99%,12年重度火烧迹地白桦生物量最大,占整个林分生物量94%;22年火烧迹地以落叶松生物量最大;31年火烧迹地只有白桦,其中轻度火烧迹地的白桦生物量最大,采伐迹地均以白桦为主要树种,但山杨的比例也不小(10%),落叶松生物量仅占整个林分的6%。图4是不同恢复阶段迹地的树种生物量器官分配,优势树种的生物量分配格局与林分总生物量的分配格局相似。树皮、树枝、树叶的生物量占比变化较小,标准偏差小于3%,而树干生物量和地下生物量比例则变化明显。
![]() 图 3 不同阶段的森林主要树种的生物量Figure 3. Biomass of main tree species in different restoration stages
图 3 不同阶段的森林主要树种的生物量Figure 3. Biomass of main tree species in different restoration stages![]() 图 4 不同恢复阶段森林的主要树种生物量分配格局L代表落叶松;B代表白桦;P代表山杨。L, Larix gmelinii; B, Betula platyphylla; P, Populus davidiana.Figure 4. Biomass distribution pattern of main forest species in different restoration stages
图 4 不同恢复阶段森林的主要树种生物量分配格局L代表落叶松;B代表白桦;P代表山杨。L, Larix gmelinii; B, Betula platyphylla; P, Populus davidiana.Figure 4. Biomass distribution pattern of main forest species in different restoration stages2.5 林分生物量与各因子间的关系及通用模型
对所有气象因子做主成分分析,提取了两个主成分,其中第1主成分的贡献率为73.3%,第2主成分的贡献率为24.2%,两个主成分的累积贡献率达97.5%,可以有效表示各气象因子所代表的信息。但不论是使用这两个主成分还是直接使用气候因子,得到的结果均与表3一致。由于林分生物量是单株生物量合计得到,所以株数密度乘以林分平均胸径的平方(
Nˉd2 )与林分生物量的关系十分密切,恢复时间、海拔、干扰类型、优势树种对林分生物量有显著影响。不同干扰类型下林分生物量恢复有显著性差异,而气候因子均对林分生物量的影响不显著,使用方差分析得到对林分生物量有显著影响的因子,建立林分生物量模型如下。表 3 林分生物量与各因子的方差分析Table 3. Variance analysis result of forest biomass with other factors影响因子 Influencing factor 平方和 Sum Sq. 自由度 df 均方 Mean Sq. F 值 F value 显著性 Sig. 恢复时间 Restoration time (T) 520.9 1 520.9 8.6 0.007 海拔 Altitude (AL) 887.7 1 887.7 14.7 0.001 干扰类型 Interference type (I) 2 920.0 3 973.3 16.1 < 0.001 优势树种 Dominant species (S) 454.7 2 227.3 3.8 0.036 变量 Variable (Nˉd2) 7 545.2 1 7 545.2 124.8 < 0.001 残差 Residual 1 633.0 27 60.5 模型 Model 48 002.0 8 6 000.2 99.2 < 0.001 M1:W=3.27E−4×Nˉd2+3.531 M2:W=2.97E−4×Nˉd2+0.941×T+0.167×AL−186.354+{26.502I=轻度火烧5.283I=中度火烧0I=重度火烧10.678I=采伐+{18.395S=落叶松林25.590S=桦树林0S=杨桦混交林 式中:W为林分地上生物量(t/hm2),N为密度(株/hm2),
ˉd 为林分平均胸径,T为恢复时间(年),AL为海拔(m),I为干扰类型,S为林分优势树种。M1的决定系数R2 = 0.856 1,只有2个变量,应用较为方便,增加了恢复时间、海拔等因子的M2的决定系数R2 = 0.957 4,表明二者都具有良好的拟合优度,模型差异性检验的F统计量为7.09 > 1.96,两个模型有显著性差异,M2可以进一步提升林分生物量的预测,可以很好的表明各因子对林分生物量的影响。从M2的结果看,林分生物量随恢复时间和海拔的增加而增加,林分生物量随火烧程度的增加而降低,采伐迹地的生物量恢复介于轻度火烧迹地和中度火烧迹地生物量恢复之间,不同林型的生物量也有差异,杨桦混交林的生物量最低,桦树林的生物量最高,而落叶松林则介于两者之间。
3. 讨 论
白桦是该地区的先锋树种,在原为针叶树种地区,迹地内均有白桦、山杨等出现。曾锋等[29]研究表明兴安落叶松天然更新的出苗频率在当年可达60%,范兆飞等[30]研究认为白桦具有很强的恢复力,白桦种源丰富,种子易随风漂流,快速侵入迹地并进行大量繁殖,进而影响林分结构,因此种源是决定天然更新的一个重要条件,它决定了迹地恢复的林型。本研究中出现30年组迹地生物量均值小于20年组均值的反常现象,可能是多方面原因造成的,既有样本数量的问题,也有林分类型的问题以及干扰前的森林状态问题,需要进一步地研究。从结果看,I 和 T 对各器官的生物量分配有影响,尤其是对树干生物量的影响较大,径阶的差异直接影响着生物量的大小与分布特征,生物量占比多在中间径阶达到一个峰值,但是不同迹地的生物量占比的峰值所在径阶并不一致,I 和T均影响林分生物量的径阶分配。这与李卓凡[31]的研究相比,除了各组分生物量的大小排序与本研究结果不同外,估计结果也相差数倍;而与王丽红等[32]的研究结果相比,地下部分较为接近,而地上部分差异较大。存在如此大差异有以下三方面的原因:一是样地大小有关,Brown等[33]研究结果表明:样方面积越小,不同样方之间的生物量误差越大。二是树高测量的误差导致生物量预测产生偏差。三是研究所采用的模型不同。
方差分析显示,恢复时间和海拔对林分生物量有显著影响,气候因子对林分生物量无显著影响。Condés等[34]研究林分生长时,发现加入年平均气温和年平均降水等气候因子,林分生长模型的R2仅从0.61变为0.65,气候对林分生物量的影响不大。产生这一现象的原因可能是样地尺度的气候因子差异较小,气候的空间变化表达不完全。王轶夫[35]的研究结果也认为森林生物量跟海拔、坡度和坡向之间存在明显的相关性,但是海拔与气候之间存在相关关系,所以海拔也代表了部分气候信息。
2个林分生物量模型中,M1决定系数为0.856,只有两个自变量,应用方便,M2决定系数达0.95以上,提升了生物量的预测精度。从M2拟合结果上看:生物量与 T 成正比,这一特征与部分研究结果相似。Taylor等[36]发现大火过后几年内,新的幼林恢复成森林;Reed等[37]的研究发现植被净生产力在火后第9年即可恢复至相同密度成熟林的生产力范围内。从 I 的影响看,林分生物量随着火烧程度的增加而降低,这与张玉红等[38]的研究结果一致。由于火烧迹地残存的大径级保留木,使林分生物量增加,而采伐迹地的所有林木均为小径级更新林木,导致林分生物量差异更大。郑丽凤等[39]研究表明,采伐强度越高,恢复所需要的年限越长,皆伐恢复较为困难。而不同林分类型生物量的累积能力有所不同,这与胡海清等[40]研究结果一致。He[41]与Fan[42]研究结果表明阔叶树的生物量大于针叶树,而有的研究却表明针叶林向针阔混交林转变时,碳储量平均增加可达16.8%[43],这与本研究中“杨桦混交林的生物量最低”这一结果并不一致。其原因一是研究的树种不同,本研究大多为阔叶混交林,不同林型的生物量恢复大小有所差异;二是林分生物量与生态因子作用机制尚不明确,还有待于深入研究;三是样本数量与样地大小的问题。
4. 结 论
本研究通过“空间代替时间”的方法对不同恢复年限和干扰类型的退化森林生态系统进行跟踪调查,并对其林分生物量特征进行分析。不同恢复阶段森林生物量差异较大,各器官的生物量分配、总生物量径阶分配以及不同树种的器官生物量分配与恢复时间和干扰类型有密切联系,而气候因子对林分生物量的影响并不显著。林分生物量模型表明,林分生物量随地形因子中的海拔的增加而增加,与恢复时间成正比,同时干扰类型和优势树种会影响林分生物量。本研究建立的林分生物量模型批判的入选各个因子对林分生物量的贡献大小和作用程度,研究结果可为大兴安岭林区长期森林碳库研究以及森林生物量预测提供借鉴。
-
![]()
图 1 1987—2016年的年平均气温与年平均降水量的变化趋势
Figure 1. Trends of mean annual temperature and precipitation in 1987−2016
![]()
图 2 不同恢复阶段的森林林分生物量分配格局
LF表示轻度火烧;MF表示中度火烧;SF表示重度火烧;C表示采伐。数字12、14、20、22、29、31表示恢复时间(年)。下同。LF, mild fire; MF, moderate fire; SF, severe fire; C, cutover. Number 12, 14, 20, 22, 29, 31 represent 12, 14, 20, 22, 29, 31 years restoration time, respectively. The same below.
Figure 2. Stand biomass distribution pattern of different restoration stages
![]()
图 3 不同阶段的森林主要树种的生物量
Figure 3. Biomass of main tree species in different restoration stages
![]()
图 4 不同恢复阶段森林的主要树种生物量分配格局
L代表落叶松;B代表白桦;P代表山杨。L, Larix gmelinii; B, Betula platyphylla; P, Populus davidiana.
Figure 4. Biomass distribution pattern of main forest species in different restoration stages
表 1 不同阶段的森林林分生物量 (t·hm− 2)
Table 1 Stand biomass of different restoration stages
(t·ha− 1) 恢复时间/年
Restoration
time/year干扰类型
Interference
type树干生物量
Stem
biomass树皮生物量
Bark
biomass树枝生物量
Branch
biomass树叶生物量
Leaf
biomass地上生物量
Aboveground
biomass地下生物量
Root
biomass总生物量
Total
biomass12 轻度火烧
Mild fire78.54 11.36 23.44 5.46 118.80 41.63 160.43 中度火烧
Moderate fire23.70 3.45 7.72 1.90 36.77 13.56 50.33 重度火烧
Severe fire1.39 0.38 0.74 0.26 2.77 1.09 3.86 22 轻度火烧
Mild fire68.92 9.39 18.87 4.19 101.37 34.93 136.30 中度火烧
Moderate fire57.45 7.60 15.06 3.31 83.42 28.81 112.23 重度火烧
Severe fire54.82 7.34 14.65 3.28 80.09 27.80 107.89 31 轻度火烧
Mild fire42.72 8.70 22.81 4.26 78.49 24.09 102.58 中度火烧
Moderate fire30.71 6.48 13.56 3.28 54.03 17.64 71.67 重度火烧
Severe fire28.69 6.00 10.77 3.08 48.54 16.17 64.71 14 采伐 Cutover 1.76 0.51 0.97 0.34 3.58 1.44 5.02 20 采伐 Cutover 52.56 10.23 15.42 4.69 82.90 21.27 104.17 29 采伐 Cutover 26.45 5.65 9.79 2.90 44.79 14.38 59.17  下载: 导出CSV
下载: 导出CSV
表 2 不同阶段森林林分生物量的径阶分配
Table 2 Forest biomass proportion in different diameter classes of varied restoration stages
% 径阶
Diameter
class/cm12年
12 years22年
22 years31年
31 years14年
14 years20年
20 years29年
29 yearsLF MF SF LF MF SF LF MF SF C C C 2 — 0.48 6.1 — — — 0.09 0.52 0.75 9.04 0.36 1.58 4 0.14 0.48 21.77 0.08 — — 0.36 1.22 5.49 43.98 4.18 9.21 6 0.52 — 47.28 0.22 0.06 0.29 0.29 1.67 16.63 36.8 11.22 21.53 8 1.91 1.33 24.85 0.12 — 0.46 0.93 4.73 26.18 10.19 19.13 32.99 10 5.25 3.07 — 0.58 0.7 1.16 1.78 13.81 17.46 — 28.35 9.11 12 6.23 5 — 1.03 — 2.12 1.91 14.67 10.08 — 22.94 2.59 14 6.08 19.66 — 2.91 2.08 5.97 5.03 21.89 3.78 — 8.35 2.35 16 11.47 11.91 — 4.77 7.31 13.04 8.97 20.45 3.01 — 4.41 — 18 8.15 29.65 — 18.25 9.31 7.25 3.68 9.57 3.44 — 1.07 5.82 20 17.93 9.11 — 17.97 15.23 18.03 14.59 — 5.33 — — 5.09 22 10.38 19.31 — 16.07 10.86 8.77 11.79 8.75 3.34 — — 3.31 24 8.81 — — 22.55 16.43 12.86 12.62 2.72 — — — 6.42 26 8.76 — — 3.95 21.37 11.51 8.6 — 4.5 — — — 28 4.36 — — 9.28 11.94 — 7.12 — — — — — 30 5.27 — — 2.21 — 7.69 — — — — — — 32 — — — — — 3 4.53 — — — — — 34 — — — — — — — — — — — — 36 — — — — — 7.85 — — — — — — 38 4.69 — — — — — — — — — — — 40 — — — — — — 7.22 — — — — — 50 — — — — — — 10.5 — — — — —
下载: 导出CSV
表 3 林分生物量与各因子的方差分析
Table 3 Variance analysis result of forest biomass with other factors
影响因子 Influencing factor 平方和 Sum Sq. 自由度 df 均方 Mean Sq. F 值 F value 显著性 Sig. 恢复时间 Restoration time (T) 520.9 1 520.9 8.6 0.007 海拔 Altitude (AL) 887.7 1 887.7 14.7 0.001 干扰类型 Interference type (I) 2 920.0 3 973.3 16.1 < 0.001 优势树种 Dominant species (S) 454.7 2 227.3 3.8 0.036 变量 Variable (Nˉd2) 7 545.2 1 7 545.2 124.8 < 0.001 残差 Residual 1 633.0 27 60.5 模型 Model 48 002.0 8 6 000.2 99.2 < 0.001
下载: 导出CSV
-
[1] Tang S Z, Liu S R. Conservation and sustainability of natural forests in China[J]. Review of China Agricultural Science and Technology, 2000, 2(1): 42−46.
[2] 夏自谦, 滕秀玲. 世界森林资源现状及前景展望[J]. 北京林业大学学报(社会科学版), 2003, 2(3):24−28. doi: 10.3969/j.issn.1671-6116.2003.03.006 Xia Z Q, Teng X L. Status quo and prospect of world forest resources[J]. Journal of Beijing Forestry University (Social Sciences), 2003, 2(3): 24−28. doi: 10.3969/j.issn.1671-6116.2003.03.006
[3] Dobson A P, Bradshaw A D, Baker A J M. Hopes for the future: restoration ecology and conservation biology[J]. Science, 1997, 277: 515−522. doi: 10.1126/science.277.5325.515
[4] 李俊清. 森林生态学[M]. 北京: 高等教育出版, 2010. Li J Q. Forest ecology[M]. Beijing: Higher Education Press, 2010.
[5] 徐化成. 中国大兴安岭森林[M]. 北京: 科学出版社, 1998. Xu H C. Daxing’anling forest, China[M]. Beijing: Science Press, 1998.
[6] 郑焕能, 温广玉. 林火灾变阈值[J]. 火灾科学, 1999, 8(3):1−5. Zheng H N, Wen G Y. Forest fire catastrophe and its threshold[J]. Fire Safety Science, 1999, 8(3): 1−5.
[7] Houghton R A. Converting terrestrial ecosystems from sources to sinks of carbon[J]. Arnbio, 1996, 25(4): 267−272.
[8] Hornbeck J W, Smith C T, Martin C W, et al. Effects of intensive harvesting on nutrient capitals of three forest types in New England[J]. Forest Ecology and Management, 1990, 30(1−4): 55−64. doi: 10.1016/0378-1127(90)90126-V
[9] 张厚华, 傅德志, 孙谷畴. 森林植被恢复重建的理论基础[J]. 北京林业大学学报, 2004, 26(1):97−99. doi: 10.3321/j.issn:1000-1522.2004.01.020 Zhang H H, Fu D Z, Sun G C. Theoretical fundamentals of restoration and reconstruction of degraded forest vegetation[J]. Journal of Beijing Forestry University, 2004, 26(1): 97−99. doi: 10.3321/j.issn:1000-1522.2004.01.020
[10] Amiro B. Net primary productivity following forest fire for Canadian ecoregions[J]. Canadian Journal of Forest Research, 2000, 30(30): 939−947.
[11] Wang C, Gower S T, Wang Y, et al. The influence of fire on carbon distribution and net primary production of boreal Larix gmelinii forests in northeastern China[J]. Global Change Biology, 2010, 7(6): 719−730.
[12] 丛燕, 魏荣华, 袁强. 黑河地区阔叶林和白桦落叶松林火烧迹地生物量动态变化的研究[J]. 林业科技, 2011, 36(2):17−19. doi: 10.3969/j.issn.1673-3290.2011.02.03 Cong Y, Wei R H, Yuan Q. Research of the biomass dynamics in burned areas of broad-leaved forest and birch and larch forest in Heihe Area[J]. Forestry Science & Technology, 2011, 36(2): 17−19. doi: 10.3969/j.issn.1673-3290.2011.02.03
[13] 解伏菊, 肖笃宁, 李秀珍, 等. 大兴安岭北坡火烧迹地森林景观恢复及其影响因子: 以郁闭度指标为例[J]. 应用生态学报, 2005, 16(9):1711−1718. doi: 10.3321/j.issn:1001-9332.2005.09.025 Xie F J, Xiao D N, Li X Z, et al. Forest landscape restoration and its affecting factors in burned area of northern Great Xing’an Mountains: taking forest coverage as an example[J]. Chinese Journal of Applied Ecology, 2005, 16(9): 1711−1718. doi: 10.3321/j.issn:1001-9332.2005.09.025
[14] 王晓莉, 常禹, 陈宏伟, 等. 黑龙江省大兴安岭主要森林生态系统生物量分配特征[J]. 生态学杂志, 2014, 33(6):1437−1444. Wang X L, Chang Y, Chen H, et al. Biomass allocation characteristics of the main forest ecosystems in the Great Xing’an Mountains, Heilongjiang Province[J]. Chinese Journal of Ecology, 2014, 33(6): 1437−1444.
[15] 孙伟迪. 乌尔旗汗林业局森林碳储量分布特征研究[J]. 内蒙古林业调查设计, 2017, 40(6):96−100, 102. Sun W D. Study on distribution characteristics of forest carbon storage in Wuerqihan Forestry Bureau[J]. Inner Mongolia Forestry Survey and Design, 2017, 40(6): 96−100, 102.
[16] 孟宪宇. 测树学[M]. 北京: 中国林业出版社, 2006. Meng X Y. Forest mensuration[M]. Beijing: China Forestry Publishing House, 2006.
[17] Wang T, Wang G, Innes J L, et al. ClimateAP: an application for dynamic local downscaling of historical and future climate data in Asia Pacific[J]. Frontiers of Agricultural Science and Engineering, 2017, 4(4): 448−458. doi: 10.15302/J-FASE-2017172
[18] Weiskittel A R, Crookston N L, Radtke P J. Linking climate, gross primary productivity, and site index across forests of the western United States[J]. Canadian Journal of Forest Research, 2011, 41(8): 1710−1721. doi: 10.1139/x11-086
[19] 国家林业局. 国家森林资源连续清查技术规定[S]. 北京: 国家林业局, 2014. State Forestry Administration. National forest resources continuous inventory technical regulations[S]. Beijing: State Forestry Administration, 2014.
[20] 国家林业局. LY/T 2655—2016立木生物量模型及碳计量参数: 云杉[S]. 北京: 中国标准出版社, 2017. State Forestry Administration. LY/T 2655—2016, Tree biomass models and related parameters to carbon accounting for Picea asperata[S]. Beijing: China Forestry Publishing House, 2017.
[21] 国家林业局. LY/T 2654—2016立木生物量模型及碳计量参数: 落叶松[S]. 北京: 中国标准出版社, 2017. State Forestry Administration. LY/T 2654—2016, Tree biomass models and related parameters to carbon accounting for Larix gmelinii[S]. Beijing: China Forestry Publishing House, 2017.
[22] 国家林业局. LY/T 2659—2016, 立木生物量模型及碳计量参数—桦树[S]. 北京: 中国标准出版社, 2017. State Forestry Administration. LY/T 2659—2016, Tree biomass models and related parameters to carbon accounting for Betula platyphylla[S]. Beijing: China Forestry Publishing House, 2017.
[23] 李海奎, 雷渊才. 中国森林植被生物量和碳储量评估[M]. 北京: 中国林业出版社, 2010. Li H K, Lei Y C. Estimation and evaluation of forest biomass carbon storage in China[M]. Beijing: China Forestry Publishing House, 2010.
[24] 唐守正, 郎奎建, 李海奎. 统计和生物数学模型计算: ForStat教程[M]. 北京: 科学出版社, 2009. Tang S Z, Lang K J, Li H K.Statistics and biomathematical models[M]. Beijing: Science Press, 2009.
[25] Kutner M H, Nachtsheim C J, Neter J, et al. Applied linear statistical models[M]. Boston: McGraw-Hill Irwin, 2005.
[26] Farrar D E, Glauber R R. Multicollinearity in regression analysis: the problem revisited[J]. Review of Economics and Statistics, 1967, 49(1): 92−107. doi: 10.2307/1937887
[27] Mcintyre S H, Montgomery D B, Srinivasan V, et al. Evaluating the statistical significance of models developed by stepwise regression[J]. Journal of Marketing Research, 1983, 20(1): 1−11. doi: 10.1177/002224378302000101
[28] Cornell J. Classical and modern regression with applications[J]. Technometrics, 1987, 29(3): 2.
[29] 曾锋, 张金池. 重金属在森林生态系统中的迁移规律研究进展[J]. 世界林业研究, 2001, 14(2):16−22. Zeng F, Zhang J C. Advances in the research on movement pattern of heavy metal ions in forest ecosystem[J]. World Forestry Research, 2001, 14(2): 16−22.
[30] 范兆飞, 徐化成, 于汝元. 大兴安岭北部兴安落叶松种群年龄结构及其与自然干扰关系的研究[J]. 林业科学, 1992, 28(1):2−11. Fan Z F, Xu H C, Yu R Y. A study on the species group age structure of Larix Gmelini population and its relation to disturbance on the North Daxing’anling Mountains[J]. Scientia Silvae Sinicae, 1992, 28(1): 2−11.
[31] 李卓凡. 兴安落叶松林生物量与碳储量的研究[D]. 呼和浩特: 内蒙古农业大学, 2013. Li Z F. Biomass and carbon storage of the Larix gmelinii forest’s research[D]. Hohhot: Inner Mongolia Agricultural University, 2013.
[32] 王丽红, 辛颖, 邹梦玲, 等. 大兴安岭火烧迹地植被恢复中植物多样性与生物量分配格局[J]. 北京林业大学学报, 2015, 37(12):41−47. Wang L H, Xin Y, Zou M L, et al. Plants diversity and biomass distribution of vegetation restoration in burned area of Great Xing’an Mountains[J]. Journal of Beijing Forestry University, 2015, 37(12): 41−47.
[33] Brown I F, Martinelli L A, Thomas W W, et al. Uncertainty in the biomass of Amazonian forests: an example from Rondonia, Brazil[J]. Forest Ecology and Management, 1995, 75(1−3): 175−189.
[34] Condés S, Roberedo F G. An empirical mixed model to quantify climate influence on the growth of Pinus halepeensis Mill. stands in southeastern Spain[J]. Forest Ecology and Management, 2012, 284(15): 59−68.
[35] 王轶夫. 基于神经网络的森林生物量估测模型研究[D]. 北京: 北京林业大学, 2013. Wang Y F. The study of forest biomass estimation model based on neural network[D]. Beijing: Beijing Forestry University, 2013.
[36] Taylor K D. Fire history of a sequoia-mixed conifer forest[J]. Ecology, 1979, 60(1): 129−142. doi: 10.2307/1936475
[37] Reed R A, Finley M E, Romme W H, et al. Above-ground net primary production and leaf area index in early postfire vegetation in Yellowstone National Park[J]. Ecosystems, 1999, 2(1): 88−94. doi: 10.1007/s100219900061
[38] 张玉红, 覃炳醒, 孙铭隆, 等. 林火对大兴安岭典型林型林下植被与土壤的影响[J]. 北京林业大学学报, 2012, 34(2):7−13. Zhang Y H, Qin B X, Sun M L, et al. Impact of forest fire on understory vegetation and soil in typical forest types of Daxing’an Mountains, northeastern China[J]. Journal of Beijing Forestry University, 2012, 34(2): 7−13.
[39] 郑丽凤, 周新年, 李丹, 等. 森林采伐对闽北天然次生林碳储量及其动态的影响[J]. 安全与环境学报, 2013, 13(6):162−167. Zheng L F, Zhou X N, Li D, et al. Impact of harvesting intensity of the natural secondary forestry on the carbon stocks reduction in northern Fujian[J]. Journal of Safety and Environment, 2013, 13(6): 162−167.
[40] 胡海清, 罗碧珍, 魏书精, 等. 小兴安岭7种典型林型林分生物量碳密度与固碳能力[J]. 植物生态学报, 2015, 39(2):140−158. doi: 10.17521/cjpe.2015.0014 Hu H Q, Luo B Z, Wei S J, et al. Biomass carbon density and carbon sequestration capacity in seven typical forest types of the Xiaoxing ’an Mountains, China[J]. Chinese Journal of Plant Ecology, 2015, 39(2): 140−158. doi: 10.17521/cjpe.2015.0014
[41] He Y J, Qin L, LI Z Y, et al. Carbon storage capacity of monoculture and mixed-species plantation in subtropical China[J]. Forest Ecology and Management, 2013, 295(1): 193−198.
[42] Fan H B, Liu W F, Wu J P, et al. Ecosystem carbon pools in mixed stands of hardwood species and Masson pine[J]. Journal of Tropical Forest Science, 2013, 25(2): 154−165.
[43] 李江, 陈宏伟, 冯弦. 云南热区几种阔叶人工林C储量的研究[J]. 广西植物, 2003, 23(4):294−298. Li J, Chen H W, Feng X. Carbon stock and rate of carbon sequestration assessment of hardwood plantations in tropical Yunnan, China[J]. Guihaia, 2003, 23(4): 294−298.
-
期刊类型引用(3)
1. 刘子洋,强波,张浩,符利勇,郭晋平. 气候和立地等级对落叶松林分生物量估计的影响. 北京林业大学学报. 2025(01): 22-28 .  本站查看
本站查看
2. 何潇,李海奎,张逸如,黄金金. 天然次生林碳储量生长模型与固碳能力驱动力研究. 北京林业大学学报. 2023(01): 1-10 . 本站查看
3. 何潇,雷相东,段光爽,丰庆荣,张逸如,冯林艳. 气候变化对落叶松人工林生物量生长的影响模拟. 南京林业大学学报(自然科学版). 2023(03): 120-128 . 百度学术
其他类型引用(4)

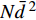
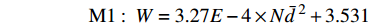


计量
- 文章访问数: 2087
- HTML全文浏览量: 613
- PDF下载量: 101
- 被引次数: 7
