
Effects of different thinning patterns on the spatial structure of Quercus mongolica secondary forests
-
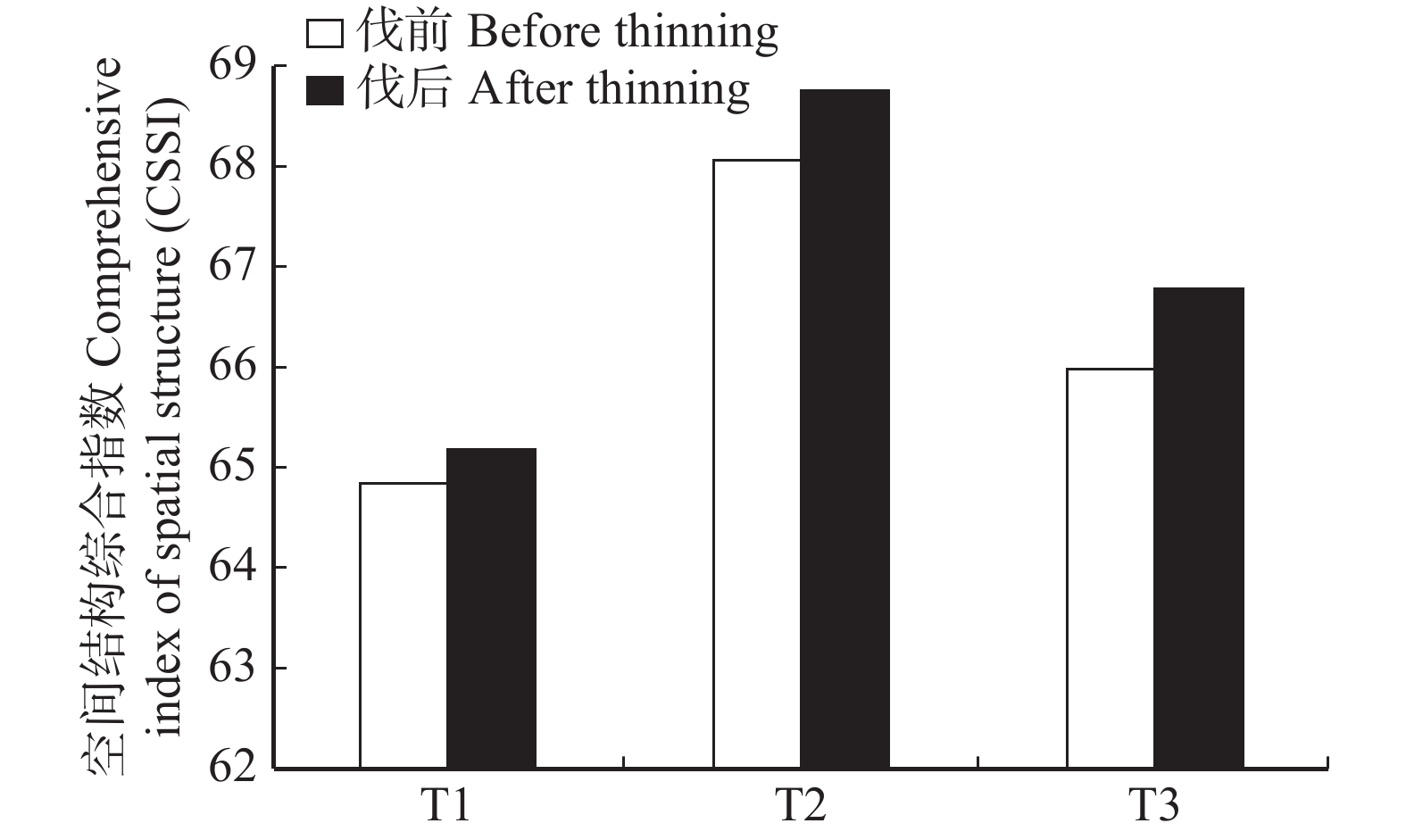
摘要:目的以长白山林区蒙古栎次生林为研究对象,分析基于目标树经营与传统经营的抚育间伐前后蒙古栎次生林空间结构主要指标及空间结构综合指数变化,评价不同抚育间伐方式对森林空间结构的影响。方法在吉林汪清设置9块面积均为1 hm2的蒙古栎次生林样地,随机区组进行3种处理,分别为传统经营(T1)、目标树经营密度1(T2)和目标树经营密度2(T3)。利用混交度、角尺度、大小比数和密集度4个林分空间结构指标,分析间伐前后目标树及林分的空间结构变化,并利用4个指标构建空间结构综合指数(CSSI)来综合评价蒙古栎次生林的空间结构对不同抚育间伐的响应。结果结果表明,从干扰树间伐对目标树的影响来看,目标树的混交度、大小比数和密集度3个指标的改善明显,角尺度改善不明显;从不同抚育间伐方式对林分空间结构的影响来看,3种处理均提高了林分内种间隔离程度、降低了密集程度,使林分空间分布格局趋向于随机分布,对林木大小分化度的影响不明显;根据CSSI综合评价得知:3种处理均提高了林分的空间结构综合指数,提高幅度为T3 > T2 > T1。结论干扰树间伐优化了目标树的空间结构,3种处理的抚育间伐均优化了林分的空间结构。基于目标树经营的抚育间伐对空间结构的改善程度优于传统的抚育间伐,T3处理,也即目标树选择密度为100株/hm2时采取的抚育间伐最有利于目标树和林分的空间结构优化。Abstract:ObjectiveThe effects of different thinning patterns on forest spatial structure were evaluated by analyzing the changes of main indicators of spatial structure and comprehensive spatial structure index of Quercus mongolica secondary forests (oak forest) before and after thinning.MethodNine permanent plots with an area of 1 ha were established in the oak forest in Wangqing, Jilin Province of northeastern China. Three kinds of treatments were carried out by random grouping, namely traditional management (T1), crop tree management with density 1 (T2) and crop tree management with density 2 (T3). The spatial structure variations of the crop trees and forest stand before and after the cutting operation were analyzed by mingling degree, uniform angle index, neighborhood comparison and crowding degree. The above four indicators were used to construct the comprehensive spatial structure index (CSSI) to comprehensively evaluate the response of oak secondary forest to the management measures.ResultThe results showed that the mean mingling degree, neighborhood comparison and crowding degree of the crop trees in T2 and T3 were improved after competitors cutting, while the change of mean uniform angle index was not obvious. From the perspective of the effects of different measures on the stand spatial structure, the mean mingling degree, uniform angle index and crowding degree of the target stands were all improved after cutting, while the neighborhood comparison variation was not pronounced. CSSI of all stands was increased by T3 > T2 > T1.ConclusionThe spatial structure of crop trees was optimized by competitor cutting, and the spatial structure of all stands was optimized by 3 kinds of thinning. The improvement of the spatial structure of the thinning based on the crop tree management is better than the traditional thinning. T3 is most beneficial to the optimization of the spatial structure of the stand.
-
在快速城镇化发展进程中,人类较大程度地改变了自然地形地貌,以不透水地面铺砌代替原有透水土壤和植被,造成下渗量与蒸发量的显著减少,从而引起同强度暴雨形成的地表径流量增大,峰值流量增加等现象[1]。为解决城市内涝等问题,“海绵城市”和低影响开发系统(Low impact development, LID)概念应运而生。目前对低影响开发系统的研究多数集中于市政道路[2]、住宅小区[3-4]、公园绿地[5-6]以及城市绿道[7]中绿地的渗透滞留能力,对其后续储蓄利用的量化研究较少。为解决上述问题,本文以位于北京市的首钢西十为研究对象,结合景观需求建设渗透滞留与储蓄利用相结合的低影响开发系统。
1. 研究区域概况
北京市位于华北平原北部,为典型的北温带半湿润大陆性季风气候,年平均降雨量(1950—2012年期间)为585.0 mm,80%降雨量集中于汛期(6—9月)[8],对城市内涝造成极大压力。本文选取首钢北区西北角的西十片区作为研究区域,西十内部及周边场地硬质化程度高达65.3%,现状下垫面对自然雨水下渗疏解能力非常有限,具有典型特征。
西十地块原为首钢料场,运输线两侧土壤含铁较高,研究表明土壤污染深度为0.5~2.0 m[15]。改造过程中将表层以下2 m的土壤替换为壤土,渗透系数为3.5×10-6 m/s[9]。
如图 1、图 2所示,西十地块四面环街,占地4.24 hm2,地势整体呈西北高东南低,内部有筒仓、转运站、料仓、泵站等多组建筑体,总占地面积约1.5 hm2。场地周边区域(包括秀池东路、秀池北街两条道路和筒仓办公楼、北七筒建筑遗址、停车场、N3-17办公楼、武警用房、员工宿舍、干法除尘等建筑及其外环境,共5.8 hm2)地势略低于西十地块,东侧和北侧有多组由厂房改建的配套服务建筑,强降雨时存在较为严重的内涝问题亟待解决,因此将其与西十地块一同作为本文的研究对象,称为外源雨水地块。
2. 低影响开发系统空间布局设计
2.1 设计目标与原则
根据《新首钢高端产业综合服务区控制性详细规划》要求,西十地块及外源雨水地块低影响开发系统雨水消纳能力应达到北京市3年一遇24 h降雨量(108 mm[9])的标准。设计在满足功能需求和景观效果的前提下,制定合理的低影响开发系统设计策略,在有限的绿地空间中因地制宜地叠加低影响开发设施层以应对降雨后地表径流产生速度加快、峰值流量加大、峰现时间提前等问题,以达到上位规划对径流消纳的要求,同时考虑将收集的地表径流用于后期灌溉。
2.2 设计策略
根据《海绵城市建设技术指南》低影响开发主要通过“源头削减、中途传输、末端调蓄”等多种手段[10]。西十地块及外源雨水场地提出的设计策略为:根据场地竖向、土地利用等条件优先构建渗透滞留系统,在不具备条件的情况下考虑将径流收集再利用,从源头、中途、末端3个方面进行低影响开发系统设计(见图 3)。
2.2.1 西十地块低影响开发系统
西十地块以建立渗透滞留系统为目标,针对“源头削减”本研究提出在满足功能使用的前提下,尽可能多地利用透水铺装、建设绿色屋顶、增加绿地面积,从源头有效削减地表径流。此次设计的“中途传输”途径为植草沟,沿硬质边缘形成线性景观带,达到收集、传输、滞留及下渗径流的目的。“末端调蓄”设施有下沉式绿地、生物滞留池两种。植草沟内的雨水排入末端调蓄设施内,雨水在此进行滞留下渗,当雨量过大时末端调蓄设施内的雨水通过溢流口排入市政雨水管网。
2.2.2 外源雨水地块低影响开发系统
外源雨水地块绿地面积较小(占总面积的28.3%),不适宜做渗透滞留设施,且建筑密度较大、人群活动量大,雨水应尽快排出场地,因此将建立储存利用系统作为设计策略。“末端调蓄”设施只有雨水调蓄池,调蓄池内收集的雨水用于西十地块浇灌。
2.3 设计内容
综上所述,西十地块及外源雨水地块低影响开发系统分为渗透滞留和储蓄利用两部部分。渗透滞留系统包含种植屋面、透水铺装、下凹式绿地、植草沟、生物滞留池5类LID设施;储蓄利用系统以雨水调蓄池为储蓄设施收集地表径流。低影响系统设计见图 4,各类LID设施设计要点见表 1,各类用地面积及用地比例见表 2和表 3。
表 1 各类LID设施设计要点Table 1. Design points of LID facilities区域Area 低影响开发设施LID facility 设计要点Design point 西十地块West 10 Plot 种植屋面Planting roof 将具备条件的平屋顶(6 495.1 m2)改建为可上人的屋顶花园,为办公人员提供多样活动空间的同时,在急降雨时可滞存一部分雨水以达到推迟峰现时间的目的The flat roof (6 495.1 m2) which can be converted into a roof garden provides office workers with a variety of activity spaces and retains rainwater in case of emergency rainfall to delay the peak time 透水铺装Permeable pavement 将场地中工业废弃物加工成透水混凝土砖,应用于地块内主园路(6 647.8 m2)Permeable concrete bricks which are processed from industrial wastes in the site applied to the main garden road (6 647.8 m2) in the plot 植草沟Vegetative swale 沿硬质边缘形成线性景观带(425.4 m2),以接纳铺装及绿地上的地表径流,将径流输送至雨水花园内部A linear landscape belt (425.4 m2) is formed along the hard edge to accept the surface runoff from pavement and green space and transport the runoff to the rain garden 下凹式绿地Rain garden 选择地块内部地势较低的区域集中布置两处下凹绿地(1 090.5 m2),一处位于建筑下沉庭院内,可就近接纳周边绿地及铺装上的地表径流;另一处利用下洼料场基址设计成90余米长的带状雨水花园,其为地表终端LID设施,接纳场地内全部径流Two concave green spaces (1 090.5 m2) are arranged centrally in the low-lying area inside the plot. One is located in the sunken courtyard of the building, which can receive the runoff of surrounding green space and pavement. Another site is designed as a belt rain garden more than 90 m long, which is the surface terminal LID facility and accepts all runoff in the site 生物滞留池Bio-retention cell 在人群活动量相对较小的地势低洼区域布置了1 035.8 m2的生物滞留池,当强降雨发生后,生物滞留池中一定时间内的雨水积蓄不影响场地的正常使用In low-lying areas with relatively small amount of population activity, 1 035.8 m2 of bio-retention cell was arranged. When heavy rainfall occurred, rainwater accumulation in the bio-retention cell within a certain period of time did not affect the normal use of the site 外源雨水地块Exogenous rainwater area 透水铺装Permeable pavement 将场地中工业废弃物加工成透水混凝土砖,应用于秀池北街、秀池东路人行道(7 422.0 m2)Permeable concrete bricks which are processed from industrial wastes in the site applied to the sidewalks of Xiuchi North Street and Xiuchi East Road (7 422.0 m2) 植草沟Vegetative swale 沿硬质边缘形成线性景观带(443.1 m2),以接纳铺装及绿地上的地表径流,将径流输送至雨水调蓄池中A linear landscape belt (443.1 m2) is formed along the hard edge to accept the surface runoff from pavement and green space and transport the runoff to the rain retention barrel 雨水调蓄池Rain retention barrel 外源雨水场地低处设置有雨水调蓄池,以接纳建筑屋顶雨水及全部地表径流。以设计目标为根据进行计算,外源雨水地块雨水调蓄池总容量为4 189 m3,方可达到消纳北京市3年重现期24 h降雨量这一目标。雨水调蓄池内雨水用于西十地块植物灌溉,并设有溢水口,当降雨强度超过地块内部LID设施消纳能力时,雨水通过溢水口排入市政雨水管网A rain retention barrel is installed at the lower level of the exogenous rainwater area to receive rainwater from the roof of the building and all surface runoff. According to the calculation based on the design objective, the total capacity of rain retention barrel in the exogenous rainwater area was 4 189 m3, which can reach the goal of absorbing 24-hour rainfall in the 3-year recurrence period of Beijing. Rainwater in the rain retention barrel is used for plant irrigation in the west 10 plot, and there is an overflow outlet. When the rainfall intensity exceeds the capacity of LID facilities inside the block, rainwater was discharged into the municipal rainwater pipe network through the overflow outlet 表 2 西十地块用地面积及用地比例表Table 2. Table of West 10 Plot of land area and land use ratio用地类型Type of land use 项目类型Item type 面积Area/m2 总面积Total area/m2 子项比例Subitem proportion/% 比例Proportion/% 绿化用地Green space 普通绿地Common green space 15 737.9 18 371.8 37.1 43.4 植草沟Vegetative swale 425.4 1.1 生物滞留池Bio-retention cell 1 035.8 2.4 下沉式绿地Rain garden 1 172.7 2.8 建筑占地Building space 屋顶花园绿地Green space of roof garden 4 871.3 14 987.3 11.5 35.7 屋顶花园铺装Impermeable pavement of roof garden 1 623.8 3.8 不上人硬质屋顶Hard roof of no people 8 492.2 20.4 园路及铺装场地用地Path and square 透水铺装Permeable pavement 6 647.8 8 954.4 15.6 20.8 不透水铺装Impermeable pavement 2 306.6 5.2 水体Water space 景观水体(不具备调蓄功能) Landscape water space (no regulating function) 52.3 52.3 0.1 0.1 合计Total 42 365.8 100 表 3 外源雨水地块用地面积及用地比例表Table 3. Table of exogenous site of land area and land use ratio用地类型Type of land use 项目类型Item type 面积Area/m2 总面积Total area/m2 子项比例Subitem proportion/% 比例Proportion/% 绿化用地Green space 普通绿地Common green space 15 967.6 16 410.7 27.5 28.3 植草沟Vegetative swale 443.1 0.8 建筑占地Building space 不上人硬质屋顶Hard roof of no people 16 865.1 16 865.1 29.1 29.1 园路及铺装场地用地Path and square 透水铺装Permeable pavement 7 422.0 24 740.1 12.8 42.6 不透水铺装Impermeable pavement 17 318.1 29.8 雨水调蓄池(地下) Rain retention barrel 雨水调蓄池(地下) Rain retention barrel (underground) 4 189 4 189 合计Total 58 015.9 100 3. 研究方法
按照《新首钢高端产业综合服务区控制性详细规划》要求完成西十地块及外源雨水地块低影响开发系统设计,并对上述设计方案进行SWMM模型构建。构建西十地块与外源雨水地块两套体系,西十地块体系用于对比传统模型和低影响开发模型,以分析低影响开发系统的雨洪调控效果;外源雨水地块体系用于对比雨水调蓄池蓄水量与植物灌溉需水量,以分析两者之间的数量关系。
3.1 渗透滞留系统模型建立
通过SWMM构建低影响开发模型和传统开发模型,对比两者之间数据以分析首钢西十低影响开发系统对区域径流总量、峰值流量和峰现时间的调控效果。
3.1.1 低影响开发模型构建
研究区域为西十地块及外源雨水地块,主要包含3类下垫面——屋顶、铺装、绿地。其中:不透水下垫面有硬质屋顶、不透水铺装;透水下垫面有绿色屋顶、透水铺装、绿地。根据上述低影响开发系统设计策略,遵循概化原则,将西十地块及外源雨水地块共划分为21个子汇水单元,末端出水口为2个。
3.1.2 传统开发模型构建
传统开发模式为通过竖向设计将绿地及屋顶径流排到铺装或道路上,再通过雨排水管线排入市政管网(图 5)。为方便后期对比分析,传统开发模型构建时绿地及铺装面积与低影响开发模型中一致(表 4)。雨排水管线图见图 6,SWMM模型概化图见图 7。
![]() 图 6 首钢西十传统雨水系统平面图Figure 6. Plan of traditional storm-water system in Shougang West 10 Plot
图 6 首钢西十传统雨水系统平面图Figure 6. Plan of traditional storm-water system in Shougang West 10 Plot![]() 图 7 首钢西十传统雨水系统概化图Figure 7. Generalized map of traditional storm-water system in Shougang West 10 Plot表 4 传统开发模式用地面积及用地比例表Table 4. Table of land area and land use ratio in traditional development model
图 7 首钢西十传统雨水系统概化图Figure 7. Generalized map of traditional storm-water system in Shougang West 10 Plot表 4 传统开发模式用地面积及用地比例表Table 4. Table of land area and land use ratio in traditional development model项目类型Item type 面积Area/m2 比例Proportion/% 绿化用地Green space 34 782.5 34.7 建筑占地Building space 31 852.4 31.7 园路及铺装场地用地Path and square 33 694.5 33.5 水体Water space 52.3 0.1 雨水调蓄池(地下) Rain retention barrel (underground) 4 189 合计Total 100 381.7 100 3.1.3 模块参数设置
本研究在推求各子汇水区的产流过程时,选用霍顿(Horton)公式进行下渗计算。根据方案设计对不同下垫面进行设定,各项参数参考SWMM用户手册[12]中的推荐值及相关文献。汇水模型为非线性水库模型,水力模型选用动力波模型[11]。根据场地土壤特性,入渗率为12.6 mm/h,衰弱常数为4 h-1。透水和不透水地表糙率系数分别为0.15和0.011,洼蓄量均为3 mm;植草沟宽度为2 000 mm,深度为500 mm;下沉式绿地平均深度为800 mm;生物滞留池平均深度为1 000 mm。
3.2 储蓄利用系统模型建立
根据北京市2017年全年降雨量日值统计(http://www.meteoblue.com/en/weather/archive/export/beijing-capital-international-airport_china_6301354),通过SWMM模拟得出西十地块及外源雨水场地每日径流总量及峰值流量;整合数据得到每周雨水调蓄池内收集的径流总量,同计算得出的浇灌需水量进行对比,以周为单位分析外源雨水地块收集的径流与西十地块的浇灌需求之间的关系。
4. 结果与分析
4.1 渗透滞留系统结果与分析
4.1.1 设计降雨选择
SWMM模型可使用的降雨数据为两种类型:(1)实测历时降雨资料;(2)设计降雨[13]。本研究根据北京《雨水控制与利用工程设计规范》(DB11/685—2013)北京地区1年一遇、2年一遇、3年一遇、5年一遇、10年一遇、20年一遇重现期降雨量分别为45、81、108、141、209、270 mm,降雨历时为24 h,模拟时间48 h。
4.1.2 结果与分析
渗透滞留系统在重现期为1、2、3、5、10、20年的24 h降雨情况下,西十地块和外源雨水地块的低影响开发模型与传统开发模型的径流总量、峰值径流量和峰现时间分别见表 5和表 6;西十地块出口径流曲线对比图见图 8~13;外源雨水地块雨水调蓄池内部容积变化曲线对比图见图 14~19。根据上述数据结构可得出以下结果:
表 5 西十地块不同降雨强度下的出水口径流量模拟结果Table 5. Simulated results of West 10 Plot of the runoff under different rainfall intensities模拟项目Simulation item 1年一遇(45 mm)24 h 1-year-once(45 mm)24 hour 2年一遇(81 mm)24 h 2-year-once(81 mm)24 hour 3年一遇(108mm)24 h 3-year-once(108 mm)24 hour 5年一遇(141 mm)24 h 5-year-once(141 mm)24 hour 10年一遇(209 mm)24 h 10-year-once(209 mm)24 hour 20年一遇(270 mm)24 h 20-year-once(270 mm)24 hour 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 径流总量Total runoff/m3 0 1 189 1 189 0 2 411 2 411 0 3 490 3 490 1 028 4 846 3 818 3 818 7 699 3 881 6 712 10 284 3 572 峰值流量Peak flow/(m3·s-1) 0 0.027 0.027 0 0.065 0.065 0 0.092 0.092 0.037 0.124 0.087 0.128 0.192 0.064 0.203 0.252 0.049 峰现时间Time of peak flow occurrence 消失Disappea- rance 09:01 消失Disappea- rance 09:00 消失Disappea- rance 09:00 15:15 09:00 375 min 14:10 09:00 310 min 13:00 09:00 240 min 表 6 外源雨水地块不同降雨强度下的出水口径流量模拟结果Table 6. Simulated results of exogenous site of the runoff under different rainfall intensities模拟项目Simulation item 1年一遇(45 mm)24 h 1-year-once(45 mm)24 hour 2年一遇(81 mm)24 h 2-year-once(81 mm)24 hour 3年一遇(108 mm)24 h 3-year-once(108 mm)24 hour 5年一遇(141 mm)24 h 5-year-once(141 mm)24 hour 10年一遇(209 mm)24 h 10-year-once(209 mm)24 hour 20年一遇(270 mm)24 h 20-year-once(270 mm)24 hour 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 低影响开发模式LID model 传统开发模式Traditional development model 差值Difference 径流总量Total runoff/m3 800 1 806 1 006 2 633 3 599 966 4 189 5 156 967 6 145 7 108 963 10 249 11 205 956 13 970 14 912 942 峰值流量Peak flow/(m3·s-1) 0.026 0.040 0.014 0.079 0.093 0.014 0.116 0.132 0.016 0.162 0.179 0.017 0.256 0.275 0.019 0.341 0.362 0.021 峰现时间Time of peak flow occurrence 12:20 09:01 119 min 10:45 09:00 105 min 10:15 09:00 75 min 10:10 09:00 70 min 10:00 09:00 60 min 09:55 08:58 57 min ![]() 图 8 西十地块1年一遇24小时降雨出口径流曲线Figure 8. Runoff curve of 1-year-once 24 hour rainfall event in West 10 Plot
图 8 西十地块1年一遇24小时降雨出口径流曲线Figure 8. Runoff curve of 1-year-once 24 hour rainfall event in West 10 Plot![]() 图 9 西十地块2年一遇24小时降雨出口径流曲线Figure 9. Runoff curve of 2-year-once 24 hour rainfall event in West 10 Plot
图 9 西十地块2年一遇24小时降雨出口径流曲线Figure 9. Runoff curve of 2-year-once 24 hour rainfall event in West 10 Plot![]() 图 10 西十地块3年一遇24小时降雨出口径流曲线Figure 10. Runoff curve of 3-year-once 24 hour rainfall event in West 10 Plot
图 10 西十地块3年一遇24小时降雨出口径流曲线Figure 10. Runoff curve of 3-year-once 24 hour rainfall event in West 10 Plot![]() 图 11 西十地块5年一遇24小时降雨出口径流曲线Figure 11. Runoff curve of 5-year-once 24 hour rainfall event in West 10 Plot
图 11 西十地块5年一遇24小时降雨出口径流曲线Figure 11. Runoff curve of 5-year-once 24 hour rainfall event in West 10 Plot![]() 图 12 西十地块10年一遇24小时降雨出口径流曲线Figure 12. Runoff curve of 10-year-once 24 hour rainfall event in West 10 Plot
图 12 西十地块10年一遇24小时降雨出口径流曲线Figure 12. Runoff curve of 10-year-once 24 hour rainfall event in West 10 Plot![]() 图 13 西十地块20年一遇24小时降雨出口径流曲线Figure 13. Runoff curve of 20-year-once 24 hour rainfall event in West 10 Plot
图 13 西十地块20年一遇24小时降雨出口径流曲线Figure 13. Runoff curve of 20-year-once 24 hour rainfall event in West 10 Plot![]() 图 14 外源雨水地块1年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 14. nstantaneous volume change curve of rain retention barrel of 1-year-once 24 hour rainfall event in exogenous site
图 14 外源雨水地块1年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 14. nstantaneous volume change curve of rain retention barrel of 1-year-once 24 hour rainfall event in exogenous site![]() 图 15 外源雨水地块2年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 15. Instantaneous volume change curve of rain retention barrel of 2-year-once 24 hour rainfall event in exogenous site
图 15 外源雨水地块2年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 15. Instantaneous volume change curve of rain retention barrel of 2-year-once 24 hour rainfall event in exogenous site![]() 图 16 外源雨水地块3年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 16. Instantaneous volume change curve of rain retention barrel of 3-year-once 24 hour rainfall event in exogenous site
图 16 外源雨水地块3年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 16. Instantaneous volume change curve of rain retention barrel of 3-year-once 24 hour rainfall event in exogenous site![]() 图 17 外源雨水地块5年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 17. Instantaneous volume change curve of rain retention barrel of 5-year-once 24 hour rainfall event in exogenous site
图 17 外源雨水地块5年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 17. Instantaneous volume change curve of rain retention barrel of 5-year-once 24 hour rainfall event in exogenous site![]() 图 18 外源雨水地块10年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 18. Instantaneous volume change curve of rain retention barrel of 10-year-once 24 hour rainfall event in exogenous site
图 18 外源雨水地块10年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 18. Instantaneous volume change curve of rain retention barrel of 10-year-once 24 hour rainfall event in exogenous site![]() 图 19 外源雨水地块20年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 19. Instantaneous volume change curve of rain retention barrel of 20-year-once 24 hour rainfall event in exogenous site
图 19 外源雨水地块20年一遇24小时降雨雨水调蓄池瞬时容积变化曲线Figure 19. Instantaneous volume change curve of rain retention barrel of 20-year-once 24 hour rainfall event in exogenous site(1) 根据表 3,通过低影响开发模型与传统开发模型结果数据对比,可知经过西十地块地表低影响开发设施后,在1年一遇、2年一遇、3年一遇、5年一遇、10年一遇及20年一遇降雨强度下出水口的径流总量分别削减100%、100%、100%、78.79%、50.41%、34.73%,峰值流量分别减少100%、100%、100%、70.12%、33.33%、19.44%,峰现时间为1年一遇、2年一遇、3年一遇峰值消失,5年一遇、10年一遇及20年一遇降雨强度下分别延迟375、310、240 min。由此得出结论:低影响开发模式可减少径流总量、峰值径流量,推迟峰现时间。外源雨水场地内LID设施除透水铺装外只有植草沟。由表 4可知,对比传统开发模式,低影响开发模式对场地径流总量的削减量较小,1年一遇、2年一遇、3年一遇、5年一遇、10年一遇、20年一遇重现期径流总量分别为800、2 633、4 189、6 145、10 249、13 970 m3,因此可将这部分径流收集到雨水调蓄池中用于西十地块浇灌。
(2) 根据图 8~19可知:低影响开发模式在降雨前期对地表瞬时径流量的削减能力较强;随着时间的推迟削减能力逐渐减弱,在降雨后期低影响开发设施承载容量趋于饱和,此时低影响开发模式下的出口径流量高于传统开发模式。
4.2 储蓄利用系统结果与分析
4.2.1 储蓄结果与分析
根据SWMM模拟结果,外源雨水地块全年总蓄水量为9 617 m3,径流收集主要集中在第25~35周;单周最大收水总量为2 078 m3,发生在第27周(6月初);第1~20周、43~52周收水量基本为0。
4.2.2 浇灌利用计算与分析
根据植物种类及北京气候类型,可推算出园林植物需水量:
PWR=ET0×KL (1) 式中:PWR为园林植物需水量,mm/d;ETO为参照作物需水量,mm/d;KL为园林系数。
根据彭曼-蒙蒂斯公式计算参照作物需水量:
ETo=0.408Δ(Rn−G)+γ900T+273u2(es−ea)Δ+γ(1+0.34u2) (2) 式中:ETO为参照作物需水量,mm/d;Rn为输入冠层净辐射量,MJ/m2·d;G为土壤热通量,MJ/m2·d;T为2 m高处日平均温度,℃;u2为2 m高处风速,m/s;es为饱和水汽压,kPa;ea为实际水汽压,kPa;Δ为饱和水汽压与温度关系曲线在某处得斜率,kPa/℃;γ为干湿温度计常数,kPa/℃。
园林系数(KL)的计算公式为:
KL=KsKdKme (3) 式中:Ks为园林植物种类因子;Kd为园林种植密度因子;Kmc为园林小气候因子。
园林植物净需水量:
PWRnet=PWR−Re (4) 式中:PWRnet为园林植物净需水量,mm/d;Re为有效降雨量,mm/d。
园林植物净灌溉需水量(IWRnet)是指存储于植物根区,可添加于土壤水库以满足净植物需水量(PWRnet)的那部分灌溉水量。
IWRnet=PWRnet (5) 为简化运算过程,将已报道的不同类型植物的种类系数、密度因子、小气候因子平均值[14](表 7)带入公式(3)得到冷季型草园林系数(KL)为0.8,其余植被类型园林系数(KL)为0.5。本研究参照北京市房山地区气象站20年(1982—2002年)常规观测气象数据,根据公式(2)计算出每月平均ETO值[16],估算出园林植物需水量PWR(表 8)。园林植物在不降雨或降雨不足时需要人工灌溉。将上述结果带入公式(1)计算得出不降雨时园林植物需水量速率;结合不同植被类型面积求得2017年每周西十场地园林植物灌溉需水量;与雨水调蓄池周容积量进行对比可知每周雨水利用率及雨水调蓄池空置率如图 20所示。由此,计算得出第25周、第27周、第29~32周、第34~35周、第41周雨水调蓄池中收集水量在满足当月灌溉需求后仍有余量;全年径流收集的利用率为43.65%;在冬季雨水调蓄池空置率较大。
![]() 图 20 2017年每周植物需水量与雨水调蓄池容积量关系图Figure 20. Diagram of weekly plant water requirement and rainwater storage of rain retention barrel in 2017表 7 各类园林植物园林系数相关指标及面积Table 7. Relevant indicators and area of KL for all kinds of garden plants
图 20 2017年每周植物需水量与雨水调蓄池容积量关系图Figure 20. Diagram of weekly plant water requirement and rainwater storage of rain retention barrel in 2017表 7 各类园林植物园林系数相关指标及面积Table 7. Relevant indicators and area of KL for all kinds of garden plants植被类型Vegetation type 种类系数Coefficient of species(Ks) 密度因子Density factor(Kd) 小气候因子Microclimatic factor(Kmc) 面积Area/m2 乔木Tree 0.9 0.5 0.2 1.3 1.0 0.5 1.4 1.0 0.5 灌木Shrub 0.7 0.5 0.2 1.1 1.0 0.5 1.3 1.0 0.5 地被植物Ground plant 0.9 0.5 0.2 1.1 1.0 0.5 1.2 1.0 0.5 12 400.5 树木、灌木、地被植物混合Mixed trees, shrubs and ground plants 0.9 0.5 0.2 1.3 1.0 0.6 1.4 1.0 0.5 冷季型草Cold season type grass 0.8 1.0 1.0 0.6 1.2 1.0 0.8 5 971.3 合计Total 18 371.8 表 8 2017年每月植物需水量Table 8. Monthly plant water requirement in 2017时间Time 1月Jan. 2月Feb. 3月Mar. 4月Apr. 5月May. 6月Jun. 7月Jul. 8月Aug. 9月Sep. 10月Oct. 11月Nov. 12月Dec. ETO/(mm·d-1) 0.72 1.20 2.17 3.38 4.26 4.72 4.21 3.74 3.02 2.00 1.19 0.72 PWR/(mm·d-1) 0.43 0.72 1.30 2.02 2.55 2.82 2.52 2.24 1.81 1.20 0.71 0.43 5. 结论
按照西十地块及外源雨水地块低影响开发系统雨水消纳能力应达到北京市3年一遇24 h降雨量(108 mm[9])的标准,经过上述设计研究及对SWMM模拟结果的定量分析可得出以下结论:
(1) 面对相同的雨洪调控目标,本研究根据基底条件和景观需求的差异制定不同的设计策略。西十地块优先构建渗透滞留系统,结合场地现状基底构建425.4 m2植草沟、1 035.8 m2生物滞留池、1 172.7 m2下沉式绿地、4 871.3 m2绿色屋顶以及6 647.8 m2透水铺装等5类LID设施;外源雨水地块以储蓄利用为目的,末端构建4 189 m3雨水调蓄池,最终均达到设计目标要求。
(2) 低影响开发设施对雨洪问题具有良好的调控效果,可以减少径流总量、峰值流量,推迟峰现时间,但随着时间的推迟削减能力逐渐减弱。由此可知,低影响开发系统的雨洪调控能力具有上限。
(3) 根据北京市2017年全年降雨量日值统计,外源雨水地块雨水调蓄池中径流收集的利用率为43.65%,有30周(210 d)雨水调蓄池空置率为100%,较为浪费。因此在进行低影响开发系统设计时,应综合考虑成本造价等因素。
-
![]()
图 2 林分空间结构综合指数(CSSI)变化情况
Figure 2. Changes of comprehensive index of spatial structure (CSSI)
表 1 样地基本概况
Table 1 General description of sample plot
样地号
Sample plot No.海拔
Altitude/m坡位
Slope position坡度
Slope gradient/(°)郁闭度
Canopy density密度/(株·hm− 2)
Stand density/(tree·ha− 1)树种组成
Species composition处理
Treatment1 705 中 Middle 8 0.70 766 6蒙1桦1杨1红1其他 T1 2 738 中 Middle 8 0.71 896 4蒙2杨1桦1色1椴1红 T2 3 721 中 Middle 7 0.72 1 038 5蒙2桦1色1杨1椴 T3 4 741 中 Middle 8 0.83 992 4蒙2桦1色1椴1红1其他 T2 5 637 中 Middle 8 0.85 998 5蒙2桦1红1黑1其他 T1 6 635 中 Middle 8 0.88 1 047 5蒙3桦1黑1红 T3 7 677 中 Middle 7 0.90 800 6蒙1黑1胡1水1其他 T1 8 685 中 Middle 7 0.77 929 6蒙2桦1黑1其他 T3 9 703 中 Middle 6 0.87 995 4蒙3桦1杨2其他 T2 注:蒙代表蒙古栎,桦代表白桦,杨代表大青杨,红代表红松,色代表色木槭,椴代表紫椴,落代表长白落叶松,黑代表黑桦,胡代表胡桃楸,水代表水曲柳,其他代表其他树种。Notes: 蒙 stands for Quercus mongolica, 桦 stands for Betula platyphylla, 杨 stands for Populus ussuriensis, 红 stands for Pinus koraiensis, 色 stands for Acer mono, 椴 stands for Tilia tuan, 落 stands for Larix olgensis, 黑 stands for Betula dahurica, 胡 stands for Juglans mandshurica, 水 stands for Fraxinus mandschurica, 其他 stands for other tree species.  下载: 导出CSV
下载: 导出CSV
表 2 蒙古栎次生林林分基本信息
Table 2 General information of oak secondary forest
样地号
Sample plot No.胸径 DBH/cm 树高 Tree height/m 优势平均高
Mean dominant height/m蓄积
Volume/m3最小 Min. 平均 Average 最大 Max. 最小 Min. 平均 Average 最大 Max. 1 5.0 14.6 66.0 2.0 11.0 29.6 22.2 150.72 2 5.0 13.7 59.1 0.5 11.2 24.1 21.5 162.31 3 5.0 13.0 55.7 1.3 10.25 28.5 21.6 172.00 4 5.0 13.6 50.2 1.5 10.9 25.8 21.2 176.10 5 5.0 13.1 50.9 1.5 8.9 22.4 21.5 162.30 6 5.0 12.8 58.5 3.2 11.5 31.1 22.2 173.91 7 5.0 14.3 52.2 1.4 9.8 24.9 21.8 154.45 8 5.0 14.3 70.1 3.1 12.6 24.2 22.9 179.20 9 5.0 14.1 58.2 2.2 11.0 26.5 22.7 186.90 注:采用汪清林业局一元立木材积表[28]计算单木材积和林分蓄积,选取样地最高的5株林木计算优势高平均值。Notes: volume of wood was calculated by the tree volume table of Wangqing Forestry Bureau, and the mean dominant height was calculated by choosing five highest trees in each sample plot.
下载: 导出CSV
表 3 采伐前后目标树结构单元的各个空间结构指标
Table 3 Spatial structure indexes of target trees before and after thinning
抚育间伐方式
Thinning treatment类型
TypeM平均值
Mean uniform angle indexW平均值
Mean neighborhood patternU平均值
Mean uniform angel indexC平均值
Mean crowding degreeT2 伐前 Before thinning 0.615 5 0.529 2 0.184 0 0.415 1 伐后 After thinning 0.638 3 0.524 1 0.177 7 0.409 4 T3 伐前 Before thinning 0.509 9 0.533 2 0.226 7 0.407 4 伐后 After thinning 0.529 7 0.536 0 0.216 7 0.395 3 注:M代表混交度;W代表角尺度;U代表大小比数;C代表密集度;T2代表目标树经营密度1;T3代表目标树经营密度2。下同。Notes: M represents for mingling degree, W represents for uniform angel index, U represents for neighborhood comparison, C represents for crowding degree, T2 represents for target tree management with density 1, T3 represents for target tree management with desity 2. Same as below.
下载: 导出CSV
表 4 不同抚育间伐方式林分采伐前后
M 频率分布及林分平均混交度(¯M )Table 4 M frequency distribution and mean mingling degree of different thinning treatments before and after thinning
抚育间伐方式
Thinning treatment伐前 Before thinning 伐后 After thinning 频率分布 Frequency distribution ¯M 频率分布 Frequency distribution ¯M 0.00 0.25 0.50 0.75 1.00 0.00 0.25 0.50 0.75 1.00 T1 0.159 2 0.190 1 0.242 1 0.248 0 0.160 5 0.515 1 0.150 3 0.192 0 0.252 7 0.243 3 0.161 7 0.518 5 T2 0.071 0 0.173 1 0.250 0 0.277 9 0.228 0 0.604 7 0.062 3 0.165 3 0.253 9 0.289 5 0.229 0 0.614 4 T3 0.107 9 0.193 9 0.258 5 0.255 1 0.184 6 0.553 7 0.092 2 0.194 5 0.267 3 0.259 2 0.186 9 0.563 5 注:T1代表传统经营措施。Note: T1 represents for traditional management.
下载: 导出CSV
表 5 不同抚育间伐方式林分采伐前后
W 频率分布及林分平均角尺度(¯W )Table 5 W frequency distribution and mean value of stand uniform angel index of different thinning treatments before and after thinning
抚育间伐方式
Tninning treatment伐前 Before thinning 伐后 After thinning 频率分布 Frequency distribution ¯W 频率分布 Frequency distribution ¯W 0.00 0.25 0.50 0.75 1.00 0.00 0.25 0.50 0.75 1.00 T1 0.005 9 0.167 8 0.553 3 0.196 1 0.077 0 0.542 6 0.006 7 0.165 1 0.558 6 0.196 8 0.072 8 0.540 9 T2 0.003 6 0.179 9 0.580 9 0.163 7 0.071 9 0.530 1 0.004 2 0.185 0 0.583 6 0.165 3 0.061 8 0.523 9 T3 0.004 9 0.164 2 0.578 2 0.191 4 0.061 2 0.535 0 0.006 1 0.168 0 0.584 5 0.184 3 0.057 0 0.529 5
下载: 导出CSV
表 6 不同抚育间伐方式林分采伐前后
U 频率分布及林分平均大小比数(¯U )Table 6 U frequency distribution and mean value of neighborhood comparison of different thinning treatments before and after thinning
抚育间伐方式
Thinning treatment伐前 Before thinning 伐后 After thinning 频率分布 Frequency distribution ¯U 频率分布 Frequency distribution ¯U 0.00 0.25 0.50 0.75 1.00 0.00 0.25 0.50 0.75 1.00 T1 0.207 2 0.196 1 0.205 9 0.194 7 0.196 1 0.494 1 0.212 3 0.196 8 0.202 2 0.195 4 0.193 4 0.490 2 T2 0.207 7 0.204 6 0.194 2 0.192 0 0.201 4 0.493 7 0.209 8 0.197 2 0.201 4 0.190 6 0.200 9 0.493 9 T3 0.204 6 0.206 5 0.191 9 0.201 2 0.195 8 0.494 3 0.209 8 0.201 6 0.193 0 0.198 6 0.197 0 0.492 9
下载: 导出CSV
表 7 不同抚育间伐方式林分采伐前后
C 频率分布及林分平均密集度(¯C )Table 7 C frequency distribution and mean value of crowding degree of different thinning treatments before and after thinning
抚育间伐方式
Thinning treatment伐前 Before thinning 伐后 After thinning 频率分布 Frequency distribution ¯C 频率分布 Frequency distribution ¯C 0.00 0.25 0.50 0.75 1.00 0.00 0.25 0.50 0.75 1.00 T1 0.059 9 0.138 8 0.166 4 0.221 7 0.413 2 0.338 3 0.063 3 0.140 8 0.175 2 0.217 0 0.403 6 0.336 3 T2 0.021 6 0.072 4 0.141 2 0.216 3 0.548 6 0.391 8 0.023 9 0.078 2 0.158 3 0.221 5 0.518 0 0.389 3 T3 0.018 5 0.074 8 0.163 8 0.221 1 0.521 9 0.384 1 0.021 9 0.080 4 0.183 3 0.235 7 0.478 6 0.378 1
下载: 导出CSV
-
[1] 陆元昌, 栾慎强, 张守攻, 等. 从法正林转向近自然林: 德国多功能森林经营在国家区域和经营单位层面的实践[J]. 世界林业研究, 2010, 23(1):1−11. Lu Y C, Luan S Q, Zhang S G, et al. From normal forest to close-to-nature forest: multi-functional forestry and its practice at national, regionaland forest management unit levels in Germany[J]. World Forestry Research, 2010, 23(1): 1−11.
[2] 陆元昌. 近自然森林经营的理论与实践[M]. 北京: 科学出版社, 2006. Lu Y C. Theory and practice of close-to-nature forest management[M]. Beijing: Science Press, 2006.
[3] 李慧卿, 江泽平, 雷静品, 等. 近自然森林经营探讨[J]. 世界林业研究, 2007, 20(4):6−11. doi: 10.3969/j.issn.1001-4241.2007.04.002 Li H Q, Jiang Z P, Lei J P, et al. Exploitation of the theory and application of close to nature forest management in Europe[J]. World Forestry Research, 2007, 20(4): 6−11. doi: 10.3969/j.issn.1001-4241.2007.04.002
[4] 王祖华, 李瑞霞, 关庆伟. 间伐对杉木人工林不同根序细根形态、生物量和氮含量的影响[J]. 应用生态学报, 2013, 24(6):1487−1493. Wang Z H, Li R X, Guan Q W. Effects of thinning on fine-root morphology, biomass and N concentration of different branch orders of Chinese fir[J]. Chinese Journal of Applied Ecology, 2013, 24(6): 1487−1493.
[5] 雷相东, 陆元昌, 张会儒, 等. 抚育间伐对落叶松云冷杉混交林的影响[J]. 林业科学, 2005, 41(4):78−85. doi: 10.3321/j.issn:1001-7488.2005.04.014 Lei X D, Lu Y C, Zhang H R, et al. Effects of thinning on mixed stands of Larix olgensis, Abies nephrolepis and Picea jazoensis[J]. Scientia Silvae Sinicae, 2005, 41(4): 78−85. doi: 10.3321/j.issn:1001-7488.2005.04.014
[6] 高云昌, 张文辉, 何景峰, 等. 黄龙山油松人工林间伐效果的综合评价[J]. 应用生态学报, 2013, 24(5):1313−1319. Gao Y C, Zhang W H, He J F, et al. Effects of thinning intensity on Pinus tabulaeformis plantation in Huanglong Mountain, Northwest China: a comprehensive evaluation[J]. Chinese Journal of Applied Ecology, 2013, 24(5): 1313−1319.
[7] 汤孟平, 唐守正, 雷相东, 等. 林分择伐空间结构优化模型研究[J]. 林业科学, 2004, 40(5):25−31. doi: 10.3321/j.issn:1001-7488.2004.05.004 Tang M P, Tang S Z, Lei X D, et al. Study on spatial structure optimizing model of stand selection cutting[J]. Scientia Silvae Sinicae, 2004, 40(5): 25−31. doi: 10.3321/j.issn:1001-7488.2004.05.004
[8] 惠刚盈, 胡艳波. 混交林树种空间隔离程度表达方式的研究[J]. 林业科学研究, 2001, 14(1):23−27. doi: 10.3321/j.issn:1001-1498.2001.01.004 Hui G Y, Hu Y B. The optimum standard angle of the uniform angle index[J]. Forest Research, 2001, 14(1): 23−27. doi: 10.3321/j.issn:1001-1498.2001.01.004
[9] Patrick J B, Andrew P R, Ewel J J. Sudden and sustained response of Acacia koa crop trees to crown release in stagnant stands[J]. Canadian Journal of Forest Research, 2008, 38(4): 656−666. doi: 10.1139/X07-137
[10] Schuler T M. Crop tree release improves competitiveness of northernred oak growing in association with black cherry[J]. Northern Journal of Applied Forestry, 2006, 23(2): 77−82.
[11] 吴瑶, 李凤日, 秦凯伦, 等. 近自然经营技术对红松林土壤化学性质的影响[J]. 东北林业大学学报, 2014, 42(1):76−79. doi: 10.3969/j.issn.1000-5382.2014.01.017 Wu Y, Li F R, Qin K L, et al. Impacts of close-to-nature management on soil chemical properties of Korean pine forest in northeastern China[J]. Journal of Northeast Forestry University, 2014, 42(1): 76−79. doi: 10.3969/j.issn.1000-5382.2014.01.017
[12] 孙冬婧, 温远光, 罗应华, 等. 近自然化改造对杉木人工林物种多样性的影响[J]. 林业科学研究, 2015, 28(2):202−208. Sun D J, Wen Y G, Luo Y H, et al. Effect of close-to-nature management on species diversity in a Cunninghamia lanceolata plantation[J]. Forest research, 2015, 28(2): 202−208.
[13] 王懿祥, 张守攻, 陆元昌, 等. 干扰树间伐对马尾松人工林目标树生长的初期效应[J]. 林业科学, 2014, 50(10):67−73. Wang Y X, Zhang S G, Lu Y C, et al. Initial effects of crop trees growth after crop tree release on Pinus massoniana plantation[J]. Scientia Silvae Sinicae, 2014, 50(10): 67−73.
[14] 周建云, 李荣, 何景峰, 等. 近自然经营对辽东栎林优势乔木更新的影响[J]. 林业科学, 2013, 49(8):15−20. Zhou J Y, Li R, He J F, et al. Regeneration of the dominant arbors after close-to-natural management of Quercus wutaishanica forest[J]. Scientia Silvae Sinicae, 2013, 49(8): 15−20.
[15] 雷相东, 唐守正. 林分结构多样性指标研究综述[J]. 林业科学, 2002, 38(3):140−146. doi: 10.3321/j.issn:1001-7488.2002.03.025 Lei X D, Tang S Z. Indicators on structural diversity within-stand: a review[J]. Scientia Silvae Sinicae, 2002, 38(3): 140−146. doi: 10.3321/j.issn:1001-7488.2002.03.025
[16] 曹小玉, 李际平. 林分空间结构指标研究进展[J]. 林业资源管理, 2016, 38(4):65−73. Cao X Y, Li J P. Research progress on indicators of the stand spatial structure[J]. Forest Resources Management, 2016, 38(4): 65−73.
[17] 李建, 彭鹏, 何怀江, 等. 采伐对吉林蛟河针阔混交林空间结构的影响[J]. 北京林业大学学报, 2017, 39(9):48−57. Li J, Peng P, He H J, et al. Effects of thinning intensity on spatial structure of multi-species temperate forest at Jiaohe in Jilin Province, northeastern China[J]. Journal of Beijing Forestry University, 2017, 39(9): 48−57.
[18] 张连金, 孙长忠, 赖光辉. 北京九龙山侧柏生态公益林空间结构分析与评价[J]. 林业科学研究, 2018, 31(4):75−82. Zhang L J, Sun C Z, Lai G H. Analysis and evaluation of stand spatial structure of Platycladus orientalis ecological forest in Jiulongshan of Beijing[J]. Forest Research, 2018, 31(4): 75−82.
[19] 吴征镒. 中国植被[M]. 北京: 科学出版社, 1980. Wu Z Y, China vegetation[M]. Beijing: Science Press, 1980.
[20] 王良民, 任宪威, 刘一樵, 等. 我国落叶栎的地理分布[J]. 北京林学院学报, 1985, 7(2):57−69. Wang L M, Ren X W, Liu Y Q, et al. The geographical distribution of deciduous oak in China[J]. Journal of Beijing Forestry University, 1985, 7(2): 57−69.
[21] 郑焕能, 贾松青, 胡海清. 大兴安岭林区的林火与森林恢复[J]. 东北林业大学学报, 1986, 14(4):1−7. Zheng H N, Jia S Q, Hu H Q. Forest fire and forest restoration in Daxinganling Forest Area[J]. Journal of Northeast Forestry University, 1986, 14(4): 1−7.
[22] 于顺利, 马克平, 陈灵芝. 中国北方蒙古栎林起源和发展的初步探讨[J]. 广西植物, 2000, 5(2):131−137. doi: 10.3969/j.issn.1000-3142.2000.02.005 Yu S L, Ma K P, Chen L Z. A preliminary study on the origin and development of Mongolian oak forest in north China[J]. Guihaia, 2000, 5(2): 131−137. doi: 10.3969/j.issn.1000-3142.2000.02.005
[23] Vose J M, Miniat C F, Luce C H, et al. Ecohydrological implications of drought for forests in the United States[J]. Forest Ecology and Management, 2016, 380: 335−345. doi: 10.1016/j.foreco.2016.03.025
[24] Andreas B, Christian A, Magnus L, et al. Adaptive forest management in central Europe: climate change impacts, strategies and integrative concept[J]. Scandinavian Journal of Forest Research, 2009, 24(6): 473−482. doi: 10.1080/02827580903418224
[25] Rigling A, Bigler C, Eilmann U, et al. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests[J]. Global Change Biology, 2012, 19(1): 229−240.
[26] Schelhaas M J, Nabuurs G J, Hengeveld G, et al. Alternative forest management strategies to account for climate change-induced productivity and species suitability changes in Europe[J]. Regional Environmental Change, 2015, 15(8): 1581−1594. doi: 10.1007/s10113-015-0788-z
[27] 殷晓洁, 周广胜, 隋兴华, 等. 蒙古栎地理分布的主导气候因子及其阈值[J]. 生态学报, 2013, 33(1):103−109. Yin X J, Zhou G S, Sui X H, et al. Dominant climatic factors of Quercus mongolica geographical distribution and their thresholds[J]. Acta Ecologica Sinica, 2013, 33(1): 103−109.
[28] 刘琪璟. 中国立木材积表[M]. 北京: 中国林业出版社, 2018. Liu Q J. Tree volume tables of China[M]. Beijing: China Forestry Publishing House, 2018.
[29] 周红敏, 惠刚盈, 赵中华, 等. 林分空间结构分析中样地边界木的处理方法[J]. 林业科学, 2009, 45(2):1−5. doi: 10.3321/j.issn:1001-7488.2009.02.001 Zhou H M, Hui G Y, Zhao Z H, et al. Treatment methods of plot boundary trees in spatial forest structure analysis[J]. Scientia Silvae Sinicae, 2009, 45(2): 1−5. doi: 10.3321/j.issn:1001-7488.2009.02.001
[30] 惠刚盈, Gadow K V. 森林空间结构量化分析方法[M]. 北京: 中国科学技术出版社, 2003. Hui G Y, Gadow K V. Quantitative analysis method of forest spatial structure[M]. Beijing: China Science and Technology Press, 2003.
[31] 汤孟平. 森林空间结构研究现状与发展趋势[J]. 林业科学, 2010, 46(1):117−122. Tang M P. Advances in study of forest spatial structure[J]. Scientia Silvae Sinicae, 2010, 46(1): 117−122.
[32] Aguirre O, Hui G Y, Gadow K V, et al. An analysis of spatial forest structure using neighbourhood-based variables[J]. Forest Ecology & Management, 2003, 183(1): 137−145.
[33] Davies O, Pommerening A. The contribution of structural indices to the modelling of Sitka spruce (Picea sitchensis) and birch (Betula spp.) crowns[J]. Forest Ecology and Management, 2008, 256(1−2): 68−77. doi: 10.1016/j.foreco.2008.03.052
[34] 岳永杰, 余新晓, 李钢铁, 等. 北京松山自然保护区蒙古栎林的空间结构特征[J]. 应用生态学报, 2009, 20(8):1811−1816. Yue Y J, Yu X X, Li G T, et al. Spatial structure of Quercus mongolica forest in Beijing Songshan Mountain Nature Reserve[J]. Chinese Journal of Applied Ecology, 2009, 20(8): 1811−1816.
[35] 段昌盛, 王军辉, 马建伟, 等. 秦岭西段锐齿栎林分经营状态评价[J]. 北京林业大学学报, 2009, 31(5):61−66. doi: 10.3321/j.issn:1000-1522.2009.05.011 Duan C S, Wang J H, Ma J W, et al. Evaluation of Quercus aliena var. acuteserrata forest at the western segment of Qinling Mountain, northwestern China[J]. Journal of Beijing Forestry University, 2009, 31(5): 61−66. doi: 10.3321/j.issn:1000-1522.2009.05.011
[36] 刘彦, 余新晓, 岳永杰, 等. 北京密云水库集水区刺槐人工林空间结构分析[J]. 北京林业大学学报, 2009, 31(5):25−28. doi: 10.3321/j.issn:1000-1522.2009.05.005 Liu Y, Yu X X, Yue Y J, et al. Spatial structure of Robinia pseudoacacia plantation in Miyun Reservoir Watershed of Beijing[J]. Journal of Beijing Forestry University, 2009, 31(5): 25−28. doi: 10.3321/j.issn:1000-1522.2009.05.005
[37] 胡艳波, 惠刚盈. 基于相邻木关系的林木密集程度表达方式研究[J]. 北京林业大学学报, 2015, 37(9):1−8. Hu Y B, Hui G Y. How to describe the crowding degree of trees based on the relationship of neighboring trees[J]. Journal of Beijing Forestry University, 2015, 37(9): 1−8.
[38] 董灵波, 刘兆刚, 马妍, 等. 天然林林分空间结构综合指数的研究[J]. 北京林业大学学报, 2013, 35(1):16−22. Dong L B, Liu Z G, Ma Y, et al. A new composite index of stand spatial structure for natural forest[J]. Journal of Beijing Forestry University, 2013, 35(1): 16−22.
[39] Gadow K V. ZurBestandesbeschreibung in der Forsteinrichtung[J]. Forst und Holz, 1993, 48(21): 602−606.
[40] Füldner K. Strukturbeschreibung von Buchen-Edellaubholz-Mischwäldern[M]. Göttingen: Cuvillier Verlag, 1995.
[41] 惠刚盈, Gadow K V, 胡艳波, 等. 林木分布格局类型的角尺度均值分析方法[J]. 生态学报, 2004, 24(6):1225−1229. doi: 10.3321/j.issn:1000-0933.2004.06.020 Hui G Y, Gadow K V, Hu Y B, et al. Characterizing forest spatial distribution pattern with the mean value of uniform angle index[J]. Acta Ecologica Sinica, 2004, 24(6): 1225−1229. doi: 10.3321/j.issn:1000-0933.2004.06.020
[42] 惠刚盈, Gadow K V, Matthias A. 角尺度−一个描述林木个体分布格局的结构参数[J]. 林业科学, 1999, 35(1):37−42. doi: 10.3321/j.issn:1001-7488.1999.01.006 Hui G Y, Gadow K V, Matthias A. The neighbourhood pattern: a new structure parameter for describing distribution of forest tree position[J]. Scientia Silvae Sinicae, 1999, 35(1): 37−42. doi: 10.3321/j.issn:1001-7488.1999.01.006
[43] 胡艳波, 惠刚盈, 王宏翔, 等. 随机分布的角尺度置信区间及其应用[J]. 林业科学研究, 2014, 27(3):302−308. Hu Y B, Hui G Y, Wang H X, et al. Uniform angle index (w) confidence interval of the random distribution and its application[J]. Forest Research, 2014, 27(3): 302−308.
[44] 惠刚盈, Gadow K V, Albert M. 一个新的林分空间结构参数: 大小比数[J]. 林业科学研究, 1999, 12(1):1−6. doi: 10.3321/j.issn:1001-1498.1999.01.001 Hui G Y, Gadow K V, Albert M. A new parameter for stand spatial structure: neighborhood comparison[J]. Forest Research, 1999, 12(1): 1−6. doi: 10.3321/j.issn:1001-1498.1999.01.001
[45] 张俊艳, 陆元昌, 成克武, 等. 近自然改造对云南松人工林群落结构及物种多样性的影响[J]. 河北农业大学学报, 2010, 33(3):72−77. doi: 10.3969/j.issn.1000-1573.2010.03.016 Zhang J Y, Lu Y C, Cheng K W, et al. Impact of near nature modification on community structure and species biodiversity of artificial Pinus yunnanensis forest[J]. Journal of Agricultural University of Hebei, 2010, 33(3): 72−77. doi: 10.3969/j.issn.1000-1573.2010.03.016
[46] 邬可义. 哈尔滨市的森林近自然和价值化经营实践[J]. 世界林业研究, 2009, 22(6):14−18. Wu K Y. Close-to-nature and value-oriented management of forests in Harbin[J]. World Forestry Research, 2009, 22(6): 14−18.
[47] 冯琪雅, 陈超凡, 覃林, 等. 不同经营模式对蒙古栎天然次生林林分结构和植物多样性的影响[J]. 林业科学, 2018, 54(1):12−21. Feng Q Y, Chen C F, Qin L, et al. Effects of different management models on stand structure and plant diversity of natural secondary forests of Quercus mongolica[J]. Scientia Silvae Sinicae, 2018, 54(1): 12−21.
[48] 陈科屹, 张会儒, 雷相东, 等. 基于目标树经营的抚育采伐对云冷杉针阔混交林空间结构的影响[J]. 林业科学研究, 2017, 30(5):718−726. Chen K Y, Zhang H R, Lei X D, et al. Effect of thinning on spatial structure of spruce-fir mixed broadleaf-conifer forest base on crop tree management[J]. Forest Research, 2017, 30(5): 718−726.
[49] 吴建强, 王懿祥, 杨一, 等. 干扰树间伐对杉木人工林林分生长和林分结构的影响[J]. 应用生态学报, 2015, 26(2):340−348. Wu J Q, Wang Y X, Yang Y, et al. Effects of crop tree release on stand growth and stand structure of Cunninghamia lanceolata plantation[J]. Chinese Journal of Applied Ecology, 2015, 26(2): 340−348.
[50] 张会儒, 武纪成, 杨洪波, 等. 长白落叶松−云杉−冷杉混交林林分空间结构分析[J]. 浙江农林大学学报, 2009, 26(3):319−325. doi: 10.3969/j.issn.2095-0756.2009.03.005 Zhang H R, Wu J C, Yang H B, et al. Spatial structure of mixed larch-spruce-fir stands[J]. Journal of Zhejiang Forestry College, 2009, 26(3): 319−325. doi: 10.3969/j.issn.2095-0756.2009.03.005
[51] 李先琨, 黄玉清, 苏宗明, 等. 元宝山南方红豆杉种群分布格局及动态[J]. 应用生态学报, 2000, 11(2):169−172. doi: 10.3321/j.issn:1001-9332.2000.02.003 Li X K, Huang Y Q, Su Z M, et al. Distribution pattern and its dynamics of Taxus chinensis var. mairei population on Yunbaoshan Mountain[J]. Chinese Journal of Applied Ecology, 2000, 11(2): 169−172. doi: 10.3321/j.issn:1001-9332.2000.02.003
[52] 赵中华, 惠刚盈, 胡艳波, 等. 基于大小比数的林分空间优势度表达方法及其应用[J]. 北京林业大学学报, 2014, 36(1):78−82. Zhao Z H, Hui G Y, Hu Y B, et al. Method and application of stand spatial advantage degree based on the neighborhood comparison[J]. Journal of Beijing Forestry University, 2014, 36(1): 78−82.
[53] 李建军, 李际平. 基于粗糙集的林分经营决策因子分类研究[J]. 福建林业科技, 2008, 35(3):4−9. doi: 10.3969/j.issn.1002-7351.2008.03.002 Li J J, Li J P. Classification rule study of forest management decision-factor based on the Rough Set Theory[J]. Journal of Fujian Forestry Science & Technology, 2008, 35(3): 4−9. doi: 10.3969/j.issn.1002-7351.2008.03.002
[54] 曾群英, 周元满, 李际平, 等. 基于生态系统经营的林分采伐决策方法[J]. 东北林业大学学报, 2010, 38(9):31−35. doi: 10.3969/j.issn.1000-5382.2010.09.010 Zeng Q Y, Zhou Y M, Li J P, et al. Decision-making methodology of forest ecosystem management based on spatial structure factors of forest[J]. Journal of Northeast Forestry University, 2010, 38(9): 31−35. doi: 10.3969/j.issn.1000-5382.2010.09.010
[55] 胡艳波, 惠刚盈. 优化林分空间结构的森林经营方法探讨[J]. 林业科学研究, 2006, 19(1):1−8. doi: 10.3321/j.issn:1001-1498.2006.01.001 Hu Y B, Hui G Y. A discussion on forest management method optimizing forest spatial structure[J]. Forest Research, 2006, 19(1): 1−8. doi: 10.3321/j.issn:1001-1498.2006.01.001
-
期刊类型引用(6)
1. 莫崇杏,董明亮,李荣生,余纽,郑显澄,杨锦昌. 米老排杂交子代苗期生长性状遗传变异及选择. 森林与环境学报. 2023(05): 555-560 .  百度学术
百度学术
2. Shuchun Li,Jiaqi Li,Yanyan Pan,Xiange Hu,Xuesong Nan,Dan Liu,Yue Li. Variation analyses of controlled pollinated families and parental combining ability of Pinus koraiensis. Journal of Forestry Research. 2021(03): 1005-1011 . 必应学术
3. 潘艳艳,许贵友,董利虎,王成录,梁德洋,赵曦阳. 日本落叶松全同胞家系苗期生长性状遗传变异. 南京林业大学学报(自然科学版). 2019(02): 14-22 . 百度学术
4. 秦光华,宋玉民,乔玉玲,于振旭,彭琳. 旱柳苗高年生长与气象因子的灰色关联度. 东北林业大学学报. 2019(05): 42-45+51 . 百度学术
5. 李峰卿,陈焕伟,周志春,楚秀丽,徐肇友,肖纪军. 红豆树优树种子和幼苗性状的变异分析及优良家系的初选. 植物资源与环境学报. 2018(02): 57-65 . 百度学术
6. 张素芳,张磊,赵佳丽,张莉,张含国. 长白落叶松小RNA测序和其靶基因预测. 北京林业大学学报. 2016(12): 64-72 . 本站查看
其他类型引用(6)























计量
- 文章访问数: 2833
- HTML全文浏览量: 962
- PDF下载量: 130
- 被引次数: 12
