
Comparative study on antioxidant capacity of heteromorphic leaves of Populus euphratica
-
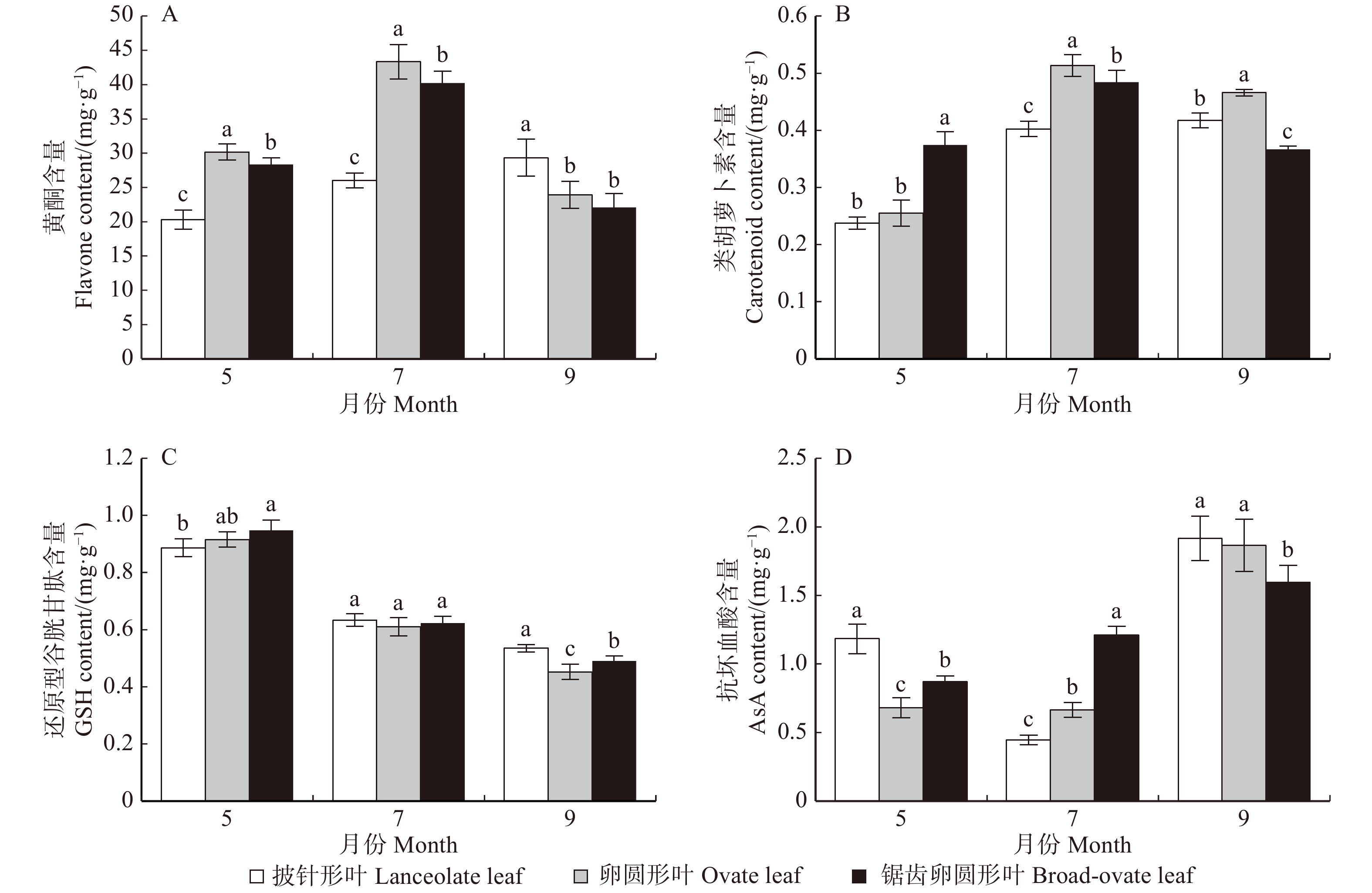
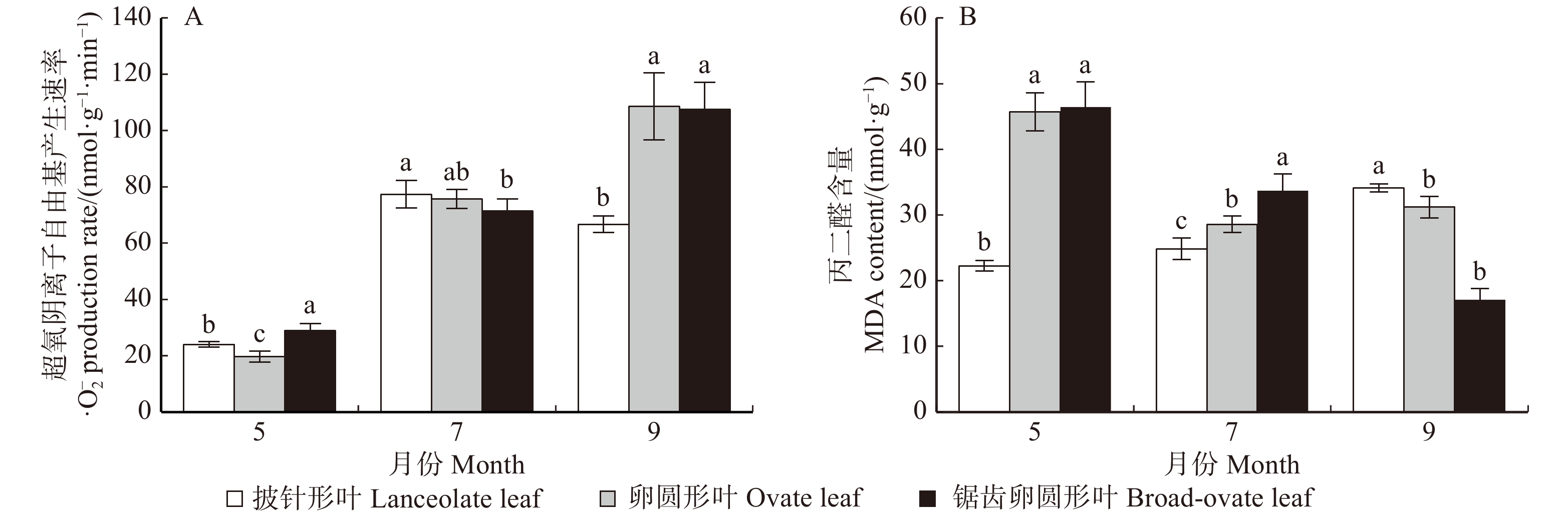
摘要:目的胡杨是我国西部荒漠地区抗逆性较强的树种,具有异叶性。前人的研究发现,胡杨叶片从披针形叶到锯齿卵圆形叶其结构趋于旱生性,且抗性逐渐增强。本文拟从活性氧清除机制着手,深入探讨胡杨异形叶的抗氧化能力,以期阐释其抗逆性的生化机制。方法以成年胡杨披针形叶、卵圆形叶和锯齿卵圆形叶3种典型形态叶为试验材料,对其·O2−产生速率、MDA含量、抗氧化酶(SOD、POD、CAT、APX、GR)活性和抗氧化物质(黄酮、类胡萝卜素、AsA、GSH)含量随季节(5、7和9月)变化的规律进行了比较研究。结果(1)·O2−产生速率随季节而变化,·O2−产生速率在锯齿卵圆形与卵圆形叶中递增,在披针形叶中先增后减;MDA含量在锯齿卵圆形叶中递减,在卵圆形叶中先减后增,而在披针形叶中持续递增。值得注意的是,9月份披针形叶中·O2−产生速率最低而MDA含量最高。(2)锯齿卵圆形叶与卵圆形叶的SOD活性先降后升而CAT活性先升后降,POD活性持续升高而GR活性递减。APX活性在锯齿卵圆形叶中先升后降,在卵圆形叶中一直升高。而披针形叶的SOD、POD、CAT、APX和GR活性均呈先升后降的变化趋势。整体而言,锯齿卵圆形叶的抗氧化酶活性 > 卵圆形叶 > 披针形叶。其中,POD活性差异最为明显。(3)黄酮和类胡萝卜素在7月份大量积累,9月份AsA含量最高而GSH含量最低。黄酮和类胡萝卜素含量在锯齿卵圆形与卵圆形叶中先增后减,在披针形叶中递增;AsA含量在锯齿卵圆形与卵圆形叶中递增,在披针形叶中先减后增;3种形态叶的GSH含量均递减。(4)锯齿卵圆形叶中·O2−产生速率与POD活性及AsA含量呈极显著正相关(P < 0.01),卵圆形叶中·O2−产生速率与POD和APX活性及类胡萝卜素和AsA含量存在极显著正相关(P < 0.01),披针形叶中·O2−产生速率则与POD、CAT和APX活性及黄酮和类胡萝卜素含量都存在极显著正相关(P < 0.01)。结论胡杨3种形态叶的抗氧化能力不同。锯齿卵圆形叶抗氧化能力最强,卵圆形叶次之,披针形叶最弱。3种形态叶随季节变化的抗氧化应答机制也存在差异。夏季锯齿卵圆形叶与卵圆形叶中以POD、CAT和APX作为主要清除ROS的抗氧化酶,秋季则以高活性的SOD和POD作为主要的抗氧化酶。同时,秋季卵圆形叶中APX活性也维持在较高水平。而披针形叶中以SOD、POD、CAT、APX和GR协同清除ROS。此外夏季胡杨叶片大量积累黄酮和类胡萝卜素消除ROS;秋季则通过AsA-GSH循环清除更多的ROS。Abstract:ObjectivePopulus euphratica, commonly known as a desert poplar, is a pioneer tree species with stress resistance and heteromorphic leaves in western China. Previous studies have demonstrated that the structural characteristics of the diversiform-leaves of P. euphratica are related to its eco-adaptability. In the present study, to elucidate the regulation mechanisms of stress tolerance, we focused on the antioxidant capacity for three types of heteromorphic leaves of P. euphratica.MethodIn this paper, the changes of ·O2− production rate, MDA content, antioxidant enzyme activities (SOD, POD, CAT, APX and GR) and antioxidative substance contents (flavone, carotenoid, AsA and GSH) in lanceolate leaves, ovate leaves and dentate broad-ovate leaves, three typical morphological leaves of adult P. euphratica, were studied in different seasons.ResultThe results were as follows: (1) the ·O2− production rate and MDA content: with the change of seasons, the rate of ·O2− production was progressively increased both in dentate broad-ovate leaves and ovate leaves. While it firstly increased and then decreased in lanceolate leaves. The content of MDA decreased in dentate broad-ovate, firstly decreased and then increased in ovate leaves, while it increased in lanceolate leaves all the time. It is noteworthy that compared with the other types of leaves, ·O2− production rate was the lowest while MDA content was the highest in lanceolate leaves in September. (2) The activities of antioxidant enzymes: with the change of seasons, both in dentate broad-ovate leaves and ovate leaves, the activity of SOD firstly decreased and then increased, while the activity of CAT firstly increased and then decreased. There was a progressively increase of the activity of POD, and a decrease of the activity of GR. The activity of APX firstly increased and then decreased in dentate broad-ovate, and it progressively increased in ovate leaves. Remarkably, the activities of SOD, POD, CAT, APX and GR in lanceolate leaves all firstly increased and then decreased. Overall, the activities of antioxidant enzymes were the highest in dentate broad-ovate leaves, followed by ovate leaves, and was the lowest in lanceolate leaves. Furthermore, the difference of POD activity was the most obvious among the three types of leaves. (3) The antioxidative substance contents: in the three heteromorphic leaves of P. euphratica, a large amount of flavone and carotenoid accumulated in July, and AsA content was the highest while GSH content was the lowest in September. The contents of carotenoid and flavone firstly increased and then decreased both in dentate broad-ovate leaves and ovate leaves, while they were increased in lanceolate leaves. The content of AsA progressively increased both in dentate broad-ovate leaves and ovate leaves, while it firstly decreased and then increased in lanceolate leaves. In addition, the content of GSH all decreased in three types of morphological leaves. (4) Correlation analysis: the ·O2− production rate was significantly and positively correlated with POD activity and AsA content in dentate broad-ovate leaves (P < 0.01). In addition to POD and AsA, there was a significant and positive correlation between the ·O2− production rate and APX activity, carotenoid content in ovate leaves (P < 0.01). While the activities of antioxidant enzymes (POD, CAT and APX) and the contents of flavone and AsA were all highly significantly and positively correlated with the ·O2− production rate in lanceolate leaves (P < 0.01).ConclusionThese results showed that there was an obvious difference of the antioxidant ability among the three morphological leaves in P. euphratica. It was the strongest in dentate broad-ovate leaves, followed by ovate leaves, and was the weakest in lanceolate leaves. Moreover, there were differences in the antioxidant response mechanisms of three morphological leaves with seasonal changes. In dentate broad-ovate leaves and oval leaves, POD, CAT and APX were the main antioxidant enzymes for scavenging ROS in summer, while the SOD and POD with high activity were the main antioxidant enzymes in autumn. Meantime, APX activity in the ovate leaves in autumn was also maintained at a high level. In addition, SOD, POD, CAT, APX and GR synergistically scavenged ROS in lanceolate leaves. And in three heteromorphic leaves, flavone and carotenoid were the main antioxidants to eliminate ROS in summer, and more ROS was removed through the AsA-GSH circulation in autumn.
-
Keywords:
- Populus euphratica /
- heteromorphic leaf /
- antioxidase /
- antioxidant
-
多倍体育种是杨树(Populus spp.)遗传改良的重要途径。‘三毛杨’(P. tomentosa ‘Sanmaoyang’)、‘中林46’(P. × euramericana ‘Zhonglin-46’)、‘银中杨’(P. alba × P. berolinensis ‘Yinzhong’)、‘北林雄株1号’(P. × ‘Beilinxiongzhu 1’)、‘中怀1号’(P. × ‘Zhonghuai 1’)等三倍体品种均在我国林业生产中广泛利用[1-5],对推动我国林业产业升级、改善生态环境做出了重要贡献。为了指导林木多倍体品种选育,朱之悌[6]曾提出了“大群体,强选择”的育种策略,认为多倍体育种同样要基于大群体,依据主要目标性状对多倍体种质进行选择,经过遗传测定,才能形成表现优良的新品种。因此,在杨树多倍体品种选育过程中,对性状变异规律进行遗传分析十分必要。
就杨树多倍体育种而言,基于2n配子的有性多倍化途径综合了杂合效应和倍性效应,提供了更加丰富的遗传变异[7]。植物生长通过叶片的光合作用产生物质和能量[8],叶片大小和形状影响植物的气体交换和蒸腾作用[9],有关植物叶片和气孔性状的研究可间接反映植物的植物生长发育及生理代谢特征[10]。Liao等[11]分析了‘哲引3号杨’ × ‘北京杨’全同胞杂种三倍体和二倍体的苗期表型,发现三倍体群体整体具有生长和光合优势,其中也存在一些光合效率高但生长较差的个体和光合效率低但生长较好的个体,可见,异源三倍体杨树的性状变异规律非常复杂。已有研究表明,雌雄异株植物在叶面积、生长势、生物量等方面存在差异[12],性状表现也会受到性别的影响。然而,关于叶片及气孔性状的研究多集中在苗期植株的分析,尚缺少对于节间生长缓慢且已进入开花成熟期的杨树枝条上的短枝功能叶片及气孔性状的变异规律的研究,且对于异源三倍体杨树,性别效应对性状表现产生的影响尚不明确。因此,为进一步丰富杨树三倍体叶片及气孔性状的遗传变异特征,本研究以‘哲引3号杨’ × ‘北京杨’全同胞杂种二倍体和三倍体的10年生种质保存林为材料,对其大树短枝功能叶片及气孔性状变异规律进行系统研究,解析倍性、基因型和性别效应对叶片及气孔性状变异的影响大小,为杨树三倍体育种提供理论依据。
1. 材料与方法
1.1 试验材料
试验材料采集自河北威县的‘哲引3号杨’ × ‘北京杨’全同胞杂种二倍体和三倍体种质保存林,4株小区,3次重复,树龄10年,其中包括二倍体无性系17个(雌株7个,编号为D9、D24、D25、D34、D44、D45、D52;雄株10个,编号为D2、D3、D4、D11、D14、D20、D21、D38、D41、D43),三倍体无性系19个(雌株9个,编号为T6、T12、T15、T16、T19、T23、T26、T32、T40;雄株10个,编号为T13、T18、T22、T27、T30、T31、T33、T36、T37、T42)。每个无性系从每小区的4株中随机选择1株作为采样树,共3次重复,采集位于树冠中部北侧的无病虫害短枝功能叶片进行叶片长度、叶片宽度、叶面积、叶柄长度、叶缘锯齿数、气孔长度、气孔宽度、气孔密度等性状测量。
1.2 试验方法
1.2.1 短枝叶片性状测量
每株采样树随机选取30片形态完整的短枝叶片,每个无性系重复3株采样树,用CI-203便携式激光叶面积仪(美国CID)测量叶片长度、叶片宽度、叶面积和叶柄长度,并对叶缘锯齿数进行统计。
1.2.2 气孔性状
每株采样树随机选取5片形态完整的短枝叶片,每个无性系重复3株采样树,采用指甲油涂抹撕取法进行气孔性状观察[13]。每个叶片随机选择50个处于关闭状态的气孔,于Olympus BX51显微镜下测量气孔长度和气孔宽度;随机选择10个视野统计气孔密度。
1.2.3 数据统计分析
利用Excel整理数据,由于各性状数据均不满足正态分布或方差齐次,应用SPSS 17.0软件的非参数Kruskal−Wallis检验进行差异显著性分析。应用R语言对各性状进行Spearman相关分析和方差贡献率的计算,将方差分量的剩余项视为环境效应。
无性系重复力(R)计算公式:
R=σc2/(σc2+σe2/k) 式中:σc2为无性系方差,σe2为随机误差,k为每个无性系采集样本数的调和平均值[14]。
2. 结果与分析
2.1 青黑杨全同胞杂种短枝叶片及气孔性状基因型间的变异分析
青黑杨全同胞杂种植株短枝叶片及气孔各性状在无性系之间均存在极显著差异(表1)。无性系之间叶片性状变异较大,变异系数介于14.23% ~ 27.20%之间,其中锯齿数变异系数最高,叶片宽度变异系数最小;气孔性状变异系数在8.73% ~ 32.73%之间,其中气孔密度变异系数最高,气孔宽度变异系数最低,表明叶片宽度和气孔宽度是所测性状中较稳定的遗传特征。
表 1 ‘哲引3号杨’ × ‘北京杨’杂种二倍体和三倍体叶片及气孔性状无性系重复力及变异系数Table 1. Repeatability and variation coefficients of leaves and stomatal traits of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis性状 Trait 二倍体 Diploid 三倍体 Triploid P值 P value 无性系重复力 Repeatability of clone 变异系数 CV/% 变化范围
Variation range均值 ± 标准误 Mean ± SE 变化范围
Variation range均值 ± 标准误 Mean ± SE 倍性 Ploidy 性别Gender 基因型Genotype 叶片长度
Leaf length/cm6.26 ~ 9.35 7.52 ± 0.14 6.59 ~ 10.51 8.65 ± 0.18 < 0.01 0.86 < 0.01 0.903 15.23 叶片宽度
Leaf width/cm4.83 ~ 7.48 6.05 ± 0.13 5.45 ~ 7.92 6.85 ± 0.11 < 0.01 0.33 < 0.01 0.836 14.23 叶面积
Leaf area/cm217.76 ~ 39.92 27.32 ± 0.97 21.61 ~ 41.97 33.41 ± 1.09 < 0.01 0.72 < 0.01 0.789 25.32 叶柄长度
Petiole length/cm3.32 ~ 5.33 4.35 ± 0.10 3.28 ~ 5.95 4.48 ± 0.12 0.41 0.29 < 0.01 0.829 16.95 锯齿数
Serration number19 ~ 74 42.1 ± 2.0 30 ~ 55 38.4 ± 1.1 0.10 0.34 < 0.01 0.924 27.20 气孔长度
Stomatal length/μm20.67 ~ 28.29 23.58 ± 0.30 24.08 ~ 30.66 27.59 ± 0.30 < 0.01 0.79 < 0.01 0.954 11.13 气孔宽度
Stomatal width/μm12.82 ~ 16.17 14.53 ± 0.14 14.15 ~ 17.66 16.27 ± 0.16 < 0.01 0.28 < 0.01 0.952 8.73 气孔密度/(个·mm−2)
Stomatal density/
(number·mm−2)87 ~ 279 188.6 ± 8.1 95 ~ 191 128.5 ± 3.8 < 0.01 0.06 < 0.01 0.947 32.73 重复力作为一个重要的遗传参数,反映了无性系优良性状受特殊环境影响的程度[15],其大小还表明了性状稳定性的高低[16]。本研究中,叶片性状无性系重复力介于0.789 ~ 0.924之间,气孔性状无性系重复力介于0.947 ~ 0.960之间,气孔性状的遗传效应略大于叶片性状。可见,青黑杨全同胞杂种植株叶片和气孔性状受到强度的遗传控制,环境影响较小,有利于无性系的评价选择,更利于较为准确地解析各效应对性状变异的影响。
2.2 青黑杨全同胞杂种植株短枝叶片性状变异规律分析
不同倍性群体间比较,三倍体植株平均叶片长度、叶片宽度、叶面积均显著大于二倍体植株,呈现出一定的巨大性特征(表1,图1a、b);而叶柄长度和锯齿数性状则无显著差异(表1)。在不同性别间,尽管雄株的叶片宽度、叶面积和叶柄长度比雌株分别大3.14%、2.02%和3.70%,叶片长度和锯齿数分别小1.10%和1.97%,但是这些叶片性状在雌、雄株之间并未呈现显著差异。
![]() 图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis
图 1 ‘哲引3号杨’ × ‘北京杨’杂种全同胞二倍体和三倍体植株叶片和气孔a. 二倍体D44叶片;b. 三倍体T22叶片;c. 二倍体植株气孔密度和气孔大小D44;d. 三倍体植株气孔密度和气孔大小T22。 a, a leaf from diploid D44; b, a leaf from triploid T22; c, stomata density and stomatal size of diploid D44; d, stomata density and stomatal size of triploid T22.Figure 1. Leaves and stomata of diploid and triploid plants of (P. pseudo-simonii × P. nigra ‘Zheyin3#’) × P. × beijingensis从叶片各性状相关分析可知(图2),叶面积与叶片长度、叶片宽度呈极显著的正相关,分别达0.86和0.94;叶柄长度与叶片长度、叶片宽度、叶面积之间呈显著弱正相关,与锯齿数相关关系不显著;锯齿数与叶片宽度、叶面积呈显著弱正相关,与叶片长度的相关关系不显著。进一步分析发现(表2),叶片长度、叶片宽度和叶面积与倍性水平呈极显著的正相关(r分别为0.44、0.43、0.38,P < 0.01),叶柄长度和锯齿数与倍性水平之间无显著相关性(分别为r = 0.09,P = 0.41和r = −0.17,P = 0.10),而性别与叶片性状之间均没有显著的相关性(P > 0.05)。
![]() 图 2 叶片和气孔性状的Spearman相关分析*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.Figure 2. Spearman correlation analysis of leaves and stomatal traits表 2 叶片及气孔性状与倍性、性别的相关性分析Table 2. Correlation analysis of leaves and stomatal traits with ploidy and gender
图 2 叶片和气孔性状的Spearman相关分析*、**分别表示在P < 0.05 和P < 0.01水平上差异显著。*, ** represent significant difference at P < 0.05 and P < 0.01 level, respectively.Figure 2. Spearman correlation analysis of leaves and stomatal traits表 2 叶片及气孔性状与倍性、性别的相关性分析Table 2. Correlation analysis of leaves and stomatal traits with ploidy and gender项目
Item叶片长度
Leaf length叶片宽度
Leaf width叶面积
Leaf area叶柄长度
Petiole length锯齿数
Serration number气孔长度
Stomatal length气孔宽度
Stomatal width气孔密度
Stomatal density倍性 Ploidy r值 r value 0.44 0.43 0.38 0.09 −0.17 0.72 0.65 −0.60 P值 P value < 0.01 < 0.01 < 0.01 0.41 0.10 < 0.01 < 0.01 < 0.01 性别 Gender r值 r value −0.02 0.10 0.04 0.11 −0.10 0.03 −0.11 0.20 P值 P value 0.86 0.33 0.72 0.29 0.34 0.80 0.28 0.06 2.3 青黑杨全同胞杂种植株短枝叶气孔性状变异规律分析
三倍体植株的平均气孔长度和气孔宽度均极显著大于二倍体,而平均气孔密度极显著小于二倍体(表1,图1c、d)。在不同性别间比较发现,雄株群体的平均气孔长度和气孔密度均大于雌株,气孔宽度小于雌株,但是均为呈现显著差异。
从气孔各性状的相关分析可知(图2),气孔长度与气孔宽度呈极显著的正相关,达0.89,气孔密度与气孔长度和气孔宽度呈极显著的负相关,分别为−0.84和−0.76。进一步分析发现(表2),气孔长度和气孔宽度与倍性水平间均呈极显著正相关(r分别为0.72和0.65,P < 0.01),气孔密度与倍性水平呈极显著负相关(r = −0.60,P < 0.01),而性别与气孔性状之间均没有显著的相关性(P > 0.05),表明气孔性状与倍性水平密切相关,可作为鉴定杨树多倍体的有效指标。
2.4 影响青黑杨全同胞杂种叶片及气孔性状各效应的方差贡献率解析
为深入解析倍性、性别和基因型效应对各性状表现的影响程度,计算各效应的方差分量及方差贡献率后发现(图3),气孔性状的变异主要来源于倍性效应,其次是基因型效应和环境效应。其中,气孔长度的倍性效应方差分量在表型总变异中所占比例高达63.79%,其次是气孔宽度的57.05%,气孔密度的47.10%,表明这些性状受倍性水平的影响最大;叶片性状变异的主要来源于基因型效应,其中,锯齿数变异的基因型效应方差贡献率最高,达80.72%,其次是叶柄长度变异的基因型效应为61.66%,表明叶片性状主要受基因型的影响。性别效应对叶片及气孔性状的影响均很小,其中,性别效应对气孔密度的方差贡献率仅为0.06%,对锯齿数和叶柄长度变异的方差贡献率甚至均为0。
3. 讨 论
多倍化是植物进化和适应性改变的重要力量[17-18]。由于基因组剂量的增加所带来的倍性效应往往导致多倍体植物产生广泛的性状变异[19-20]。同源四倍体苏丹凤仙花(Impatiens walleriana)在叶面积、叶片厚度、子房宽度、花粉粒长度和气孔大小等性状上增加,花瓣总花青素含量较高,而株高、气孔保卫细胞密度和花数减少[21]。滇杨(P. yunnanensis)同源四倍体和嵌合体的苗期长枝叶分析发现,与叶片长度、宽度和气孔性状大于二倍体,而气孔密度则显著小于二倍体[22]。本研究发现青黑杨杂种的叶柄长度和锯齿数变异受倍性效应影响较小,而三倍体植株平均叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度均极显著大于二倍体,气孔密度则极显著小于二倍体,且气孔性状变异的倍性效应方差贡献率最高,表明倍性效应对短枝叶的性状表现同样产生重要影响。而且,叶片长度、叶片宽度、叶面积、气孔长度和气孔宽度与倍性水平之间均存在极显著正相关性,气孔密度与倍性水平呈极显著负相关,叶柄长度和锯齿数与倍性水平无显著相关性,因此,在倍性检测时可通过叶片大小和气孔性状相结合作为初步鉴定。此外,值得注意的是,本研究测得的气孔长度和气孔宽度大小与王君等[23]测得的数据相比均偏小,可能是由于杨树长枝和短枝叶片的气孔长宽大小存在差异。
基因型差异也是影响植物性状的重要变异来源。白凤莹等[24]发现,受基因型的影响,不同天然三倍体毛白杨无性系间气孔性状的差异达到了极显著水平,其中存在部分个体表现出类似二倍体的特征。本研究发现,青黑杨全同胞杂种无性系间短枝叶片和气孔各性状均存在极显著差异,其中部分二倍体无性系的叶片和气孔性状表现甚至优于一些三倍体无性系,可见三倍体也并非株株皆优,无性系基因型对性状表现的影响也较大。
对于雌雄异株植物而言,植株性别会对表型产生一定的影响。相关研究表明,多年生植物雄株在叶片数量及叶面积、生长势、树冠体积、生物量等方面都超过雌株[12, 25]。陈珂等[26]发现成年银杏(Ginkgo biloba)雌株叶片长度、叶片宽度、叶面积显著小于雄株,而叶柄长度和气孔密度显著大于雄株。两年生青杨(P. cathayana)雌株叶片总数显著高于雄株,雌株叶片性状较雄株有优势[27];毛白杨雄株叶片长度和叶面积显著大于雌株,而叶片宽度则无显著差异[28]。本研究中,青黑杨全同胞杂种叶片和气孔性状在雌、雄株之间虽存在一定差异,但均未达显著性水平,性状变异与性别之间也不存在显著相关性,且性别效应方差贡献率几乎为0,说明性别对叶片及气孔性状的影响较小。
异源多倍化在植物遗传改良和种质创新中发挥着重要作用,特别是利用2n配子杂交的异源多倍化途径,结合了倍性优势和杂种优势,在杨树多倍体育种已取得了极大的成就[29]。Liao等[11]发现,杨树异源三倍体群体苗期整体具有生长和光合优势,但也存在一些光合效率高但生长较差的个体和光合效率低但生长较好的个体。本研究利用一个包含杂种二倍体和三倍体的青黑杨全同胞群体,证明了杨树经历异源多倍化后,倍性效应和基因型效应均对性状变异产生重要影响,而性别对性状变异的贡献相对较小,并初步明确了倍性、基因型和性别效应对功能叶片和气孔性状的贡献大小,为我们进一步开展多倍体性状变异的遗传解析提供了借鉴。显然,在进行杨树三倍体育种时,对大量变异材料进行“大群体,强选择”是进一步品种选育的必要环节,异源三倍体遗传变异的复杂性也为杨树三倍体选育提供了丰富的选择基础。
-
![]()
图 1 胡杨异形叶·O2−产生速率和MDA含量的变化
图中误差棒为平均值的标准偏差;同一月份内不同小写字母表示不同叶形在P < 0.05水平差异显著。下同。Error bars in figure represent standard deviation of the average value; different lowercase letters in the same month indicate significant difference among heteromorphic leaves at P < 0.05 level. The same as below.
Figure 1. Changes of·O2− production rate and MDA content of heteromorphic leaves of P. equhratica
![]()
图 2 胡杨异形叶抗氧化酶活性的变化
Figure 2. Changes of antioxidant enzyme activities of heteromorphic leaves of P. equhratica
-
[1] 王世绩, 陈炳浩, 李护群. 胡杨林[M]. 北京: 中国环境科学出版社, 1995: 141−144. Wang S J, Chen B H, Li H Q. Populus euphratica forest[M]. Beijing: China Environmental Science Press, 1995: 141−144.
[2] 吴平, 赵健, 王文丽, 等. 额济纳绿洲胡杨生物生态学特性的调查[J]. 内蒙古草业, 2005, 17(3):4−6. doi: 10.3969/j.issn.2095-5952.2005.03.002 Wu P, Zhao J, Wang W L, et al. Investigation on the biological and ecological characteristics of Populus euphratica in Ejina Oasis[J]. Inner Mongolia Prataculture, 2005, 17(3): 4−6. doi: 10.3969/j.issn.2095-5952.2005.03.002
[3] Li Z X, Zheng C X. Structural characteristics and eco-adaptability of heteromorphic leaves of Populus euphratica[J]. Forestry Studies in China, 2005, 7(1): 11−15. doi: 10.1007/s11632-005-0050-8
[4] Liu Y, Li X, Chen G, et al. Epidermal micromorphology and mesophyll structure of Populus euphratica heteromorphic leaves at different development stages[J/OL]. PloS One, 2015, 10(9): e137701 (2018−05−24) [2018−03−01]. http://doi.org/10.1371/journal.pone.0137701.
[5] Hao J Q, Yue N, Zheng C X. Analysis of changes in anatomical characteristics and physiologic features of heteromorphic leaves in a desert tree, Populus euphratica[J/OL]. Acta Physiologiae Plantarum, 2017, 39(8) (2017−07−10) [2018−05−20]. https://dor.org/10.1007/s11738-017-246-9.
[6] 陈少良, 李金克, 王天华, 等. 氯盐胁迫下胡杨木质部汁液ABA、离子浓度和叶片气体交换的变化[J]. 植物学报, 2003, 45(5):561−566. doi: 10.3321/j.issn:1672-9072.2003.05.009 Chen S L, Li J K, Wang T H, et al. Gas exzhange, xylem ions and abscisic acid response to Na+-salts and Cl−-salts in Populus equhratica[J]. Acta Botanica Sinica, 2003, 45(5): 561−566. doi: 10.3321/j.issn:1672-9072.2003.05.009
[7] 郭学民, 刘挨枝, 王华芳. 胡杨种子微形态结构特征及其耐旱性[J]. 河北科技师范学院学报, 2013, 27(1):23−26. doi: 10.3969/J.ISSN.1672-7983.2013.01.005 Guo X M, Liu A Z, Wang H F. Morphological characteristics of seed surface of Populus equhratica under scanning electron microscope and their drought resistance[J]. Collge of Life Science and Technology, Hebei Nonral University of Science & Technology, 2013, 27(1): 23−26. doi: 10.3969/J.ISSN.1672-7983.2013.01.005
[8] 姚琨, 练从龙, 王菁菁, 等. 胡杨PePEX11基因参与调节盐胁迫下拟南芥的抗氧化能力[J]. 北京林业大学学报, 2018, 40(5):19−28. Yao K, Lian C L, Wang J J, et al. PePEX11 functions in regulating antioxidant capacity of Arabidopsis thaliana under salt stress[J]. Journal of Beijing Forestry University, 2018, 40(5): 19−28.
[9] 夏振华, 陈亚宁, 朱成刚, 等. 干旱胁迫环境下的胡杨叶片气孔变化[J]. 干旱区研究, 2018, 35(5):1111−1117. Xia Z H, Chen Y N, Zhu C G, et al. Stomatal change in leaves of Population euphratica under drought stress[J]. Arid Zone Research, 2018, 35(5): 1111−1117.
[10] 白雪, 张淑静, 郑彩霞, 等. 胡杨多态叶光合和水分生理的比较[J]. 北京林业大学学报, 2011, 33(6):47−52. Bai X, Zhang S J, Zheng C X, et al. Comparative study on photosynthesis and water physiology of polymorphic leaves of Populus equhratica[J]. Journal of Beijing Forestry University, 2011, 33(6): 47−52.
[11] Zheng C X, Qiu J, Jiang C N, et al. Comparison of stomatal characteristics and photosynthesis of polymorphic Populus euphratica leaves[J]. Frontiers of Forestry in China, 2007, 2(1): 87−93. doi: 10.1007/s11461-007-0014-3
[12] 郝建卿, 吕娜, 杨扬, 等. 内蒙乌海胡杨异形叶水分及叶绿素荧光参数的比较[J]. 北京林业大学学报, 2010, 32(5):41−44. Hao J Q, Lü N, Yang Y, et al. Comparative study of chlorophyll fluorescence parameters and water physiological characters of heteromorphic leaves for Populus euphratica[J]. Journal of Beijing Forestry University, 2010, 32(5): 41−44.
[13] 蒋少伟, 周多多, 吴桂林, 等. 不同地下水埋深下胡杨枝条水力导度及其季节变化[J]. 干旱区研究, 2017, 34(3):648−654. Jiang S W, Zhou D D, Wu G L, et al. Hydraulic conductivity and its seasonal variation of Populus euphratica shoot at the sites with varying groundwater depths[J]. Arid Zone Research, 2017, 34(3): 648−654.
[14] Li W H, Zheng C X, He S H. The analysis of xylem vessel structure of Populus euphratica Oliv. fresh branch by HRCT[J]. Communications in Soil Science and Plant Analysis, 2018, 50(1): 10−19.
[15] 岳宁. 胡杨异形叶蛋白质表达的电泳分析[J]. 安徽农业科学, 2010, 38(32):18058−18061. doi: 10.3969/j.issn.0517-6611.2010.32.029 Yue N. Electrophoresis analysis on the expressed proteins in heteromorphic leaves of Populus euphratica Oliv.[J]. Journal of Anhui Agricultural Sciences, 2010, 38(32): 18058−18061. doi: 10.3969/j.issn.0517-6611.2010.32.029
[16] Zhang J, Jia H X, Li J B, et al. Molecular evolution and expression divergence of the Populus euphratica Hsf genes provide insight into the stress acclimation of desert poplar[J/OL]. Scientific Reports, 2016, 6 (2016−07−18) [2018−06−21]. https://dor.org/10.1038/srep30050.
[17] 秦少伟, 李才林, 郭忠忠, 等. 参与胡杨异形叶发生的发育相关基因表达模式及作用[J]. 基因组学与应用生物学, 2017, 36(11):4785−4792. Qin S W, Li C L, Guo Z Z, et al. Expression patterns and functions of genes related to the development of Populus euphratica Oliv. heteromorphic leaves morphogenesis[J]. Genomics and Applied Biology, 2017, 36(11): 4785−4792.
[18] Mittler R. ROS are good[J]. Trends in Plant Science, 2016, 22(1): 11−19.
[19] Sharma S S, Dietz K J. The relationship between metal toxicity and cellular redox imbalance[J]. Trends in Plant Science, 2009, 14(1): 43−50. doi: 10.1016/j.tplants.2008.10.007
[20] Anjum N A, Sofo A, Scopa A, et al. Lipids and proteins-major targets of oxidative modifications in abiotic stressed plants[J]. Environmental Science and Pollution Research, 2015, 22(6): 4099−4121. doi: 10.1007/s11356-014-3917-1
[21] 郑清岭, 杨忠仁, 张凤兰, 等. 沙芥属植物活性氧清除系统对干旱胁迫的响应[J]. 西北植物学报, 2018, 38(9):1674−1682. Zheng Q L, Yang Z R, Zhang F L, et al. Responses of reactive oxygen species scavenging system to drought stress in Pugionium Gaertn.[J]. Acta Botanica Boreali-Occidentalia Sinica, 2018, 38(9): 1674−1682.
[22] 常二梅, 史胜青, 刘建锋, 等. 古侧柏针叶活性氧产生及其清除机制[J]. 东北林业大学学报, 2011, 39(11):8−11. doi: 10.3969/j.issn.1000-5382.2011.11.003 Chang E M, Shi S Q, Liu J F, et al. ROS production and its elimination in old Platycladus orientalis’s leaves[J]. Journal of Northeast Forestry University, 2011, 39(11): 8−11. doi: 10.3969/j.issn.1000-5382.2011.11.003
[23] 施征, 史胜青, 姚洪军, 等. 植物线粒体中活性氧的产生及其抗氧化系统[J]. 北京林业大学学报, 2009, 31(1):150−154. doi: 10.3321/j.issn:1000-1522.2009.01.026 Shi Z, Shi S Q, Yao H J, et al. Production of ROS and its antioxidant mechanism in plant mitochondria[J]. Journal of Beijing Forestry University, 2009, 31(1): 150−154. doi: 10.3321/j.issn:1000-1522.2009.01.026
[24] 邵旭平, 万建宏, 万东石. 不同生态型胡杨异形叶对虫害的抗氧化反应[J]. 草业科学, 2011, 28(7):1396−1399. Shao X P, Wan J H, Wan D S. The anti-xidation responses of two ecotype leaves of diversifolious poplar (Populus euphratica) to pest[J]. Pratacultural Science, 2011, 28(7): 1396−1399.
[25] 王海珍, 徐雅丽, 张翠丽, 等. 干旱胁迫对胡杨和灰胡杨幼苗渗透调节物质及抗氧化酶活性的影响[J]. 干旱区资源与环境, 2015, 29(12):125−130. Wang H Z, Xu Y L, Zhang C L, et al. Effects of drought stress on osmotic adjustment substances and antioxidant enzymes activity of Populus euphratica and Populus pruinosa seedlings[J]. Journal of Arid Land Resources & Environment, 2015, 29(12): 125−130.
[26] 王爱国, 罗广华. 植物的超氧物自由基与羟胺反应的定量关系[J]. 植物生理学报, 1990, 26(6):55−57. Wang A G, Luo G H. Quantiative relation between the reaicton of hydroxylamine and superoxide anion radicals in plants[J]. Plant Physiology Journal, 1990, 26(6): 55−57.
[27] Heath R L, Lester P. Effect of light on lipid peroxidation in chloroplasts[J]. Biochemical and Biophysical Research Communications, 1965, 19(6): 716−720. doi: 10.1016/0006-291X(65)90316-5
[28] Giannopolitis C N, Ries S K. Superoxide dismutases: Ⅰ. occurrence in higher plants[J]. Plant Physiology, 1977, 59(2): 309−314. doi: 10.1104/pp.59.2.309
[29] 陈建勋, 王晓峰. 植物生理学实验指导 [M]. 2版. 广州: 华南理工大学出版社, 2002. Chen J X, Wang X F. Plant physiology experiment instruction[M]. 2nd ed. Guangzhou: South China University of Technology Press, 2002.
[30] Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts[J]. Plant and Cell Physiology, 1981, 22(5): 867−880.
[31] Schaedle M. Chloroplast glutathione reductase[J]. Plant Physiology, 1977, 59(5): 1011−1012. doi: 10.1104/pp.59.5.1011
[32] Kampfenkel K, Vanmontagu M, Inze D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue[J]. Analytical Biochemistry, 1995, 225(1): 165−167. doi: 10.1006/abio.1995.1127
[33] 李玲. 植物生理学模块实验指导[M]. 北京: 科学出版社, 2009: 100−102. Li L. Plant physiology module experiment instruction[M]. Beijing: Science Press, 2009: 100−102.
[34] 冯涛, 曹东旭, 吕晓玲. 竹叶总黄酮含量的测定[J]. 中国食品添加剂, 2002(6):85−87, 35. doi: 10.3969/j.issn.1006-2513.2002.06.022 Feng T, Cao D X, Lü X L. The determination of bamboo leaves total flavonoids content[J]. China Food Additives, 2002(6): 85−87, 35. doi: 10.3969/j.issn.1006-2513.2002.06.022
[35] Li X, Yang Y, Sun X, et al. Comparative physiological and proteomic analyses of poplar (Populus yunnanensis) plantlets exposed to high temperature and drought[J/OL]. PloS One, 2014, 9(9) (2014−09−09) [2018−03−09]. http://doi.org/10.1371/journal.pone.0107605.
[36] Solti A, Sárvári E, Szöllősi E, et al. Stress hardening under long-term cadmium treatment is correlated with the activation of antioxidative defence and iron acquisition of chloroplasts in Populus[J]. Zeitschrift für Naturforschung C, 2016, 71(9−10): 323−334. doi: 10.1515/znc-2016-0092
[37] 张淑静, 王强, 郑彩霞, 等. 文冠果花药发育与活性氧平衡的关系[J]. 农业科学, 2012, 2(3):37−42. doi: 10.3969/j.issn.1009-2196.2012.03.008 Zhang S J, Wang Q, Zheng C X, et al. Studies on the relationship between anther development and reactive oxygen balancing in Xanthoceras sorbifolia Bunge[J]. Hans Journal of Agricultural Science, 2012, 2(3): 37−42. doi: 10.3969/j.issn.1009-2196.2012.03.008
[38] 马剑, 刘贤德, 孟好军, 等. 水分胁迫对文冠果幼苗生长及生理特性的影响[J]. 干旱区资源与环境, 2018, 32(1):128−132. Ma J, Liu X D, Meng H J, et al. Effects of water stress on growth and physiological characteristics of seedlings of the Xanthoceras sorbifolia[J]. Journal of Arid Land Resources & Environment, 2018, 32(1): 128−132.
[39] Saini R K, Keum Y S. Progress in microbial carotenoids production[J]. Indian Journal of Microbiology, 2017, 57(1): 129−130. doi: 10.1007/s12088-016-0637-x
[40] Nan J, Andrea D, Erich G. Flavones: from biosynthesis to health benefits[J/OL]. Plants, 2016, 5(2): 27 (2016−06−15)[2017−10−23]. https://dor.org/10.3390/plants5020027.
[41] Gest N, Gautier H, Stevens R. Ascorbate as seen through plant evolution: the rise of a successful molecule[J]. Journal of Experimental Botany, 2013, 64(1): 33−53. doi: 10.1093/jxb/ers297
[42] Gallie D R. The role of L-ascorbic acid recycling in responding to environmental stress and in promoting plant growth[J]. Journal of Experimental Botany, 2013, 64(2): 433−443. doi: 10.1093/jxb/ers330
[43] 韩刚, 党青, 赵忠. 干旱胁迫下沙生灌木花棒的抗氧化保护响应研究[J]. 西北植物学报, 2008, 28(5):1007−1013. doi: 10.3321/j.issn:1000-4025.2008.05.024 Han G, Dang Q, Zhao Z. Response of antioxidation protection system of Hedysarum scoparium to drought stress[J]. Acta Botanica Boreali-Occidentalia Sinica, 2008, 28(5): 1007−1013. doi: 10.3321/j.issn:1000-4025.2008.05.024
[44] 张涛, 安黎哲, 陈拓, 等. 不同海拔青海云杉与祁连圆柏叶片抗氧化系统[J]. 植物生态学报, 2009, 33(4):802−811. doi: 10.3773/j.issn.1005-264x.2009.04.019 Zhang T, An L Z, Chen T, et al. Antioxidative system in leaves of Picea crassifolia and Sabina przewalskii along an altitudinal gradient[J]. Chinese Journal of Plant Ecology, 2009, 33(4): 802−811. doi: 10.3773/j.issn.1005-264x.2009.04.019
-
期刊类型引用(4)
1. 赵钰婷,陈冬瑶,杨柳,李晶楠,宁广亮,姜静. 白桦四倍体×紫雨桦二倍体杂交种子活力及杂种子代生长特性分析. 温带林业研究. 2025(01): 1-8 .  百度学术
百度学术
2. 马鸿文,任宇昕,龙羿辛,王楠,冯祥元,俞天泉,华晓琴,王君. 青黑杨杂种全同胞二倍体与三倍体长枝叶性状变异研究. 北京林业大学学报. 2024(01): 27-34 . 本站查看
3. 王舒扬,田力,周顺陶,储月娥,梅迪,袁佳秋,余延浩,洑香香. 多倍化对青钱柳叶形态、光合性能和次生代谢产物积累的影响. 林业科学. 2024(08): 120-131 . 百度学术
4. 卞瑶,耿丽妍,冯彦博,陈福燕,张雯,王宏伟. 不同进化历程下的异源四倍体小麦核型进化及其稳定性分析. 辽宁师范大学学报(自然科学版). 2023(03): 363-369 . 百度学术
其他类型引用(5)

 下载:
下载:




计量
- 文章访问数: 2506
- HTML全文浏览量: 811
- PDF下载量: 57
- 被引次数: 9
