
Partitioning of water vapor flux on the ecosystem scale: a case study on larch boreal forests
-
摘要:目的蒸散发(ET)包括蒸发(E)和蒸腾(T), 是生态系统降雨(P)返回大气的最主要形式, 在气候变化背景下, 了解大兴安岭北部多年冻土区的寒温带兴安落叶松林的ET特征及其分配状况, 有助于进一步理解北方森林对气候变化的响应模式。方法在2015年7月10日至8月10日期间, 利用模型与野外实测的方法对寒温带兴安落叶松林蒸发(E)、蒸腾(T)及蒸散发(ET)进行研究。E包括林地蒸发(Ef)和林冠截留(Ec), 而林分蒸腾总量(Ttot)则为优势木(Td)、中等木(Ti)、劣势木(Ts)蒸腾量之和。分析非降水和降水日的ET及其组分特征和分配, 探讨水汽通量对气象因子的响应。结果非降雨和降雨日的ET及其组分的日变化均呈单峰格局, 且非降雨日曲线的日峰值均高于降雨日。非降雨日, Ef、Td、Ti、Ts和ET分别为10.3、25.6、15.2、10.8和66.3mm; 降雨日, Ef、Ec、Td、Ti、Ts和ET则分别为2.2、24.3、11.2、5.1、3.8和47.8mm。非降雨日, Ef/ET为15.5%, 而Ttot/ET为78.0%, 其中Td/ET、Ti/ET和Ts/ET分别贡献38.7%、23.0%和16.4%;降雨日, Ef/ET低至4.6%, Ec/ET则可以达到50.9%, 而Ttot/ET降低至42.2%, 其中Td/ET、Ti/ET和Ts/ET分别为23.5%、10.6%和8.0%;表明非降雨日ET以T为主(具体为Td), 降雨日则以E(具体为Ec)为主。观测期间94.7%的P主要以ET形式返回大气, 其中由T贡献57%, E贡献38%。总体上, 无论降雨与否, ET与23m处净辐射(Rn)的相关性均高于其与水汽压亏缺(VPD)的相关性, Ttot与二者的相关性则差异不大, 而Ef的表现则与ET相反, 说明Rn是生态系统能量循环和物质交换的最主要驱动力, Ttot同时主要受到Rn和VPD的约束, 而Ef优先受VPD的限制。结论兴安落叶松优势木的蒸腾能力强于中等木和劣势木, 以往研究多采用Td(或包括较大径级的Ti)为林分尺度上推计算过程的基准值的方法会高估林分整体的蒸腾能力, 实际误差的大小取决于林分的分化程度以及是否降雨等因素。非降雨日的气象条件更有利于植被-大气界面的水汽交换, 降雨的发生会影响生态系统ET的分配模式。Abstract:ObjectiveEvapotranspiration (ET) including evaporation (E) and transpiration (T), was the main way that ecosystem precipitation (P) returned to the atmosphere. Under the background of climate change, understanding the characteristics and distribution of ET in a cold-temperate Larix gmelinii natural forest at permafrost region of northern part of Daxing'an Mountains, northeastern China could help to further comprehend the response modes of the boreal forest ecosystem to the climate change.MethodModel simulation and field experiments were applied for estimating and measuring ecosystem scale evaporation (E), transpiration (T), and evapotranspiration (ET) in a boreal larch forest. E includes forest floor evaporation (Ef) and canopy interception (Ec) which only occurred during rainfall events. Total forest transpiration (Ttot) in this study is the sum of dominant tree transpiration (Td), intermediate tree transpiration (Ti) and suppressed tree transpiration (Ts). Furthermore, we analyzed the variations and allocation proportions of ET and components under conditions of non-rainy or rainy. Then we discussed the responses of Ef, Ttot and ET to the net radiation (Rn) and the vapor pressure deficit (VPD) under different water input conditions.ResultDiurnal variations of ET and its components all performed as single peak patterns in both non-rainy and rainy days, and peak value in each curve at non-rainy day grouping was higher than in rainy day grouping. Ef, Td, Ti, Ts and ET in non-rainy days were 10.3, 25.6, 15.2, 10.8 and 66.3mm, respectively. Meanwhile, Ef, Ec, Td, Ti, Ts and ET in rainy days were 2.2, 24.3, 11.2, 5.1, 3.8 and 47.8mm, respectively.In non-rainy days, Ef/ET was 15.5%, and Ttot/ET was 78.0%, while Td/ET, Ti/ET and Ts/ET contributed 38.7%, 23.0% and 16.4%, respectively.In rainy days, Ef/ET could go low to 4.6%, Ec/ET reached 50.9%, and Ttot/ET decreased to 42.2%, in which Td/ET, Ti/ET and Ts/ET were 23.5%, 10.6% and 8.0%, respectively.These results represented that ET was dominated by T (specifically for Td) in non-rainy days. However, E (specifically for Ec) contributed the highest proportion of ET in rainy days. 94.7% of P returned to the atmosphere through ET during whole observation days, there into, T and E accounted for nearly 57% and 38%, respectively. Overall, without regard to rainfall events, ET had better correlation with net radiation at 23m height (Rn) than with VPD and Ttot owned similar correlations with both Rn and VPD, but Ef expressed opposite result with ET. This result demonstrated that Rn was obviously the main driver for ecosystem's energy cycle and material exchange, Ttot restrained by Rn and VPD at the same time, and Ef was more susceptible to VPD compared to Rn.ConclusionThe transpiration capacity of dominant trees of Larix gemelinii species was stronger than intermediate and suppressed trees. So that, forest transpiration upscale from the individual tree could be overestimated if this method only considers in situ measured Td (even including Ti with larger DBH) and ignores Ts. The actual error depends on the degree of forest differentiation and moisture income conditions. The meteorological condition of non-rainfall days is more propitious to the water-vapor exchange at vegetation-atmosphere interface, and the occurrence of rainfall will affect the distribution pattern of ecosystem ET.
-
Keywords:
- boreal forest /
- evaporation /
- canopy interception /
- transpiration /
- evapotranspiration
-
森林生态系统的降水(P, precipitation)分配去向主要包括地表径流、土壤入渗和蒸散发(ET, evapotranspiration)[1]。ET由蒸发(E, evaporation)和蒸腾(T, transpiration)两个过程主导[2-3],是生态系统水分返回大气的最主要形式[4],也是地表能量平衡和全球水循环的重要组分[1, 5]。E包括林地蒸散发(Ef)和林冠截留(Ec)[6-7]。T则包括林下植被和乔木的蒸腾耗水,其是最大的地表水汽来源,在全球尺度上约占陆地ET总量的80%~90%[3]。同时,天然林中林下植被蒸腾远低于乔木[8-9],在生态系统尺度下常忽略不计[6-7, 10-11]。不同森林生态系统的降水量通过ET的形式返还大气的比例存在差异,例如:热带海洋气候的加勒比松(Pinus caribaea)人工林可高达93.8%[12],寒温带半湿润森林草原气候的青海云杉(Picea crassifolia)天然林为82.7%[11],而温带森林气候的日本扁柏(Chamaecyparis obtusa)人工林仅为47.5%[6]。此外,从ET组分的分配角度,Lin等[13]发现亚高山气候的冷杉(Abies fabri)林生长季(4—10月)Ec/ET超过70%,T次之,Ef仅占ET的6%;Waterloo等[12]发现年尺度上加勒比松林的ET以T为主(70%),Ef和Ec均比例较低(分别为8.5%和20%)。而Schlesinger等[1]通过对全球尺度的T/ET进行整合分析,发现热带雨林最高(70%),北方针叶林次之(65%),地中海灌木林最低(47%)。因此,森林生态系统ET及其分配模式受气候条件、森林生态系统类型、研究的时空尺度等多种生物和非生物因子的综合、交互影响,变异规律和机理复杂,有必要对此开展深入研究。
当前,Ef数据获取主要以间接估算为主[4, 7, 11, 14],原因是直接观测易受降水干扰[7, 14],产生明显误差;Ec、T及ET可分别采用林内外降水差值法[15]、树干液流监测和通量观测[1, 2, 16]进行测算。但以往对T的研究多忽略了林分内小径级的劣势木[17-20],由于不同等级林木T存在较大差异[21-22],在尺度扩展时会造成一定误差。随着气候变化不断加剧,地处高纬度的寒温带森林生态系统的水循环易受到显著影响[23]。此外,本研究以2015年7月10日至8月10日为观测时段,由于研究地的气候特征为冬季寒冷漫长、夏季凉爽短暂,根据以往对同一区域的兴安落叶松(Larix gmelinii)蒸腾特性的研究结果[17],该时间段为兴安落叶松的典型生长季旺盛期,林分的生理生态特征稳定,选择该时段有助于排除展叶期和落叶期林木生理急剧变化对水汽通量定量研究的干扰。该研究的主要目的是以大兴安岭北部兴安落叶松天然林为对象,运用模型模拟与野外观测相结合的手段,对ET及其组分进行测算,并同步测定相关环境因子,研究揭示其水汽通量变异规律及其关键性控制因子,为大尺度上综合分析森林生态系统水循环对气候变化的响应提供理论依据和数据支持。
1. 研究地概况与研究方法
1.1 研究地概况
研究地设在大兴安岭地区漠河的黑龙江漠河森林生态系统定位研究站(53°28′02.94″ N、122°20′16.98″ E),属寒温带大陆性气候,多年冻土区,平均海拔290m,年均温-4.9℃,多年平均降水量350~500mm,全年无霜期85~110d。地带性土壤为棕色针叶林土,地带性植被是以兴安落叶松为优势建群种的明亮针叶林。林内其他常见树种为樟子松(Pinus sylvestris var. mongolica)、白桦(Betula platyphylla)和山杨(Populus davidiana)等;常见林下植物为杜香(Ledum palustre)、笃斯越桔(Vaccinium uliginosum)、兴安杜鹃(Rhododendron dauricum)和越桔(Vaccinium vitisi-idaea)。该研究于2015年7月在通量塔东北侧设置15m × 20m的样地,样地内有兴安落叶松84株、白桦14株、樟子松1株。根据Kljun模型进行通量足迹分析,通量塔的通量贡献区70%和90%的平均距离分别为846和1133m,观测期间的主风向为东西向和东西偏北向,因此通量观测的湍流范围(footprint)包括树干液流法的监测区域。同时,漠河通量塔的搭建考虑了下垫面状况,其观测足迹区域内为几乎无倾斜坡度的兴安落叶松天然林,下垫面的空间异质性较低。
漠河通量观测塔配置一体式三维超声风与红外CO2/H2O分析仪(IRGASON, Campbell Scientific, USA),安装高度36m。梯度气象系统包括:空气温、湿度(HMP45C, Vaisala, Finland)和风速(010C, Met One, USA)传感器,安装高度分别为1.5、9、13、20、28和35m;风向传感器(020C, Met One, USA),安装高度28m;净辐射传感器(NR01, Hukseflux, Netherlands)、光合有效辐射传感器(LI190SB, Li-Cor, USA)、雨滴谱仪(Parsivel2, OTT, Germany)和雨量筒(TE525MM, Texas Electronics, USA),安装高度23m。大气压传感器(CS100, Campbell Scientific, USA),安装高度23m。土壤热通量传感器(HFP01, Hukseflux, Netherlands),分别安装于距通量塔东、西两侧10m远,距离地面10cm深处;土壤水分、温度和电导率传感器(CS650, Campbell Scientific, USA),安装于距通量塔东侧10m远,距离地面5、10、20和40cm深处。采用CR3000数据采集器(Campbell Scientific, USA)以10Hz频率记录通量的观测数据;气象数据每隔30min保存其平均数据;所有监测仪器均同步运行。
1.2 林分特征
采用生长锥(CO200, Haglof, Sweden)对林内60株落叶松样树自胸高处取深至髓心的树芯;处理后利用WinDENDRO年轮分析软件(Regent Instruments Inc., Canada)估测树龄和边材宽度。胸径5~10cm林木树龄为(37.3 ± 11.8)年,10~20cm为(43.7±9.5)年,20~25cm为(53.4 ± 8.7)年。兴安落叶松天然林林龄差在一个龄级内(20年),可视为相对同龄林[24]。同龄林中,林木等级主要受林木个体竞争优势及空间生态位决定[25],采用相对直径法[24-25]划分林木等级,优势木、中等木和劣势木数量分别为28、30和25株。选择不同等级林木各3株作为树干液流观测样树(表 1)。
表 1 液流测定样木特征Table 1. Sample tree features of sap flow measurement林木等级
Forest grade样本数
Sample tree number平均胸径
Average DBH/cm平均树高
Average height/m平均边材面积
Average sapwood area/cm2优势木Dominant tree 3 23.53±2.10 20.75±1.20 131.00±18.94 中等木Intermediate tree 3 11.17±2.73 12.82±2.03 39.15±15.33 劣势木Suppressed tree 3 6.01±0.82 8.66±0.76 13.79±3.03 1.3 环境因子数据
23m处净辐射(Rn,MJ/(m2·s))、土壤热通量G和降水量(P,mm),1.5和20m两个高度的气温(Ta,℃)、相对湿度(RH,%)、风速(w,m/s)和水汽压亏缺(VPD)数据均获取自通量塔的梯度气象系统。不同深度土层的土壤组成存在差异,其持水性、渗透性也均不同,本研究中θ为5、15、20和40cm的4个土壤深度的加权土壤容积含水量,计算方法详见Sun等[4, 6-7]。此外,该研究定义P>0.1mm的观测日为降水日。
1.4 水汽通量数据
1.4.1 蒸散发
ET获取自通量塔,其计算过程经过一系列的质量控制。本研究中,ET及其组分的关系描述为式(2)。该研究中的显热与潜热通量之和与净辐射与土壤热通量之差的线性回归结果显示:斜率为1.67,截距为18.87,R2为0.80,与FLUXNET研究站点[30]相比(R2范围0.64~0.95),该研究站点的能量闭合水平可以保证较好的数据质量。涡度相关技术容易低估夜间湍流不充分时的通量[9],该研究通过对摩擦风速的敏感性测试剔除了湍流强度较弱时的数据[31-32],保证涡度相关系统的测定是处于强湍流条件下的结果。经质量控制后的潜热通量的缺失率约22%。同时利用MATLAB(R2015b, MathWorks, USA)进行插补[26]。BP网络模型采用输入层、隐含层和输出层的3层式结构。输入层的气象因子以Penman-Monteith模型参数为基础,包括代表能量分配与变化的Rn、Ta和G,代表研究区域上层大气湍流变化的w和RH,以及代表土壤水分状况的θ。
l=√n+m+a (1) 式中:l为隐含层节点数,n为输入节点数(该研究取6),m为输出节点数(该研究取1);a为1~10之间的调节常数。最终,l值取10,LE拟合结果的R2为0.87。
ET=Ef+Ec+Td+Ti+Ts (2) 式中:ET为生态系统蒸散发,Ef为林地表面蒸发,Ec为林冠截留,Td为优势木蒸腾,Ti为中等木蒸腾,Ts为劣势木蒸腾。需要强调的是,该公式指的是ET及其组分间的分配关系,而并非其数值上的计算公式。本研究中ET、Td、Ti和Ts均为实际测定,而Ef和Ec均为模型模拟计算。
1.4.2 林分蒸腾量
边材宽度获取自生长锥取样调查法。结果显示:优势木各方位边材宽度均大于2cm,而中等木、劣势木北向边材宽度分别为(1.3±0.1)cm和(1.1±0.1)cm。因此,为尽量减小树干液流方位变异和太阳辐射增温现象的影响,本研究中优势木在东、西、北向分别安装液流探针,而中等木和劣势木则仅安装在北向;中等木和劣势木的探针安装时需斜向插入边材(即与茎轴夹角 < 90°),保证探针只与边材接触。液流密度(Fd, cm3/(cm2·h))、液流速率(Fs, g/h)与T(指Td、Ti和Ts,mm)的计算公式见式(4)~(6);采用边材面积对单木尺度蒸腾耗水进行扩展(式(3))[10]。
SA=0.71×DBH1.65,R2=0.93,P<0.01 (3) Fd=0.0119×3600×(ΔVmax (4) F_{\mathrm{s}}=F_{\mathrm{d}} \cdot \mathrm{SA} (5) T=F_{\mathrm{d}} /\left(10^{3} A\right) (6) 式中:DBH为树木的胸径,cm;ΔVmax为液流为0时的电压,ΔV是某一测定时刻的电压,A为样地面积(300m2),相关计算采用Baseliner软件。
T_{\mathrm{tot}}=T_{\mathrm{d}}+T_{\mathrm{i}}+T_{\mathrm{s}} (7) 式中:Td、Ti、Ts和Ttot表示优势木、中等木、劣势木和林分总蒸腾量,林分内优势木的边材面积占林分总边材面积的54.1%,中等木占31.6%,被压木为14.3%,因而以边材面积为基础的尺度上推方法所计算的Ttot已将各等级林木蒸腾耗水的权重比例分配考虑在内。
1.4.3 林地表面蒸发
该研究采用改进的Penman-Monteith方程模拟计算Ef[4, 7, 11, 14]。Ef包括林下植被蒸腾和蒸发两部分[7],以往研究认为天然林林下植被蒸腾远小于乔木蒸腾和蒸发[8],因而尽管研究中的Ef代表林地表面蒸发,但在较大尺度的研究中往往可以考虑等同为林地表面蒸发量。由于研究地点通量塔的梯度气象系统未安装地表(1.5m)净辐射相关传感器,该研究以Beer-Bouguer-Lambert Law为基础[27],利用式(9),采用LAI-2200C(Li-Cor, USA)每隔3d测定的叶面积指数(LAI)计算Rf。同时,式(9)结果显示,观测期间的Rf平均为Rn的15.04%。es(Ta)和ea的计算方法参见式(10)和式(11)。同时,ρa、cp、γ和λ分别取1.293、1.03、0.066和2.26[14]。ra由空气动力学参数ca(通常为0.01)和林地1.5m处w通过式(12)计算而得;rc为与θ相关的经验函数见式(13)。k为消光系数,通常取-0.05[14, 28]。此外,以E代表广义的水分蒸发(包括Ef和Ec)。同时,该研究根据梯度气象系统记录的半小时尺度的P,并结合以往[15]在同一研究区域、针对同一森林生态系统、在林冠稳定期(7—8月)所构建的多年林外降水量与落叶松林冠截留量的经验模型(详见盛后财等[15]),估算Ec。
E_{\mathrm{f}}=\frac{\Delta\left(R_{\mathrm{f}}-G\right)+\rho_{\mathrm{a}} c_{\mathrm{p}}\left\{e_{\mathrm{s}}\left(T_{\mathrm{a}}\right)-e_{\mathrm{a}}\right\} / r_{\mathrm{a}}}{\left[\Delta+\gamma \times\left(1+r_{\mathrm{c}} / r_{\mathrm{a}}\right)\right] / \lambda} (8) R_{\mathrm{f}}=R_{\mathrm{n}}\left[\exp \left(-k \cdot \mathrm{LAI}_{\mathrm{c}}\right)\right] (9) e_{\mathrm{s}}\left(T_{\mathrm{a}}\right)=0.6108 \times \exp \left[17.27 \times T_{\mathrm{a}} /\left(T_{\mathrm{a}}+237.3\right)\right] (10) e_{\mathrm{a}}=e_{\mathrm{s}}\left(T_{\mathrm{a}}\right) \times \mathrm{RH} / 100 (11) r_{\mathrm{a}}=1 /\left(c_{\mathrm{a}} w\right) (12) r_{\mathrm{e}}=\max \{0,-1.29 \times 104(\theta-0.199)\} (13) 式中:Δ为饱和水汽压曲线在地表 1.5m处Ta的斜率(kPa/℃),Rf即林地净辐射(MJ/(m2·s))为经林冠层消减作用后达到地表的Rn值,ρa为空气密度(kg/m3),cp为空气比热容(J/(kg·℃)),es(Ta)为地表 1.5m处某Ta时的饱和水汽压(kPa),ea为地表 1.5m处蒸汽温度(kPa),γ为经验常数(kPa/℃),λ为水的汽化潜热(MJ/kg),ra和rc分别为林地空气动力学和地表阻力系数(s/m)。
2. 结果与分析
2.1 环境变量特征
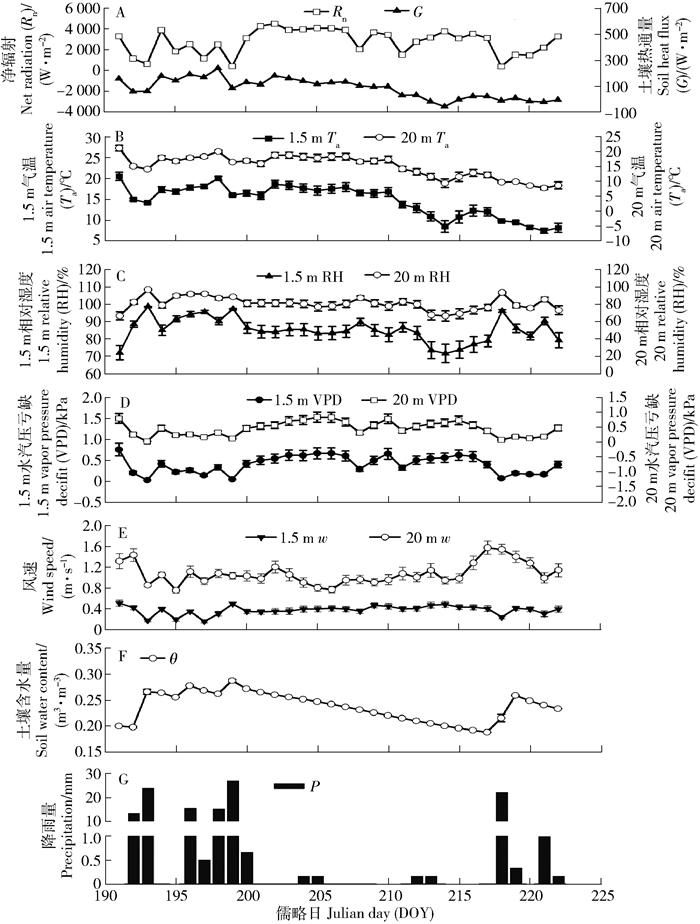
观测期间,共有15d发生降水,累计P为119.46mm(图 1)。降水发生时,生态系统能量分配发生改变,降水量级较高时Rn和G的波动均较为明显(图 1A、1G)。非降水日的Rn(3255.35W/(m2·d))与G(86.76W/(m2·d))均高于降水日(2191.34与77.40W/(m2·d))。RH与θ的变化受降水影响显著(图 1C、1F),尽管非降水日的θ(0.23m3/m3)较降水日(0.25m3/m3)略低(图 1F),但观测期间的土壤水分供给均比较充足(>0.20m3/m3)。VPD与空气水热状况(即Ta与RH)波动十分一致(图 1D),非降水日1.5m处VPD(0.42kPa)和20m处VPD(0.53kPa)均高于降水日(依次分别为0.24和0.34kPa)。同时,由于生长季旺期林木遮蔽度较高,林地表面w明显较林冠上方低(图 1E)。
![]() 图 1 观测期间能量通量和气象因子日变化格局Figure 1. Daily variations of energy flux and meteorological factors during observation periods
图 1 观测期间能量通量和气象因子日变化格局Figure 1. Daily variations of energy flux and meteorological factors during observation periods观测期间,除RH外,林地表面的气象因子均表现为低于林冠上层且变化相对平缓的森林小气候特点,林冠层对于太阳辐射的反射和吸收降低了到达林地表面的Rn强度,而林木蒸腾耗水过程的吸热,也使林内温度变幅减小;同时由于林木表面的摩擦和阻挡等作用,导致林内w迅速降低,不利于水汽的扩散。气象因子在观测期间的趋势变化发现,降水日林冠层的Rn降低,但在降水停止后(即非降水日)反弹迅速,而整个观测期间的G与林地表面和林冠层的Ta均呈下降趋势,体现了生态系统的能量分配对气象因子的驱动作用[29]。随着观测时间的增加,G与显热通量在Rn的分配比例逐渐降低,即LE(潜热通量,即生态系统内用于水汽蒸发的能量总量)逐渐占优,兴安落叶松生态系统趋向于分配更多的能量用于蒸发,抵消降水产生的短期θ升高[10]。此外,由于研究地位于大兴安岭北部,西伯利亚南缘寒温带气候,冬季寒冷漫长、夏季凉爽短暂,年无霜期极短。而根据以往对于同一区域内兴安落叶松蒸腾特征的研究结果[17],发现7—8月是其典型生长旺盛期,相较于展叶期和落叶期,该时期林分的生理生态特征较为稳定。而对该时段的研究结果有利于更清晰的发掘生态系统水分输入变化(降水和非降水)对生态系统蒸散发分配的影响。
2.2 蒸散发及其组分的昼夜变化特征
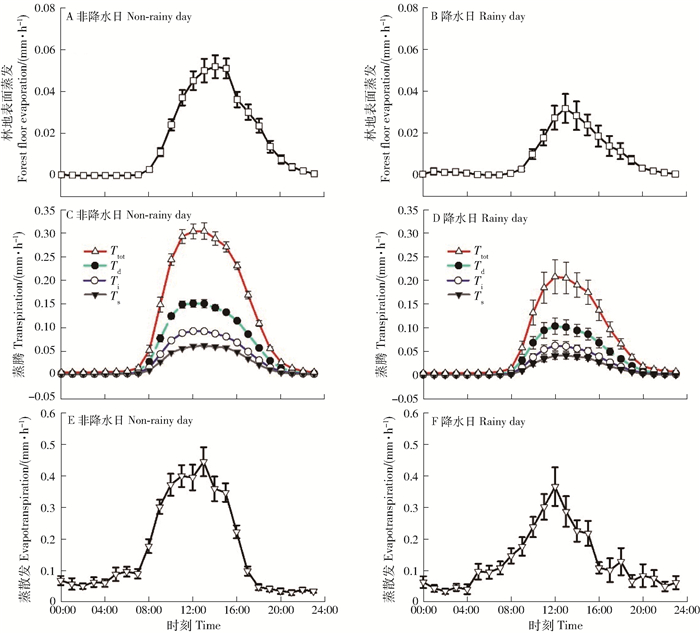
方差分析表明,非降水和降水日的Ef(F1766=29.9)、Td(F1766=24.0)、Ti(F1766=25.3)、Ts(F1766=22.9)、Ttot(F1766=24.3)和ET(F1766=6.6)均差异显著(P < 0.05)。非降水和降水日ET及其组分的昼夜变化均呈单峰格局(图 2)。非降水日Ef峰值出现在14:00,降水日则出现在13:00,分别为(0.05±0.01)mm/h和(0.03±0.01)mm/h(图 2A、2B),非降水日的Td、Ti、Ts和Ttot的峰值均出现在13:00,分别为(0.15±0.01)mm/h、(0.09±0.01)mm/h、(0.06±0.00)mm/h和(0.34±0.02)mm/h(图 2C);降水日,蒸腾峰值出现12:00,Td、Ti、Ts和Ttot的峰值分别降至(0.10±0.02)mm/h、(0.06±0.01)mm/h、(0.04±0.01)mm/h和(0.21±0.04)mm/h(图 2D),可见林分在非降水日的蒸腾需求更高(总量大、峰值较高、峰值出现时间较早)。非降水日ET的峰值为(0.44±0.05)mm/h,同样高于降水日的(0.37± 0.06)mm/h(图 2E、2F)。此外,无论降水与否,优势木的蒸腾量均高于中等木和劣势木。
![]() 图 2 观测期间林地表面蒸发和林木蒸腾以及生态系统蒸散发的昼夜变化格局Figure 2. Diurnal variations of forest floor evaporation (Ef), forest transpiration (T) and ecosystem evapotranspiration (ET) during observation periods
图 2 观测期间林地表面蒸发和林木蒸腾以及生态系统蒸散发的昼夜变化格局Figure 2. Diurnal variations of forest floor evaporation (Ef), forest transpiration (T) and ecosystem evapotranspiration (ET) during observation periods值得一提的是,本研究中ET、Td、Ti和Ts均为实际测定,而Ef和Ec均为模型模拟结果。因此,不同组分之和与实际测定的ET间存在客观误差(式2两端不闭合),考虑到闭合调整的方法误差有可能高于不同测算方法本身的系统误差,因此该研究中均采用了原始结果进行计算和比较。兴安落叶松全部观测日时的兴安落叶松林蒸腾能力为1.92mm/d,高于张彦群等[33]总结的北方针叶林生长季1.24mm/d的蒸腾能力,表明以兴安落叶松为优势种群的北方森林的蒸腾耗水能力较强。同时,由于以Rn和VPD为代表的气象因子是林木T的主要驱动力[34],且兴安落叶松的T对气象因子响应敏感[22],因而不同区域的气象因子的差别、树种组成与林龄的差异[35]以及由竞争引起的林木等级的不确定性[21, 36]均不利于对全球尺度研究结果的交叉对比。ET的日变化格局与T更为接近,主要原因与T是生态系统主要的水汽通量来源有关[3],同时该研究结果也证明对兴安落叶松林的全部观测日Ttot均明显高于Ef,与以往研究结果相近[1, 4, 7, 14]。此外,全部观测日的林木Td均高于Ti和Ts,与林内竞争优势地位有关[22, 36],优势木冠层位于林分平均冠层之上,可获取较多太阳辐射能量以充分驱动蒸腾耗水[37]。非降水和降水日Ef的峰值均滞后树干液流约1h,出现该时滞的原因可能与冠层对Rn的消减作用有关,从太阳辐射在林内的垂直分布角度,林冠层不仅在时间上先于林地表面,同时其接触到的辐射量级也较高,从而优先驱动蒸腾。
2.3 蒸散发及其组分的日变化特征
图 3为观测期内日水汽通量的连续变化特征。结果表明:非降水日,Ef、Td、Ti、Ts、Ttot和ET总量分别为10.3、25.6、15.2、10.8、51.6和66.3mm,降水日则分别降低至2.2、11.2、5.1、3.8、20.1和47.8mm;此外,在降水日,Ec最高,为24.3mm。非降水日Ef、Td、Ti、Ts、Ttot和ET日均值分别为0.6、1.5、0.9、0.6、3.0、3.9mm/d,降水日则降低至0.2、0.8、0.3、0.3、1.3和3.2mm/d,而Ec的日交换能力为1.6mm/d。可见降水对森林生态系统—大气界面的水汽通量交换具有明显的削弱效应;非降水和降水日,Ttot均由Td显著占优;同时,优势木、中等木和劣势木的非降水日水汽交换能力为降水日的1.9、3.0和2.0倍,表明中等木蒸腾对气象变化最为敏感,其次为劣势木和优势木。此外,ET的测定基于涡度通量观测技术,实时的对生态系统水汽通量进行观测,因此通量塔的获取的ET与其组分经模型模拟(如Ef、Ec)和实际观测如(Td、Ti和Ts)的累加值存在不能完全闭合的现象[10]。降水发生时,由于RH的逐渐饱和,水分的蒸发过程较为困难。以2015年第197天(DOY 197)的观测为例,该日发生的降水为观测期间最高值,而ET则明显降低。这首先体现了林分对水分输入的响应过程,DOY197之后,由于天气晴朗,空气湿度降低,出现蒸散发需求,ET迅速升高。Ec和Ef均为物理过程,从日变化特征的相关数值较为容易理解,而ET和T(Td、Ti、Ts)则包括了植被的生物学过程,其对环境的响应存在滞后效应(如树干水分动态调整),日变化动态更多的体现林分对水分输入的动态响应过程。
![]() 图 3 观测期间林地表面蒸发、林冠截留、林木蒸腾和蒸散发的日变化特征Figure 3. Daily variations of forest floor evaporation (Ef), canopy interception (Ec), forest transpiration (T) and evapotranspiration (ET) during observation periods
图 3 观测期间林地表面蒸发、林冠截留、林木蒸腾和蒸散发的日变化特征Figure 3. Daily variations of forest floor evaporation (Ef), canopy interception (Ec), forest transpiration (T) and evapotranspiration (ET) during observation periods2.4 蒸散发及其组分的分配特征
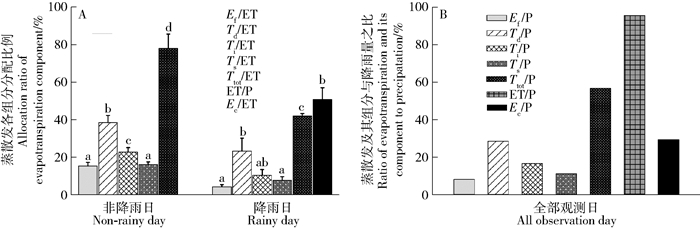
LSD多重比较分析表明,非降水和降水日的组内各ET组分与ET之比均差异显著(P < 0.05)(图 4A)。非降水日,T是水汽通量返回大气的主要形式,Ttot/ET可以达到78.0%,其中Td/ET、Ti/ET和Ts/ET分别为38.7%、23.0%和16.4%;降水日,E为ET的主要组分,Ec/ET为50.9%,Td/ET、Ti/ET、Ts/ET和Ttot/ET分别为23.5%、10.6%、8.0%和42.2%。同时,非降水日Ef/ET高于降水日(15.5%> 4.6%),但其对ET的贡献程度较低。图 4B为ET及其组分与P的比值。全部观测日的ET/P为94.7%,约5.3%的P产生径流或下渗;ET组分与P之比由小到大依次为Ef/P、Ts/P、Ti/P、Ec/P、Td/P,数值分别为8.3%、11.3%、16.8%、29.6%和28.7%;绝大部分的P通过T的形式返回大气(Ttot/P为56.8%),E(Ef和Ec)则占37.9%。以上结果说明:非降水日ET主要以T为主(具体为Td),降水日则以E(具体为Ec)占优,蒸腾耗水是该系统水分输出最主要形式。
![]() 图 4 不同降水状况下蒸散发及其组分的分配比例不同小写字母表示非降水或降水日组内各水汽通量组分与ET比值间差异的显著性(P < 0.05)。Figure 4. Evapotranspiration and its components under different precipitation conditionsDifferent lowercase letters mean the significance at P < 0.05 level in difference of water vapor flux components to ET within group in non-rainy days or rainy days.
图 4 不同降水状况下蒸散发及其组分的分配比例不同小写字母表示非降水或降水日组内各水汽通量组分与ET比值间差异的显著性(P < 0.05)。Figure 4. Evapotranspiration and its components under different precipitation conditionsDifferent lowercase letters mean the significance at P < 0.05 level in difference of water vapor flux components to ET within group in non-rainy days or rainy days.对于兴安落叶松林生态系统,非降水日的ET明显更高,表明该时期的气象条件更有利于植物—大气界面的水汽通量交换。同时,结果显示降水日ET主要以Ec占优势,而非降水日则以Td为主,与以往的研究结果相符[4, 7, 10, 38];说明生态系统会随着水分输入条件的改变对ET组分(即E和T的分配比例)进行动态调节[1, 10],主要原因是叶片气孔对空气湿度增加的响应较为强烈从而限制了植物的蒸腾耗水[34, 39]。全部观测日的Ttot约占ET的60.1%,略低于Schlesinger等[1]的全球统计结果(平均为65%),主要原因可能是观测期间的降水事件较多,不利于维持较高的T[22],但对E则影响相对较弱。同时,结果发现,观测期间约95%的P主要以ET的形式返回大气,其中T约为57%,而E约为38%,进一步验证了兴安落叶松林生态系统的ET分配模式,即非降水日的ET以T为主(具体为Td),降水日则以E(具体为Ec)占优;其余5%的P的去向则包括地表径流和下渗。
此外,非降水和降水日E(Ef和Ec)与Ttot(Td、Ti和Ts)之和略低于ET(误差在7%以内),这与所采用模型的模拟精度[40]、水分输入状况对通量观测传感器的干扰[11]以及通量数据插补的方法有关[26]。此外,非降水日,Td/ET分别可以达到Ti/ET和Ts/ET的1.7和3.0倍,而在降水日则分别为2.2和2.9倍,因此仅以Td(包括较大径级的Ti)代表林木平均蒸腾能力进行尺度上推的方法容易高估林分的蒸腾能,具体误差的大小则取决于林分内中等木和劣势木的株树以及林木边材面积大小,而如果观测期间出现降水日,该误差可能还会进一步增大。此外,该研究建议在研究E、T以及ET对主要气象因子的响应关系时,需考虑环境水分条件的差异[10-11, 38]。
2.5 蒸散发及其组分对主要环境因子的响应
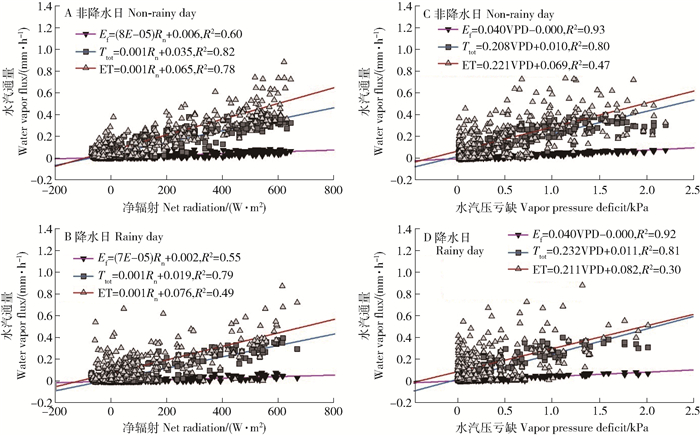
以降水的发生情况为固定变量,Rn和VPD(1.5、20m处VPD的平均值)为协变量进行方差分析,结果表明非降水和降水日的Ef、Ttot和ET均受Rn、VPD的显著影响(P < 0.01)。选取Ef、Ttot以及ET作为对气象因子响应的水汽变量,非降水和降水日Ef、Ttot和ET与Rn均呈正相关关系(图 5A、5B),且非降水日时的R2均较高,表明水分的输入影响水汽通量对Rn的响应。同时,非降水日和降水日的Ef和Ttot对VPD响应程度均较高(R2分别为0.9和0.8;图 5C、5D),表明非降水和降水日的VPD均为Ef和Ttot的限制性因子;非降水日Ttot随VPD的增加而逐渐增强的范围较降水日时明显,表明水分输入较低时利于维持林木的蒸腾需求;ET对VPD的响应水平在非降水日略高(图 5C、5D)。总体上,非降水和降水日Ttot对Rn的响应水平均高于Ef和ET,但Ef对Rn响应在非降水日略高于ET,而在降水日反之;Ef和Ttot对VPD的响应水平高于ET,该现象在降水日更为明显。此外,在非降水或降水日,ET与Rn的相关性均高于其与VPD的相关性,Ttot则差异不大,而Ef则表现出与ET相反的特点。上述结果表明Rn是生态系统能量循环和物质交换的最主要驱动力[29],Rn和VPD均可以对Ttot进行有效的限制[22, 37],而Ef则优先受VPD的限制[41]。
![]() 图 5 非降水和降水日的蒸散发及其组分与净辐射和水汽压亏缺的相关性线性相关性均显著(P < 0.01);VPD为1.5与20m两个高度处的平均值。Figure 5. Correlations of evapotranspiration and its components with net radiation (Rn) and vapor pressure deficit (VPD) in both non-rainy and rainy daysLinear correlation is all significant(P < 0.01); VPD is the average of 1.5 and 20m height.
图 5 非降水和降水日的蒸散发及其组分与净辐射和水汽压亏缺的相关性线性相关性均显著(P < 0.01);VPD为1.5与20m两个高度处的平均值。Figure 5. Correlations of evapotranspiration and its components with net radiation (Rn) and vapor pressure deficit (VPD) in both non-rainy and rainy daysLinear correlation is all significant(P < 0.01); VPD is the average of 1.5 and 20m height.3. 结论
非降水和降水日的林地表面蒸散发、优势木蒸腾、中等木蒸腾、被压木蒸腾和蒸散发的日变化呈单峰格局,且非降水日曲线峰值高于降水日。蒸散发是观测期间降雨返回大气的最主要形式,其中由T贡献较多。非降水日蒸散发以蒸腾为主(具体为优势木蒸腾),降水日则以蒸腾(具体为林冠截留)为主。兴安落叶松优势木的蒸腾能力强于被压木,以往研究常用的以优势木和中等木为代表木进行尺度上推的方法会高估林分的蒸腾能力。而无论降水与否,蒸散发与净辐射的相关性均高于其与水汽压亏缺的相关性,林分总蒸腾与二者的相关性则差异不大,而林地表面蒸散发的表现则与生态系统蒸散发相反。
-
![]()
图 1 观测期间能量通量和气象因子日变化格局
Figure 1. Daily variations of energy flux and meteorological factors during observation periods
![]()
图 2 观测期间林地表面蒸发和林木蒸腾以及生态系统蒸散发的昼夜变化格局
Figure 2. Diurnal variations of forest floor evaporation (Ef), forest transpiration (T) and ecosystem evapotranspiration (ET) during observation periods
![]()
图 3 观测期间林地表面蒸发、林冠截留、林木蒸腾和蒸散发的日变化特征
Figure 3. Daily variations of forest floor evaporation (Ef), canopy interception (Ec), forest transpiration (T) and evapotranspiration (ET) during observation periods
![]()
图 4 不同降水状况下蒸散发及其组分的分配比例
不同小写字母表示非降水或降水日组内各水汽通量组分与ET比值间差异的显著性(P < 0.05)。
Figure 4. Evapotranspiration and its components under different precipitation conditions
Different lowercase letters mean the significance at P < 0.05 level in difference of water vapor flux components to ET within group in non-rainy days or rainy days.
![]()
图 5 非降水和降水日的蒸散发及其组分与净辐射和水汽压亏缺的相关性
线性相关性均显著(P < 0.01);VPD为1.5与20m两个高度处的平均值。
Figure 5. Correlations of evapotranspiration and its components with net radiation (Rn) and vapor pressure deficit (VPD) in both non-rainy and rainy days
Linear correlation is all significant(P < 0.01); VPD is the average of 1.5 and 20m height.
表 1 液流测定样木特征
Table 1 Sample tree features of sap flow measurement
林木等级
Forest grade样本数
Sample tree number平均胸径
Average DBH/cm平均树高
Average height/m平均边材面积
Average sapwood area/cm2优势木Dominant tree 3 23.53±2.10 20.75±1.20 131.00±18.94 中等木Intermediate tree 3 11.17±2.73 12.82±2.03 39.15±15.33 劣势木Suppressed tree 3 6.01±0.82 8.66±0.76 13.79±3.03  下载: 导出CSV
下载: 导出CSV
-
[1] Schlesinger W H, Jasechko S. Transpiration in the global water cycle[J]. Agricultural and Forest Meteorology, 2014, 189(6): 115-117. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=43a92230a6d9b8a4f9c94139e9bdffeb
[2] Xiao J F, Sun G, Chen J Q, et al. Carbon fluxes, evapotranspiration, and water use efficiency of terrestrial ecosystems in China[J].Agriculturaland Forest Meteorology, 2013, s182-183(22): 76-90. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=0bf5bf98fd07ab7f2930ba9f69004fcc
[3] Jasechko S, Sharp Z D, Gibson J J, et al. Terrestrial water fluxes dominated by transpiration[J]. Nature, 2013, 496(7445): 347-350. doi: 10.1038/nature11983
[4] Sun X C, Onda Y, Kato H, et al. Partitioning of the total evapotranspiration in a Japanese cypress plantation during the growing season[J].Ecohydrology, 2014, 7(3): 1042-1053. doi: 10.1002/eco.v7.3
[5] Zhang B Z, Xu D, Liu Y, et al. Multi-scale evapotranspiration of summer maize and the controlling meteorological factors in north China[J]. Agricultural and Forest Meteorology, 2016, 216: 1-12. doi: 10.1016/j.agrformet.2015.09.015
[6] Sun X C, Onda Y, Otsuki K, et al. The effect of strip thinning on tree transpiration in a Japanese cypress (Chamaecyparis obtusa Endl.) plantation[J]. Agricultural and Forest Meteorology, 2014, 197: 123-135. doi: 10.1016/j.agrformet.2014.06.011
[7] Sun X C, Onda Y, Otsuki K, et al. The effect of strip thinning on forest floor evaporation in a Japanese cypress plantation[J]. Agricultural and Forest Meteorology, 2016, 216: 48-57. doi: 10.1016/j.agrformet.2015.10.006
[8] Yepez E A, Williams D G, Scott r L, et al. Partitioning overstory and understory evapotranspiration in a semiarid savanna woodland from the isotopic composition of water vapor[J]. Agricultural and Forest Meteorology, 2003, 119(1-2): 53-68. doi: 10.1016/S0168-1923(03)00116-3
[9] Fisher J B, Baldocchi D D, Misson L, et al. What the towers don't see at night: nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California[J]. Tree Physiology, 2007, 27(4): 597-610. doi: 10.1093/treephys/27.4.597
[10] 刘晨峰, 张志强, 孙阁, 等.基于涡度相关法和树干液流法评价杨树人工林生态系统蒸发散及其环境响应[J].植物生态学报, 2009, 33(4):706-718. doi: 10.3773/j.issn.1005-264x.2009.04.009 Liu C F, Zhang Z Q, Sun G, et al. Quantifying evapotranspiration and biophysical regulations of a poplar plantation assessed by eddy covariance and sap-flow methods[J]. Chinese Journal of Plant Ecology, 2009, 33(4): 706-718. doi: 10.3773/j.issn.1005-264x.2009.04.009
[11] Tian F X, Zhao C Y, Feng Z D. Simulating evapotranspiration of Qinghai spruce (Picea crassifolia) forest in the Qilian Mountains, northwestern China[J]. Journal of Arid Environments, 2011, 75(7): 648-655. doi: 10.1016/j.jaridenv.2011.02.001
[12] Waterloo M J, Bruijnzeel L A, Vugts H F, et al. Evaporation from Pinus caribaea plantations on former grassland soils under maritime tropical conditions[J]. Water Resources Research, 1999, 35(7): 2133-2144. doi: 10.1029/1999WR900006
[13] Lin Y, Wang G X, Guo J Y, et al. Quantifying evapotranspiration and its components in a coniferous subalpine forest in Southwest China[J]. Hydrological Processes, 2012, 26(20): 3032-3040. doi: 10.1002/hyp.v26.20
[14] Bond-lamberty B, Gower S T, Amiro B, et al. Measurement and modelling of bryophyte evaporation in a boreal forest chronosequence[J]. Ecohydrology, 2011, 4(1): 26-35. doi: 10.1002/eco.v4.1
[15] 盛后财, 蔡体久, 李奕, 等.大兴安岭北部兴安落叶松林降雨截留再分配特征[J].水土保持学报, 2014, 28(6):101-105. http://d.old.wanfangdata.com.cn/Periodical/trqsystbcxb201406019 Sheng H C, Cai T J, Li Y, et al. Rainfall redistribution in Larix gmelinii forest on Northern of Daxing'an Mountains, northeast of China[J].Journal of Soil and Water Conservation, 2014, 28(6):101-105. http://d.old.wanfangdata.com.cn/Periodical/trqsystbcxb201406019
[16] Wilson K B, Hanson P J, Mulholland P J, et al. A comparison of methods for determining forest evapotranspiration and its components: sap-flow, soil water budget, eddy covariance and catchment water balance[J]. Agricultural and Forest Meteorology, 2001, 106(2): 153-168. doi: 10.1016/S0168-1923(00)00199-4
[17] 池波, 蔡体久, 满秀玲, 等.大兴安岭北部兴安落叶松树干液流规律及影响因子分析[J].北京林业大学学报, 2013, 35(4):21-26. http://j.bjfu.edu.cn/article/id/9894 Chi B, Cai T J, Man X L, et al. Effects of influencing factors on stem sap flow in Larix gmelinii in northern Da Hinggan Mountains, northeastern China[J]. Journal of Beijing Forestry University, 2013, 35(4):21-26. http://j.bjfu.edu.cn/article/id/9894
[18] 孙鹏森, 马履一, 王小平, 等.油松树干液流的时空变异性研究[J].北京林业大学学报, 2000, 22(5):1-6. doi: 10.3321/j.issn:1000-1522.2000.05.001 Sun P S, Ma L Y, Wang X P, et al. Temporal and spacial variation of sap flow of Chinese pine (Pinus tabuliformix) [J]. Journal of Beijing Forestry University, 2000, 22(5):1-6. doi: 10.3321/j.issn:1000-1522.2000.05.001
[19] 张璇, 张会兰, 王玉杰, 等.缙云山典型树种树干液流日际变化特征及与气象因子关系[J].北京林业大学学报, 2016, 38(3):11-20. doi: 10.13332/j.1000-1522.20150389 Zhang X, Zhang H L, Wang Y J, et al. Characteristics of daily sap flow for typical species in Jinyun Mountain of Chongqing in relation to meteorological factors[J]. Journal of Beijing Forestry University, 2016, 38(3):11-20. doi: 10.13332/j.1000-1522.20150389
[20] 陈宝强, 张建军, 张艳婷, 等.晋西黄土区辽东栎和山杨树干液流对环境因子的响应[J].应用生态学报, 2016, 27(3): 746-754. http://d.old.wanfangdata.com.cn/Periodical/yystxb201603010 Chen B Q, Zhang J J, Zhang Y T, et al. Whole-tree sap flow of Quercus liaotungensis and Populus davidiana in response to environmental factors in the Loess Plateau area of western Shanxi Province, northern China [J]. Chinese Journal of Applied Ecology, 2016, 27(3): 746-754. http://d.old.wanfangdata.com.cn/Periodical/yystxb201603010
[21] 李振华, 王彦辉, 于澎涛, 等.华北落叶松液流速率的优势度差异及其对林分蒸腾估计的影响[J].林业科学研究, 2015, 28(1): 8-16. http://d.old.wanfangdata.com.cn/Periodical/lykxyj201501002 Li Z H, Wang Y H, Yu P T, et al. Variation of sap flow density of Larix principis-rupprechtii with dominances and its impact on stand transpiration estimation [J]. Forest Research, 2015, 28(1): 8-16. http://d.old.wanfangdata.com.cn/Periodical/lykxyj201501002
[22] 刘家霖, 满秀玲, 胡悦.兴安落叶松天然林不同分化等级林木树干液流对综合环境因子的响应[J].林业科学研究, 2016, 29(5): 726-734. doi: 10.3969/j.issn.1001-1498.2016.05.015 Liu J L, Man X L, Hu Y. Response of tree sap flow of Larix gmelinii with various differentiation classes to multiple environmental factors [J]. Forest Research, 2016, 29(5): 726-734. doi: 10.3969/j.issn.1001-1498.2016.05.015
[23] Soja A J, Tchebakova N M, French N H F, et al. Climate-induced boreal forest change: predictions versus current observations[J]. Global and Planetary Change, 2007, 56(3/4): 274-296. http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ027078691/
[24] 玉宝, 乌吉斯古楞, 王百田, 等.大兴安岭兴安落叶松(Larix gmelinii)天然林分级木转换特征[J].生态学报, 2008, 28(11): 5750-5757. doi: 10.3321/j.issn:1000-0933.2008.11.062 Yu B, Wujisiguleng, Wang B T, et al.Analysis on classified stem transformation characteristic of Larix gmelinii natural forest at Daxingan Mountains [J]. Acta Ecologica Sinica, 2008, 28(11): 5750-5757. doi: 10.3321/j.issn:1000-0933.2008.11.062
[25] 玉宝, 乌吉斯古楞, 王百田, 等.兴安落叶松天然林树冠生长特性分析[J].林业科学, 2010, 46(5): 41-48. http://d.old.wanfangdata.com.cn/Periodical/lykx201005007 Yu B, Wujisiguleng, Wang B T, et al.Analysis of crown growth characteristics in natural Larix gmelinii forests [J]. Scientia Silvae Sinicae, 2010, 46(5): 41-48. http://d.old.wanfangdata.com.cn/Periodical/lykx201005007
[26] Van Wijk M T, Bouten W. Water and carbon fluxes above European coniferous forests modelled with artificial neural networks[J]. Ecological Modelling, 1999, 120(2/3): 181-197. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=2032594c3aaa44fc6c6e7e03bd617ec4
[27] Sinclair T R. A reminder of the limitations in using beer's law to estimate daily radiation interception by vegetation[J]. Crop Science, 2006, 46(6): 2343-2347. doi: 10.2135/cropsci2006.01.0044
[28] Landsberg J J, Gower S T. Applications of physiological ecology to forest management[M]. San Diego:Academic Press, 1997:354.
[29] 谭正洪, 于贵瑞, 周国逸, 等.亚洲东部森林的小气候特征(1):辐射和能量的平衡[J].植物生态学报, 2015, 39(6): 541-553. http://d.old.wanfangdata.com.cn/Periodical/zwstxb201506001 Tan Z H, Yu G R, Zhou G Y, et al. Microclimate of forests across East Asia biomes (1): radiation and energy balance[J]. Chinese Journal of Plant Ecology, 2015, 39(6): 541-553. http://d.old.wanfangdata.com.cn/Periodical/zwstxb201506001
[30] Wilson K, Goldstein A, Falge E, et al. Energy balance closure at FLUXNET sites[J]. Agricultural and Forest Meteorology, 2002, 113(1/4): 223-243. http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ0229230263/
[31] Goulden M L, Munger J W, Fan S M, et al. Measurements of carbon sequestration by long-term eddy covariance: methods and a critical evaluation of accuracy[J]. Global Change Biology, 1996, 2(3): 169-182. doi: 10.1111-j.1365-2486.1996.tb00070.x/
[32] 徐自为, 刘绍民, 宫丽娟, 等.涡动相关仪观测数据的处理与质量评价研究[J].地球科学进展, 2008, 23(4): 357-370. doi: 10.3321/j.issn:1001-8166.2008.04.005 Xu Z W, Liu S M, Gong L J, et al. A study on the data processing and quality assessment of the eddy covariance system[J]. Advances in Earth Science, 2008, 23(4): 357-370. doi: 10.3321/j.issn:1001-8166.2008.04.005
[33] 张彦群, 王传宽.北方和温带森林生态系统的蒸腾耗水[J].应用与环境生物学报, 2008, 14(6): 838-845. http://d.old.wanfangdata.com.cn/Periodical/yyyhjswxb200806020 Zhang Y Q, Wang C K. Transpiration of boreal and temperate forests[J]. Chinese Journal of Applied & Environmental Biology, 2008, 14(6): 838-845. http://d.old.wanfangdata.com.cn/Periodical/yyyhjswxb200806020
[34] Meinzer F C, Goldstein G, Jackson P, et al. Environmental and physiological regulation of transpiration in tropical forest gap species: the influence of boundary layer and hydraulic properties[J]. Oecologia, 1995, 101(4): 514-522. doi: 10.1007/BF00329432
[35] Lagergren F, Lindroth A. Variation in sapflow and stem growth in relation to tree size, competition and thinning in a mixed forest of pine and spruce in Sweden[J]. Forest Ecology and Management, 2004, 188(1-3): 51-63. doi: 10.1016/j.foreco.2003.07.018
[36] 刘家霖, 王传宽, 张全智.不同分化等级兴安落叶松树干心材和边材空间变异[J].林业科学, 2014, 50(12): 114-121. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=lykx201412016 Liu J L, Wang C K, Zhang Q Z. Spatial variations in stem heartwood and sapwood for Larix gmelinii trees with various differentiation classes [J]. Scientia Silvae Sinicae, 2014, 50(12): 114-121. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=lykx201412016
[37] O'brien J J, Oberbauer S F, Clark D B. Whole tree xylem sap flow responses to multiple environmental variables in a wet tropical forest[J].Plant, Cell & Environment, 2004, 27(5): 551-567. doi: 10.1111-j.1365-3040.2003.01160.x/
[38] 白岩, 朱高峰, 张琨, 等.基于树干液流及涡动相关技术的葡萄冠层蒸腾及蒸散发特征研究[J].生态学报, 2015, 35(23): 7821-7831. http://d.old.wanfangdata.com.cn/Periodical/stxb201523026 Bai Y, Zhu G F, Zhang K, et al. Research of transpiration and evapotranspiration from a grapevine canopy combining the sap flow and eddy covariance techniques [J]. Acta Ecologica Sinica, 2015, 35(23): 7821-7831. http://d.old.wanfangdata.com.cn/Periodical/stxb201523026
[39] Whitehead D. Regulation of stomatal conductance and transpiration in forest canopies[J]. Tree Physiology, 1998, 18(8-9): 633-644. doi: 10.1093/treephys/18.8-9.633
[40] Massonnet F, Bellprat O, Guemas V, et al. Using climate models to estimate the quality of global observational data sets[J].Science, 2016, 354: 425-455. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=9b0c5ae97d08fd17adca62219db65f30
[41] Deguchi A, Hattori S, Daikoku K, et al. Measurement of evaporation from the forest floor in a deciduous forest throughout the year using microlysimeter and closed-chamber systems[J]. Hydrological Processes, 2008, 22(18): 3712-3723. doi: 10.1002/hyp.v22:18
-
期刊类型引用(1)
1. 伊丽茜,岳永杰,张军,舒洋,王雅倩. 额尔古纳市兴安落叶松中龄林植被碳储量研究. 现代园艺. 2021(23): 55-56+74 .  百度学术
百度学术
其他类型引用(6)

计量
- 文章访问数: 2134
- HTML全文浏览量: 307
- PDF下载量: 62
- 被引次数: 7
