
Spatiotemporal distribution and seed germination characteristics of Betula costata seed rain in the spruce-fir mixed forest
-
摘要:目的 研究东北云冷杉针阔混交林枫桦种子雨时空分布规律和种子萌发状况,为云冷杉针阔混交林物种多样性保护提供理论依据。方法 基于在吉林省汪清林业局设立的9块1 hm2固定样地,按对角线五点取样法在每块样地布设5个(9块样地共布设45个)种子收集器收集枫桦种子,采用方差/均值(Cx
)、负二项指数(K)、聚集度指标(David和Moore丛生指标,I)、Cassie. R. M.指标(CA)等4种空间分布类型评价方法判别种子雨空间分布状态,此外,从9、10月份收集的种子中,每块样地随机抽取300粒枫桦种子进行种子萌发试验,分别测定种子千粒质量、发芽势和发芽率。 结果 从7—11月份,共收集到69 100粒种子;枫桦落种高峰期为10月份;种子分布方差/均值(Cx)达1116.30、负二项指数(K)达4.75、聚集度指标(David和Moore丛生指标,I)达1115.30、Cassie. R. M.指标(CA)达0.41,4个指标均显示种子空间分布呈聚集分布;9月份与10月份收集的平均种子千粒质量分别为(0.398 ± 0.064)g和(0.353 ± 0.040)g,9月份种子千粒质量显著高于10月份(P < 0.01);9月份收集的种子平均发芽率为(29.84 ± 11.68)%,显著高于10月份(23.52 ± 8.23)%(P < 0.05)。 结论 在云冷杉针阔混交林中,枫桦种子小结实量大,且具有明显的时间分布特征,由于在一定程度上种子扩散受到限制,使得枫桦种子呈聚集分布状态。该区域枫桦种源丰富但种子萌发能力差,这些特性将会影响枫桦分布格局,从而影响该地区云冷杉针阔混交林群落结构和物种多样性。Abstract:Objective The population of Betula costata in spruce-fir mixed forest in northeastern China was taken as the research object. The spatiotemporal distribution of Betula costata seed rain and seed germination characteristics were studied to provide theoretical basis for species diversity protection of the forest.Method Based on the 9 sample plots, 1 ha (100 m × 100 m) each set in Wangqing Forestry Bureau, Jilin Province of northeastern China, sampling by diagonal five points, we set 5 seed traps in each sample plot, totally 45 in 9 sample plots to collect Betula costata seeds. We used 4 evaluation methods on spatial distribution types to distinguish the spatial distribution state of seed rain, including variance-mean ratio (Cx), negative binomial parameter (K), aggregation index (I), Cassie. R.M. index (CA). In addition, 300 Betula costata seeds collected in September and October were randomly selected from each sample plot for seed germination experiment, and the 1 000-grain mass, germination potential and germination percentage were measured, respectively.Result From July to November, 69 100 seeds were collected in total. The peak period of seed falling for Betula costata was October. We found that the seeds of Betula costata presented aggregation distribution at the spatial level based on 4 indexes, i.e. Cx reaching 1 116.30, K reaching 4.75, I reaching 1 115.30, CA reaching 0.41, respectively. The average 1000-grain mass value of Betula costata seeds in September was 0.398 g, which was significantly higher than that in October (0.353 g) (P<0.01). Similarly, the average germination rate in September was (29.84 ± 11.68)%, which was significantly higher than that in October (23.52 ± 8.23)% (P < 0.05).Conclusion In spruce-fir coniferous-broadleaved mixed forest, the seeds are small and fruiting for Betula costata and have obvious time distribution characteristics. Because the diffusion of Betula costata seeds is limited, as a result the seeds show an aggregated distribution pattern. In this region, the characteristics of rich provenance and poor germination ability of Betula costata will affect the distribution pattern of Betula costata, thus affect the community structure and species diversity of spruce-fir coniferous-broadleaved mixed forest in this area. -
种子雨指植物的种子或果实从母树向地表扩散的过程[1],它是森林土壤种子库的主要来源[2]。种子雨的时空特征反映了树种对环境的适应以及树种自身的天然更新策略。树种能否更新取决于该树种从种子到成树阶段所经历的由环境带来的死亡风险[3],其中种子扩散是植物群落建立幼苗库的第一步[4]。种子的扩散方式会改变森林群落结构,影响物种多样性[5-7]。由于物种本身的遗传特性和周围环境的变化使得种子雨的发生时间、强度、空间分布特征等方面存在较大差异。一般来说,林木的高度、种子的大小和数量、种子形态及落种时期的环境条件都会影响种子的扩散方式[8]。同时种子产量的变化也会影响当地以植物果实或种子为食的动物和昆虫种群的变化[9-10],捕食者饱和假说(seed predator satiation hypothesis)认为在种子歉收季节,捕食者种群数量会随着种子数量减少而减少;在种子丰收年,植物结实量大,使得捕食者的食物饱和,从而增加了更多的种子存活几率[11-13]。种子作为森林更新的起点,其产量和萌发能力对森林更新起着决定性的作用[14],从母树落种到幼苗库建立期间,各种因素都会影响种子的存活、萌发及幼苗的空间格局[15]。因此,对种子雨时空分布特征及其种子萌发特性的研究对一些重要树种的保护和人工辅助更新具有重要意义[16-19]。
东北云冷杉针阔混交林在20世纪30年代,由于日本侵略者在该地区的大规模砍伐优质红松,使得原始阔叶红松(Pinus koraiensis)林向云冷杉针阔混交林转变,目前是我国东北林区最常见的森林类型之一[20]。桦木属(Betula)树种是云冷杉林中常见树种,它们材质较硬,纹理美观,被广泛应用于家具和地板行业。另一方面,桦木属树种具有较强的自我稀疏能力,对森林生态系统的稳定性和森林更新具有重要意义[15],其中枫桦(Betula costata)是天然云冷杉针阔混交林重要的伴生树种[21],在森林演替过程中承担着不可替代的角色,其与林分主要优势种的比例变化是判断森林群落所处演替阶段的重要标志[22],而现阶段针对枫桦的天然更新研究还属于空白。本研究以东北云冷杉针阔混交林枫桦种群为研究对象,对其种子雨的时空分布规律及种子萌发能力进行研究,研究结果将有助于揭示枫桦自然更新规律、物种共存机制以及预测森林群落演替方向等方面,并为维持温带森林生态系统物种多样性提供科学参考。
1. 研究区概况和研究方法
1.1 研究区概况
研究区位于吉林省汪清林业局的金沟岭林场(130°05′ ~ 130°20′E,43°17′ ~ 43°25′N),属长白山系老爷岭山脉雪岭支脉。该区地貌属低山丘陵,海拔300 ~ 1 200 m,坡度5° ~ 25°。该区属季风型气候,1月份气温最低,平均气温−32 ℃,7月份最高,平均气温32 ℃,全年平均气温为4 ℃,年降水量600 ~ 700 mm,具有明显的季节差异,降水多集中在6—8月份。研究区土壤类型为暗棕壤。研究区主要乔木树种有红皮云杉(Picea koraiensis)、鱼鳞云杉(Picea jezoensis)、长白落叶松(Larix olgensis)、臭松(Abies nephrolepis)、红松、白桦(Betula platyphylla)、枫桦、紫椴(Tilia amurensis)、色木槭(Acer mono)、水曲柳(Fraxinus mandshurica)、青楷槭(Acer tegmentosum)等。
1.2 研究方法
1.2.1 样地设置和调查
2013年在研究区同一林班内设立9块面积为1 hm2(100 m × 100 m)的永久样地,样地基本概况见表1,按相邻格子法将每块样地划分成10 m × 10 m的网格,并对样地内胸径大于1 cm的所有木本植物进行每木检尺,测量并记录林木树种、树高、胸径、冠幅、林木相对空间位置等信息。
表 1 云冷杉针阔混交林样地基本概况Table 1. Basic condition of mixed spruce-fir coniferous-broadleaved forest样地
Sample plot地理坐标
Geographical coordinate海拔
Altitude/m坡向
Slope
aspect坡度
Slope
degree/(°)平均胸径
Mean
DBH/cm平均树高
Mean tree
height/m林分密度/
(株·hm−2)
Stand density/
(tree·ha−1)枫桦密度
(DBH > 15 cm)/(株·hm−2)
Density of Betula costata
(DBH > 15 cm)/(tree·ha−1)YLK-1 130°15′09″E 43°22′46″N 760 东北 Northeast 15 19.0 15.1 1 232 72 YLK-2 130°15′16″E 43°22′50″N 773 东北 Northeast 16 18.2 15.1 1 165 56 YLK-3 130°15′14″E 43°22′46″N 780 东北 Northeast 5 17.4 15.8 1 328 83 YLK-4 130°15′13″E 43°22′43″N 771 东北 Northeast 5 18.5 15.5 1 539 123 YLK-5 130°15′00″E 43°23′05″N 732 东北 Northeast 5 18.6 14.9 1 309 27 YLK-6 130°15′05″E 43°23′04″N 749 东北 Northeast 5 19.9 16.5 1 436 60 YLK-7 130°15′09″E 43°23′04″N 759 东北 Northeast 5 18.8 15.7 1 195 73 YLK-8 130°15′15″E 43°23′04″N 769 东北 Northeast 5 18.4 16.3 1 301 56 YLK-9 130°15′20″E 43°23′05″N 773 东北 Northeast 3 17.6 16.5 1 437 70 1.2.2 种子收集器布设和种子收集
在每块样地内按照对角线五点取样法布设种子收集器,即在四角与样地中心的对角线中点和样地中心位置各布设1个种子收集器(如图1),共布设种子收集器45个,其中每个收集器由收集框和立柱组成,收集框由4根长为80 cm的PVC管和网孔为1 mm的纱网组成,立柱是由4根长1 m的PVC管组成,用于支撑收集框,使用大小合适的PVC立体三通将立柱和收集框连接在一起组成种子收集器。于2018年7月末开始种子的第一次收集,以后每月月末收集一次,直到11月末种子完全从母树上脱落时结束。枫桦母树的胸径基本均超过15 cm,故以此统计样地内枫桦母树密度。
1.2.3 种子处理、鉴定与计数
将收集到的种子样品于实验室内进行分类、筛选、鉴定、计数。由于枫桦种子个体小数量大,故采用四分法计数种子。将种子样品放置在光滑玻璃板上,用分样板将种子混合均匀后摊平,要求厚度不超过1 cm,沿对角线将样品分成4个三角形,再取两个对顶三角形内的样品继续按以上方法分取,直到分到样品既能代表整个样本又方便计数为止。
1.2.4 种子萌发能力测定
先将两层滤纸置于培养皿底部,加水浸泡,使滤纸充分湿润。将每个样地收集的枫桦种子混合均匀后随机抽取300粒种子,随机分成3组(即3个重复),每组100粒,称质量标号后放入培养皿中。将培养皿放置在(30 ± 2)℃的培养箱内,在自然光条件下进行发芽试验,第2天起开始记录发芽数目,连续7 d无种子萌发时停止发芽试验,分别计算种子千粒质量、发芽势和发芽率[23]。
P0=N0N×100% (1) 式中:P0为种子发芽率,N0为发芽种子数,N为种子总数。
P1=N1N×100% (2) 式中:P1为种子发芽势,N1为发芽高峰期的累计发芽总数。
1.3 数据分析
D=NnumA (3) 式中:D为该收集器种子雨密度,Nnum为收集器收集到的种子数量,A为收集器面积。
种子雨的分布类型主要有聚集分布、随机分布和均匀分布。本文采用方差/均值(Cx
)、负二项指数(K)、聚集度指标(David和Moore丛生指标,I)[24]、Cassie. R. M.指标(CA )4种空间分布类型评价方法判别种子雨空间分布状态,具体分级见表2。 表 2 各空间变异参数计算公式与分级Table 2. Formulas and classification of spatial variation parameters参数 Parameter 公式 Formula 分级 Classification 方差/均值 Variance/mean (Cx) Cx=S2ˉx Cx < 1 (均匀分布 Regular distribution);
Cx > 1 (聚集分布 Aggregated distribution);
Cx = 1 (随机分布 Random distribution)负二项指数 Negative binomial parameter (K) K=ˉx2(S2−ˉx) K < 0 (非聚集分布 Non-aggregated distribution);
K > 0 (聚集分布 Aggregated distribution)聚集度指标 Aggregation index (I) I=S2ˉx−1 I < 0 (均匀分布 Regular distribution);
I > 0 (聚集分布 Aggregated distribution);
I = 0 (随机分布 Random distribution)Cassie. R. M.指标 Cassie. R. M. index (CA) CA=(S2−ˉx)ˉx2 CA < 0 (均匀分布 Regular distribution);
CA > 0 (聚集分布 Aggregated distribution);
CA = 0 (随机分布 Random distribution)注:S2为样地所有收集器收集到种子总数的方差,ˉx为样地收集器收集到种子数量的均值。Notes: S2 is the variance of total number of seeds collected by all collectors in the sample plot, ˉx is the average number of seeds collected by all collectors in the sample plot. 对于不同样地不同时间采集的种子数量,首先验证数据是否符合正态分布和方差齐性,若符合,利用方差分析进行不同时间采集的种子雨数量显著性差异分析(P < 0.05);若不符合,利用非参数检验法(Kruskal-Wallis检验法)进行显著性差异分析。以上统计分析均由软件SPSS19.0完成。
2. 结果与分析
2.1 种子雨时间分布特征
在整个研究期间,共收集到69 100粒种子,落种时间从7月末8月初开始,直至10月末结束,持续向地面输入种子时间约60 d,且落种高峰期在10月份。Kruskal-Wallis检验结果表明不同时间收集的种子密度(D)差异显著(P < 0.05),10月份种子雨密度(2 155.73 粒/m2)显著高于8月份(9.20 粒/m2)(如表3)。此外,不同样地间种子雨密度差异较大,9月份9号样地种子雨密度最大,为(678.13 ± 199.32)粒/m2,1号样地最低,为(87.50 ± 32.07)粒/m2;10月份9号样地种子雨密度最大,为(4 856.25 ± 558.52)粒/m2,2号样地最低,为(108.98 ± 71.75)粒/m2。
表 3 各样地枫桦种子雨密度 粒/m2Table 3. Density of Betula costata seed rain of each sample plotseed/m2 样地号 Sample plot No. 7月 July 8月 August 9月 September 10月 October 11月 November 1 0.00 ± 0.00b 0.00 ± 0.00b 87.5 ± 32.07ab 222.92 ± 9.36a 0.00 ± 0.00b 2 0.00 ± 0.00b 4.69 ± 4.19b 158.75 ± 14.37a 108.98 ± 71.75ab 0.00 ± 0.00b 3 0.00 ± 0.00b 2.81 ± 1.68b 172.27 ± 7.21ab 661.88 ± 212.92a 0.00 ± 0.00b 4 0.00 ± 0.00b 15.31 ± 13.35b 118.36 ± 34.18ab 1 645.63 ± 481.73a 0.00 ± 0.00b 5 0.00 ± 0.00b 1.25 ± 1.12b 492.19 ± 139.33ab 1 405.94 ± 487.45a 0.00 ± 0.00b 6 0.00 ± 0.00b 2.19 ± 1.96b 212.5 ± 172.62ab 1 882.81 ± 455.40a 0.00 ± 0.00b 7 0.00 ± 0.00b 1.56 ± 1.40b 309.06 ± 110.07ab 4 303.75 ± 980.00a 0.00 ± 0.00b 8 0.00 ± 0.00b 53.13 ± 21.47ab 426.25 ± 96.68ab 4 313.44 ± 1 302.08ab 0.00 ± 0.00b 9 0.00 ± 0.00b 1.88 ± 1.12b 678.13 ± 199.32ab 4856.25 ± 558.52a 0.00 ± 0.00b 平均值 Mean 0.00 9.20 295.00 2155.73 0.00 注:数据为平均值±标准误。不同字母表示同一样地不同收集时间之间种子密度差异显著(P < 0.05)。Notes: data is mean ± SE. Different letters mean significant differences in same sample plot at varied collecting time (P < 0.05). 2.2 种子雨空间分布特征
经4种空间分布类型评价方法分析,Cx
、K、I、CA的平均值分别为1 116.3、4.75、1 115.3、0.41,不同指标均显示该研究区枫桦种子呈聚集分布(表4),所有样地4个指标值均远大于临界值,表明枫桦种子扩散时聚集程度很高。不同样地间指标差异较大,表示种子空间聚集程度差异较大,其中4号样地Cx值高达3 166.31,种子聚集程度要远高于其他样地;1号样地最小,Cx 值为20.24,说明种子空间聚集程度最小。 表 4 枫桦种子雨空间分布特征Table 4. Spatial distribution characteristics of seed rain of Betula costata样地号
Sample plot No.Cx 分布格局
Distribution patternK 分布格局
Distribution patternI 分布格局
Distribution patternCA 分布格局
Distribution pattern1 20.24 聚集 Aggregated 17.19 聚集 Aggregated 19.24 聚集 Aggregated 0.06 聚集 Aggregated 2 86.13 聚集 Aggregated 3.01 聚集 Aggregated 85.13 聚集 Aggregated 0.33 聚集 Aggregated 3 462.49 聚集 Aggregated 1.78 聚集 Aggregated 461.49 聚集 Aggregated 0.56 聚集 Aggregated 4 3 166.31 聚集 Aggregated 1.04 聚集 Aggregated 3 165.31 聚集 Aggregated 0.96 聚集 Aggregated 5 971.74 聚集 Aggregated 1.96 聚集 Aggregated 970.74 聚集 Aggregated 0.51 聚集 Aggregated 6 763.18 聚集 Aggregated 2.82 聚集 Aggregated 762.18 聚集 Aggregated 0.35 聚集 Aggregated 7 1 532.86 聚集 Aggregated 3.01 聚集 Aggregated 1 531.86 聚集 Aggregated 0.33 聚集 Aggregated 8 2 489.16 聚集 Aggregated 1.93 聚集 Aggregated 2 488.16 聚集 Aggregated 0.52 聚集 Aggregated 9 554.61 聚集 Aggregated 10.00 聚集 Aggregated 553.61 聚集 Aggregated 0.10 聚集 Aggregated 均值 Mean 1 116.30 4.75 1 115.30 0.41 2.3 种子萌发特性
由于种子集中分布在9月和10月,其他月份较少,故仅选择9月和10月的种子进行种子萌发试验分析。
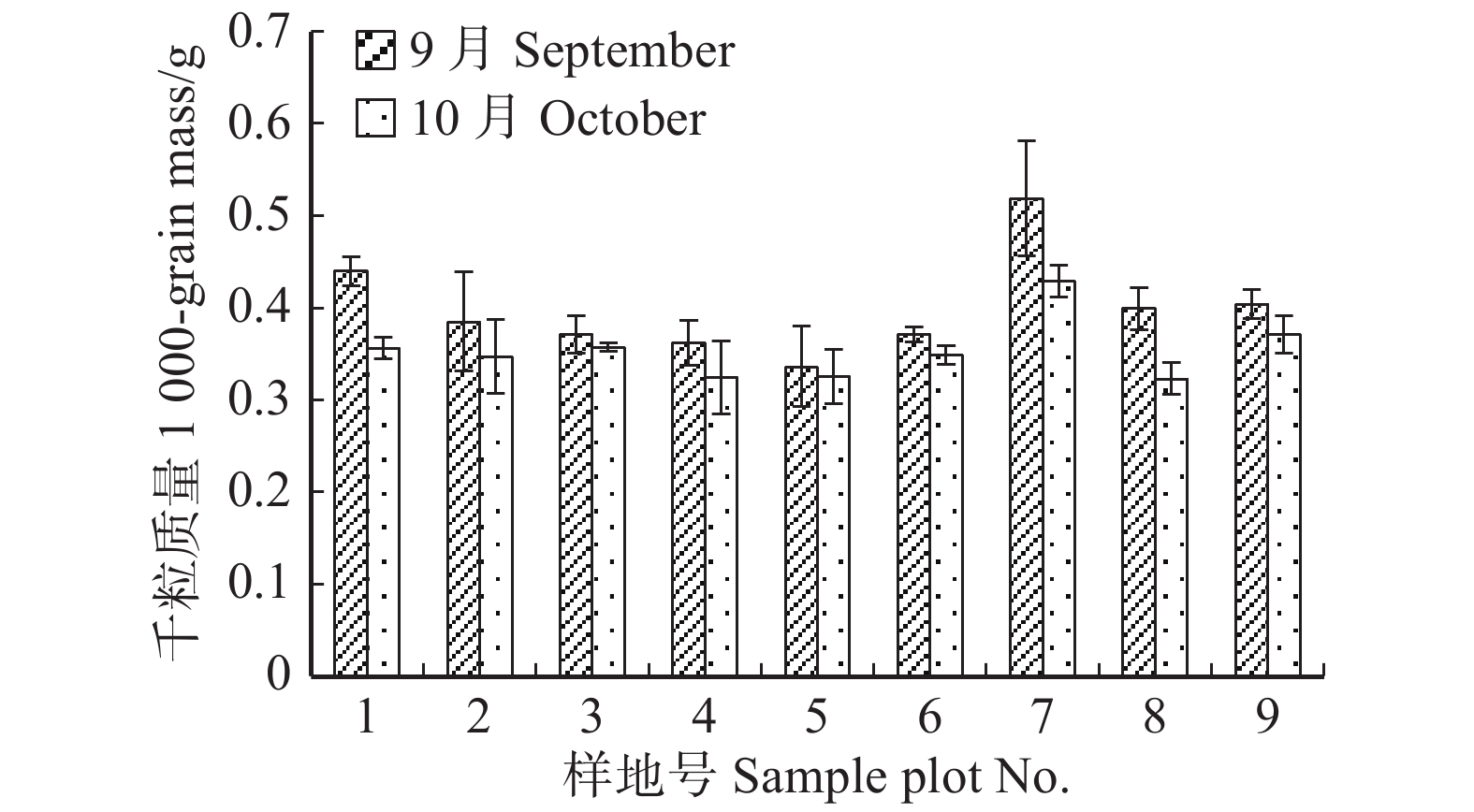
T检验结果表明,不同收集时间种子千粒质量差异极显著(P < 0.01),其中9月种子质量显著大于10月份种子质量。对不同收集时间种子千粒质量进行分析表明(如图2),9月平均千粒质量为(0.398 ± 0.064)g(平均值 ± 标准差,下同),其中7号样地最大,为(0.519 ± 0.062)g,5号样地最小,为(0.336 ± 0.044)g;10月份种子平均千粒质量为(0.353 ± 0.040)g,其中7号样地最大,为(0.429 ± 0.018)g,8号样地最小,为(0.323 ± 0.017)g。
![]() 图 2 云冷杉针阔混交林各样地种子千粒质量Figure 2. 1 000-grain mass of spruce-firconiferous-broadleaved forest in each sample plot
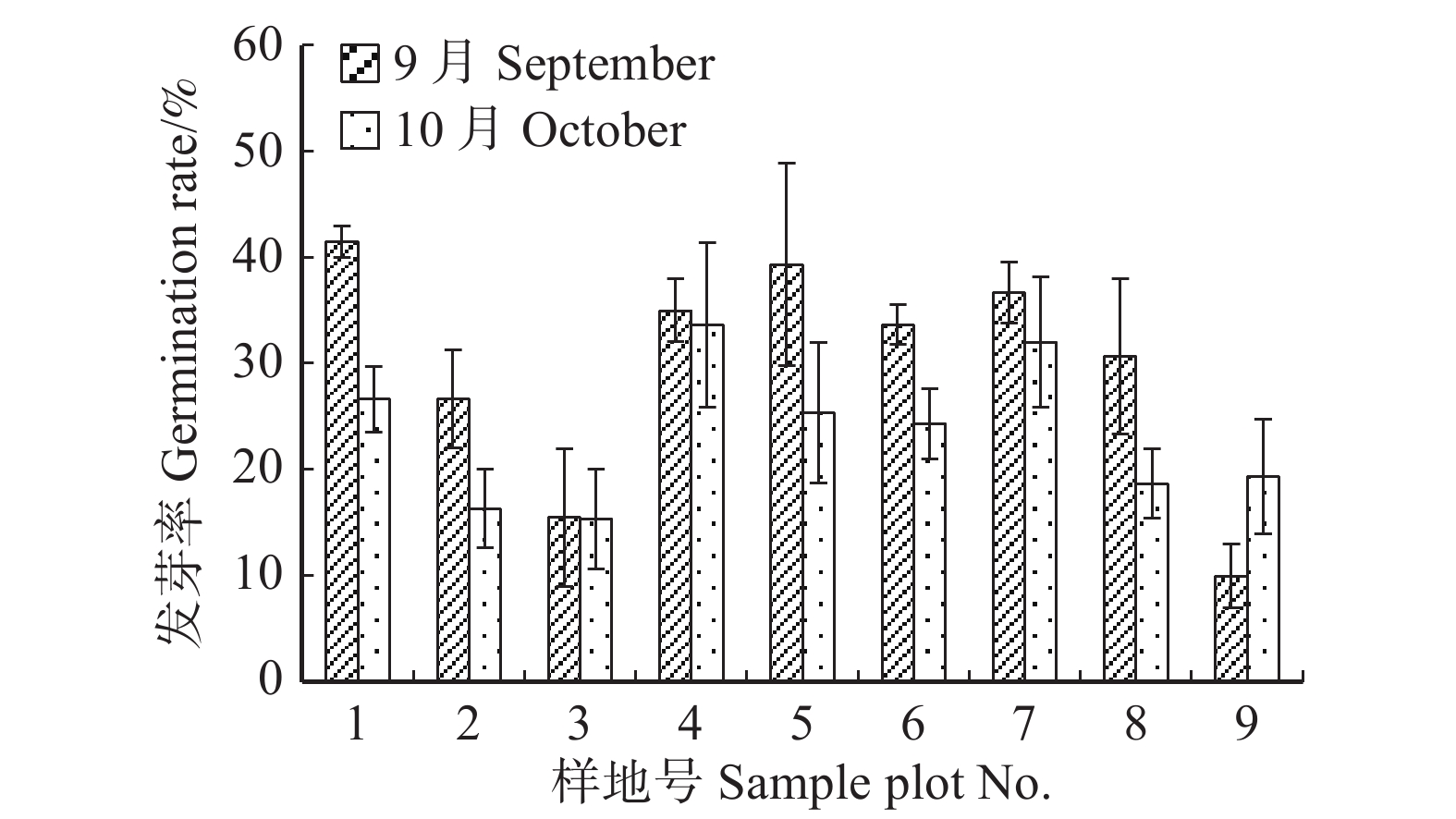
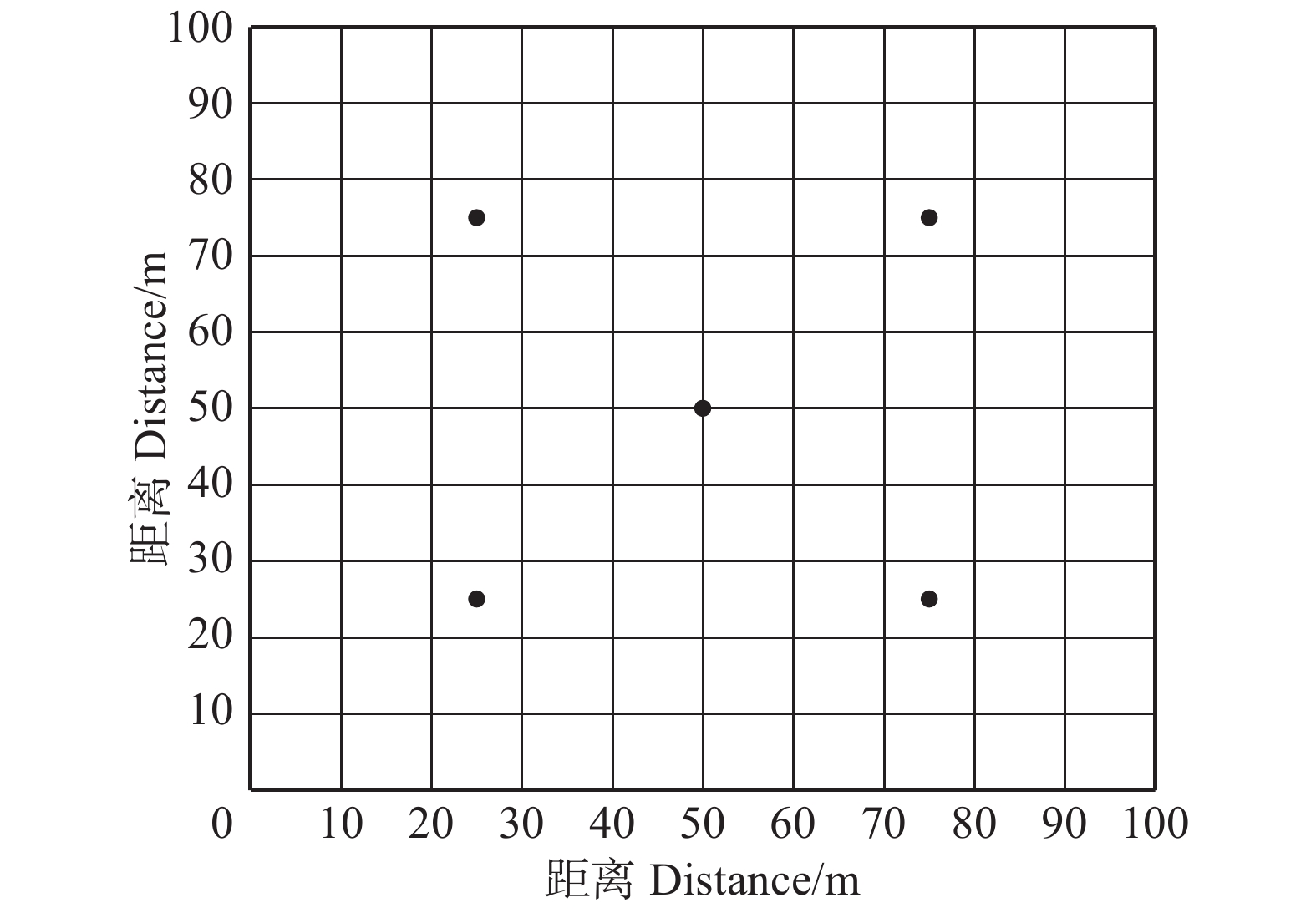
图 2 云冷杉针阔混交林各样地种子千粒质量Figure 2. 1 000-grain mass of spruce-firconiferous-broadleaved forest in each sample plot枫桦种子发芽快,种子无休眠期,第2天开始萌发。种子开始萌发至种子萌发结束约14 d,第6天时种子发芽势(P1)达到最高,9月最大发芽势为26.25%,10月为21.22%。T检验结果表明,不同时间收集的枫桦种子发芽率(P0)差异显著(P = 0.036 < 0.05)。萌发试验结果表明(如图3),在9月份收集的种子平均发芽率为(29.84 ± 11.68)%,其中1号地发芽率最大,为(41.50 ± 1.50)%,9号地最小,为(10.00 ± 3.00)%;在10月份收集的种子平均发芽率为(23.52 ± 8.23)%,4号地种子发芽率最大,为(33.67 ± 7.76)%,3号地最小,为(15.33 ± 4.71)%。
![]() 图 3 云冷杉针阔混交林各样地种子发芽率Figure 3. Seed germination rate of spruce-fir coniferous-broadleaved forest in each sample plot
图 3 云冷杉针阔混交林各样地种子发芽率Figure 3. Seed germination rate of spruce-fir coniferous-broadleaved forest in each sample plot3. 讨 论
3.1 种子雨时间动态
一般来说,种子大小和种子产量呈显著负相关[25],种子越小,种子产量越高,在7—11月期间,共收集到枫桦种子69 100粒,种子产量远高于长白山地区内大部分树种的种子产量[26],说明该地区枫桦种源充足,天然更新潜力较高。时间动态是种子雨的主要特征之一[27],主要表现为落种起始期、高峰期和消退期,本研究中枫桦种子雨持续时间为60 d左右,8月份开始落种,高峰期出现在10月份,枫桦种子成熟时间与当地主要树种(如紫椴、槭树、和怀槐(Maackia amurensis))的成熟时间一致[28]。杜彦君等[29]认为种子在秋季成熟与幼苗定居有关联,种子散落后要经历寒冷的冬天,萌发的幼苗不能过冬。另一方面,种子秋季成熟这一特性有利于种子扩散,动物和鸟类会在秋季寻找食物以准备入冬,从而将种子扩散到更远的地方。另外,虽然枫桦种子雨密度远超过长白山次生针阔混交林其他物种,但在调查中并未发现较多的枫桦幼苗,这与陈香茗等[30]在长白山地区调查的幼苗结果一致,表明枫桦虽然具有充足的种源,但种子野外萌发率不高,说明该林分条件已经不太适合枫桦种子萌发,可能面临着在群落演替进程中被逐步淘汰的风险。
3.2 种子雨空间分布
种子雨空间分布反映了种群对环境的适应性[31],同一林分内枫桦种子分布不均匀,种子扩散与母树位置、数量、高度、结实量,落种时林内风速温度及林分坡度等环境因素有关[8],是由多种因素共同作用的结果。1号和2号地枫桦种子密度远小于其他样地,但1号和2号样地坡度高于其他样地,可能是由于坡度较大导致土壤养分和水分流失,从而母树结实量出现较大差异。
一般来说,种子会围绕母树呈聚集性分布,云冷杉针阔混交林中枫桦种子在整个林分中呈聚集性分布,且聚集程度较大。Masaki等[32]研究发现日本温带森林中的桦木科种子也呈聚集分布,同样以风力传播为主的紫椴[30]和油松(Pinus tabuliformis)[16]种子也存在类似的现象。但无论种子具有怎样的传播方式,种子围绕母树的聚集分布会导致当地群落产生较大的空间变异,从而影响森林的群落结构和物种多样性。聚集分布会增大种内竞争,种子密度越高的区域,种子转化为幼苗的概率越低[27]。枫桦种子的高强度聚集分布表明枫桦种子扩散受到限制,可能是由于林分郁闭度和林分密度都较大,导致种子扩散时风速较小,种子不能远距离扩散,从而呈现聚集性分布。
3.3 种子萌发特性
研究表明枫桦母树结实量大,但不同落种时间种子萌发能力不同。种子发芽差异与贮藏蛋白含量差异有关[33],通常情况下,种子越大,种子内相对贮藏蛋白含量越高[34],种子萌发能力越强。9月份枫桦种子千粒质量与发芽能力均高于10月份,这可能和9月份枫桦种子内贮藏蛋白更高有关系。在森林演替进程中随着树木年龄的增长,小种子物种数量减少,大种子物种数量增加[35],本文研究结果也显示,枫桦种子小,在实验室内,种子萌发率低,而在林分内,受更多因素限制,枫桦种子萌发率更会远远低于实验室条件下[36]。这可能是枫桦数量减少的原因之一。
4. 结 论
东北云冷杉针阔混交林枫桦种子雨具有明显的时间分布特征,10月份为种子雨高峰期。由于枫桦种子的扩散能力受到限制使得种子在空间上呈聚集分布。枫桦结实量大,种源充足,有着良好的天然更新潜力,但枫桦种子小,萌发能力较差。枫桦种群在森林演替过程中将可能面临着被淘汰的风险,为了更好地把握森林演替动态,多区域的持续监测和调查是非常有必要的。
-
![]()
图 2 云冷杉针阔混交林各样地种子千粒质量
Figure 2. 1 000-grain mass of spruce-firconiferous-broadleaved forest in each sample plot
![]()
图 3 云冷杉针阔混交林各样地种子发芽率
Figure 3. Seed germination rate of spruce-fir coniferous-broadleaved forest in each sample plot
表 1 云冷杉针阔混交林样地基本概况
Table 1 Basic condition of mixed spruce-fir coniferous-broadleaved forest
样地
Sample plot地理坐标
Geographical coordinate海拔
Altitude/m坡向
Slope
aspect坡度
Slope
degree/(°)平均胸径
Mean
DBH/cm平均树高
Mean tree
height/m林分密度/
(株·hm−2)
Stand density/
(tree·ha−1)枫桦密度
(DBH > 15 cm)/(株·hm−2)
Density of Betula costata
(DBH > 15 cm)/(tree·ha−1)YLK-1 130°15′09″E 43°22′46″N 760 东北 Northeast 15 19.0 15.1 1 232 72 YLK-2 130°15′16″E 43°22′50″N 773 东北 Northeast 16 18.2 15.1 1 165 56 YLK-3 130°15′14″E 43°22′46″N 780 东北 Northeast 5 17.4 15.8 1 328 83 YLK-4 130°15′13″E 43°22′43″N 771 东北 Northeast 5 18.5 15.5 1 539 123 YLK-5 130°15′00″E 43°23′05″N 732 东北 Northeast 5 18.6 14.9 1 309 27 YLK-6 130°15′05″E 43°23′04″N 749 东北 Northeast 5 19.9 16.5 1 436 60 YLK-7 130°15′09″E 43°23′04″N 759 东北 Northeast 5 18.8 15.7 1 195 73 YLK-8 130°15′15″E 43°23′04″N 769 东北 Northeast 5 18.4 16.3 1 301 56 YLK-9 130°15′20″E 43°23′05″N 773 东北 Northeast 3 17.6 16.5 1 437 70  下载: 导出CSV
下载: 导出CSV
表 2 各空间变异参数计算公式与分级
Table 2 Formulas and classification of spatial variation parameters
参数 Parameter 公式 Formula 分级 Classification 方差/均值 Variance/mean (Cx) Cx=S2ˉx Cx < 1 (均匀分布 Regular distribution);Cx > 1 (聚集分布 Aggregated distribution);Cx = 1 (随机分布 Random distribution)负二项指数 Negative binomial parameter (K) K=ˉx2(S2−ˉx) K < 0 (非聚集分布 Non-aggregated distribution);K > 0 (聚集分布 Aggregated distribution)聚集度指标 Aggregation index (I) I=S2ˉx−1 I < 0 (均匀分布 Regular distribution);I > 0 (聚集分布 Aggregated distribution);I = 0 (随机分布 Random distribution)Cassie. R. M.指标 Cassie. R. M. index (CA) CA=(S2−ˉx)ˉx2 CA < 0 (均匀分布 Regular distribution);CA > 0 (聚集分布 Aggregated distribution);CA = 0 (随机分布 Random distribution)注:S2为样地所有收集器收集到种子总数的方差, ˉx 为样地收集器收集到种子数量的均值。Notes: S2 is the variance of total number of seeds collected by all collectors in the sample plot,ˉx is the average number of seeds collected by all collectors in the sample plot.
下载: 导出CSV
表 3 各样地枫桦种子雨密度 粒/m2
Table 3 Density of Betula costata seed rain of each sample plot
seed/m2 样地号 Sample plot No. 7月 July 8月 August 9月 September 10月 October 11月 November 1 0.00 ± 0.00b 0.00 ± 0.00b 87.5 ± 32.07ab 222.92 ± 9.36a 0.00 ± 0.00b 2 0.00 ± 0.00b 4.69 ± 4.19b 158.75 ± 14.37a 108.98 ± 71.75ab 0.00 ± 0.00b 3 0.00 ± 0.00b 2.81 ± 1.68b 172.27 ± 7.21ab 661.88 ± 212.92a 0.00 ± 0.00b 4 0.00 ± 0.00b 15.31 ± 13.35b 118.36 ± 34.18ab 1 645.63 ± 481.73a 0.00 ± 0.00b 5 0.00 ± 0.00b 1.25 ± 1.12b 492.19 ± 139.33ab 1 405.94 ± 487.45a 0.00 ± 0.00b 6 0.00 ± 0.00b 2.19 ± 1.96b 212.5 ± 172.62ab 1 882.81 ± 455.40a 0.00 ± 0.00b 7 0.00 ± 0.00b 1.56 ± 1.40b 309.06 ± 110.07ab 4 303.75 ± 980.00a 0.00 ± 0.00b 8 0.00 ± 0.00b 53.13 ± 21.47ab 426.25 ± 96.68ab 4 313.44 ± 1 302.08ab 0.00 ± 0.00b 9 0.00 ± 0.00b 1.88 ± 1.12b 678.13 ± 199.32ab 4856.25 ± 558.52a 0.00 ± 0.00b 平均值 Mean 0.00 9.20 295.00 2155.73 0.00 注:数据为平均值±标准误。不同字母表示同一样地不同收集时间之间种子密度差异显著(P < 0.05)。Notes: data is mean ± SE. Different letters mean significant differences in same sample plot at varied collecting time (P < 0.05).
下载: 导出CSV
表 4 枫桦种子雨空间分布特征
Table 4 Spatial distribution characteristics of seed rain of Betula costata
样地号
Sample plot No.Cx 分布格局
Distribution patternK 分布格局
Distribution patternI 分布格局
Distribution patternCA 分布格局
Distribution pattern1 20.24 聚集 Aggregated 17.19 聚集 Aggregated 19.24 聚集 Aggregated 0.06 聚集 Aggregated 2 86.13 聚集 Aggregated 3.01 聚集 Aggregated 85.13 聚集 Aggregated 0.33 聚集 Aggregated 3 462.49 聚集 Aggregated 1.78 聚集 Aggregated 461.49 聚集 Aggregated 0.56 聚集 Aggregated 4 3 166.31 聚集 Aggregated 1.04 聚集 Aggregated 3 165.31 聚集 Aggregated 0.96 聚集 Aggregated 5 971.74 聚集 Aggregated 1.96 聚集 Aggregated 970.74 聚集 Aggregated 0.51 聚集 Aggregated 6 763.18 聚集 Aggregated 2.82 聚集 Aggregated 762.18 聚集 Aggregated 0.35 聚集 Aggregated 7 1 532.86 聚集 Aggregated 3.01 聚集 Aggregated 1 531.86 聚集 Aggregated 0.33 聚集 Aggregated 8 2 489.16 聚集 Aggregated 1.93 聚集 Aggregated 2 488.16 聚集 Aggregated 0.52 聚集 Aggregated 9 554.61 聚集 Aggregated 10.00 聚集 Aggregated 553.61 聚集 Aggregated 0.10 聚集 Aggregated 均值 Mean 1 116.30 4.75 1 115.30 0.41
下载: 导出CSV
-
[1] 杜彦君, 马克平. 森林种子雨研究进展与展望[J]. 生物多样性, 2012, 20(1):94−107. doi: 10.3724/SP.J.1003.2012.14161. Du Y J, Ma K P. Advancements and prospects in forest seed rain studies[J]. Biodiversity Science, 2012, 20(1): 94−107. doi: 10.3724/SP.J.1003.2012.14161.
[2] 于顺利, 郎南军, 彭明俊, 等. 种子雨研究进展[J]. 生态学杂志, 2007, 26(10):1646−1652. Yu S L, Lang N J, Peng M J, et al. Research advances in seed rain[J]. Chinese Journal of Ecology, 2007, 26(10): 1646−1652.
[3] Martinez R M, Castro A S. Seed rain and advanced regeneration in a tropical rain forest[M]//Frugivory and seed dispersal: ecological and evolutionary aspects. Dordrecht: Springer, 1993: 299−318.
[4] Wang B C, Smith T B. Closing the seed dispersal loop[J]. Trends in Ecology & Evolution, 2002, 17: 379−386.
[5] Perini M, Dias H M, Kunz S H. The role of environmental heterogeneity in the seed rain pattern[J]. Florestae Ambiente, 2019, 26(9): 1−10.
[6] Seidler T G, Plotkin J B. Seed dispersal and spatial pattern in tropical trees[J]. PLoS Biology, 2006, 4(11): 2132−2137.
[7] Traveset A, Heleno R, Nogales M. The ecology of seed dispersal[M]// Seeds: the ecology of regeneration in plant communities. New York: CAB International Publishing, 2014: 85−110.
[8] Snell R S, Beckman N G, Fricke E, et al. Consequences of intraspecific variation in seed dispersal for plant demography, communities, evolution and global change[J]. AoB Plants, 2019, 11(4): 1−19.
[9] Jara-Guerrero A, Escribano-Avila G, Espinosa C I, et al. White tailed deer as the last megafauna dispersing seeds in Neotropical dry forests: the role of fruit and seed traits[J]. Biotropica, 2018, 50(1): 169−177. doi: 10.1111/btp.12507.
[10] Urrea-Galeano L A, Andresen E, Coates R, et al. Dung beetle activity affects rain forest seed bank dynamics and seedling establishment[J]. Biotropica, 2019, 51(2): 186−195. doi: 10.1111/btp.12631.
[11] Janzen D H. Seed predation by animals[J]. Annual Review of Ecology and Systematics, 1971, 2: 465−492. doi: 10.1146/annurev.es.02.110171.002341.
[12] Solvertown J W. The evolutionary ecology of mast seeding in trees[J]. Biological Journal of the Linnean Society, 1980(14): 235−250.
[13] Silvertown J W. The evolutionary ecology of mast seeding in trees[J]. Biological Journal of the Linnean Society, 1980, 14(2): 235−250.
[14] 程积民, 赵凌平, 程杰. 子午岭60年辽东栎林种子质量与森林更新[J]. 北京林业大学学报, 2009, 31(2):10−16. doi: 10.3321/j.issn:1000-1522.2009.02.002. Cheng J M, Zhao L P, Cheng J. Seed quality and forest regeneration of a 60-year Quercus liaotungensis forest in the Ziwuling region, northwestern China[J]. Journal of Beijing Forestry University, 2009, 31(2): 10−16. doi: 10.3321/j.issn:1000-1522.2009.02.002.
[15] Houle G. Seed dispersal and seedling recruitment of Betula alleghaniensis: spatial inconsistency in time[J]. Ecology, 1998, 79(3): 807−818. doi: 10.1890/0012-9658(1998)079[0807:SDASRO]2.0.CO;2.
[16] 陈智平, 张涛, 赵万奎, 等. 黄土高原子午岭林区油松林种子雨强度及时空动态特征[J]. 中国沙漠, 2020(3):1−9. Chen Z P, Zhang T, Zhao W K, et al. Seed rain intensity and spatio-temporal dynamic characteristics of Pinus tabulifornis in Ziwu Mountain forest area of Loess Plateau of China[J]. Journal of Desert Research, 2020(3): 1−9.
[17] 钱多多, 匡旭, 王绪高, 等. 2006—2017年长白山阔叶红松林木本植物种子雨的时空动态[J]. 应用生态学报, 2019, 30(5):1487−1493. Qian D D, Kuang X, Wang X G, et al. Spatio-temporal dynamics of woody plants seed rains in broad-leaved Korean pine mixed forest in Changbai Mountains form 2006 to 2017, China[J]. The Journal of Applied Ecology, 2019, 30(5): 1487−1493.
[18] 杨彬, 王玉, 郝清玉. 木麻黄海防林种子雨的时空动态[J]. 热带亚热带植物学报, 2019, 27(4):367−375. doi: 10.11926/jtsb.4033. Yang B, Wang Y, Hao Q Y. Spatial and temporal dynamics of seed rain of Casuarina equisetifolia coastal protection forest[J]. Journal of Tropical and Subtropical Botany, 2019, 27(4): 367−375. doi: 10.11926/jtsb.4033.
[19] 肖静, 黄力, 杨超, 等. 缙云山常绿阔叶林种子雨组成及其时空动态[J]. 林业科学, 2019, 55(7):163−169. doi: 10.11707/j.1001-7488.20190718. Xiao J, Huang L, Yang C, et al. Composition and spatial-temporal variation of the seed rain in an evergreen broad-leaved forest on Jinyun Mountain[J]. Scientia Silvae Sinicae, 2019, 55(7): 163−169. doi: 10.11707/j.1001-7488.20190718.
[20] 张春雨, 赵秀海, 夏富才. 长白山次生林树种空间分布及环境解释[J]. 林业科学, 2008, 44(8):1−8. doi: 10.3321/j.issn:1001-7488.2008.08.001. Zhang C Y, Zhao X H, Xia F C. Spatial distribution of tree species and environmental interpretations of secondary forest in Changbai Mountains[J]. Scientia Silvae Sinicae, 2008, 44(8): 1−8. doi: 10.3321/j.issn:1001-7488.2008.08.001.
[21] 杨亮庆, 刘一星, 蔡英春. PEG400预处理枫桦圆盘干燥特性研究[J]. 西北林学院学报, 2013, 28(4):153−157. doi: 10.3969/j.issn.1001-7461.2013.04.31. Yang L Q, Liu Y X, Cai Y C. Drying characteristics of Betula costata disk pretreated with PEG400[J]. Journal of Northwest Forestry University, 2013, 28(4): 153−157. doi: 10.3969/j.issn.1001-7461.2013.04.31.
[22] 安慧君. 阔叶红松林空间结构研究[D]. 北京: 北京林业大学, 2003. An H J. Study on the spatial structure of the broad-leaved Korean pine forest[D]. Beijing: Beijing Forestry University, 2003.
[23] 刘宇, 徐焕文, 姜静, 等. 基于种子活力及苗期生长性状的白桦四倍体半同胞家系初选[J]. 北京林业大学学报, 2014, 36(2):74−80. Liu Y, Xu H W, Jiang J, et al. Family selection of birch tetraploid half-sibling based on seed vigor and seedling growth traits[J]. Journal of Beijing Forestry University, 2014, 36(2): 74−80.
[24] David F N, Moore P G. Notes on contagious distributions in plant populations[J]. Annals of Botany, 1954, 18(1): 47−53. doi: 10.1093/oxfordjournals.aob.a083381.
[25] Comita L S, Hubbell S P. Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank[J]. Ecology, 2009, 90(2): 328−334. doi: 10.1890/08-0451.1.
[26] 王蕾. 长白山针阔混交林种子雨动态及扩散限制研究[D]. 北京: 北京林业大学, 2010. Wang L. Seed rain dynamics and dispersal limitations in mixed broadleaf-conifer forest in Changbai Mountain[D]. Beijing: Beijing Forestry University, 2010.
[27] Yang Y C, Huang L, Qian S H, et al. Completing the life history of Castanopsis fargesii: changes in the seed dispersal, seedling and sapling recruitment patterns[J]. European Journal of Forest Research, 2015, 134(6): 1143−1154. doi: 10.1007/s10342-015-0916-9.
[28] 张健, 郝占庆, 李步杭, 等. 长白山阔叶红松(Pinus koraiensis)林种子雨组成及其季节动态[J]. 生态学报, 2008, 28(6):2445−2454. doi: 10.3321/j.issn:1000-0933.2008.06.002. Zhang J, Hao Z Q, Li B H, et al. Composition and seasonal dynamics of seed rain in broad-leaved Korean pine (Pinus koraiensis) mixed forest, Changbai Mountain[J]. Acta Ecologica Sinica, 2008, 28(6): 2445−2454. doi: 10.3321/j.issn:1000-0933.2008.06.002.
[29] 杜彦君, 马克平. 浙江古田山自然保护区常绿阔叶林种子雨的时空变异[J]. 植物生态学报, 2012, 36(8):717−728. Du Y J, Ma K P. Temporal and spatial variation of seedfall in a broad-leaved evergreen forest in Gutianshan Nature Reserve of Zhejiang Province, China[J]. Chinese Journal of Plant Ecology, 2012, 36(8): 717−728.
[30] 陈香茗, 赵秀海, 夏富才, 等. 长白山紫椴种子雨的时空分布格局[J]. 东北林业大学学报, 2011, 39(1):7−10. doi: 10.3969/j.issn.1000-5382.2011.01.003. Chen X M, Zhao X H, Xia F C, et al. Spatiotemporal dynamics of seed rain of Tilia amurensis in forest stands in Changbai Mountain[J]. Journal of Northeast Forestry University, 2011, 39(1): 7−10. doi: 10.3969/j.issn.1000-5382.2011.01.003.
[31] Saastamoinen M, Bocedi G, Cote J, et al. Genetics of dispersal[J]. Biological Reviews, 2018, 93(1): 574−599. doi: 10.1111/brv.12356.
[32] Masaki T, Nakashizuka T, Niiyama K, et al. Impact of the spatial uncertainty of seed dispersal on tree colonization dynamics in a temperate forest[J]. Oikos, 2019, 128(12): 1816−1828. doi: 10.1111/oik.06236.
[33] Lin L, Fu J. Relationship between the accumulation of storage proteins and formation of vigor of peanut seed[J]. Journal of Tropical and Subtropical Botany, 1996, 4(1): 57−60.
[34] 姜文, 姚大年, 张文明, 等. 小麦种子萌发过程贮藏蛋白变化及其与活力关系的研究[J]. 种子, 2006, 25(7):16−19. doi: 10.3969/j.issn.1001-4705.2006.07.005. Jiang W, Yao D N, Zhang W M, et al. Study on relationship between seed vigor and storage protein in germinating wheat seeds[J]. Seed, 2006, 25(7): 16−19. doi: 10.3969/j.issn.1001-4705.2006.07.005.
[35] Wendt A L. Seed dispersal, seed size and seed predation in tropical regrowth forests[D]. Connecticut: University of Connecticut-Storrs, 2014.
[36] 任坚毅, 林玥, 岳明. 太白山红桦种子的萌发特性[J]. 植物生态学报, 2008, 32(4):883−890. doi: 10.3773/j.issn.1005-264x.2008.04.017. Ren J Y, Lin Y, Yue M. Seed germination characteristics of Betula albosinensis at Mountain Taibai, China[J]. Chinese Journal of Plant Ecology, 2008, 32(4): 883−890. doi: 10.3773/j.issn.1005-264x.2008.04.017.
-
期刊类型引用(12)
1. 包崇寅,孙永玉,李敏敏,邢洪铭,戚建华. 不同生境濒危植物龙棕种群结构及其动态特征. 西北植物学报. 2024(03): 479-490 .  百度学术
百度学术
2. 赵鑫,陈虹,赵善超,陈兵权,郭来珍,周昊亮. 新疆天山云杉种子雨时空动态分布及种子萌发特性. 浙江农林大学学报. 2024(03): 542-548 . 百度学术
3. 靳旭红,于聪,张庭耀,吕松瞳,刘扬,陈乐,龙生,穆怀志. 基于种子活力和苗期生长的枫桦半同胞家系初选. 植物研究. 2024(05): 763-773 . 百度学术
4. 刘志宇,张忠辉,杨凯麟,张军,姜润华,吴则甫,王琦,李文华,夏富才. 不同经营方式的云冷杉针阔混交林土壤真菌群落结构1). 东北林业大学学报. 2023(03): 124-129 . 百度学术
5. 强亚琪,范春雨,张春雨. 长白山暗针叶林群落物种多样性维持机制. 生态学报. 2023(05): 1884-1891 . 百度学术
6. 黄梓良,徐子恒,孙操稳. 青钱柳种子雨的季节动态及土壤种子库特征. 南京林业大学学报(自然科学版). 2023(02): 18-26 . 百度学术
7. 陈士刚,邹建军,山昌林,李秀红,芦静,王岗. 硕桦优树选择及不同种源、家系苗期评价. 吉林林业科技. 2023(02): 1-5+23 . 百度学术
8. 王莹,文淑均,罗定明,覃延闯,丁涛,刘世男. 珍稀濒危植物元宝山冷杉种实特征及种子萌发特性研究. 广西科学院学报. 2023(02): 161-168 . 百度学术
9. 李继祥,余登利,肖息,杨雪,田晓光,龚文斌,邓坦. 贵州宽阔水国家级自然保护区亮叶水青冈种子雨特征. 绿色科技. 2022(07): 67-69 . 百度学术
10. 李云红 ,田松岩 ,沃晓棠 ,邵英男 ,刘延坤 ,韩丽冬 ,陈瑶 ,刁云飞 ,刘玉龙 . 伴生东北红豆杉针阔混交林种子雨时空动态. 中南林业科技大学学报. 2022(05): 109-118 . 百度学术
11. 江海都,谢伟玲,柴胜丰,唐健民,蒋运生,秦惠珍,韦霄. 喀斯特地区珍贵树种黄枝油杉的种子萌发特性. 广西植物. 2022(06): 951-960 . 百度学术
12. 焦洁洁,李领寰,汪建民,孙杰杰,吴初平,姚良锦,王志高,袁位高. 午潮山杉阔混交林种子雨与土壤种子库特征研究. 林业资源管理. 2022(04): 28-35 . 百度学术
其他类型引用(2)
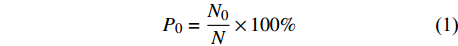
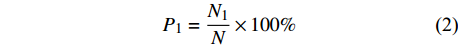
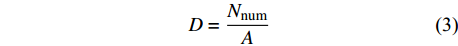
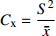
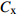
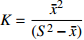
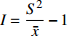
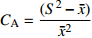
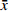
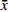
计量
- 文章访问数: 2151
- HTML全文浏览量: 607
- PDF下载量: 76
- 被引次数: 14
