
Numerical simulation of the safety influence of defects on Qijia-beams of ancient timber building
-
摘要:目的七架梁作为大型木结构古建筑的主要承重构件,其承载力安全性直接影响古建筑木结构整体的安全性。由于周围环境和使用长久等原因造成木梁出现不同程度的缺陷,会影响木梁弯曲拉应力和剪切应力的分布,进而影响其承载力安全性。因此,研究不同缺陷类型、尺寸、位置对古建筑七架梁承载力安全性的影响很有必要。方法采用Abaqus有限元软件模拟计算梁上存在的裂纹、腐朽和空洞等不同缺陷时的应力分布状态,通过量化缺陷大小和缺陷位置,对不同残损因素进行单参量数值模拟分析,确定带有缺陷木梁的最大工作应力位置,分析木梁破坏的敏感位置,探究木梁承载力安全性的变化规律。结果不同缺陷类型对七架梁安全性的影响不同,外部腐朽对七架梁承载力的影响最大,空洞缺陷次之,裂纹缺陷的影响相对最小;对于弹性阶段的受弯木梁而言,缺陷位于两下金瓜柱之间的受拉区域时,对七架梁承载力安全性的影响程度最大;不同缺陷大小对七架梁承载力的影响不同,随着木梁开裂深度、腐朽区域深度、空洞缺陷大小的增加,木梁安全性逐步降低。结论局部缺陷的存在会降低七架梁安全性。数值模拟结果可以精确算出木梁最大拉应力值,是定量研究缺陷对七架梁安全性影响和确定七架梁安全性监测位置点的良好方法。Abstract:ObjectiveThe Qijia-beams are used as the main load-bearing members for Chinese ancient timber building. The safety performance of whole wood structure is directly affected by the bearing capacity safety of Qijia-beam. Factors, such as surrounding environment and load for a long period, lead to different degrees of the defects in the wooden beam. These defects will affect the tensile stress and shear stress distribution, as well as the bearing capacity safety of the wooden beam. Therefore, it is necessary to study the influence of different defect types, sizes and locations on the safety of bearing capacity of the Qijia-beam.MethodAbaqus finite element software was used to simulate the stress state of the beam with different defects such as crack, decay and hole. By quantifying the size and location of the defects, the single parameter numerical simulation analysis of different damage factors was carried out to determine the maximum working stress location of the wood beam with defects, analyze the sensitive location of the damage of the wood beam, and investigate the variation patterns of the bearing capacity of wood beams.ResultThe results showed that the types of different defects had different influence on the safety of the Qijia-beam. External decay had the greatest influence on the bearing capacity of the Qijia-beam, followed by hole defect, and the impact of crack defect was relatively minimal. For the bending beams in the elastic stage, the defects had the greatest influence on the bearing capacity safety of the Qijia-beam when it was located in the tension zone between two under Jiagua-columns. The influence of different defect sizes on the bearing capacity of the Qijia-beam was different. With the increase of the crack depth, the decay depth, and the hole size, the safety of the wooden beam was gradually reduced.ConclusionDue to the existence of local defects, the bearing capacity of the Qijia-beams would be decreased. The research of this paper provides numerical simulation, which could accurately determine the maximum tensile stress of the wooden beam. It is a good method to quantitatively study the influence of defect on the safety of Qijia-beam and to determine the location of Qijia-beam safety monitoring.
-
随着全球气候的变化,干旱日益成为农林领域最为关注的问题之一。植物在生长过程中,生理会产生一定的响应[1−2],干旱环境可对植物生产造成显著影响,直接导致植物产量下降,限制农林产业的发展,同时也对生态环境带来一定的破环。干旱问题越来越受到人们的重视,加快干旱地区造林是解决干旱地区环境问题的有效途径之一,研究植物的抗旱机理和选育抗旱品种也是国内外专家一直研究的热点和方向。由此,在淡水资源匮乏、水土流失严重和干旱面积增加的情况下,对重要造林树种进行抗旱性综合评价,研究植物的抗旱机理,了解干旱胁迫下植物的响应和适应机制,选育优良的抗旱品种,对于改善干旱贫瘠地区的生态环境具有重要意义。
干旱胁迫会使植物的外部形态和生理生化特性发生改变,在生长指标方面,干旱环境会影响植物的根以及叶片的生长发育。光合作用是植物重要的生理过程,干旱胁迫会通过影响叶绿素的合成,进而影响光合作用。干旱还会导致植物细胞的结构和功能发生改变,细胞膜透性增加,细胞内物质大量外流,相对电导率上升,丙二醛(malondialdehyde,MDA)含量增加,同时会导致植物抗氧化酶活性和渗透调节物质发生相应变化[3−4]。
杨属(Populus)树种速生性强、繁殖速度快,其中黑杨派(Sect. Aigeiros)的美洲黑杨(P. deltoides)及其杂交种欧美杨(P. × euramericana)是优良的防护林和用材林树种,在我国林业建设中具有重要地位[5]。杨树在涵养水源、水土保持等方面发挥着不可替代的作用。发展杨树人工林是缓解全球森林资源短缺、满足社会发展需要以及促进经济发展的一项重大举措。目前,关于干旱胁迫对杨树的研究已有许多报道,大多集中在生长性状[6−7]、光合性状[8]和生理生化[9]研究等方面,但缺乏对黑杨派无性系抗旱性各项指标的综合评价。本试验利用盆栽试验方法,对10个黑杨派无性系进行干旱胁迫,研究在持续控水处理下,不同黑杨派无性系的生长及生理生化特性的变化规律,对10个无性系的抗旱性进行综合评价,筛选出抗旱能力强的无性系,以期为黑杨派无性系在干旱立地的推广应用提供依据。
1. 材料与方法
1.1 试验材料
本试验以10个黑杨派无性系为试验材料(表1),包括山东省林业科学研究院根据3年生试验林的表型性状初选的8个优良无性系,以及2个目前推广的无性系I-107和中林2025,其中I-107和中林2025作为对照。试验材料来自山东省单县国有高韦庄苗圃。
表 1 试验材料Table 1. Experimental materials无性系 Clone 拉丁学名 Latin name 母本 Female parent 父本 Male parent 1627 Populus × euramericana ‘1627’ 美洲黑杨 P. deltoides 欧洲黑杨 P. nigra 1640 P. × euramericana ‘1640’ 美洲黑杨 P. deltoides 欧洲黑杨 P. nigra 1641 P. × euramericana ‘1641’ 美洲黑杨 P. deltoides 欧洲黑杨 P. nigra 1716 P. deltoides ‘1716’ 美洲黑杨 P. deltoides 美洲黑杨 P. deltoides 1722 P. deltoides ‘1722’ 美洲黑杨 P. deltoides 美洲黑杨 P. deltoides 1723 P. deltoides ‘1723’ 美洲黑杨 P. deltoides 美洲黑杨 P. deltoides 1725 P. deltoides ‘1725’ 美洲黑杨 P. deltoides 美洲黑杨 P. deltoides 1733 P. × euramericana ‘1733’ 美洲黑杨 P. deltoides 欧洲黑杨 P. nigra I-107 P. × euramericana ‘Neva’ 美洲黑杨 P. deltoides 欧洲黑杨 P. nigra 2025 P. deltoides ‘2025’ 美洲黑杨 P. deltoides 美洲黑杨 P. deltoides 1.2 试验设计
试验在山东省林业科学研究院试验苗圃内的干旱棚中进行,采用盆栽试验方法。2021年4月中旬将各供试无性系硬枝插穗扦插于规格为350 mm × 280 mm的花盆中,每盆扦插1个插穗,每个无性系20盆。盆栽基质由苗圃熟土、草炭土、细沙按7∶2∶1的体积比例混合而成。扦插时以插穗最上面的芽刚露出土壤基质表面为宜,对试验苗木常规管理至测试时间。8月下旬每个无性系选择12株生长状况相近的苗木参加干旱试验。试验设计4个水分处理,3次重复,每盆苗木为1个重复。该试验通过人工供水后自然耗水的方法模拟干旱胁迫,预实验发现分别在自然耗水的第0、5、10、15天后,可以得到4组水分梯度:正常供水(土壤相对含水量(RWC)为80%以上)、轻度干旱(RWC为50% ~ 70%)、中度干旱(RWC为30% ~ 50%)、重度干旱(RWC为30%以下)[10]。试验前对盆栽苗木充分浇水,使土壤含水量达到饱和状态,然后通过持续控水的方法,逐渐降低土壤含水量达到水分胁迫梯度。取样时每个无性系每次选取3盆长势均一的植株,剪取从顶端数第4 ~ 6片的功能叶用于测定各项生理生化指标,取过叶片的植株下次不再取样,同时测定实时土壤含水量。
1.3 指标及其测定方法
1.3.1 生长指标的测定
生长指标包括苗高、地径、生物量等,用于研究干旱胁迫对黑杨派各无性系植株生长状况的影响。在试验开始前和结束后分别测定所有参试无性系的苗高和地径,无性系苗高(cm)用刻度尺测量,地径(mm)使用游标卡尺测量,胁迫后减去胁迫前的生长量即为胁迫期间的苗高和地径增长量。
每个无性系各定植20盆,选择12盆长势较为一致的参加试验,试验开始时,根据参试无性系的苗高均值,从剩余的8盆植株中选择3株与苗高均值相近的完整植株,用蒸馏水清洗表面杂物,将植株的根、茎、叶分开并用吸水纸迅速吸干,放入烘箱于105 ℃杀青,在75 ℃下烘至质量恒定,测定其生物量,取其均值为生物量1。试验结束时,每个无性系从参试的12盆植株中选择3株长势较为一致的完整植株,用蒸馏水清洗表面杂物,将植株的根、茎、叶分开后用吸水纸迅速吸干,放入烘箱于105 ℃杀青,并在75 ℃下烘至质量恒定,测定其生物量,记为生物量2,生物量增量则为生物量2与生物量1的差值。
1.3.2 生理生化指标的测定
采用乙醇提取比色法测定叶绿素含量,采用相对电导率法测定细胞膜透性,采用硫代巴比妥酸法测定丙二醛含量,采用氮蓝四唑光还原法测定超氧化物歧化酶(superoxide dismutase,SOD)活性,采用愈创木酚法测定过氧化物酶(peroxidase,POD)活性,采用茚三酮染色法测定游离脯氨酸含量[11]。
1.4 数据分析
运用Excel 2010软件对试验数据进行统计整理和图表绘制。用SPSS 22.0软件进行多重比较(Duncan’s新复极差法)、相关分析和主成分分析。
2. 结果与分析
2.1 干旱胁迫对黑杨派无性系生长指标的影响
在重度干旱胁迫下,10个供试无性系的各项生长指标见表2。苗高增量最大的无性系是1733、I-107,最小的是1722、2025;地径增量最大的无性系是1733、1627,最小的是1722、1640;生物量增量最大的无性系是1733、1627,最小的是2025、1722。无性系1733和1627的3个生长指标与对照2025的差异均达到显著水平,说明其具有较强的抗旱性。
表 2 土壤重度干旱对黑杨派无性系生长指标的影响Table 2. Effects of soil severe drought on differentgrowth of clones无性系
Clone苗高增量
Ramet height
increment/cm地径增量
Basal diameter
increment/mm生物量增量
Biomass
increment/g1641 16.3 ± 0.6ab 0.75 ± 0.13bc 5.67 ± 0.68abc 1733 19.6 ± 2.3a 1.02 ± 0.08a 7.04 ± 0.40a 1627 16.3 ± 0.7ab 0.89 ± 0.08ab 6.58 ± 0.43ab 2025 12.1 ± 0.9cd 0.63 ± 0.06c 4.38 ± 0.57c 1725 15.4 ± 1.4bc 0.73 ± 0.08bc 4.85 ± 1.07bc 1716 13.5 ± 1.0bcd 0.66 ± 0.07bc 5.56 ± 0.47abc 1722 10.1 ± 1.0d 0.58 ± 0.02c 4.78 ± 0.20bc I-107 16.6 ± 1.4ab 0.82 ± 0.09abc 6.28 ± 0.40abc 1723 14.9 ± 1.3bc 0.72 ± 0.05bc 5.01 ± 0.58bc 1640 15.0 ± 0.6bc 0.61 ± 0.07c 4.94 ± 0.60bc 注:同列中不同小写字母表示差异显著(P < 0.05)。下同。Notes: different lowercase letters in the same column indicate significant differences (P < 0.05). The same below. 2.2 干旱胁迫对黑杨派无性系叶绿素含量的影响
植物的叶绿素含量随着干旱胁迫的加剧而下降,不同植物在干旱胁迫下叶绿素含量出现不同程度的降低。从表3可以看出:无性系1716的叶片叶绿素含量随干旱程度加剧一直处于下降趋势,其余各无性系的叶绿素含量随干旱胁迫加剧表现出先上升后下降的规律,均在轻度干旱时叶片的叶绿素含量有所增加。随自然耗水时间的增加,土壤干旱胁迫加剧,叶绿素含量下降,在中度干旱和重度干旱处理下,所有无性系叶片的叶绿素含量均小于正常供水时。在重度干旱时,叶绿素含量较高的无性系为I-107和1627,显著高于对照2025。
表 3 土壤干旱对各无性系叶绿素含量的影响Table 3. Effects of soil drought on chlorophyll content of each clone无性系
Clone叶绿素含量 Chlorophyll content/(mg·g−1) 正常供水
Normal water
supply轻度干旱
Mild
drought中度干旱
Moderate
drought重度干旱
Severe
drought1641 2.50 ± 0.09a 3.71 ± 0.29a 1.84 ± 0.14abc 1.47 ± 0.08bcd 1733 2.32 ± 0.24abc 3.53 ± 0.14a 2.09 ± 0.14a 1.52 ± 0.12abc 1627 2.62 ± 0.07a 3.72 ± 0.25a 2.04 ± 0.11ab 1.68 ± 0.08ab 2025 1.83 ± 0.04d 2.02 ± 0.16e 1.41 ± 0.07e 1.29 ± 0.04cd 1725 2.04 ± 0.09bcd 2.85 ± 0.25bcd 1.73 ± 0.08bcd 1.41 ± 0.03cd 1716 2.42 ± 0.07ab 2.24 ± 0.11de 1.46 ± 0.06de 1.24 ± 0.05d 1722 2.44 ± 0.12ab 2.74 ± 0.22cd 1.45 ± 0.09de 1.22 ± 0.06d I-107 2.23 ± 0.20abcd 3.42 ± 0.22ab 1.93 ± 0.13ab 1.76 ± 0.14a 1723 1.99 ± 0.11cd 2.41 ± 0.16de 1.74 ± 0.04bcd 1.32 ± 0.07cd 1640 2.04 ± 0.13bcd 3.36 ± 0.23abc 1.55 ± 0.07cde 1.30 ± 0.07cd 2.3 干旱胁迫对黑杨派无性系细胞膜透性的影响
由表4可知,各供试无性系在不同干旱处理条件下,细胞膜透性变化差别明显。随着干旱胁迫的加剧,10个无性系的细胞膜透性呈明显的升高趋势,且在重度干旱时达到最大值。其中无性系1716和1722升高最明显,与正常供水时相比均升高117.0%以上,增幅最小的是无性系1733和1641,都在94.0%以下,说明干旱胁迫对无性系1733和1641的影响较小,对1716和1722的影响较大。
表 4 土壤干旱对各无性系细胞膜透性的影响Table 4. Effects of soil drought on membrane permeability of each clone无性系
Clone细胞膜透性 Membrane permeability/% 正常供水
Normal water
supply轻度干旱
Mild
drought中度干旱
Moderate
drought重度干旱
Severe
drought1641 12.5 ± 1.1bc 13.4 ± 0.4b 17.3 ± 1.3cd 24.2 ± 1.1cd 1733 11.1 ± 0.6c 13.5 ± 0.4b 16.9 ± 0.7cd 21.2 ± 1.8d 1627 11.2 ± 1.0c 13.1 ± 1.0b 15.9 ± 1.3d 22.1 ± 1.8d 2025 12.3 ± 0.7bc 14.1 ± 1.0b 18.6 ± 1.8bcd 24.9 ± 2.9bcd 1725 14.7 ± 0.9ab 15.5 ± 0.4ab 20.5 ± 2.9bcd 29.8 ± 2.8abc 1716 11.3 ± 0.4c 14.5 ± 0.2b 21.3 ± 1.1abc 33.1 ± 2.4a 1722 14.5 ± 1.2ab 17.6 ± 0.7a 25.7 ± 1.0a 31.5 ± 1.4ab I-107 12.8 ± 0.3bc 14.4 ± 1.3b 16.9 ± 0.5cd 24.9 ± 2.1bcd 1723 16.6 ± 0.6a 17.7 ± 1.2a 23.2 ± 1.7ab 33.0 ± 1.6a 1640 12.1 ± 0.9bc 14.3 ± 0.7b 19.1 ± 1.2bcd 25.2 ± 2.9bcd 2.4 干旱胁迫对黑杨派无性系MDA含量的影响
表5显示:随干旱胁迫程度的增加,10个无性系的叶片MDA含量表现出先升高后下降的变化规律,在中度干旱时达到最高水平,变幅在3.40 ~ 4.85 mmol/kg之间,其中MDA含量最高的是无性系1716,达到4.85 mmol/kg;MDA含量最低的是无性系1733,为3.40 mmol/kg。在重度干旱时,MDA含量下降,此时无性系1641和1627的MDA含量最低,显著低于对照2025。
表 5 土壤干旱对各无性系MDA含量的影响Table 5. Effects of soil drought on MDA content of each clone无性系
CloneMDA含量 MDA content/(mmol∙kg−1) 正常供水
Normal water
supply轻度干旱
Mild
drought中度干旱
Moderate
drought重度干旱
Severe
drought1641 1.87 ± 0.26a 2.36 ± 0.20b 3.79 ± 0.14b 1.11 ± 0.23e 1733 2.31 ± 0.12a 2.87 ± 0.22ab 3.40 ± 0.37b 1.65 ± 0.14bcde 1627 2.46 ± 0.21a 3.03 ± 0.36ab 3.50 ± 0.41b 1.28 ± 0.14de 2025 2.27 ± 0.13a 2.99 ± 0.27ab 4.12 ± 0.15ab 2.69 ± 0.23a 1725 2.31 ± 0.26a 3.47 ± 0.34a 4.24 ± 0.27ab 1.44 ± 0.15cde 1716 2.22 ± 0.16a 2.94 ± 0.22ab 4.85 ± 0.24a 1.74 ± 0.27bcde 1722 2.21 ± 0.13a 2.78 ± 0.35ab 4.65 ± 0.19a 2.18 ± 0.12ab I-107 2.16 ± 0.22a 3.04 ± 0.20ab 3.75 ± 0.27b 1.80 ± 0.13bcd 1723 1.86 ± 0.23a 2.54 ± 0.21b 4.21 ± 0.21ab 1.60 ± 0.35bcde 1640 2.13 ± 0.30a 2.73 ± 0.27ab 3.76 ± 0.14b 2.07 ± 0.16abc 2.5 干旱胁迫对黑杨派无性系SOD活性的影响
表6显示:在整个干旱胁迫过程中,各供试无性系的SOD活性表现出先升后降的变化规律。在干旱胁迫初期,植物体内会产生SOD来抵御干旱胁迫造成的伤害,所有无性系的SOD活性在中度干旱时最高,与正常处理相比,升高幅度较大的无性系是1627(28.8%)和1733(28.4%),升高幅度较小的是1725(12.8%)和1722(12.5%)。随干旱时间增加,保护酶活性下降,在重度干旱胁迫时降至最低,该处理下无性系1627和1733的SOD活性较高,无性系1716和1722的SOD活性较低,说明无性系1627的1733适应重度干旱环境的能力较强。
2.6 干旱胁迫对黑杨派无性系POD活性的影响
由表7可知:随干旱程度的加强,10个黑杨派无性系的POD活性先增大后减小,除无性系1716在轻度干旱POD活性达到峰值以外,其他无性系的POD活性均在中度干旱达到最高值,之后随着胁迫处理程度增加,在重度干旱时土壤水分严重匮乏,POD活性降低。中度干旱时,无性系1733和1641的POD活性升高幅度较大,增幅在265.0%以上;在重度干旱时,相比其他无性系,1733和1627的POD活性维持在较高水平,与对照2025和I-107差异显著。
表 6 土壤干旱对各无性系SOD活性的影响Table 6. Effects of soil drought on SOD activity of each clone无性系
CloneSOD活性 SOD activity/(U∙g−1) 正常供水
Normal water supply轻度干旱
Mild drought中度干旱
Moderate drought重度干旱
Severe drought1641 642.5 ± 32.8abc 728.5 ± 48.6ab 783.1 ± 20.5bcd 526.1 ± 25.5de 1733 619.4 ± 22.0bc 687.8 ± 16.6b 795.0 ± 11.5abc 667.5 ± 25.2ab 1627 637.3 ± 17.9abc 792.1 ± 11.4a 821.1 ± 23.7ab 700.7 ± 25.5a 2025 584.0 ± 18.7c 678.1 ± 13.3b 746.8 ± 8.1cd 603.5 ± 22.0abcd 1725 658.0 ± 27.0ab 736.3 ± 25.2ab 742.2 ± 31.4cd 547.4 ± 25.3cde 1716 684.4 ± 16.5ab 746.8 ± 9.4ab 795.4 ± 19.0abc 456.7 ± 15.2e 1722 645.9 ± 17.7abc 695.3 ± 12.7b 726.5 ± 20.8d 522.6 ± 39.0de I-107 619.0 ± 19.7bc 706.2 ± 12.9b 781.1 ± 22.2bcd 593.9 ± 14.3bcd 1723 703.0 ± 8.6a 785.9 ± 17.7a 855.3 ± 18.0a 605.1 ± 58.1abcd 1640 647.4 ± 4.9abc 682.0 ± 24.2b 777.5 ± 14.4bcd 629.8 ± 30.4abc 表 7 土壤干旱对各无性系POD活性的影响Table 7. Effects of soil drought on POD activity of each clone无性系
ClonePOD活性 POD activity/(U∙g−1) 正常供水
Normal water
supply轻度干旱
Mild
drought中度干旱
Moderate
drought重度干旱
Severe
drought1641 2 808 ± 197ab 6 108 ± 379b 10 275 ± 1032ab 4 633 ± 240bc 1733 2 358 ± 140ab 3 575 ± 388d 9 625 ± 516abc 5 075 ± 123ab 1627 3 392 ± 508a 7 692 ± 251a 11 400 ± 353a 5 433 ± 412a 2025 2 125 ± 205b 3 800 ± 486cd 6 617 ± 262ef 2 658 ± 183fg 1725 2 283 ± 243ab 3 258 ± 271d 7 142 ± 226def 3 075 ± 113ef 1716 3 417 ± 726a 8 092 ± 246a 4 092 ± 435g 2 183 ± 210g 1722 2 325 ± 194ab 4 983 ± 446bc 8 058 ± 130cde 3 567 ± 466de I-107 2 733 ± 269ab 5 792 ± 180b 9 225 ± 1318bc 4 192 ± 235cd 1723 2 325 ± 166ab 3 842 ± 718cd 5 825 ± 175fg 2 950 ± 181efg 1640 2 550 ± 238ab 3 917 ± 171cd 8 608 ± 484bcd 3 075 ± 101ef 2.7 干旱胁迫对黑杨派无性系游离脯氨酸含量的影响
从表8可以看出,各供试无性系叶片的游离脯氨酸含量随控水时间的延长而不断增加。10个无性系叶片的游离脯氨酸含量在中度干旱时明显上升,变幅在24.6 ~ 40.7 mg/g之间,该水平下游离脯氨酸含量最高的是无性系1733,达到40.7 mg/g;最低的是无性系1716,为24.6 mg/g,说明各无性系对干旱胁迫比较敏感。随着土壤含水量的不断降低,植物体内的游离脯氨酸含量持续升高,渗透调节能力逐渐增强。在重度干旱下,游离脯氨酸含量及升高幅度最大的均是无性系1627和1733,与对照2025呈显著性差异。
表 8 土壤干旱对各无性系游离脯氨酸含量的影响Table 8. Effects of soil drought on free proline content of each clone无性系
Clone游离脯氨酸含量 Free proline content/(mg·g−1) 正常供水
Normal water
supply轻度干旱
Mild
drought中度干旱
Moderate
drought重度干旱
Severe
drought1641 19.2 ± 0.6a 24.7 ± 0.7ab 32.0 ± 2.1abc 40.6 ± 2.7cd 1733 18.7 ± 1.6ab 26.0 ± 2.3ab 40.7 ± 3.4a 52.5 ± 4.9a 1627 15.0 ± 1.2cd 28.9 ± 0.7a 38.0 ± 5.4a 52.1 ± 3.4a 2025 20.0 ± 0.9a 23.8 ± 0.8abc 30.8 ± 2.7abc 35.5 ± 2.7cde 1725 18.1 ± 1.1abc 22.9 ± 1.4bcd 31.1 ± 2.7abc 42.1 ± 4.1bc 1716 13.4 ± 1.2d 18.3 ± 1.2cd 24.6 ± 1.3c 31.1 ± 2.5de 1722 14.2 ± 0.3d 17.2 ± 0.8d 24.9 ± 2.9c 28.8 ± 1.7e I-107 18.9 ± 1.2ab 25.2 ± 2.6ab 36.5 ± 3.8ab 50.9 ± 2.7ab 1723 15.5 ± 0.8bcd 22.0 ± 1.5bcd 25.7 ± 1.8c 35.2 ± 3.1cde 1640 15.1 ± 1.2cd 25.2 ± 3.3ab 28.1 ± 1.1bc 39.1 ± 1.0cd 2.8 相关性分析
对重度干旱胁迫下10个黑杨派无性系的各项指标进行相关性分析,根据表9可知:其苗高增量与地径增量、生物量增量和游离脯氨酸含量呈极显著正相关,与叶绿素含量、POD活性呈显著正相关;地径增量与生物量增量、POD活性和游离脯氨酸含量呈极显著正相关,与叶绿素含量呈显著正相关;生物量增量与POD活性、游离脯氨酸含量呈极显著正相关,与叶绿素含量呈显著正相关;叶绿素含量与游离脯氨酸含量呈极显著正相关,与POD活性呈显著正相关,与细胞膜透性呈显著负相关;细胞膜透性与游离脯氨酸含量呈极显著负相关,与SOD活性和POD活性呈显著负相关;SOD活性与游离脯氨酸含量呈显著正相关;POD活性与游离脯氨酸含量呈极显著正相关。
表 9 干旱胁迫下无性系各指标间相关性分析Table 9. Correlation analysis among indicators of clones under drought stress指标 Index X1 X2 X3 X4 X5 X6 X7 X8 X9 X1 1 X2 0.880** 1 X3 0.793** 0.906** 1 X4 0.695* 0.755* 0.757* 1 X5 −0.615 −0.627 −0.585 −0.651* 1 X6 −0.617 −0.541 −0.547 −0.482 0.116 1 X7 0.518 0.586 0.441 0.530 −0.704* −0.033 1 X8 0.634* 0.782** 0.791** 0.757* −0.752* −0.537 0.593 1 X9 0.857** 0.882** 0.807** 0.913** −0.789** −0.446 0.709* 0.788** 1 注:X1. 苗高增量;X2. 地径增量;X3. 生物量增量;X4. 叶绿素含量;X5. 细胞膜透性;X6. MDA含量;X7. SOD活性;X8. POD活性;X9. 游离脯氨酸含量。*表示在0.05水平上显著相关,**表示在0.01水平上极显著相关。Notes: X1, ramet height increment; X2, basal diameter increment; X3, biomass increment; X4,chlorophyll content; X5, membrane permeability; X6, MDA content; X7, SOD activity; X8, POD activity; X9, free proline content. * indicates significant correlation at the 0.05 level, ** indicates highly significant correlation at the 0.01 level. 根据相关性分析可知:各测定指标具有一定的相关性,大部分指标间存在着显著的正负相关关系,说明各指标间存在相同的或者相反的变化关系,表中指标所反映的信息有一定的重叠性,直接利用这些指标会比较片面,因此需要用综合评价方法对其进行评价分析。
2.9 主成分分析
对10个无性系在严重缺水条件下的生长及生理指标进行主成分分析,将多个相关指标转换为几个独立的综合指标,并对各个无性系进行综合评价。主成分分析结果如表10所示,第1、第2主成分的累积贡献率达83.962%,表明这2个主成分可以把原来9项指标83.962%的信息表达出来。由表10可知:第1主成分中地径增量、生物量增量、游离脯氨酸含量、苗高增量、POD活性、叶绿素含量的特征向量绝对值较大,且均为正值,说明第1主成分主要反映无性系的生长量、渗透调节物质和抗氧化酶信息;第2主成分中丙二醛含量、SOD活性、细胞膜透性的特征向量绝对值较大,其中除细胞膜透性为负值,其余均为正值,说明第2主成分主要反映膜脂过氧化、抗氧化酶信息。
表 10 干旱胁迫下各指标的主成分特征向量、特征值及累计贡献率Table 10. Principal component characteristic vector, characteristic value and cumulative contribution rate of each index under drought stress测定指标
Measurement index第1主成分
Principal
component 1第2主成分
Principal
component 2苗高增量 Ramet height increment 0.351 −0.169 地径增量 Basal diameter increment 0.371 −0.100 生物量增量 Biomass increment 0.354 −0.182 叶绿素 Chlorophyll 0.349 −0.017 细胞膜透性 Membrane permeability −0.313 −0.424 丙二醛 MDA −0.222 0.678 超氧化物岐化酶 SOD 0.271 0.528 过氧化物酶 POD 0.351 0.012 游离脯氨酸 Free proline 0.384 0.097 特征值 Characteristic value 6.335 1.221 累计贡献率 Cumulative contribution rate/% 70.393 83.962 根据主成分分析得到10个无性系的第1主成分值F1和第2主成分值F2,根据公式F = 0.703 93F1 + 0.135 69F2计算综合主成分值,并将其排序,对各供试无性系的抗旱能力进行综合评价比较。由表11可知:10个黑杨派无性系的抗旱能力由强到弱依次为1733、1627、I-107、1641、1640、1725、1723、2025、1716、1722,其中无性系1733和1627的抗旱性优于I-107。
表 11 干旱胁迫下10个无性系的综合主成分值Table 11. Intergrated principal component values of 10 clones under drought stress无性系
CloneF1 F2 F 排序
Ranking1641 0.936 −1.223 0.493 4 1733 3.904 0.302 2.789 1 1627 3.567 0.306 2.552 2 2025 −2.188 2.135 −1.250 8 1725 −0.580 −0.876 −0.527 6 1716 −2.545 −1.497 −1.995 9 1722 −3.092 0.202 −2.149 10 I-107 2.150 0.092 1.526 3 1723 −1.218 −0.596 −0.938 7 1640 −0.723 1.155 −0.352 5 注:F1和F2代表第1主成分值和第2主成分值,F代表综合主成分值。Notes: F1 and F2 represent the 1st and 2nd principal component values and F represents the composite principal component value. 3. 讨论与结论
外部环境和自我调控机制共同作用于植物的生长发育,两者相互促进,对植物的正常生命活动产生作用[12]。当植物受到干旱胁迫的影响时,植物的生长量和生物量分配发生变化,以适应干旱环境带来的伤害。在本试验中,不同程度的干旱胁迫对10个黑杨派无性系的生长产生抑制,并导致其叶绿素含量、细胞膜透性、抗氧化酶活性、游离脯氨酸含量等生理生化指标发生不同程度的变化,说明不同无性系对干旱环境的适应能力也不同。重度干旱胁迫下各无性系的苗高增量、地径增量和生物量增量均有所不同,其中无性系1733和1627的生物量增量较大,无性系2025和1722的生物量增量较小。
干旱会使植物叶片受到损伤,影响植物正常进行光合作用。研究表明,植物在受到干旱胁迫时,叶绿素含量呈下降趋势[13−14]。在本试验中,随着干旱胁迫时间的延长,各无性系的叶绿素含量在轻度干旱下较对照轻微增加,这可能是因为植物为保持光合作用的稳定和适应干旱环境,自我调节增加了体内叶绿素含量,也有可能是由于干旱导致了植株的叶片发育受到阻碍,造成叶片水分流失,从而叶绿素含量升高;而在干旱胁迫后期土壤水分降低,叶绿素含量下降,表明干旱胁迫对10个黑杨派无性系叶片的叶绿素合成具有明显的抑制作用,同时原有的叶绿素会加速分解,导致光合作用能力降低,进而影响其生长量和生物量积累。这与之前对小叶杨(P. simonii)无性系[15]、白杨派(Sect. Leuce)无性系[16]的研究中得出的结果相同。
干旱同样会危害植物的细胞膜系统,发生膜脂过氧化。植物细胞膜在正常供水条件下选择透过性比较稳定,当土壤水分匮乏,细胞膜受到损伤,透性增大。持续干旱胁迫下,10个黑杨派无性系叶片的细胞膜透性呈增加的变化趋势,这与邱兴等[17]对4个1年生杨树无性系的研究结果基本一致,说明干旱胁迫对各无性系的植株幼苗细胞膜系统均造成了破坏。但不同无性系的升高幅度不一致,表明干旱条件下植物细胞膜受到的损伤程度不同。重度干旱时,无性系1716叶片的细胞膜透性增加幅度最大,说明其受到的损伤最为严重;无性系1733叶片的细胞膜相对透性增加幅度最小,说明其受到的伤害最小,对干旱环境的更有耐受性。膜脂过氧化的最终产物丙二醛,会导致生物膜的结构和功能发生改变,对植物产生毒害作用,其含量的高低可以反映膜脂过氧化作用的强弱。前人[18]对美洲黑杨的研究结果表明:不同美洲黑杨在遇到干旱胁迫后,其MDA含量存在着较大的差异。在本试验中,10个无性系在受到干旱胁迫时,MDA含量大幅增加,这说明土壤水分亏缺导致细胞膜脂过氧化作用逐渐加强,细胞膜受到伤害,胞内电解质外溢。在干旱胁迫后期,MDA含量达到最高值后不再增加,并随着水分匮乏严重,其含量开始降低,这可能是细胞代谢紊乱所致[19]。从MDA的含量来看,无性系1716和1722受到干旱胁迫的膜伤害更早出现,而无性系1733和1627的细胞膜相对受害程度较轻,其抗旱能力较强。这与鲁俊倩等[20]对‘84K’杨(P. alba × P. glandulosa ‘84K’)的研究结果相同。
干旱能够促进植物增强抗氧化酶的活性来缓解氧化损伤[21−22],提高保护能力。SOD和POD是植物体内重要的保护酶,逆境条件下,SOD和POD活性增强可以提高其抗氧化能力,降低胁迫对膜系统造成的损伤。相关研究发现,在不同程度的干旱胁迫下,杨树苗叶片的SOD和POD活性呈先升高后降低的趋势[23]。在本试验中,干旱胁迫前期SOD、POD活性较对照明显上升,说明一定程度的干旱胁迫会加快植物体内抗氧化酶的积累以适应干旱环境;随着干旱胁迫时间增加和干旱胁迫程度加重,这2种保护酶的活性均出现下降,说明黑杨派无性系对干旱胁迫的忍耐能力有一定的限度,当干旱程度超出自身的承受范围时,抗氧化酶活性降低,对活性氧和自由基的清除能力减弱,从而使膜系统受到破坏,对植物造成伤害。这与对蔷薇(Rosa multiflora)[24]、柽柳(Tamarix amarisk)[25]及小青杨(P. pseudo-simonii)[26]等的研究结论相似。在重度干旱时,无性系1733和1627的SOD和POD活性相对较高,均排在前2位,可以更好地抵抗干旱胁迫,减少活性氧的积累,增强抗旱能力。
干旱还会引起植物体内渗透调节物质的含量变化,植物在缺水状态下能够通过积累大量有机物质和无机离子,以调节渗透压[27],提高抗旱能力,避免其遭受干旱胁迫引起的损伤[28−29]。在本试验中,10个黑杨派无性系叶片中游离游离脯氨酸含量均随干旱胁迫程度的加重而不断增加,并且高于对照,说明游离脯氨酸的积累与黑杨派无性系抵抗干旱胁迫的能力密切相关,这与李敏等[30]对3种杨树的研究结果一致。游离脯氨酸作为一种重要的渗透调节物质,可以在一定程度上保持细胞渗透压平衡,10个无性系的游离脯氨酸含量升高幅度各不一致,表明其在干旱环境下维持渗透压稳定的能力不同。在重度干旱时,游离脯氨酸含量增幅最大的无性系是1627和1733,说明这2个无性系在干旱条件下细胞维持渗透压能力较强,对干旱环境的适应性强。
植物的抗旱能力受多种因素的影响,且不同植物面对干旱胁迫的响应不同,为应对干旱胁迫的伤害,黑杨派无性系启动自身的调节机制,体内抗氧化酶活性和渗透调节物质含量产生变化。本试验结果表明:在重度干旱胁迫下,10个无性系的各项指标与抗旱性存在不同相关性,其中细胞膜透性和MDA含量与抗旱性呈负相关,其余指标与抗旱性呈正相关。运用主成分分析法对各无性系进行综合评价,可以较为全面地评价干旱胁迫对无性系的影响,本研究初步认为无性系1733和1627具有较强的抗旱性,这2个无性系均属于欧美杨,另外3个欧美杨无性系I-107、1641和1640均排在前5位,5个美洲黑杨无性系1725、1723、2025、1716和1722均排在后5位,由此推测欧美杨的抗旱能力强于美洲黑杨。本试验采用盆栽模拟干旱试验,对10个黑杨派无性系的抗旱性常用指标进行测试和分析,并对其抗旱能力强弱进行综合评价,筛选出2个抗旱能力较强的欧美杨无性系,该试验结果为干旱立地杨树新品种选育和推广提供了重要参考依据,对林业生产具有重要指导意义。后期将对这些无性系进行田间抗旱能力测定,进一步验证其抗旱能力。
-
![]()
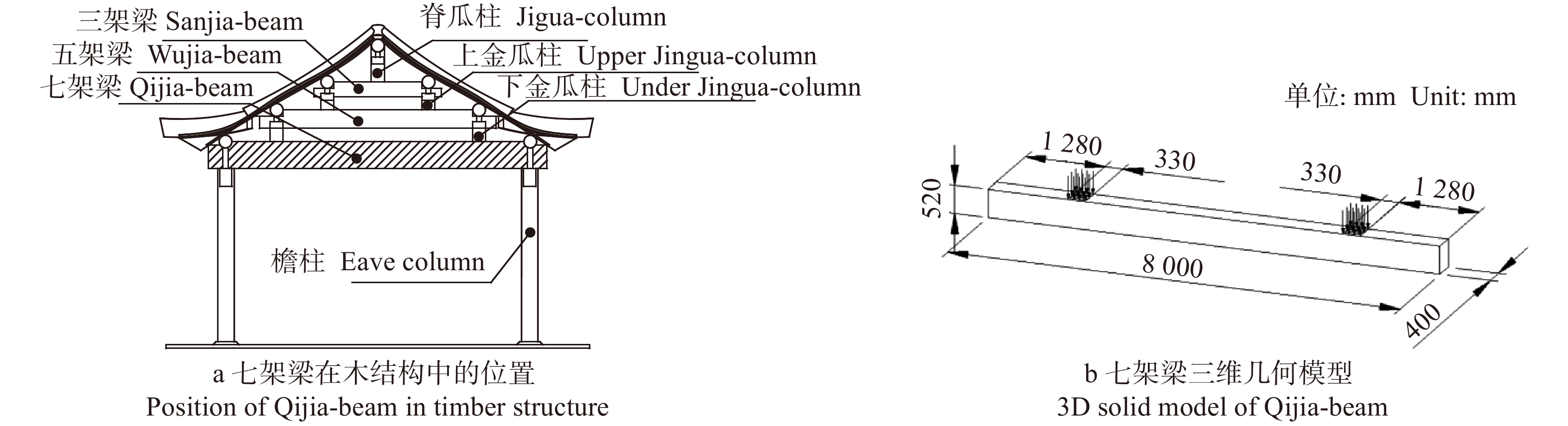
图 3 七架梁裂纹缺陷几何模型示意图
l1. 缝长Crack length; w1. 缝宽Crack width; d1. 缝深Crack depth
Figure 3. Sketch map of geometric model for Qijia-beam specimens with crack defect
![]()
图 4 七架梁外部腐朽截面示意图
d2. 腐朽深度Decay depth
Figure 4. Section diagram of Qijia-beam specimens with external decay defect
![]()
图 5 七架梁内部腐朽和空洞缺陷示意图
l3. 空洞长度Hole length; h3. 空洞高度Hole height; D. 空洞直径Hole diameter
Figure 5. Diagram of Qijia-beam specimens with internal decay and hole defect
![]()
图 6 七架梁荷载与边界约束示意图
UL为木梁纵向位移,UR为径向位移,UT为弦向位移。UL is the longitudial displacement, and UR is the radial displacement, UT is the tangential displacement.
Figure 6. Sketch map of load and boundary constraint for Qijia-beam
![]()
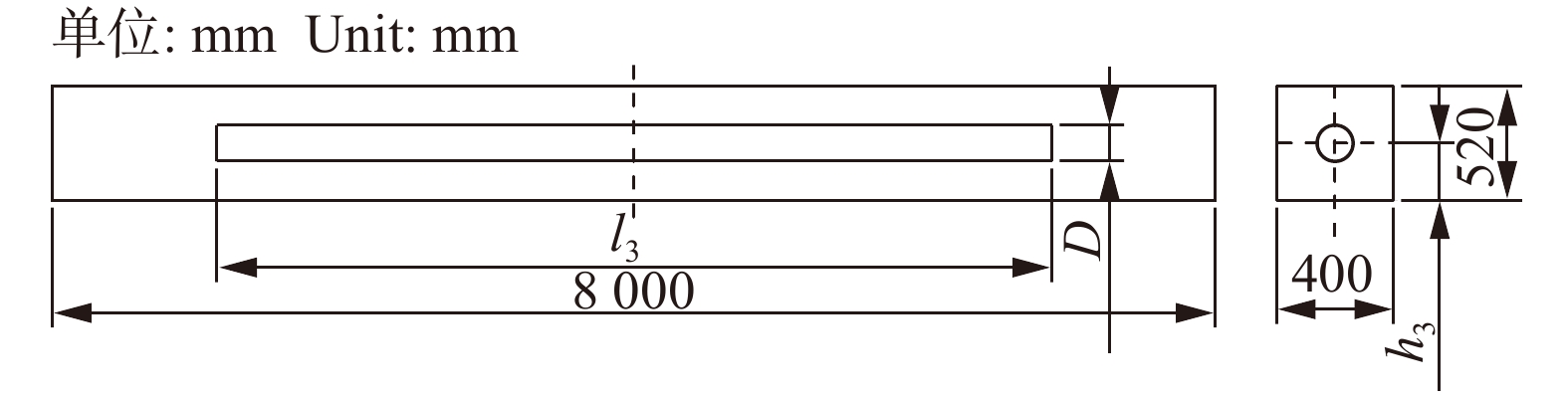
图 8 完好七架梁线弹性应力计算结果
Figure 8. Calculation results of linear elastic of healthy Qijia-beam
![]()
图 9 七架梁BC1-1线弹性应力计算结果
Figure 9. Calculation results of linear elastic of Qijia-beam BC1-1
![]()
图 10 不同开裂深度d1七架梁线弹性应力计算结果(l1 = 8 000 mm、w1 = 10 mm、h1 = 260 mm)
Figure 10. Calculation results of linear elastic stress of wood Qijia-beam with different d1 (l1 = 8 000 mm,w1 = 10 mm,h1 = 260 mm)
![]()
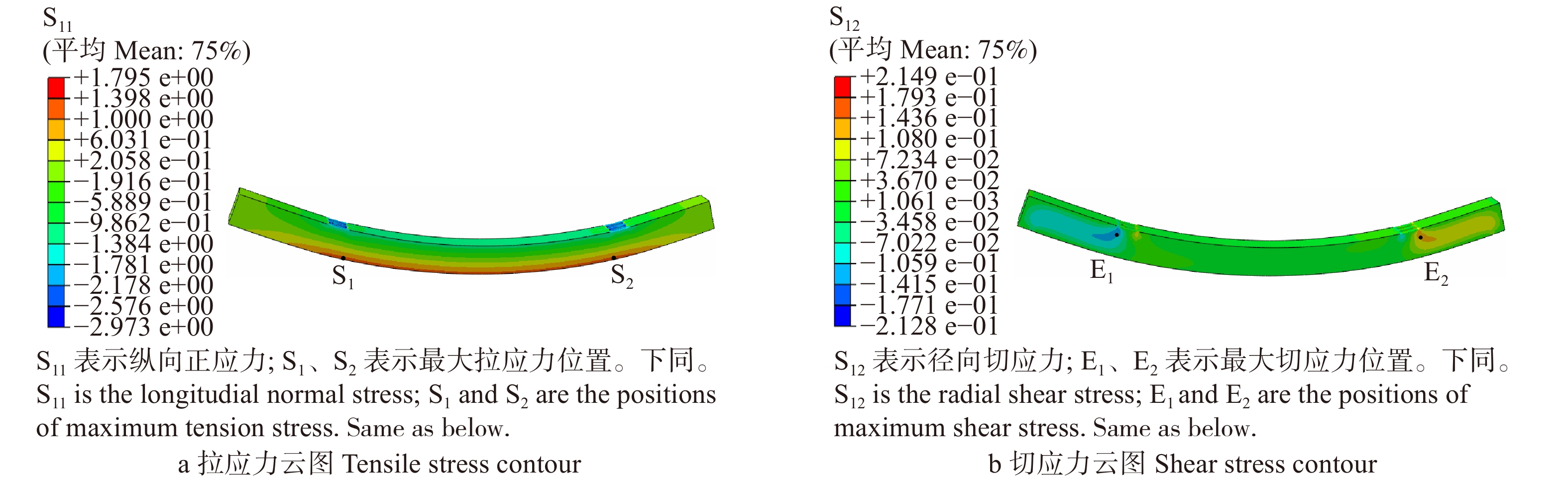
图 11 七架梁BD1-4线弹性应力计算结果
Figure 11. Calculation results of linear elastic stress of Qijia-beam BD1-4
![]()
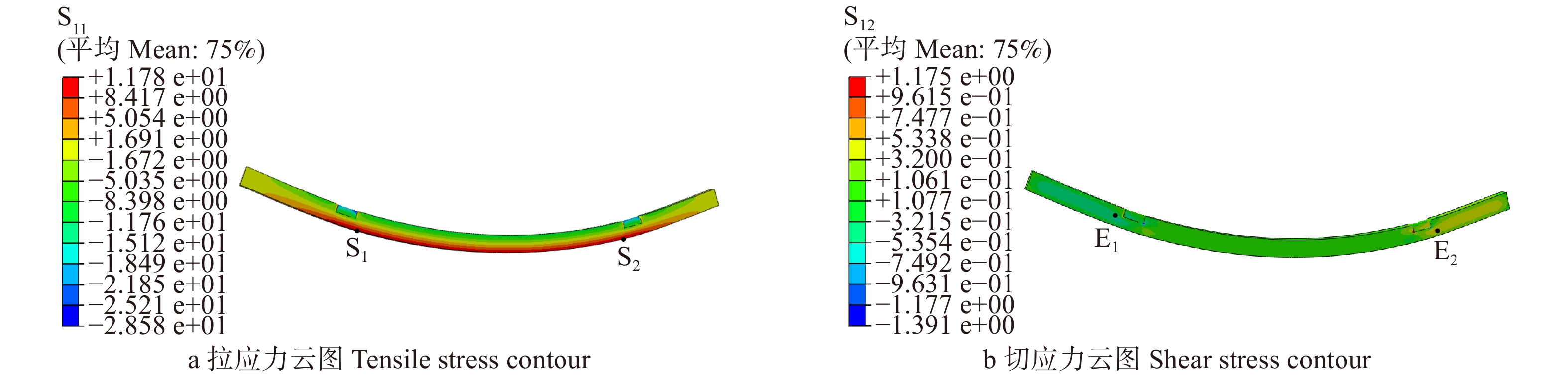
图 12 不同腐朽深度d2七架梁线弹性应力计算结果
Figure 12. Calculation results of linear elastic stress of Qijia-beam with different d2
![]()
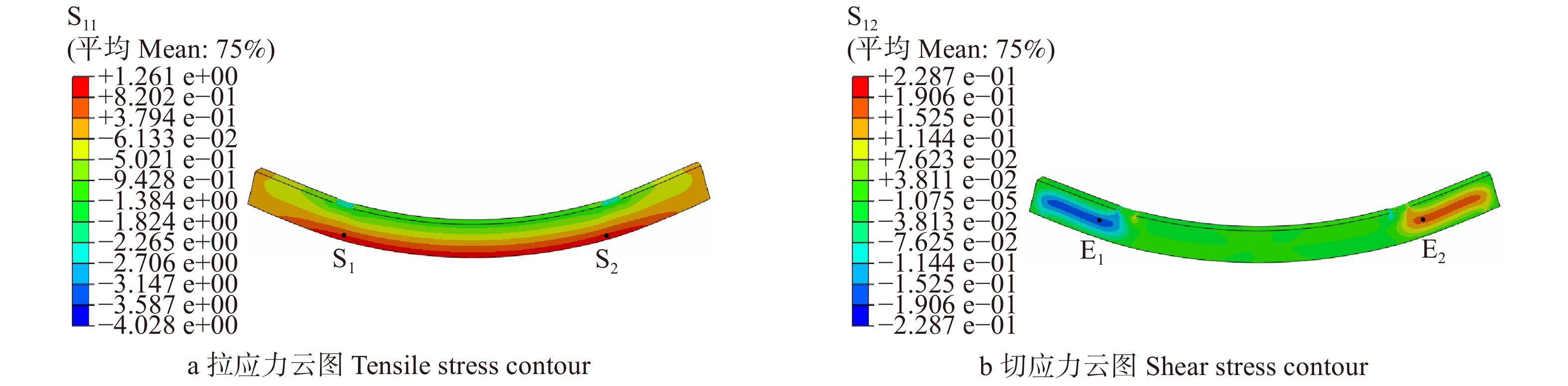
图 13 七架梁BH1-4线弹性应力计算结果
Figure 13. Calculation results of linear elastic stress of Qijia-beam BH1-4
![]()
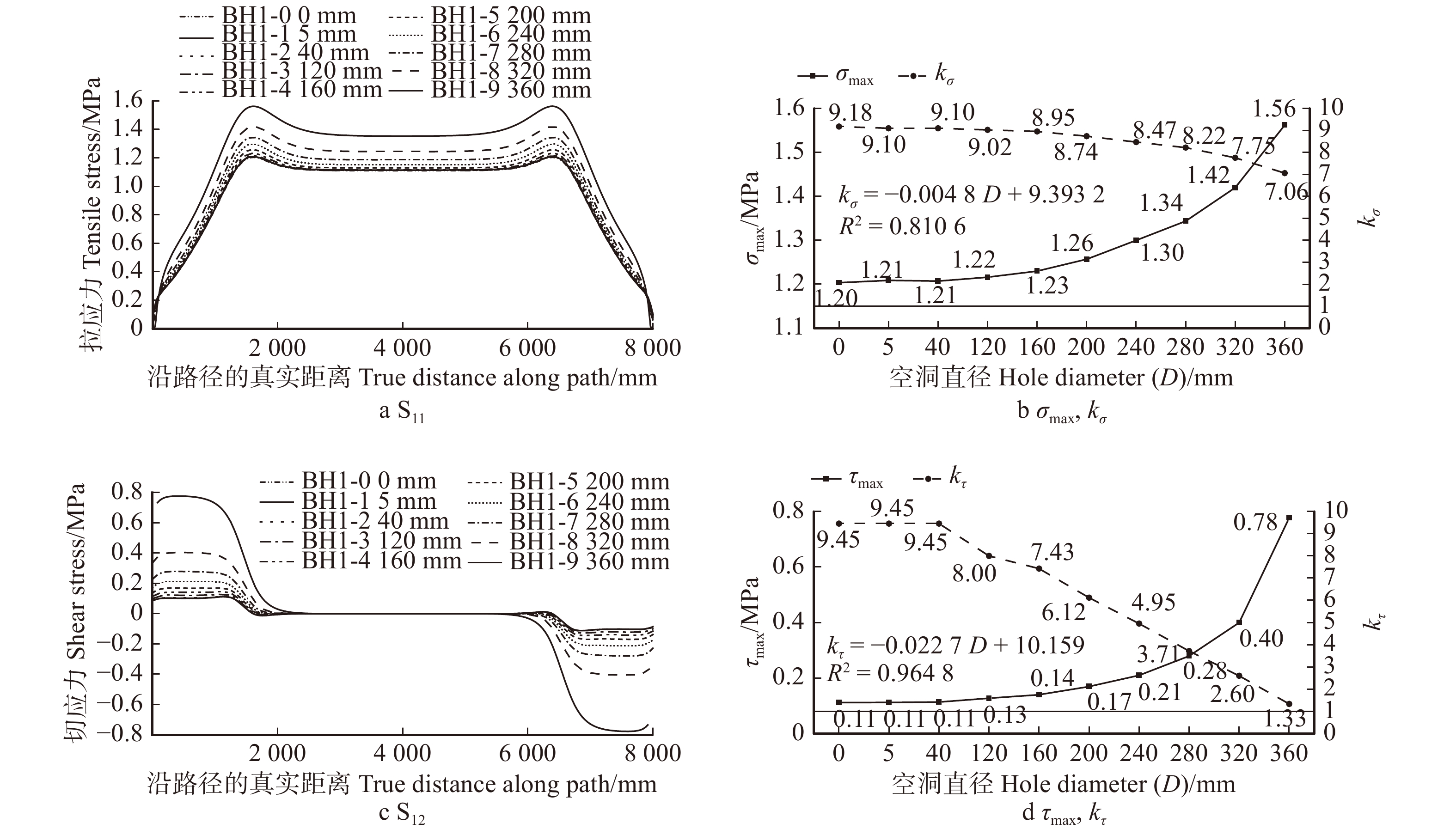
图 14 不同空洞直径D七架梁线弹性应力计算结果(h3 = 260 mm,l3 = 8 000 mm)
Figure 14. Calculation results of linear elastic stress of Qijia-beam with different D (h3 = 260 mm,l3 = 8 000 mm)
![]()
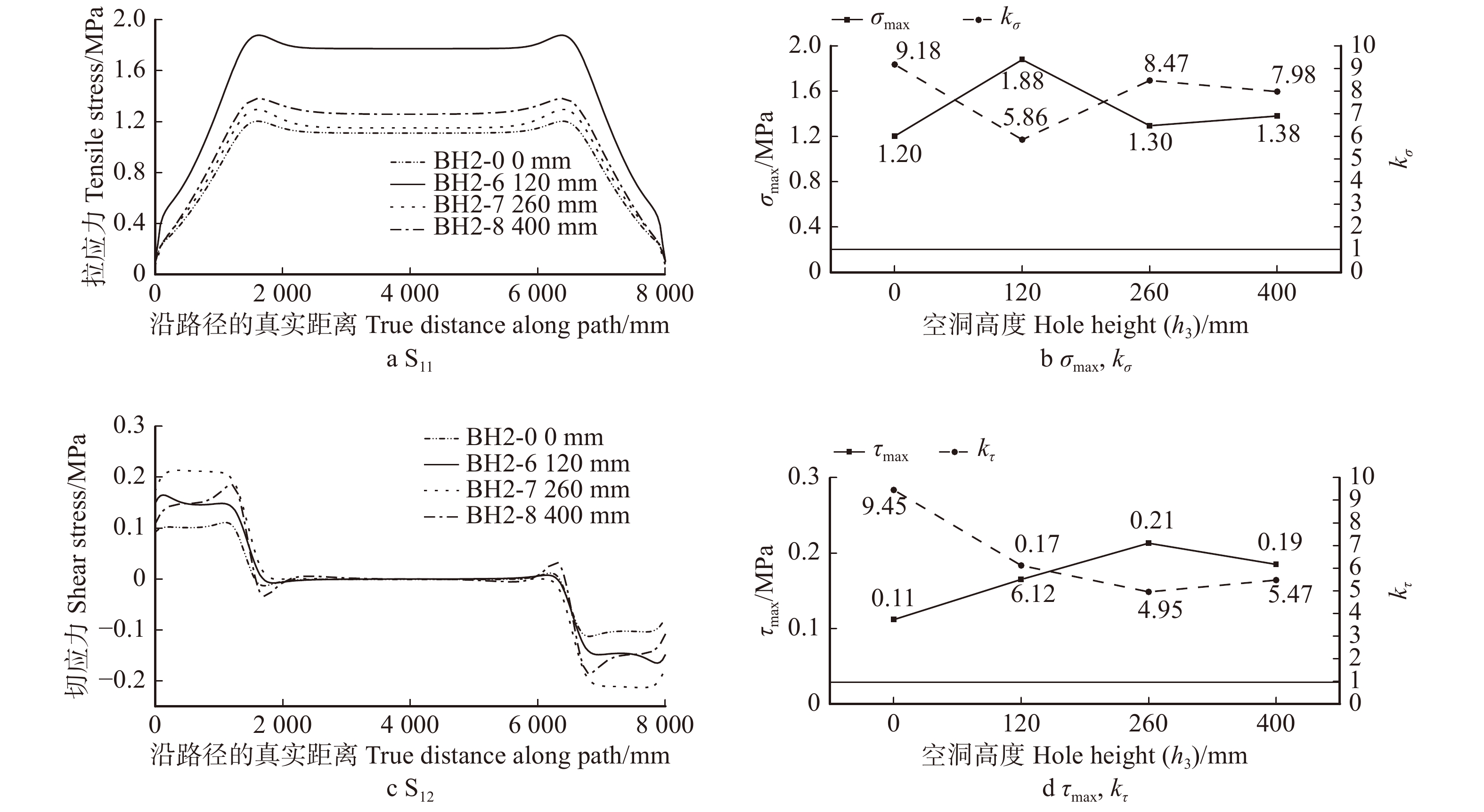
图 15 不同空洞高度h3七架梁线弹性应力计算结果(D = 40 mm,l3 = 8 000 mm)
Figure 15. Calculation results of linear elastic stress of Qijia-beam with different h3 (D=40 mm, l3 = 8 000 mm)
![]()
图 16 不同空洞高度h3七架梁线弹性应力计算结果(D = 240 mm,l3 = 8 000 mm)
Figure 16. Calculation results of linear elastic stress of Qijia-beam with different h3 (D = 240 mm, l3 = 8 000 mm)
![]()
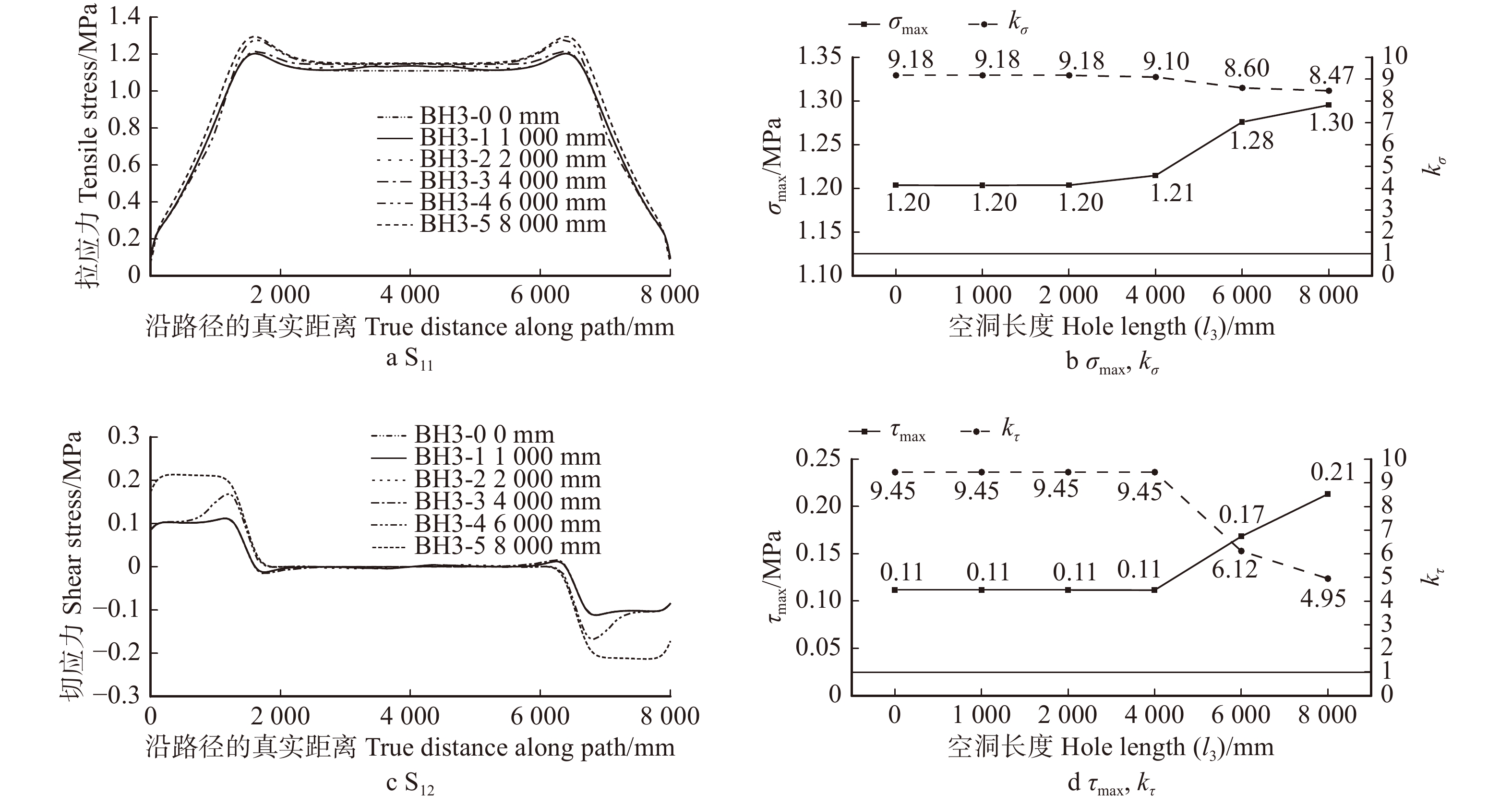
图 17 不同空洞长度l3七架梁线弹性应力计算结果(D = 240 mm,h3 = 260 mm)
Figure 17. Calculation results of linear elastic stress of Qijia-beam with different l3 (D = 240 mm, h3 = 260 mm)
![]()
图 18 某古建筑木结构七架梁线弹性应力计算结果
Figure 18. Calculation results of linear elastic stress of Qijia-beam for an ancient timber builiding
表 1 七架梁裂纹缺陷几何模型参数
Table 1 Modeling parameters of Qijia-beam specimens with crack defect
mm 模型编号
Model No.缝长
Crack
length (l1)缝宽
Crack
width (w1)缝深
Crack
depth (d1)缝高
Crack
height (h1)BC1-0 0 0 0 0 BC1-1 8 000 10 50 260 BC1-2 8 000 10 100 260 BC1-3 8 000 10 150 260 BC1-4 8 000 10 200 260 BC1-5 8 000 10 250 260 BC1-6 8 000 10 300 260 BC1-7 8 000 10 350 260  下载: 导出CSV
下载: 导出CSV
表 2 七架梁外部腐朽缺陷模型参数
Table 2 Modeling parameters of Qijia-beam specimens with external decay defect
mm 模型编号
Model No.腐朽深度
Decay
depth (d2)有效截面尺寸
Effective section size腐朽长度
Decay
length (l2)宽 Width (w2) 高 Height (h2) BD1-0 0 0 0 0 BD1-1 50 300 420 8 000 BD1-2 75 250 370 8 000 BD1-3 100 200 320 8 000 BD1-4 125 150 270 8 000 BD1-5 150 100 220 8 000
下载: 导出CSV
表 3 七架梁内部腐朽和空洞缺陷模型参数
Table 3 Modeling parameters of Qijia-beam specimens with internal decay and hole defect
mm 模型编号
Model No.空洞直径
Hole diameter (D)空洞高度
Hole height (h3)空洞长度
Hole length (l3)BH1-0 0 0 0 BH1-1 5 260 8 000 BH1-2 40 260 8 000 BH1-3 120 260 8 000 BH1-4 160 260 8 000 BH1-5 200 260 8 000 BH1-6 240 260 8 000 BH1-7 280 260 8 000 BH1-8 320 260 8 000 BH1-9 360 260 8 000 BH2-0 0 0 0 BH2-1 40 60 8 000 BH2-2 40 160 8 000 BH2-3 40 260 8 000 BH2-4 40 360 8 000 BH2-5 40 460 8 000 BH2-6 240 120 8 000 BH2-7 240 260 8 000 BH2-8 240 400 8 000 BH3-0 0 0 0 BH3-1 240 260 1 000 BH3-2 240 260 2 000 BH3-3 240 260 4 000 BH3-4 240 260 6 000 BH3-5 240 260 8 000
下载: 导出CSV
表 4 楠木和落叶松强度设计值
Table 4 Strength design value of Phoebe zhennan and Larix gmelinii
MPa 树种 Tree species 抗弯强度设计值 Designed bending strength (fm) 抗剪强度设计值 Designed shear strength (fv) 楠木Phoebe zhennan 7.13 0.84 落叶松Larix gmelinii 11.01 1.04 注:表4引自参考文献[13]。表中数据为正常条件下的木材强度设计值乘以不同使用条件和不同使用年限的调整系数所得。Notes:Tab. 4 is quoted from reference [13]. The data in the table are obtained by multiplying the wood designed strength under normal conditions by the adjustment factors of different used conditions and service life.
下载: 导出CSV
表 5 落叶松木材材性参数
Table 5 Property parameters of larch wood
EL/MPa ER/MPa ET/MPa μLR μLT μRT GLR/MPa GLT/MPa GRT/MPa 14 190 1 419 709.50 0.03 0.02 0.43 1 064.25 851.40 255.42 注:表5引自参考文献[2, 14]。EL为木梁纵向弹性模量,MPa; ER为径向弹性模量,MPa; ET为弦向弹性模量,MPa; μLR为LR面的泊松比; μLT为LT面的泊松比; μRT为RT面的泊松比; GLR为LR面内的剪切模量,Mpa;GLT为LT面内的剪切模量,Mpa;GRT为RT面内的剪切模量,Mpa。下同。Notes: Tab. 5 is quoted from reference [2, 14]. EL is the longitudinal elastic modulus of the wood beam, MPa; ER is the radial elastic modulus, MPa; ET is the tangential elastic modulus, MPa; μLR is the Poisson’s ratio of LR-plane; μLT is the Poisson’s ratio of LT-plane; μRT is the Poisson’s ratio of RT-plane; GLR is the shear modulus of LR-plane, Mpa; GLT is the shear modulus of LT-plane, Mpa; GRT is the shear modulus of RT-plane, Mpa. Same as below.
下载: 导出CSV
-
[1] 石四军. 古建筑营造技术细部图解[M]. 沈阳: 辽宁科学技术出版社, 2010. Shi S J. Ancient building construction technology detail[M]. Shenyang: Liaoning Science and Technology Publishing House, 2010.
[2] 成俊卿. 木材学[M]. 北京: 中国林业出版社, 1985. Cheng J Q. Wood science[M]. Beijing: China Forestry Publishing House, 1985.
[3] 廖春晖, 张厚江, 黎冬青, 等. 古建筑圆柱形木构件内部缺陷筛查方法研究[J]. 北京林业大学学报, 2013, 35(1):123−126. Liao C H, Zhang H J, Li D Q, et al. Screening method of internal defects in cylindrical wood members of ancient architectures[J]. Journal of Beijing Forestry University, 2013, 35(1): 123−126.
[4] 林丽. 古建筑木梁架常见破坏形式及修缮方案—以浙江古民居为例[J]. 价值工程, 2015, 34(29):155−157. Li L. The wooden beams renovation scheme of Zhejiang ancient architecture[J]. Value Engineering, 2015, 34(29): 155−157.
[5] 张厚江, 管成, 文剑. 木质材料无损检测的应用与研究进展[J]. 林业工程学报, 2016, 1(6):1−9. Zhang H J, Guan C, Wen J. Applications and research development of nondestructive testing of wood based materials[J]. Journal of Forestry Engineering, 2016, 1(6): 1−9.
[6] Akbiyik A, Lamanna A J, Hale W M. Feasibility investigation of the shear repair of timber stringers with horizontal splits[J]. Construction and Building Materials, 2007, 21(5): 991−1000. doi: 10.1016/j.conbuildmat.2006.03.004
[7] Nagai H, Murata K, Nakano T. Defect detection in lumber including knots using bending deflection curve: comparison between experimental analysis and finite element modeling[J]. Journal of Wood Science, 2009, 55(3): 169−174. doi: 10.1007/s10086-008-1016-y
[8] Xu H, Wang L. Curvature modal analysis of defective wood beams based on finite element method[C]// Changsha University of Science & Technology. International Conference on Measuring Technology & Mechatronics Automation. Washington: IEEE Computer Society, 2010: 874−877.
[9] Zhang J, Xu Q F, Xu Y X, et al. Research on residual bending capacities of used wood members based on the correlation between non-destructive testing results and the mechanical properties of wood[J]. Journal of Zhejiang University-Science A (Applied Physics & Engineering), 2015, 16(7): 541−550.
[10] 孙艳玲, 张青, 赵东. 木材裂纹尖端应力、应变场的数值分析[J]. 北京林业大学学报, 2010, 32(1):103−107. Sun Y L, Zhang Q, Zhao D. Stress and strain filed at wood crack[J]. Journal of Beijing Forestry University, 2010, 32(1): 103−107.
[11] 温宇鑫, 赵健, 赵东. 孔洞对木梁弯曲应变分布影响的试验研究[J]. 北京林业大学学报, 2017, 39(11):106−113. Wen Y X, Zhao J, Zhao D. Experimental study on the effects of holes on bending strain distribution of wood beams[J]. Journal of Beijing Forestry University, 2017, 39(11): 106−113.
[12] 周乾, 闫维明, 纪金豹. 古建残损木梁受弯性能数值模拟研究[J]. 山东建筑大学学报, 2012, 27(6):570−574. doi: 10.3969/j.issn.1673-7644.2012.06.006 Zhou Q, Yan W M, Ji J B. Numerical simulation of bending performance of the unsound Chinese ancient wooden beam[J]. Journal of Shandong Jianzhu University, 2012, 27(6): 570−574. doi: 10.3969/j.issn.1673-7644.2012.06.006
[13] 中华人民共和国建设部. 木结构设计规范: GB50005—2017[S]. 北京: 中国建筑工业出版社, 2017. Ministry of Development of the People’s Republic of China. Code for design of timber structures: GB50005−2017[S]. Beijing: China Architecture & Building Press, 2017.
[14] 龙卫国, 杨学兵. 木结构设计手册[M]. 北京: 中国建筑工业出版社, 2005. Long W G, Yang X B. Manual for design of timber structure[M]. Beijing: China Architecture & Building Press, 2005.
[15] 刘大可. 古建筑屋面荷载编汇(上)[J]. 古建园林技术, 2001(3):58−64. Liu D K. Assembly of roof loads in ancient buildings[J]. Traditional Chinese Architecture and Gardens, 2001(3): 58−64.
[16] 中华人民共和国建设部. 古建筑木结构维护与加固技术规范: GB50165—92[S]. 北京: 中国建筑工业出版社, 1992. Ministry of Development of the People’s Republic of China. Technical code for maintenance and strengthening of ancient timber buildings: GB50165−92[S]. Beijing: China Architecture & Building Press, 1992.
[17] 朱忠漫, 邱洪兴, 王靖翔. 干缩裂缝对木梁抗弯性能影响的试验研究[J]. 淮海工学院学报(自然科学版), 2015, 24(增刊1):74−79. Zhu Z M, Qiu H X, Wang J X. Flexural performance of timber beams with shrinkage crack[J]. Journal of Huaihai Institute of Technology (Natural Science Edition), 2015, 24(Suppl.1): 74−79.
-
期刊类型引用(12)
1. 包崇寅,孙永玉,李敏敏,邢洪铭,戚建华. 不同生境濒危植物龙棕种群结构及其动态特征. 西北植物学报. 2024(03): 479-490 .  百度学术
百度学术
2. 赵鑫,陈虹,赵善超,陈兵权,郭来珍,周昊亮. 新疆天山云杉种子雨时空动态分布及种子萌发特性. 浙江农林大学学报. 2024(03): 542-548 . 百度学术
3. 靳旭红,于聪,张庭耀,吕松瞳,刘扬,陈乐,龙生,穆怀志. 基于种子活力和苗期生长的枫桦半同胞家系初选. 植物研究. 2024(05): 763-773 . 百度学术
4. 刘志宇,张忠辉,杨凯麟,张军,姜润华,吴则甫,王琦,李文华,夏富才. 不同经营方式的云冷杉针阔混交林土壤真菌群落结构1). 东北林业大学学报. 2023(03): 124-129 . 百度学术
5. 强亚琪,范春雨,张春雨. 长白山暗针叶林群落物种多样性维持机制. 生态学报. 2023(05): 1884-1891 . 百度学术
6. 黄梓良,徐子恒,孙操稳. 青钱柳种子雨的季节动态及土壤种子库特征. 南京林业大学学报(自然科学版). 2023(02): 18-26 . 百度学术
7. 陈士刚,邹建军,山昌林,李秀红,芦静,王岗. 硕桦优树选择及不同种源、家系苗期评价. 吉林林业科技. 2023(02): 1-5+23 . 百度学术
8. 王莹,文淑均,罗定明,覃延闯,丁涛,刘世男. 珍稀濒危植物元宝山冷杉种实特征及种子萌发特性研究. 广西科学院学报. 2023(02): 161-168 . 百度学术
9. 李继祥,余登利,肖息,杨雪,田晓光,龚文斌,邓坦. 贵州宽阔水国家级自然保护区亮叶水青冈种子雨特征. 绿色科技. 2022(07): 67-69 . 百度学术
10. 李云红 ,田松岩 ,沃晓棠 ,邵英男 ,刘延坤 ,韩丽冬 ,陈瑶 ,刁云飞 ,刘玉龙 . 伴生东北红豆杉针阔混交林种子雨时空动态. 中南林业科技大学学报. 2022(05): 109-118 . 百度学术
11. 江海都,谢伟玲,柴胜丰,唐健民,蒋运生,秦惠珍,韦霄. 喀斯特地区珍贵树种黄枝油杉的种子萌发特性. 广西植物. 2022(06): 951-960 . 百度学术
12. 焦洁洁,李领寰,汪建民,孙杰杰,吴初平,姚良锦,王志高,袁位高. 午潮山杉阔混交林种子雨与土壤种子库特征研究. 林业资源管理. 2022(04): 28-35 . 百度学术
其他类型引用(2)









计量
- 文章访问数: 3213
- HTML全文浏览量: 1769
- PDF下载量: 84
- 被引次数: 14
