
Establishment of an effective protocol for cultivation of tissue cultured seedlings in Pinus massoniana superior provenance
-
摘要:目的马尾松是我国南方生态建设和造林用材的主要树种,受种苗良种化限制,人工林生产力水平整体不高,产业发展缓慢。因此,构建高效马尾松优良种质繁育技术体系对推动良种推广利用,加快产业发展,提升行业竞争力很有必要。方法以马尾松骨干育种资源桐棉种源为研究对象,未成熟球果合子胚为外植体,通过体胚发生技术获取成熟胚状体作为本试验供试材料,针对马尾松体胚萌发率低、芽苗活性差、生根成苗困难等技术瓶颈,系统分析了活性炭(AC)、基本培养基、植物激素对体胚萌发及萌发体胚芽苗复壮与不定根诱导的影响。结果(1)AC能显著提升成熟胚状体萌发培养效果,但过高浓度AC会导致培养效果减弱,其中以0.83 mol/L AC效果最佳。在附加AC基础上,利用高N、低NH4+/NO3− 比,K、Ca量适中的1/2MMS基本培养基能进一步提升萌发效果,萌发率可达94.1%。(2)0.42 mol/L AC能有效促进萌发体胚芽苗伸长,在4 μmol/L TDZ作用下,腋芽诱导效果好,有效芽增殖系数5.6/35 d,芽高9.2 cm/50 d。(3)经1.2 μmol/L NAA + 2 μmol/L PBZ处理60 d,单芽生根率达94.3%、根条数6.4,移栽3个月后成活率为95.8%。结论本研究首次通过体胚发生与器官发生途径相结合的技术构建了高效组培繁育体系,可用于马尾松优良种质的快速繁殖以及遗传转化方面的研究,为马尾松良种产业化以及进一步开展基因工程分子育种方面的研究奠定了坚实基础。Abstract:ObjectivePinus massoniana is a main tree species for ecological construction and timber in southern areas of China. However, the productivity of plantation is generally low resulting from the limitation of improved varieties, which leads to the slow development of industrialization in P. massoniana. It is necessary to develop an effective propagation system of elite germplasm for P. massoniana in order to promote the use of improved varieties, accelerate the industrial development, and enhance the competitiveness of industry.MethodIn this study, ‘Tongmiansong’ (TM), the backbone of P. massoniana breeding resources was used as the research object, zygotic embryos excised from immature cones were applied to explants, and mature somatic embryos (SE) obtained via somatic embryogenesis were used for testing materials. Concerning the technical bottlenecks of P. massoniana tissue culture, including low germination rate, poor shoot growth, and recalcitrance to rooting, effects of active charcoal (AC), basal media, and plant hormones on SE germination as well as reinvigoration and adventitious rooting of shoots were investigated in the present study.Result(1) AC significantly improved germination of TM mature SE, while a high level of AC was able to weaken their germination effects, and the best effect was observed at the 0.83 mol/L AC treatment. Based on the application of AC in the medium, the basal medium composed of high N, low ratio of NH4+/NO3− , and moderate K and Ca furtherly enhanced the germination of SE, reaching 94.1% of germinating rate. (2) 0.42 mol/L AC effectively promoted the elongation of germinated SE. Under the treatment of 4 μmol/L TDZ, induction of axillary buds was better, achieving effective bud proliferation coefficient of 5.6/35 d, and shoot height of 9.2 cm/50 d. (3) After 60 days of 1.2 μmol/L NAA + 2 μmol/L PBZ application in the rooting medium, rooting rate was 94.3%, root number was 6.4, and survival rate was 95.8% after 3-month transplanting.ConclusionThe effective breeding system by tissue culture for TM was firstly established via a combined approach of somatic embryogenesis and organogeneis in this study, which would be used for the rapid propagation of elite germplasm for P. massoniana as well as for the research on genetic transformation, providing solid foundation for industrialization of improved varieties and molecular breeding in P. massoniana.
-
Keywords:
- Pinus massoniana /
- tissue culture /
- somatic embryo /
- adventitious rooting /
- plant regeneration
-
赤霉素(GAs)是一种重要植物激素,在植物整个生命周期中对生长发育的各个阶段具有广泛的调控作用[1-2]。GAs通过促进细胞分裂和细胞伸长[3-6]进而促进茎部节间的伸长生长。李哲馨[7]的研究表明GAs在烟草茎部形成层发挥重要作用,对植株生长和木质部发育具有明显的促进作用。因此,GAs代谢调控在速生林林木育种中具有重要的应用潜力;然而,以往的研究都是基于施加外源活性GAs或全株组成型调控GAs含量的分析,并没有考虑活性GAs在活体内的运输特性对研究结果的影响,有研究表明移动的GAs对茎的次生生长与木质部形成层的发育具有直接的促进作用[8]。另外,由于GAs参与调控多个发育过程,特异下调茎部GAs含量在延缓茎部生长的同时,也会抑制不定根的发生[9],影响根尖的径向生长[10]。有研究表明茎部GAs在低温胁迫的响应过程中具有反馈调控作用[11-12],因此,开展对茎部特异性GAs含量的调控研究,为GAs代谢调控植物生长发育在林木育种中的实际应用提供理论基础。
形成层位于木质部和韧皮部之间,是一种重要的分生组织,细胞分裂旺盛,向外分裂形成新的韧皮部细胞,向茎的中轴方向分裂形成新的木质部细胞,大量调控木材合成的基因在此过程中表达[13]。研究发现在形成层细胞分裂时,GAs加快了形成层细胞的分裂速度,促进了细胞的伸长,从而促进导管及筛管的伸长[14]。此后的研究中进一步证实了形成层GAs对植株生长发育有促进作用,但在外源施加GAs时,尚不能排除由于植物组织损伤而造成植株自身激素含量变化的误差,并且植物某一发育时间或某一组织中的外源激素含量的测定也难以精准,给实验带来很多不确定性[15]。因此,特异性降低植物形成层GAs含量对植物生长发育的调控作用有待于进一步深入研究。
已有研究表明,在植物体内自由运输的可能并不是活性GAs,而是其前体GA12[16]。植物嫁接是研究活性物质长距离运输特性的有效手段[17]。本实验通过嫁接不同激素含量的转基因植株及野生型WT植株再配合外施赤霉素进行实验分析,验证活性GAs在烟草中的运输机制,更深入地探讨组织特异性GAs含量对植株生长发育的调控作用,对今后GAs调控植物各组织发育的研究具有重要意义,为林木速生优质新品种定向培育策略的制定提供理论基础。
1. 材料与方法
1.1 实验材料
以本实验室遗传转化获得的烟草转基因株系LMX5:: PtGA2ox1、35S:: PtGA20ox、35S:: PtGA2ox1及野生型烟草为研究材料(其中35S:: PtGA20ox、35S:: PtGA2ox1来自林木遗传育种实验室的保存材料),分别记作L:G2、35:G20、35:G2与WT。PtGA2ox1基因克隆于毛白杨,转基因受体为野生型烟草W38株系(Nicotiana tabacum cultivar Wisconsin 38)。所有植物材料均在不含任何激素的MS培养基上,组培室温度为25 ℃,光照周期为16 h光照/8 h黑暗,将继代培养2月苗龄的生根苗移栽到温室,在30 ℃自然光照下培养。
1.2 烟草形成层赤霉素特异性调控转基因株系的获得
用毛白杨的成熟叶片提取高质量RNA,反转录获得cDNA,再以cDNA为模板克隆赤霉素代谢中的关键基因PtGA2ox1,构建表达载体LMX5::PtGA2ox1,并转化表达载体到农杆菌中。通过农杆菌介导的转基因技术获得以木质部形成层特异性启动子(LMX5)启动的烟草转基因株系L:G2[18]。
1.3 基因表达分析及GAs含量的测定
分别提取转基因烟草L:G2、35:G2及WT植株茎部、叶片和根部组织的总RNA,反转录得到cDNA,根据差异片段序列设计引物,半定量RT-PCR反应条件:95 ℃变性3 min,随后进行以下40个循环:95 ℃变性3 s,60 ℃退火20 s,72 ℃延伸30 s。以Actin为内标基因,扩增引物序列为FW:5′-CACAAGCCAGCACTTCAACAG-3′,RV:5′-ATGCCTTAACCAGGAGGTGC-3′。将每个扩增反应做3次重复实验,每次均设置阴性对照,并利用软件Bio-Rad CFXManager 2.0读取相关数据。
分别取组织培养30 d的35:G20、35:G2、L:G2与WT植株的叶、茎、根3个部位的样品各3 g,将其放在研钵中进行研磨,用LC-MS(液相色谱−质谱联用仪)对样品进行测定分析[19],每个样本进行3次重复实验。
1.4 烟草的微嫁接
以35:G2烟草转基因株系的顶芽作接穗、35:G20植株茎杆的一段作砧木,从砧木的横截面中心向下垂直的劈开切接口,使劈口长度与接穗的削面相吻合,再用锡箔纸将接穗与砧木紧密包裹,得到以35:G2为接穗,35:G20为砧木的独立嫁接苗(以下写作为35:G2//35:G20),以同样方法分别获得嫁接苗35:G20//35:G2、35:G2//WT以及WT//WT。
1.5 植株各部分生长性状的测定
挑选培养45 d的无菌生根苗L:G2、35:G2、35:G20与WT,分别对其茎部进行切片,用0.5%番红染液染色后对木质部进行显微观察并拍照。
将烟草转基因株系L:G2、35:G2和35:G20的顶芽扦插在无激素的MS培养基上,设置WT为对照,培养20 d后对各组的生根情况进行观察,再从4组株系中各选取30株,对其不定根的长度进行测量,计算平均值后与对照组一同进行t检验。
2. 结果与分析
2.1 转基因株系的获得及基因表达分析
通过农杆菌介导的转基因技术获得了以木质部形成层特异性启动子LMX5介导的烟草转基因株系L:G2,提取植株的DNA进行PCR扩增反应进行初步鉴定,成功获得21个转基因株系,与野生型WT相比,转基因植株(图1A)生长矮小,不定根数目减少,差异明显。
![]() 图 1 不同烟草植株及基因表达水平A是不同烟草植株,标尺长度为1 cm;B是转基因株系各组织中PtGA2ox1基因的表达水平;C是转基因株系不同组织中烟草NtGA2ox1基因本底表达水平,将35:G2植株中的PtGA2ox1基因的表达量设置为1。A is different tobacco plant with a scale length of 1 cm; B is the expression level of PtGA2ox1 gene in each tissue of transgenic lines; C is the background expression level of tobacco NtGA2ox1 gene in different tissues of transgenic lines, which will be in 35:G2 plants, the expression level of the PtGA2ox1 gene was set to 1.Figure 1. Different tobacco plants and gene expression levels
图 1 不同烟草植株及基因表达水平A是不同烟草植株,标尺长度为1 cm;B是转基因株系各组织中PtGA2ox1基因的表达水平;C是转基因株系不同组织中烟草NtGA2ox1基因本底表达水平,将35:G2植株中的PtGA2ox1基因的表达量设置为1。A is different tobacco plant with a scale length of 1 cm; B is the expression level of PtGA2ox1 gene in each tissue of transgenic lines; C is the background expression level of tobacco NtGA2ox1 gene in different tissues of transgenic lines, which will be in 35:G2 plants, the expression level of the PtGA2ox1 gene was set to 1.Figure 1. Different tobacco plants and gene expression levels通过测定转基因烟草L:G2、35:G2外源基因的表达水平(图1B、1C),发现L:G2、35:G2转基因植株的表达模式不同,PtGA2ox1转录水平只在L:G2的茎部出现了明显的上升趋势,而叶片和根部的上升趋势不明显。通过测定转基因烟草的叶片、根部及茎部中NtGA2ox1基因的本底表达水平,发现转基因烟草株系L:G2在叶片和根部受自身调控的NtGA2ox1基因表达水平与野生型WT对照比并无明显变化,而茎部NtGA2ox1基因的表达水平和野生型WT相比却明显降低;同时发现转基因烟草株系35:G2各组织中的NtGA2ox1基因表达水平与WT相比均有明显的下降。
在烟草中,主要活性赤霉素GA1在L:G2的茎部含量明显低于野生型WT植株,而非活性的GA8在茎部的含量与WT相比有显著的提高(图2);而转基因株系35:G20与WT相比,根、茎以及叶片内活性GAs(主要是GA1)含量显著提高,即植株整体GAs含量上调(图2);转基因植株35:G2茎、叶部分的活性GAs含量均有显著的下调,而非活性GAs含量增多,植株整体GAs含量下调,可能是GAs含量下降负反馈调节导致活性GA2-氧化酶含量下降 (图2)。
![]() 图 2 转基因植株根部、茎部、叶片中GAs的含量A. 转基因植株根部GAs的含量;B. 转基因植株茎部GAs的含量;C. 转基因植株叶片中GAs的含量。A, content of GAs in roots of transgenic plants; B, content of GAs in the stem of transgenic plants; C, content of GAs in leaves of transgenic plants.Figure 2. GAs content in stems, roots and leaves of transgenic plants
图 2 转基因植株根部、茎部、叶片中GAs的含量A. 转基因植株根部GAs的含量;B. 转基因植株茎部GAs的含量;C. 转基因植株叶片中GAs的含量。A, content of GAs in roots of transgenic plants; B, content of GAs in the stem of transgenic plants; C, content of GAs in leaves of transgenic plants.Figure 2. GAs content in stems, roots and leaves of transgenic plants2.2 活性GAs在烟草茎中的运输特性
对35:G20//35:G2、35:G2//35:G20、及WT//WT 3组嫁接苗进行观察,发现接穗和砧木的表型性状与其各自基因型植株的性状并无较大差异(图3)。表明35:G20植株体内产生的大量活性GAs无法在嫁接植株中自由转运到35:G2中,而35:G2植株中的GA2-氧化酶也无法转化35:G20中的大量活性GAs;但培养在添加有GA4+7培养基中的WT//WT嫁接植株,相同生长时间内接穗和砧木的性状都受GAs影响,与35:G20表型相似,均表现为生长迅速,节间和叶片快速伸长,叶片颜色呈浅绿色细长的状态。
![]() 图 3 不同基因型组成的嫁接苗和不同基因型的烟草植株A.普通MS培养基中的WT//WT嫁接植株; B.添加有GA4+7的WT//WT嫁接植株; C.普通MS培养基中的35:G2//35:G20嫁接植株; D.普通MS培养基中的35:G20//35:G2嫁接植株; E.不同基因型的烟草植株。箭头代表嫁接处;A ~ D标尺长度为2 cm;E标尺长度为1 cm。The E scale is 1 cm in length; A,WT//WT grafted plants in normal MS medium; B,WT//WT grafted plants with GA4+7 were added; C, G2//35: G20 grafted plants in normal MS medium; D, 35:G20//35:G2 grafted plants in normal MS medium; E,different genotypes of tobacco plants. The arrow represents the graft; the A−D scale is 2 cm in length.Figure 3. Grafted seedlings of different genotypes and tobacco plants of different genotypes
图 3 不同基因型组成的嫁接苗和不同基因型的烟草植株A.普通MS培养基中的WT//WT嫁接植株; B.添加有GA4+7的WT//WT嫁接植株; C.普通MS培养基中的35:G2//35:G20嫁接植株; D.普通MS培养基中的35:G20//35:G2嫁接植株; E.不同基因型的烟草植株。箭头代表嫁接处;A ~ D标尺长度为2 cm;E标尺长度为1 cm。The E scale is 1 cm in length; A,WT//WT grafted plants in normal MS medium; B,WT//WT grafted plants with GA4+7 were added; C, G2//35: G20 grafted plants in normal MS medium; D, 35:G20//35:G2 grafted plants in normal MS medium; E,different genotypes of tobacco plants. The arrow represents the graft; the A−D scale is 2 cm in length.Figure 3. Grafted seedlings of different genotypes and tobacco plants of different genotypes2.3 下调烟草形成层GAs对茎部木质部生长发育的影响
通过转基因方法上调GAs含量促进植株(35:G20)生长,而下调GA含量的植株(35:G2、L:G2)生长缓慢[7],且下调植株整体GAs含量(35:G2)与特异下调形成层中的GAs含量对植株(L:G2)木质部发育的影响程度相似,说明植株GAs含量下降会使木质部的生长发育变延缓,植株的伸长生长也会受到抑制。通过茎部切片观察(图4),发现上调植株内源GAs含量(35:G20),木质部成熟提前,细胞排列整齐紧密且层数增加;下调GAs含量(35:G2、L:G2),木质部分化滞后,成熟缓慢,依据染色区域的大小发现同时期茎部相同位置的初生木质部多于WT,且细胞排列疏松不规则(图4)。转基因植株35:G2与L:G2无论是茎顶部亦或是茎基部组织中木质部的形成与发育的滞后程度、木质部细胞的排列方式等都极为相似,且下调木质部形成层的GAs含量可以达到下调植株整体GAs含量对木质部发育的影响,说明GAs影响植株木质部生长发育主要是在其形成层中发挥作用,形成层中的GAs含量对植株木质部的生长发育起关键作用。
![]() 图 4 内源调控GAs含量对植株木质部发育的影响PV. 初生导管组织;C. 形成层;X. 木质部;P. 韧皮部。标尺长度为100 μm。PV, primary ductal tissue; C, forming layer; X, xylem; P, phloem. Scale length is 100 μm.Figure 4. Effects of modification of endogenous GAs levels on xylem development
图 4 内源调控GAs含量对植株木质部发育的影响PV. 初生导管组织;C. 形成层;X. 木质部;P. 韧皮部。标尺长度为100 μm。PV, primary ductal tissue; C, forming layer; X, xylem; P, phloem. Scale length is 100 μm.Figure 4. Effects of modification of endogenous GAs levels on xylem development2.4 下调烟草形成层GAs对叶片生长的影响
对转基因植株和WT的叶片进行观察,发现L:G2和35:G2的叶片均呈墨绿色且生长紧密,且L:G2的叶片数有一定增加(图5)。此外,L:G2的叶片与其他株系相比出现生长不对称和皱化的现象,推测可能是形成层内GAs含量的变化调控了植株木质部的生长,根据叶片面积发生的明显变化判断当特异性降低形成层GAs的含量时延缓了叶脉的正常发育(图5)。
![]() 图 5 内源调控GAs含量对植株叶片发育的影响每行分别为各植株的全株叶片;标尺长度为1 cm。Each row is the whole plant leaf of each plant; the length of the scale is 1 cm.Figure 5. Effects of endogenous regulation of GAs content on leaf development of plants
图 5 内源调控GAs含量对植株叶片发育的影响每行分别为各植株的全株叶片;标尺长度为1 cm。Each row is the whole plant leaf of each plant; the length of the scale is 1 cm.Figure 5. Effects of endogenous regulation of GAs content on leaf development of plants2.5 下调烟草形成层GAs对不定根的影响
与对照组相比,不定根的数量在不同程度上均有所减少(图6A),通过t检验,发现其在0.01水平上差异显著,说明GAs含量的升高或降低对不定根的发育均有显著影响[9]。L:G2株系和35:G2株系不定根的生根性状在不定根数量和长度方面均相似(图6A、6B),表明特异性降低形成层中GAs的含量与降低植株整体GAs的含量对植株不定根的发育影响程度几乎相同,即木质部形成层中的活性GAs的含量是影响不定根发育的主要因素之一。
![]() 图 6 内源调控GAs含量对植株根生长发育的影响A、B为野生型和转基因植物不定根的平均数量和平均长度;C ~ F为不同转基因型植株根部,标尺长度为1 cm。C为WT株系;D为35:G2株系;E为35:G20株系;F为L:G2株系。A, B, the number and length of average adventitious roots of wild-type and transgenic plants. C−F, roots of different transgenic plants, the length of the scale is 1 cm. C, WT strain; D, 35: G2 strain; E, 35: G20 strain; F, L: G2 strain.Figure 6. Effects of endogenous regulation of GAs content on root growth and development of plants
图 6 内源调控GAs含量对植株根生长发育的影响A、B为野生型和转基因植物不定根的平均数量和平均长度;C ~ F为不同转基因型植株根部,标尺长度为1 cm。C为WT株系;D为35:G2株系;E为35:G20株系;F为L:G2株系。A, B, the number and length of average adventitious roots of wild-type and transgenic plants. C−F, roots of different transgenic plants, the length of the scale is 1 cm. C, WT strain; D, 35: G2 strain; E, 35: G20 strain; F, L: G2 strain.Figure 6. Effects of endogenous regulation of GAs content on root growth and development of plants在植株的侧根生长性状方面(图6C ~ 6F),L:G2株系几乎没有侧根产生,而35:G2的侧根与野生型WT相比有明显的增多,进一步说明特异性下调形成层内活性GAs含量对植株生长发育具有一定的调控作用,会抑制侧根的发生。
3. 讨 论
已有研究表明,赤霉素对茎部形成层、根尖的生长[9]及抗低温胁迫都有调控作用,可促进植物的生长[11-12],然而相关研究皆是以施加外源GA3或上调GAs含量为实验依据,以全株组成型调控赤霉素含量进行分析,未考虑活性赤霉素在活体内的运输特性对研究结果的影响。本研究通过对转基因及野生型烟草进行大量的观察与分析,发现特异性下调形成层赤霉素含量对植物的生长会有一定影响,如叶片不能正常伸展,木质部发育延缓,侧根和不定根的发生也会受到抑制。说明特异性调控植物局部的GAs含量对植株整体的生长发育造成了影响,且特异下调形成层区域的GAs可以达到下调植株整体GAs对木质部发育影响的效果,表明GAs在形成层中发挥了关键调控作用。
此外,利用烟草微嫁接技术将两种不同基因型的烟草嫁接后依然能生根并进行营养生长,且接穗与砧木的生长表型分别与其自身表型相同,说明活性内源GAs并不能在二者之间自由运输;而将WT//WT嫁接苗在添加有GA4+7的培养基中培养,发现嫁接苗的接穗与砧木都表现为GAs含量上调植株的表型,说明了外源的GAs通过根部吸收后,又可在体内自由运输。从以上两种结果可以判断,内源GAs和外源GAs在植物体内的运输途径可能不同,外源施加的活性GAS可能并不是通过主动运输而是通过导管组织随矿物质进行运输,而内源激素是通过形成层进行运输,这就是为什么微嫁接后内源活性GAs无法在植物体内自由运输的原因,具体机制有待于进一步的深入解析。本研究系统地证实了特异性调控形成层GAs含量在植株生长发育调控中的重要作用,为茎部赤霉素含量对植物生长发育的特异性调控机制的研究提供了理论基础,特别是赤霉素对木质部分化的特异性调控作用,为其在林木育种中的实际应用提供了依据。
-
![]()
图 1 桐棉松体胚发生途径植株再生
A.体胚诱导;B. 体胚增殖;C. 成熟培养;D. 诱导阶段胚性细胞显微观察;E. 增殖阶段胚性细胞显微观察;F. 成熟培养胚性细胞显微观察;G. 形成优势、成熟胚的胚性细胞团;H. 成熟体细胞胚;I. 体胚萌发培养;J. 生长中的萌发体胚;K. 再生体萌苗;L. 体胚苗移栽。标尺: 2 cm(A), 1cm(B, I−L), 0.5 cm(C), 2 mm(G, H), 500 μm(D, E), 200 μm(F). A, induction of somatic embryos; B, proliferation of somatic embryos; C, maturation culture; D, microscopic observation on induced somatic embryos; E, microscopic observation on proliferated somatic embryos; F, microscopic observation on somatic embryos during mature culture; G, embryogenic cells with dominative and mature somatic embryos; H, mature somatic embryos; I, germination of somatic embryos; J, growth of germinated somatic embryos; K, regenerated somatic seedlings; L, transplanting of somatic seedlings. Scale bars: 2 cm (A), 1cm (B, I−L), 0.5 cm (C), 2 mm (G, H), 500 μm (D, E), 200 μm (F).
Figure 1. Plant regeneration via somatic embryogenesis in Pinus massoniana ‘Tongmiansong’
![]()
图 2 通过芽苗复壮与不定根诱导的桐棉松萌发体胚离体植株再生
A.切除胚根的萌发体胚;B. 切根处理后的萌发体胚复壮培养;C. 复壮的体胚苗;D. 复壮体胚苗腋芽诱导;E. 形成丛生芽的复壮体胚苗;F. 复壮体胚苗的大量增殖与扩繁;G. 形成发达根系的生根苗;F. 生根苗苗圃移栽;I. 移栽3个月后成活的生根苗。标尺: 10 cm(F), 5 cm(I), 2 cm(H), 1cm(A−E, G). A, germinated somatic embryos with radicle cutting; B, reinvigoration of germinated somatic embryos after radicle cutting; C, reinvigorated shoots originated from somatic embryos; D, induction of axillary buds for reinvigorated shoots originated from somatic embryos; E, clustered shoots from reinvigorated shoots originated from somatic embryos; F, massive proliferation of reinvigorated shoots originated from somatic embryos; G, rooted shoots with robust root system; H, transplanting of rooted shoot in the nursery; I, survival seedlings after 3-month transplanting. Scale bars: 10 cm (F), 5 cm (I), 2 cm (H), 1 cm (A−E, G).
Figure 2. In vitro regeneration of plants originated from germinated somatic embryos of Pinus massoniana ‘Tongmiansong’ through shoot reinvigoration and adventitious root induction
![]()
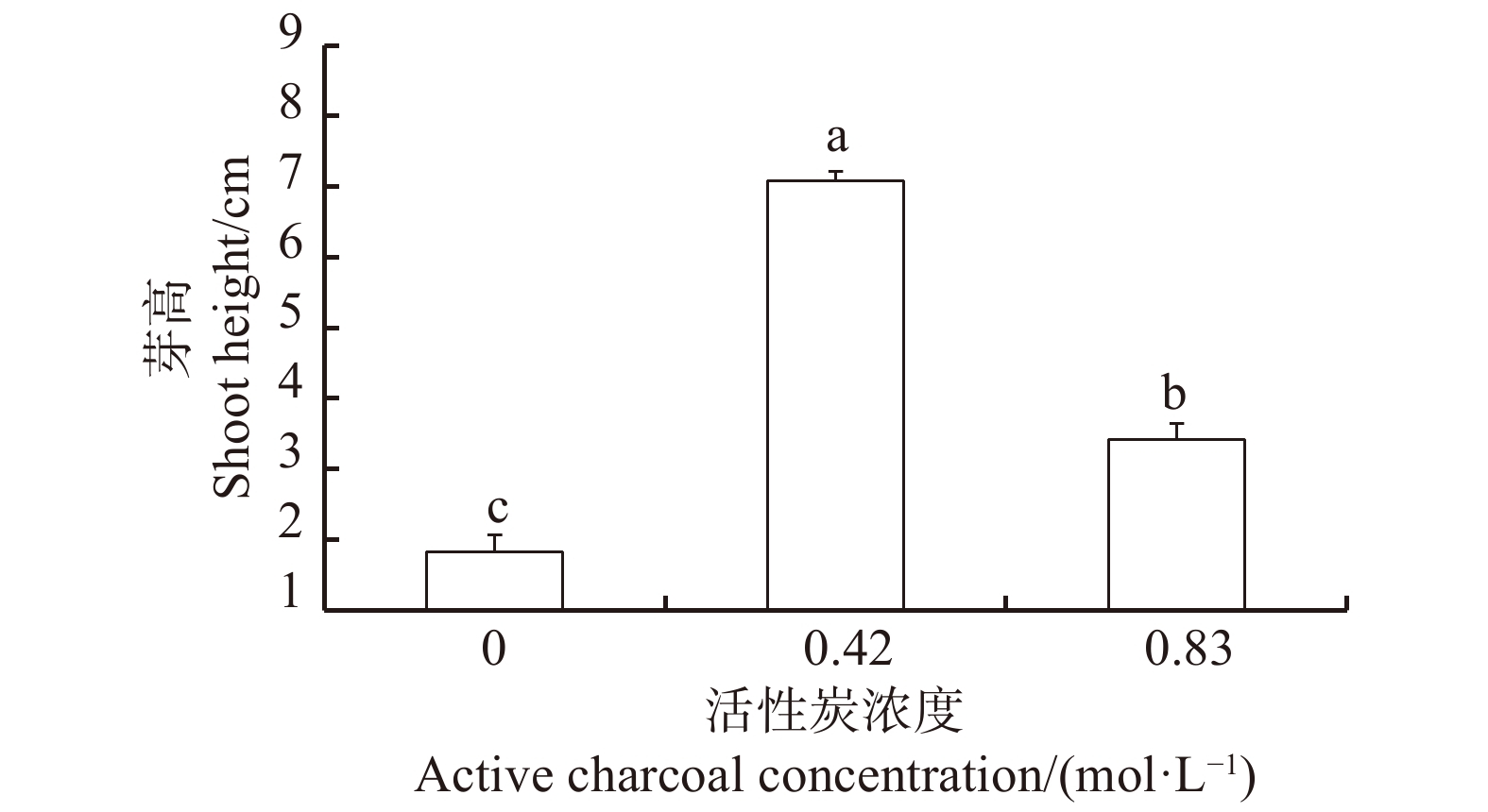
图 3 活性炭对桐棉松体胚萌发的影响
不同小写字母表示不同AC浓度处理间差异显著(P < 0.05)。Different lowercase letters indicate significant differences among varied AC concentration treatments at P < 0.05 level.
Figure 3. Effects of active charcoal on germination of somatic embryos in Pinus massoniana ‘Tongmiansong’
![]()
图 4 活性炭对桐棉松萌发体胚芽苗生长的影响
不同小写字母表示不同AC浓度处理间差异显著(P < 0.05)。Different lowercase letters indicate significant differences among varied AC concentration treatments at P < 0.05 level.
Figure 4. Effects of active charcoal on growth of shoots from germinated somatic embryos in Pinus massoniana ‘Tongmiansong’
表 1 基本培养基对桐棉松体胚萌发的影响
Table 1 Effects of basal media on germination of somatic embryos in Pinus massoniana ‘Tongmiansong’
基本培养基 Basal media 萌发率 Germination rate/% 萌发芽苗生长情况 Growth of shoots from germinated somatic embryos 1/2LP 68.5 ± 2.1b 叶片颜色深绿,顶芽形成缓慢,有褐化现象
Leaves are dark green, the formation of top shoots is slow, and the browning of shoots is occasionally observed1/2DCR 10.7 ± 2.2e 生长缓慢,叶片颜色发黄,褐化严重
Growth of shoots is slow, their leaves are yellow, and the browning of shoots is serious1/2GD 37.7 ± 2.6d 叶片短小、颜色发黄,顶芽形成困难,褐化现象明显
Leaves are short and yellow, the formation of top shoots is difficult, and the browning of shoots is obvious1/2MS 42.4 ± 3.2c 叶片呈浅绿色,形成顶芽水渍化现象明显
Leaves are light green, and the formed top shoots are easy to be water-soaking1/2MMS 94.1 ± 2.8a 叶片颜色翠绿,顶芽形成快、生长健壮,无褐化、玻璃化现象
Leaves are green, the formation of top shoots is efficient, and the shoots are robust without browning and vitrification注:同一列中不同小写字母表示不同基本培养基间差异显著(P < 0.05)。Note: different lowercase letters from the same column indicate significant differences among varied basal media at P < 0.05 level.  下载: 导出CSV
下载: 导出CSV
表 2 细胞分裂素对桐棉松萌发体胚芽苗增殖和生长的影响
Table 2 Effects of cytokinins on proliferation and growth of shoots originated from germinated somatic embryos in Pinus massoniana ‘Tongmiansong’
激素浓度
Hormone concentration/
(µmol·L− 1)有效芽增殖系数
Proliferation coefficient of effective shoots丛芽高度
Height of clustered shoots/cm芽生长情况
Shoot growth6-BA TDZ 6-BA TDZ 6-BA TDZ 0 0dA 0eA — — 未形成丛芽,针叶短小、颜色深绿,茎节间短,植株矮小
Cluster shoots are not found, leaves are short and dark green, internodes of shoots are short, and the whole plant is dwarf2 0.9 ± 0.2cA 1.1 ± 0.5dA 4.8 ± 0.4bB 8.1 ± 0.8bA 多呈单芽,大部分芽节间短、针叶密且呈深绿色
Most of shoots are single, their internodes are short, and leaves are compact and dark green丛生芽少,针叶细长,叶片为浅绿色
Cluster shoots are rare, leaves are tenuous and light green4 2.9 ± 0.7bB 5.6 ± 0.7aA 7.8 ± 0.8aB 9.2 ± 0.8aA 丛芽多,芽苗节间长,叶片颜色翠绿,生长健壮
Plenty of cluster shoots are observed, with long internodes, green leaves, and they grow well丛芽多,针叶长,叶片颜色翠绿,茎节间长,生长健壮
Plenty of cluster shoots are investigated, with long and green leaves, long internodes, and the whole shoots are thriving6 4.1 ± 0.5aA 3.3 ± 1.0bA 5.0 ± 0.8bB 6.7 ± 0.9cA 短簇状丛生芽较多,茎节间短,叶片颜色发黄,有褐化现象
Short cluster shoots are usually found, with short internodes, yellow leaves, and the browning of shoots is observed丛芽多,顶稍针叶较短,有部分叶片颜色发黄
Cluster shoots are sufficient, leaves of top shoots are short, and part of leaves are yellow8 2.5 ± 0.8bA 2.1 ± 0.7cA 3.6 ± 0.8cA 3.7 ± 0.4dA 丛芽呈短簇状,叶子卷曲,短小,褐化、玻璃化现象明显
Cluster shoots are short and small, leaves are cured and short, and the vitrification of shoots is remarkable丛芽矮小,叶片颜色发黄,有玻璃化现象
Cluster shoots are shorts, leaves are yellow, and the vitrification of shoots is occasionally found注:同一列中不同小写字母表示不同激素浓度处理间差异显著,同一行中不同大写字母表示两种激素处理间差异显著(P < 0.05)。Notes: different lowercase letters from the same column indicate significant differences among varied hormone concentration treatments, while different capital letters from the same row indicate significant differences between two hormone treatments at P < 0.05 level.
下载: 导出CSV
表 3 生长调节剂对桐棉松萌发体胚复壮芽苗生根能力的影响
Table 3 Effects of plant growth regulators on rooting capacity of reinvigorated shoots originated fromgerminated somatic embryos in Pinus massoniana ‘Tongmiansong’
激素处理
Hormone treatment生根率
Rooting rate/%生根时间
Rooting time/d根条数
Number of roots根长
Root length/cm成活率
Survival rate/%对照 Control 0c — — — — NAA 87.7 ± 4.5b 28.7 ± 2.1a 2.3 ± 0.4d 0.6 ± 0.2b 50.3 ± 3.6d NAA + IAA 91.4 ± 2.7ab 20.8 ± 1.5c 4.2 ± 0.7b 2.1 ± 0.4a 83.4 ± 3.2b NAA + IBA 90.5 ± 2.2ab 24.0 ± 2.3b 3.5 ± 0.8c 2.7 ± 0.4a 70.8 ± 4.6c NAA + PBZ 94.3 ± 3.8a 18.6 ± 1.2c 6.4 ± 0.7a 2.3 ± 0.6a 95.8 ± 2.4a 注:同一列中不同小写字母表示不同激素处理间差异显著(P < 0.05)。Note: different lowercase letters from the same column indicate significant differences among varied hormone treatments at P < 0.05 level.
下载: 导出CSV
-
[1] 黄健秋, 卫志明. 松属树种的组织培养和原生质体培养[J]. 植物学通报, 1994, 11(1):34−42. Huang J Q, Wei Z M. Tissue and protoplast culture of Pinus species[J]. Chinese Bulletin of Botany, 1994, 11(1): 34−42.
[2] 丁贵杰, 周志春, 王章荣, 等. 马尾松纸浆用材林培育与利用[M]. 北京: 中国林业出版社, 2006: 1–10. Ding G J, Zhou Z C, Wang Z R, et al. Cultivation and utilization of pulpwood stand for Pinus massoniana[M]. Beijing: China Forestry Publishing House, 2006: 1–10.
[3] 杨章旗, 刘达峰. 马尾松: 广西优良用材树种[J]. 广西林业, 2011, 29(8):41−42. doi: 10.3969/j.issn.1004-0390.2011.08.020 Yang Z Q, Liu D F. Pinus massoniana: superior timber tree species of Guangxi[J]. Guangxi Forestry, 2011, 29(8): 41−42. doi: 10.3969/j.issn.1004-0390.2011.08.020
[4] 杨模华, 张冬林, 李志辉, 等. 马尾松幼胚体细胞胚胎发生研究[J]. 植物生理学报, 2011, 47(9):904−912. Yang M H, Zhang D L, Li Z H, et al. Somatic embryogenesis with immature embryos of masson pine (Pinus massoniana Lamb.)[J]. Plant Physiology Journal, 2011, 47(9): 904−912.
[5] 季孔庶, 王章荣, 陈天华, 等. 马尾松扦插繁殖年龄效应及继代扦插复壮效果[J]. 浙江林学院学报, 1996, 16(4):341−345. Ji K S, Wang Z R, Chen T H, et al. Cyclophysis and effect of rejuvenation with continued cuttage in Pinus massoniana cutting propagation[J]. Journal of Zhejiang Forestry College, 1996, 16(4): 341−345.
[6] Klimaszewska K, Hargreaves C, Lelu-Walter M, et al. Advances in conifer somatic embryogenesis since year 2000[M]//Germanà M A, Lambardi M. In vitro embryogenesis in higher plants, methods in molecular biology. New York: Springer Science Business Media, 2016: 131–166.
[7] Pullman G S, Zeng X, Copeland-Lamp B, et al. Conifer somatic embryogenesis: improvements by supplementation of medium with oxidation-reduction agents[J]. Tree Physiology, 2015, 35(2): 209−224. doi: 10.1093/treephys/tpu117
[8] Thomas T D. The role of activated charcoal in plant tissue culture[J]. Biotechnology Advances, 2008, 26(6): 618−631. doi: 10.1016/j.biotechadv.2008.08.003
[9] De Diego N, Montalbán I A, Fernández E, et al. In vitro regeneration of Pinus pinaster adult trees[J]. Canadian Journal of Forestry Research, 2008, 38(10): 2607−2615. doi: 10.1139/X08-102
[10] Pan J J, Van Staden J. The use of activated charcoal in in vitro culture: a review[J]. Plant Growth Regulation, 1998, 26(3): 155−163. doi: 10.1023/A:1006119015972
[11] Yao R L, Wang Y. An effective protocol for regenerating mature Pinus massoniana L. trees by tissue culture[J]. Research Journal of Biotechnology, 2016, 11(12): 75−80.
[12] 黄健秋, 卫志明, 许智宏. 马尾松成熟合子胚的体细胞胚胎发生和植株再生[J]. 植物学报, 1995, 37(4):289−294, 338. Huang J Q, Wei Z M, Xu Z H. Somatic embryogenesis and plantlet regeneration from callus of of mature zygotic embryos of masson pine[J]. Acta Botanica Sinica, 1995, 37(4): 289−294, 338.
[13] 李校雨, 吕成群, 黄宝灵, 等. 马尾松组培苗不定根诱导及不定根解剖观察[J]. 西北林学院学报, 2009, 24(3):80−84. Li X Y, Lü C Q, Huang B L, et al. Adventitious roots’ induction of Pinus massoniana shoots in test tubes and anatomical observation[J]. Journal of Northwest Forestry College, 2009, 24(3): 80−84.
[14] 伊书亮, 张冬林, 杨模华, 等. 外植体采集时期与冷藏处理对马尾松愈伤组织诱导的影响[J]. 广西林业科学, 2013, 42(1):8−13. doi: 10.3969/j.issn.1006-1126.2013.01.002 Yin S L, Zhang D L, Yang M H, et al. Effects of explant collecting time and storage duration on callus induction of Pinus massoniana[J]. Guangxi Forestry Science, 2013, 42(1): 8−13. doi: 10.3969/j.issn.1006-1126.2013.01.002
[15] Wang Y, Yao R L. Plantlet regeneration of adult Pinus massoniana Lamb. trees using explants collected in March and thidiazuron in culture medium[J]. Journal of Forestry Research, 2017, 28(6): 1169−1175. doi: 10.1007/s11676-017-0412-9
[16] Aitken-Christie J, Singh A P, Davies H. Multiplication of meristematic tissue: a new tissue culture system for radiata pine[M]//Hanover J W, Keathley D E. Genetic manipulation of woody plants. New York: Plenum, 1988: 413–432.
[17] Gupta P K, Durzan D J. Shoot multiplication from mature trees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinus lambertiana)[J]. Plant Cell Reports, 1985, 4(4): 177−179. doi: 10.1007/BF00269282
[18] Gresshoff P M, Doy C H. Development and differentiation of haploid Lycopersicon esculentum (tomato)[J]. Planta, 1972, 107(2): 161−170. doi: 10.1007/BF00387721
[19] Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures[J]. Physiologia Plantarum, 1962, 15(3): 473−497. doi: 10.1111/j.1399-3054.1962.tb08052.x
[20] Wang Y, Yao R L. Increased endogenous indole-3-acetic acid:abscisic acid ratio is a reliable marker of Pinus massoniana rejuvenation[J]. Biotechnic & Histochemistry, 2019, 94(7): 546−553.
[21] Van Winkle S C, Pullman G S. The combined impact of pH and activated carbon on the elemental composition of a liquid conifer embryogenic tissue initiation medium[J]. Plant Cell Reports, 2003, 22(5): 303−311. doi: 10.1007/s00299-003-0686-6
[22] 李国树, 徐成东, 王波, 等. 植物组织培养节能降耗研究进展[J]. 植物学研究, 2014, 3(3):105−110. doi: 10.12677/BR.2014.33015 Li G S, Xu C D, Wang B, et al. Research progress on plant tissue culture system of saving energy and reducing consumption[J]. Botanical Research, 2014, 3(3): 105−110. doi: 10.12677/BR.2014.33015
[23] 曾嬿冰, 周运超, 张伟, 等. 马尾松优良种源对N肥的响应[J]. 贵州林业科技, 2016, 44(1):1−8. Zeng Y B, Zhou Y C, Zhang W, et al. Response of superior provenance of Pinus massoniana to N fertilize[J]. Guizhou Forestry Science and Technology, 2016, 44(1): 1−8.
[24] 潘瑞炽. 植物生理学[M]. 7版. 北京: 高等教育出版社, 2012. Pan R Z. Plant physiology[M]. 7th ed. Beijing: Higher Education Press, 2012.
[25] 刘忠新, 刘莉梅. 浅议植物生长所必须的营养元素与其生理功能[J]. 现代农业研究, 2007, 13(12):8. doi: 10.3969/j.issn.1674-0653.2007.12.007 Liu Z X, Liu L M. Discussion on vital nutrient elements for plant growth and their physiological function[J]. Modern Agriculture Research, 2007, 13(12): 8. doi: 10.3969/j.issn.1674-0653.2007.12.007
[26] Yolande P, Patrick D, Ludovic L, et al. Endogenous cytokinins as biochemical markers of rubber-tree (Hevea brasiliensis) clone rejuvenation[J]. Plant Cell, Tissue and Organ Culture, 1997, 47(3): 239−245. doi: 10.1007/BF02318978
[27] 曾少玲, 方良全, 吉文, 等. 桉树组织培养中的玻璃化现象及克服措施[J]. 桉树科技, 2002, 25(1):30−31. doi: 10.3969/j.issn.1674-3172.2002.01.006 Zeng S L, Fang L Q, Ji W, et al. Vitrification and its countermeasures during tissue culture of eucalypts[J]. Eucalypt Science & Technology, 2002, 25(1): 30−31. doi: 10.3969/j.issn.1674-3172.2002.01.006
[28] 段娜, 贾玉奎, 徐军, 等. 植物内源激素研究进展[J]. 中国农学通报, 2015, 31(2):159−165. doi: 10.11924/j.issn.1000-6850.2014-2335 Duan N, Jia Y K, Xu J, et al. Research progress on plant endogenous hormones[J]. Chinese Agricultural Science Bulletin, 2015, 31(2): 159−165. doi: 10.11924/j.issn.1000-6850.2014-2335
[29] 姚瑞玲, 王胤, 吴幼媚. 马尾松组培生根关键因子分析[J]. 广西植物, 2016, 36(11):1288−1294. Yao R L, Wang Y, Wu Y M. Analysis for key factors affecting rooting in Pinus massoniana by tissue culture[J]. Guihaia, 2016, 36(11): 1288−1294.
[30] Watson G. Effect of transplanting and paclobutrazol on root growth of ‘Green Column’ black maple and ‘Summit’ green ash[J]. Journal of Environmental Horticulture, 2004, 22(4): 209−212.
[31] Kamran M, Wennan S, Ahmad I, et al. Application of paclobutrazol affect maize grain yield by regulating root morphological and physiological characteristics under a semi-arid region[J]. Scientific Reports, 2018, 8(1): 4818−4832. doi: 10.1038/s41598-018-23166-z
[32] Fu X, Harberd N P. Auxin promotes Arabidopsis root growth by modulating gibberellin response[J]. Nature, 2003, 421: 740−743. doi: 10.1038/nature01387
[33] 钮世辉, 李伟, 陈晓阳. 赤霉素对根尖径向生长的调节作用研究[J]. 北京林业大学学报, 2013, 35(3):71−76. Niu S H, Li W, Chen X Y. Negative regulation of gibberellin on root tip diameter[J]. Journal of Beijing Forestry University, 2013, 35(3): 71−76.
[34] Mauriat M, Petterle A, Bellini C, et al. Gibberellins inhibit adventitious rooting in hybrid aspen and Arabidopsis by affecting auxin transport[J]. Plant Journal, 2014, 78(3): 372−384. doi: 10.1111/tpj.12478
[35] Vaičiukynė M, Žiauka J, Žūkienė R, et al. Abscisic acid promotes root system development in birch tissue culture: a comparison to aspen culture and conventional rooting-related growth regulators[J]. Physiologia Plantarum, 2019, 165(1): 114−122. doi: 10.1111/ppl.12860
[36] Takáč T, Obert B, Rolčík J, et al. Improvement of adventitious root formation in flax using hydrogen peroxide[J]. New Biotechnology, 2016, 33(5): 728−734. doi: 10.1016/j.nbt.2016.02.008
[37] Cano A, Sánchez-García A B, Albacete A, et al. Enhanced conjugation of auxin by GH3 enzymes leads to poor adventitious rooting in carnation stem cuttings[J]. Frontiers in Plant Science, 2018, 9(4): 1−17.
-
期刊类型引用(7)
1. 王为,雷俊杰,简佶沛,王利宝. 基于云平台智能灌溉控制系统的油茶苗水分管理研究. 现代农业科技. 2023(20): 110-113+121 .  百度学术
百度学术
2. 董诗芬,王自洪,李丽华,李看清. 不同覆盖措施对初植腾冲红花油茶生长的影响研究. 林业调查规划. 2022(03): 118-121+142 . 百度学术
3. 谢胤,余祖华,尹必期,王自洪,寸明辉,徐志映,吴兴波,杨忠品. 腾冲红花油茶主要营养器官含水率年内变化分析. 林业与环境科学. 2021(01): 25-28 . 百度学术
4. 胡玉玲,蔡芳丽,卢海燕,罗海秀,贺姣凤. 油茶林地夏季水分管理对油茶产量指标的影响. 江苏林业科技. 2018(03): 23-27+45 . 百度学术
5. 刘嘉翔,赵丹,杨建伟,史宝胜. 不同土壤水分条件下北京山梅花生长与耗水特性研究. 河北农业大学学报. 2018(05): 84-89 . 百度学术
6. 何小三,徐林初,龚春,王玉娟,刘新亮,赵攀,左继林,俞元春. 干旱胁迫对‘赣无12’苗期光合特性的影响. 中南林业科技大学学报. 2018(12): 52-61 . 百度学术
7. 樊星火,樊文勇,黄辉,施重阳,郑永红. 夏季不同灌溉方式对油茶叶片生理指标和花期的影响. 林业科技通讯. 2017(12): 12-14 . 百度学术
其他类型引用(4)









计量
- 文章访问数: 1963
- HTML全文浏览量: 598
- PDF下载量: 78
- 被引次数: 11
