
Relationship between tree growth and tree size, competition as well as species diversity in spruce-fir natural forests
-
摘要:目的
林分内林木大小、竞争和树种多样性等多方面因素影响着林木的生长,而胸高断面积生长量通常被用来描述树木生长状态。本文利用长白山云冷杉针阔混交林主要树种的单木胸高断面积生长量建立随机森林模型,研究和量化影响树木生长的环境因素,旨在为该地区的云冷杉针阔混交林生长预估提供理论依据。
方法连续24年(1987—2010年)对总样木数为6 903株的固定样地进行数据调查,应用随机森林算法,选取单木、竞争因子、多样性和气候方面共11个调查因子,对混交林中6个主要树种建立胸高断面积生长量模型,并使用10折交叉验证法来优化超参数mtry和评估模型结果。
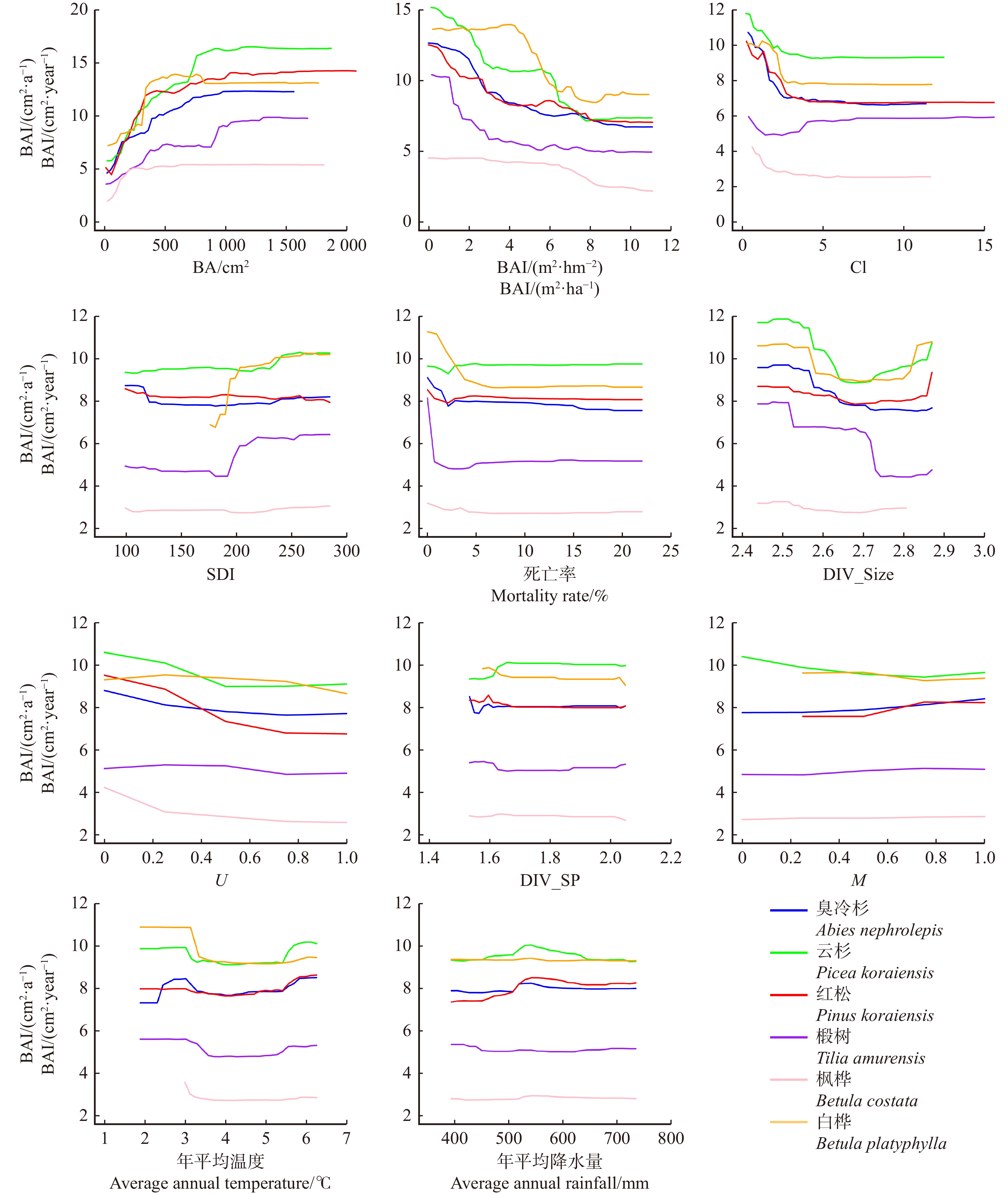
结果(1)臭冷杉、云杉、红松、椴树、枫桦、白桦6个主要树种胸高断面积生长量模型的决定系数分别为0.663、0.683、0.695、0.459、0.384和0.568。(2)单木胸高断面积是最重要的因子,对树木生长有着很强的正向影响;竞争因子和树木大小多样性是影响树木生长的主要因素,单木胸高断面积生长量随着竞争因子、树木大小多样性增加而下降。(3)树种多样性对树木生长的影响比较有限,树种多样性指数和混交度的增加会一定程度加快云杉、臭冷杉和红松的生长速度;而气候因子对树木生长的影响则相对较小。
结论树木生长在很大程度上依赖于其自身的生长潜力,在外界环境中主要受到来自林木间竞争和树木大小多样性的抑制,而树种多样性的增加也能在一定程度促进林内优势树种的生长;随机森林模型能够很好地量化和显出各变量与单木胸高断面积生长量之间复杂的关系,可以作为森林管理评价工具,为森林生长收获预估提供新的方法。
Abstract:ObjectiveTree size, competition, tree species diversity and other factors in the forest stand affect the growth of forest trees, while the individual basal area increment (BAI) is often used to describe tree growth. A random forest model was established to study the individual tree BAI of main tree species in the mixed forest of Changbai Mountain of northeastern China, study and quantify the environmental mechanisms affecting tree growth, aiming to provide a theoretical basis for growth projections.
MethodData were investigated for 24 consecutive years (1987−2010) in a fixed sample plot with a total sample size of 6 903 trees. Random forest algorithm was used to build individual tree BAI model with 11 independent variables as individual tree, competition factor, diversity, and climate for 6 main tree species in mixed forest. And 10-fold cross-validation was used to optimize hyper parameter mtry and evaluate these models.
Result(1) The coefficients of determination for the model of BAI for the six major species of Abies nephrolepis, Picea koraiensis, Pinus koraiensis, Tilia amurensis, Betula costata, and Betula platyphylla were 0.663, 0.683, 0.695, 0.459, 0.384, and 0.568, respectively. (2) The basal area (BA) of individual tree was the most important independent variable, and had a strong positive effect on the growth of trees. Competitive factors and tree size diversity were the main factors affecting tree growth, and individual BAI decreased with increasing competitive factors and tree size diversity. (3) The effects of species diversity on tree growth were relatively limited, with increases in species diversity index and mingling degrees accelerating the growth of Abies nephrolepis, Picea koraiensis and Pinus koraiensis to some extent; whereas the effects of climatic factors on tree growth were relatively small.
ConclusionTree growth is largely depends on its own growth potential, which is mainly inhibited by competition and tree size heterogeneity in the external environment, while increased species diversity also promotes the growth of dominant species within the forest to some extent. The random forest model can well quantify and express the complex relationship between the variables and BAI of individual tree. It can be used as a tool for forest management practice and providing a new method for forest growth and yield prediction.
-
Keywords:
- tree growth /
- tree size /
- competition /
- tree species diversity /
- random forest model
-
荒漠化是全球性的重大环境问题,我国是世界上荒漠化危害最严重的国家之一。目前我国为防治荒漠化进一步的扩展,采取了最有效、最稳定的植被建设措施—封沙育林育草恢复天然植被[1]。在生态环境恶劣,水资源短缺的荒漠化地区进行植被恢复,需要种植耐旱节水型树种。在毛乌素沙地区域,沙木蓼(Atraphaxis bracteata)是该地区主要造林树种之一,具有较好的固沙作用。但在沙木蓼生长到第4年以后,其株高和地径的生长量均下降,并有枝条枯死现象,需要平茬更新[2];这不仅会增大荒漠化地区造林工程的工作量,也会减缓荒漠化地区植被恢复速度,减弱人工沙木蓼群落的稳定性。此外,沙木蓼虽能在土壤水分含量较低的沙地中生长,但由于其根系分布较广,消耗水量多,深层及周边土壤水分得不到补充[3],导致沙木蓼周围植物群落物种数减少,周围物种较为单一,影响着沙木蓼造林区的植物多样性。因此,有必要实施一些措施促进沙木蓼生长,增加群落植物多样性,改善荒漠化地区的土壤理化性质。
在荒漠生态系统中,水分和养分共同限制着植物的生长和繁殖[4]。施肥就是促进植物生长和繁殖的有效措施,也是恢复荒漠化草地植物群落多样性的主要途径之一[5-9]。目前在荒漠地区的植被恢复过程中,水分与养分的调控措施大多都是施用氮、磷肥等无机肥料,但长时间过量施用容易造成土壤酸碱度不平衡、土壤板结、环境污染[10]。而有机肥不仅可以改善土壤的物理、化学和生物学性状,还可以扩大土壤的有效养分库,从而改善土壤的养分状况,提高养分的供给力[11]。马红梅等研究发现:对荒漠区造林植物进行专用有机肥和化学肥料的混合施用,能够显著增加其株高、分枝数、层数、主茎直径、冠幅、鲜质量和干质量,提高其耐旱性[12]。更有研究表明施肥改变了土壤的养分含量分布,影响着造林区的植物群落多样性的变化[13-14]。
虽然有机肥的养分全面,种类多,但目前使用的有机肥大多为粪尿肥、堆沤肥、绿肥和杂肥,其养分释放慢,常常会带某些有害成分,例如臭味、病原菌、寄生虫卵和重金属等。而鲜海带生物酶解有机液肥是一种新型的有机液肥,采用100%新鲜海带为原料,应用海藻超微破碎技术和微生物酶解技术,提高了各营养物质和天然植物活性因子的提取率,保留了海带提取物的多样性和生物活性。富含植物所需的多种营养物质、微量元素和天然植物生长刺激素,对水果类、蔬菜类和粮油类都具有增产的效果。根据Xu等对藻类有机肥研究表明,叶面喷施藻类有机肥可促进冬小麦智能化生长,增粗抗倒状,显著提高了单株穗质量的同时,增加了土壤密度,提高小麦籽粒的物理品质,有利于叶片叶绿素的合成和缓解叶绿素的分解[15]。Liu等利用藻类有机肥对蔬菜种子萌发和幼苗生长的研究表明,藻类有机肥对提高发芽率和细苗壮苗具有较好的效果[16]。目前藻类有机肥主要应用在果蔬类农作物上, 并具有显著的效果,但在荒漠化地区沙生植物上应用的效果尚待进一步研究。
因此,本文选择宁夏盐池植被恢复区中的5年生沙木蓼作为研究对象,通过人工控制不同梯度的鲜海带生物酶解有机液肥施用量的试验处理,探讨该有机液肥对荒漠化地区土壤理化性质以及沙木蓼生长和群落物种多样性的影响,以期为荒漠化地区植被恢复提供理论支持,为在荒漠化造林区沙木蓼的管理和植被恢复提供实践依据。
1. 材料与方法
1.1 试验区概括
试验位于宁夏盐池荒漠生态系统国家定位观测研究站(107°20′~107°26′ E、37°04′~37°10′ N),该站位于我国半干旱区毛乌素沙地西南缘的盐池县沙泉湾,总面积2 000 hm2,地处黄土高原南部水蚀区与鄂尔多斯高原风蚀区的边界线附近,属鄂尔多斯台地。该区海拔为1 354 m,常年干旱少雨,蒸发量大,风大沙多,属典型的中温带大陆性季风气候,年平均温度为8.1 ℃,年降雨量280 mm左右,降水主要集中在夏秋两季,7—9月降水合计占全年总降水量的62%;光能丰富,热量较适中,年太阳辐射值为586 J/cm2,年蒸发量为2 100 mm,年无霜期为165 d。植被类型有灌丛、草原、沙地植被、草甸和荒漠植被。该地区土壤为风沙土,地表物质疏松,土壤贫瘠,沙源物质丰富,风大且频,风沙活动强烈[17]。为防治该地区沙漠化进一步的扩展,主要实行封沙育林育草恢复天然植被和造林种草固沙措施,沙木蓼是造林固沙的主要灌木之一。
试验开始时,土壤0~20 cm的全氮含量为15 g/kg,土壤0~20 cm的有机质含量为9.51 g/kg,土壤0~10 cm的含水率在4%~6%之间,样方中沙木蓼调查情况如表 1所示,沙木蓼的地径和株高差异不显著(P>0.05)。
表 1 沙木蓼地径与株高的原始值分析Table 1. Analysis on original values of ground diameter and tree height of Atraphaxis bracteatacm 施肥处理
Fertilization treatment植物指标
Plant index肥料用量Fertilizing amount 0 3 mL/m2 6 mL/m2 9 mL/m2 12 mL/m2 15 mL/m2 冲施肥Flush fertilizer 地径Ground diameter 0.50a 0.49a 0.53a 0.54a 0.57a 0.53a 株高Plant height 104.87a 103.87a 119.67a 104.93a 106.40a 104.93a 叶面肥Foliar fertilizer 地径Ground diameter 0.50a 0.53a 0.49a 0.54a 0.56a 0.55a 株高Plant height 104.87a 103.33a 100.33a 108.73a 108.60a 104.67a 注:不同小写字母表示不同处理之间差异显著(P>0.05)。Note: different lowercases mean significant difference among varied treatments at P>0.05 level. 1.2 试验设计
1.2.1 试验肥料
本试验中使用的鲜海带生物酶解有机液肥是由山东省威海市世代海洋生物科技股份有限公司提供,依据鲜海带生物酶解有机液肥种类,分为叶面肥和冲施肥2种,冲施肥随灌溉水施加,叶面肥利用喷雾器对植被叶片喷施。
表 2 鲜海带生物酶解有机液肥主要成分Table 2. Main ingredients of fresh seaweed bio-enzymatic liquid organic fertilizer液肥
Liquid fertilizer主要成分
Main component叶面肥
Foliar fertilizer海带酶解液、海藻多糖、海藻蛋白、甘露醇、甜菜碱、赤霉素、吲哚乙酸、脱落酸、氨基酸等Kelp hydrolysate, seaweed polysaccharide, seaweed protein, mannitol, betaine, gibberellin, indoleacetic acid, abscisic acid, amino acid, etc 冲施肥
Flush fertilizer海带纤维、有机质、海藻多糖、吲哚乙酸、海藻蛋白、甘露醇、甜菜碱等Kelp fiber, organic matter, seaweed polysaccharide, indoleacetic acid, seaweed protein, mannitol, betaine, etc 1.2.2 试验设置
2016年4月,在研究区域内选择地势平坦且5年生沙木蓼,设定2个区组,每个区组包含18个3 m×3 m的样方,每个样方之间有2 m的缓冲带。试验设置2种施肥方式,6个施肥量处理,每种施肥处理3个重复,水肥配比为1:200,6种肥料用量分别为:0(对照CK)、3、6、9、12、15 mL/m2。于春季植物生长开始,每40 d施1次鲜海带生物酶解有机液肥,共施4次,不施肥的对照处理施等量的水分,施肥结束后进行补水措施,确保水分是控制变量,同时为了保证补水量不对施肥有影响,补水在施肥结束后第2天进行。施入的总水量是根据该研究区近10年降水量集中的7—9月日平均降水量换算确定的。
表 3 小区施肥用量Table 3. Fertilizing amount in 3 plots肥料用量
Fertilizing amount/(mL·m-2)小区肥料用量
Community fertilizing amount/mL小区用水量
Community water consumption/mL小区补水量
Community water recharge/mL0 0 27 000 0 3 27 5 400 21 600 6 54 10 800 16 200 9 81 16 200 10 800 12 108 21 600 5 400 15 135 27 000 0 ![]() 图 1 试验小区布设图CK.对照组;Y.叶面肥;C.冲施肥。数字表示肥料用量,单位为mL/m2。Figure 1. Layout of experimental plotsCK, control; Y, foliar fertilizer; C, flush fertilizer. The numbers represent the amount of fertilizer, the unit is mL/m2.
图 1 试验小区布设图CK.对照组;Y.叶面肥;C.冲施肥。数字表示肥料用量,单位为mL/m2。Figure 1. Layout of experimental plotsCK, control; Y, foliar fertilizer; C, flush fertilizer. The numbers represent the amount of fertilizer, the unit is mL/m2.1.2.3 试验监测与采样
定期调查沙木蓼的地径、株高、新梢的生长情况,并在8月中旬植物生物量达到最大时,调查、记录各样方内植被的物种数、株高、密度和盖度。在7月初施肥结束后,对土壤0~10 cm含水量按对角线定点测量,测量频率是在施肥结束后的第1天(7月2日)、第4天(7月5日)、第9天(7月13日)、第16天(7月24日)和第25天(8月9日)的一个周期监测。同时,在生长期结束之后,进行土壤0~20 cm的取样,每个样方随机选取5个取样点,用直径3.5 cm的土钻取样,风干后过2 mm筛,带回实验室进行土壤化学性质的测定。
(1) 土壤理化性质的测定。土壤物理性质:土壤含水率用土壤水分测定仪(仪器型号:ML3,生产商:北京易科泰生态技术有限公司)测定。土壤化学性质:土壤全氮采用凯氏定氮仪法;土壤有机质采用重铬酸钾氧化-外加热法。
(2) 物种丰富度的测定。物种丰富度用记名计算法测定,本研究中物种丰富度用1 m×1 m=1 m2样方内出现的物种数表示,即物种丰富度指数=出现在1 m2样方内的物种数[18]。为了消除植物的不均衡分布的影响,本试验采用包含杂草密度和频度的相对丰度值(RA)作为衡量某种杂草重要程度的指标,其计算方式为:
RA=(RD+RF)/2 式中:RD表示相对密度,小区中某种杂草的密度除以小区中所有杂草的密度之和;RF表示相对频度,小区中某种杂草的频度除以小区中所有杂草的频度之和。
(3) 物种多样性的测定。本研究中物种多样性选用Margalef物种丰富度指数、Simpson优势度指数、Shannon-Wiener多样性和Pielou均匀度指数来测定。
Margalef物种丰富度指数:
DMG=(S−1)/lnN 式中:S为样方内物种种类数量,N为样方内中所有物种的个体数量。
Simpson优势度指数:
Di=1−∑P2i 式中:Pi为物种相对重要性,即物种的个体数在样方内所有物种个体数中所占的比例。
Shannon-Wiener多样性:
H′=−∑PilnPi Pielou均匀度指数:
J=(−∑PilnPi)/lnS,即J=H′/lnS 1.3 数据处理
数据用SPSS20.0进行单因素方差分析检验不同水平鲜海带生物酶解有机液肥的添加对荒漠化土壤理化性质、沙木蓼生长以及其群落物种多样性的影响,多重比较采用LSD检验,利用EXCEL2010制图。
2. 结果与分析
2.1 鲜海带生物酶解有机液肥对土壤理化性质的影响
2.1.1 土壤含水率
土壤含水率关系到土壤养分的形成、转移和吸收,以及植株的正常生长[19],适宜的土壤含水率能够促进有机肥的分解,提高肥料的利用率。而土壤含水率又与降雨量、土壤蒸发量有着密切的关系,据研究表明,在毛乌素沙地降水对土壤蒸发影响较大,降水使土壤水分增加,土壤中可供蒸发的水分充足,土壤蒸发就会增大[20]。而从图 2~4看出,7月4日降水为2.2 mm,蒸发量却由前一天的8.62 mm降为5.77 mm,在叶面肥研究区内,7月5日的0~10 cm土壤含水率高于其他测量时间的土壤含水率,因此,鲜海带酶解有机液肥有利于土壤保水。由图 3、4可以看出,不同的施肥方式对土壤含水率具有不同的影响,冲施肥研究区的土壤含水率较叶面肥研究区提高了10.6%,且冲施肥土壤含水率较叶面肥的土壤含水率稳定,各处理间变化幅度在0.1%~0.4%之间,说明冲施肥比叶面肥更有利于土壤保水。
![]() 图 2 2016年7—8月降雨量与土壤蒸发量Figure 2. Rainfall and soil evaporation during July to August in 2016
图 2 2016年7—8月降雨量与土壤蒸发量Figure 2. Rainfall and soil evaporation during July to August in 2016![]() 图 3 冲施肥对沙木蓼土壤含水率的影响不同字母表示处理间差异显著(P<0.05)。下同。Figure 3. Effects of flush fertilizer on the soil water content of Atraphaxis bracteataDifferent letters indicate significant difference among treatments at P < 0.05 level. The same below.
图 3 冲施肥对沙木蓼土壤含水率的影响不同字母表示处理间差异显著(P<0.05)。下同。Figure 3. Effects of flush fertilizer on the soil water content of Atraphaxis bracteataDifferent letters indicate significant difference among treatments at P < 0.05 level. The same below.![]() 图 4 叶面肥对沙木蓼土壤含水率的影响Figure 4. Effects of foliar fertilizer on soil water content of Atraphaxis bracteata
图 4 叶面肥对沙木蓼土壤含水率的影响Figure 4. Effects of foliar fertilizer on soil water content of Atraphaxis bracteata冲施肥研究区的土壤含水率日观测表明(图 3),冲施肥可以增加土壤含水率,减小沙木蓼的耗水与蒸发。经过施肥处理的土壤含水率在施肥后第1天(7月2日)迅速增长之后,在7月5日、7月13日、7月24日和8月9日保持较稳定的状态,日变化幅度小于0.6%,施肥处理除了15 mL/m2的土壤含水率比对照低0.11%,其他施肥处理的土壤含水率均高于对照,是对照的109.39%~117.35%,并且随着冲施肥用量的增加,土壤含水率呈现先增加后减小的趋势。这是由于在高用量的情况下,植被因生理过程对水分的需求增大,土壤含水率就会降低。方差分析表明,除了8月9日的不同施肥处理间的土壤含水率的差异不显著(P>0.05)之外,其他时间的不同施肥处理间的土壤含水率均显著(P < 0.05)。
叶面肥研究区的土壤含水率的变化幅度较冲施肥土壤含水率的大(图 4),变化幅度在3.50%~10.00%之间。除了12 mL/m2的土壤含水率比对照高0.14%,其他施肥处理土壤含水率为对照的86.64%~96.45%。随着叶面肥用量的增加,土壤含水率呈现先降低后增加的趋势,可能是因为叶面肥利用喷施的方式,叶面吸收较多,用量小的叶面肥只能提供叶片的使用,无法进入土壤,导致叶面肥用量大的土壤含水率较高。方差分析表明,不同叶面肥处理土壤含水率差异不显著(P>0.05)。
2.1.2 土壤全氮、有机质含量
毛乌素沙地的土壤母质不含氮,土壤中的氮主要靠施肥(包括有机肥和化学氮肥)和植物残体归还而补充[21]。鲜海带生物酶解有机液肥增加了沙木蓼土壤0~20 cm层的全氮含量(图 5),方差分析表明,冲施肥除了处理12 mL/m2与对照不显著,其余处理均与对照间差异显著(P < 0.05),较对照增加了4.40%~37.14%。叶面肥除了处理3 mL/m2和12 mL/m2,其余处理均与对照差异显著(P < 0.05),较对照增加了5.56%~31.43%。从土壤全氮的增加量来看,施冲施肥和叶面肥的效果一样,没有明显的差异。
土壤0~20 cm层的有机质含量随着鲜海带生物酶解有机液肥用量的增大而显著增加(图 6),冲施肥的每个处理都与对照差异显著(P < 0.05),处理3和6 mL/m2的有机质含量分别比对照低8.33%和8.57%,其他处理均高于对照,分别高出100%、200%和125%。叶面肥除了处理15 mL/m2的以外,其余处理的有机质含量均与对照差异显著(P < 0.05),而处理3 mL/m2低于对照4.16%,其他处理的有机质含量分别比对照的高出28.57%~214.28%。且冲施肥和叶面肥在肥料用量为15 mL/m2时,土壤有机质含量达到最大值。说明施鲜海带生物酶解有机液肥能有效促进沙化土壤有机质含量的增加,增加了土壤有效养分,且叶面肥的效果要优于冲施肥,其有机质增加量要比冲施肥高0.46%。
2.2 鲜海带生物酶解有机液肥对沙木蓼生长的影响
沙木蓼地径增长量是随着鲜海带生物酶解有机液肥用量的增加呈先增加后减小的趋势(图 7),冲施肥处理的增长量比叶面肥高25.58%。在冲施肥和叶面肥用量为6 mL/m2时,沙木蓼的地径增长量均达到最高,分别为0.34和0.23 cm;且与最高肥料用量的地径增长量存在显著差异(P < 0.05)。冲施肥用量为3、15 mL/m2的地径增长量均小于对照,分别是对照的83.35%、93.35%,且与6 mL/m2的地径增长量差异显著(P < 0.05),其他处理均不存在显著差异(P>0.05);不同用量的冲施肥对地径增长量的效果依次是6 mL/m2>9 mL/m2>12 mL/m2>CK>15 mL/m2>3 mL/m2。叶面肥所有施肥处理的地径增长量均大于对照,分别比对照高0.10、0.13、0.13、0.04和0.02 cm。方差分析表明,在叶面肥用量为6和9 mL/m2的地径增长量都与用量为15 mL/m2差异显著(P < 0.05),其他处理间不存在显著差异(P>0.05)。不同用量的叶面肥对地径增长量的效果依次是6 mL/m2>9 mL/m2>3 mL/m2>12 mL/m2>15 mL/m2>CK。说明施鲜海带生物酶解有机液肥能够有效地促进沙木蓼地径的增长,用量为6 mL/m2的地径增长量最高,且在两种施肥方式中,冲施肥比叶面肥的地径增长量更高。
![]() 图 7 施肥对沙木蓼地径增长量的影响Figure 7. Effects of fertilization on the growth of ground diameter of Atraphaxis bracteata
图 7 施肥对沙木蓼地径增长量的影响Figure 7. Effects of fertilization on the growth of ground diameter of Atraphaxis bracteata沙木蓼株高增长量随着鲜海带生物酶解有机液肥用量的增加呈先增加后减小的趋势(图 8),冲施肥的株高平均增长量比叶面肥的高71.6%。冲施肥用量为6 mL/m2的株高增长量与对照存在显著差异(P < 0.05),且各处理的株高增长量均高于对照处理,较对照高7.33~23.28 cm;不同冲施肥对沙木蓼株高增长量的效果依次是6 mL/m2>9 mL/m2>12 mL/m2>3 mL/m2>15 mL/m2>CK,各处理平均株高增长量是对照株高增长量的169.88%。叶面肥对沙木蓼株高增长量也具有增长趋势,但不存在显著差异(P>0.05)。说明鲜海带生物酶解有机液肥能够有效地促进沙木蓼株高的增长,当鲜海带生物酶解有机液肥用量为6 mL/m2时的株高增长量最高,且冲施肥比叶面肥效果更优。
![]() 图 8 施肥对沙木蓼株高增长量的影响Figure 8. Effects of fertilization on the tree height increment of Atraphaxis bracteata
图 8 施肥对沙木蓼株高增长量的影响Figure 8. Effects of fertilization on the tree height increment of Atraphaxis bracteata沙木蓼新梢生长量随着鲜海带生物酶解有机液肥用量的增加呈先增加后减小的趋势(图 9),冲施肥的新梢生长量要比叶面肥高17.56%。冲施肥用量为6 mL/m2的新梢生长量最大,与对照、12和15 mL/m2处理差异显著(P < 0.05),与其他处理不存在显著差异(P>0.05)且不同施肥量处理的新梢生长量均比对照高9.39~31.73 cm。叶面肥用量为6 mL/m2的新梢生长量最大,与对照差异显著(P < 0.05),其他处理均不具有显著性(P>0.05),各处理的新梢生长量分别比对照高13.77~33.85 cm。不同处理的冲施肥和叶面肥对沙木蓼新梢生长量的效果依次是6 mL/m2>3 mL/m2>9 mL/m2>12 mL/m2>15 mL/m2>CK。说明鲜海带生物酶解有机液肥能够有效地促进沙木蓼新梢的增长,当鲜海带生物酶解有机液肥用量为6 mL/m2时,新梢生长量最高,且冲施肥比叶面肥效果更好。
![]() 图 9 施肥对沙木蓼新梢生长的影响Figure 9. Effects of fertilization on the growth of Atraphaxis bracteata new shoots
图 9 施肥对沙木蓼新梢生长的影响Figure 9. Effects of fertilization on the growth of Atraphaxis bracteata new shoots2.3 鲜海带生物酶解有机液肥对沙木蓼周围物种相对丰度的影响
施肥对沙木蓼周围植物群落组成具有明显的影响。施冲施肥后物种数增加,随着冲施肥用量的增加而增加(表 4),增加物种数一般为2~4种;按照物种相对丰度≥10%为优势种群,冲施肥研究区主要优势种为杨柴(Hedysarum fruticosum)、白蒿(Artimisiae sphaerocephala)、油蒿(Artemisia ordosica),这些优势种的相对丰度值随着施肥量的增加而减小,相对丰度值≥5%的伴生植物冰草(Agropyron cristatum)的物种丰度随着施肥量的增加而增大,而主要优势种白蒿和油蒿在施肥量为6 mL/m2下消失,也有一些植物如芦苇(Phragmites australis)、狗尾巴草(Setaria viridis)、砂蓝刺头(Echinops gmelini)、沙蓬(Agriophyllum squarrosum)、角蒿(Incarvillea sinensis)会在施肥量为3 mL/m2时,在群落中出现。
表 4 不同梯度冲施肥处理的物种相对丰度Table 4. Relative abundance of species in different gradient treatments of flush fertilizer物种名称Species name CK 3 mL/m2 6 mL/m2 9 mL/m2 12 mL/m2 15 mL/m2 杨柴Hedysarum fruticosum 44.84 23.88 22.09 19.06 16.08 14.53 白蒿Artimisiae sphaerocephala 25.82 21.70 0 6.36 22.99 39.40 油蒿Artemisia ordosica 20.92 9.20 0 10.00 0 0 冰草Agropyron cristatum 8.42 0 22.42 24.57 0 9.53 芦苇Phragmites australis 0 14.48 6.30 0 0 12.15 狗尾草Setaria viridis 0 17.70 19.76 18.66 15.54 12.37 砂蓝刺头Echinops gmelini 0 13.49 0 0 0 6.36 沙蓬Agriophyllum squarrosum 0 14.56 5.39 0 0 0 角蒿Incarvillea sinensis 0 0 0 0 4.53 0 施叶面肥后各处理物种数均也增加(表 5),随着叶面肥用量的增加,增加物种数一般为2~5种;根据物种相对丰度,叶面肥的研究区主要优势种为杨柴和油蒿,随着施肥量的增大,主要优势种的相对丰度值减小,群落中增加其他物种,如冰草、芦苇、狗尾巴草和针茅(Stipa capillata)等。
表 5 不同梯度叶面肥处理的物种相对丰度Table 5. Relative abundance of species in different gradient treatments of foliar fertilizer物种名称Species name CK 2 mL/m2 4 mL/m2 6 mL/m2 8 mL/m2 10 mL/m2 杨柴Hedysarum fruticosum 58.33 29.67 15.06 18.62 13.47 12.75 白蒿Artimisiae sphaerocephala 0 34.71 7.10 31.36 27.99 19.52 油蒿Artemisia ordosica 41.67 6.56 8.87 23.11 11.92 17.27 冰草Agropyron cristatum 0 24.56 21.89 21.07 22.57 16.53 芦苇Phragmites australias 0 0 12.32 0 6.57 12.52 狗尾草Setaria viridis 0 9.56 11.42 14.18 15.94 7.28 针茅Stipa capillata 0 0 10.59 0 0 6.40 花棒Hedysarum scoparium 0 0 0 10.61 0 0 2.4 鲜海带生物酶解有机液肥对沙木蓼群落物种多样性的影响
沙木蓼群落Margalef物种丰富度指数在不同施肥方式下具有不同的变化趋势(图 10),冲施肥对Margalef物种丰富度指数的影响不显著(P>0.05),但用量为3 mL/m2的Margalef物种丰富度指数与对照、用量为12 mL/m2有显著差异(P0.05);冲施肥各处理的Margalef物种丰富度指数分别是对照的209.98%、144.37%、138.81%、119.02%和157.68%。叶面肥对Margalef物种丰富度指数具有显著影响(P < 0.05),随着叶面肥用量的增加而增加,除了用量为9和12 mL/m2处理间无显著差异,其他处理均差异显著(P < 0.05);叶面肥各处理的Margalef物种丰富度指数分别是对照的115.83%、137.81%、164.09%、193.11%和221.37%。
![]() 9 施肥对沙木蓼群落物种多样性的影响不同字母表示处理间差异显著(P<0.05)。9. Effects of fertilization on the species diversity of Atraphaxis bracteataDifferent letters indicate significant difference among treatments at P < 0.05 level.
9 施肥对沙木蓼群落物种多样性的影响不同字母表示处理间差异显著(P<0.05)。9. Effects of fertilization on the species diversity of Atraphaxis bracteataDifferent letters indicate significant difference among treatments at P < 0.05 level.不同的施肥方式对沙木蓼群落Simpson优势度指数的影响各异,冲施肥对Simpson优势度指数的影响显著(P < 0.05),随着施肥量的增加呈先增加后减小的趋势,在施肥量为6 mL/m2的Simpson优势度指数达到最大,与对照和其余处理间差异显著(P < 0.05);冲施肥各处理的Simpson优势度指数分别是对照的132.80%、142.43%、126.93%、106.16%和124.69%。叶面肥对Simpson优势度指数影响不显著(P>0.05),与冲施肥Simpson优势度指数变化趋势相反,在施肥量为6 mL/m2时Simpson优势度指数最低。
不同的施肥方式对沙木蓼群落Shannon-Wiener多样性指数具有不同的影响,但冲施肥和叶面肥对群落Shannon-Wiener多样性指数的影响均不显著(P>0.05)。冲施肥的Shannon-Wiener多样性指数除了用量为12 mL/m2的处理以外,其他各处理均与对照有显著差异(P < 0.05),而叶面肥对照处理与高用量(12、15 mL/m2)的处理差异显著(P < 0.05),其他处理间无显著差异(P>0.05)。
不同的施肥方式对沙木蓼群落Pielou均匀度指数影响各异,冲施肥和叶面肥对群落Pielou均匀度指数的影响都不显著(P>0.05)。冲施肥的Pielou均匀度指数在用量为12 mL/m2时与对照、用量为6 mL/m2有显著差异(P < 0.05);叶面肥的Pielou均匀度指数仅在用量为3、6 mL/m2之间形成显著差异(P < 0.05)。
3. 讨论
3.1 鲜海带生物酶解有机液肥对土壤理化性质的影响
本研究中,鲜海带生物酶解有机液肥改变了植物的营养循环过程,并对土壤的理化性质产生一定的影响。土壤水分随着鲜海带生物酶解有机液肥的增加而增加,这与其他人所研究的土壤水分随着氮素水平增加而降低[22]不一致,主要是因为施加的氮素进入土壤后只有溶解到土壤水溶液中转化为离子状态才能被植物根系有效吸收,且氮肥含量越高,植被因生理过程对水分的需求越大,土壤含水率越低,而鲜海带生物酶解有机液肥是液体肥料,消耗土壤中的水分较少,可以使养分被植物根系吸收,并且鲜海带生物酶解有机液肥具有保水保肥的作用,所以增加了荒漠区土壤的含水量。并且本次研究发现,冲施肥土壤含水率较面肥研究区的土壤含水率稳定,变化幅度在0.1%~0.4%之间,说明冲施肥比叶面肥更有利于土壤保水,出现这样的结果原因可能有二:一、冲施肥与叶面肥的主要成分不同,冲施肥中含有海带纤维,据研究海带纤维的本质具有阻燃、电磁波屏蔽效应等功能以及良好的纺织加工性能[23],那么海带纤维也可能会在土壤中像保水剂一样抑制水分蒸发,提高土壤含水量,减少可溶性养分的损失,达到保水保肥的效果;二、因为冲施肥利用灌溉的方式,直接将有机液肥漫灌渗入土壤,沙地孔隙度大,入渗快,一部分被根部吸收,一部分直接被土壤吸收,使其更好的发挥保水保肥的作用;而叶面肥利用喷施的方式,先到植物再到土壤,液肥大部分都被沙木蓼叶片吸收,只有少部分进入土壤,导致叶面肥研究区土壤浅层含水率没有冲施肥研究区的高,这与赵岩等研究结果一致。同时,在高肥(12、15 mL/m2)的作用下,土壤水分也会降低,原因可能是由于荒漠地区地表植被覆盖率低、土壤水分蒸发快。而较低的含水率使植物不能有效的吸收养分,枯落物分解率减缓,生态系统中的养分循环速率减弱[24],鲜海带生物酶解有机液肥中各种营养元素在土壤中累积,造成土壤全氮含量随着肥料用量的增大而增加。另外,土壤中氮含量增加会缓解该研究区土壤氮素的限制作用,氮素与有机质结合会降低土壤中的C/N,其在影响植被凋落物返还量的同时,也促进有机质的增加[25-26]。
3.2 鲜海带生物酶解有机液肥对沙木蓼生长的影响
增加土壤养分会改变植物各方面的适应对策,而生长和形态学适应是一种最基本的机制[27-28]。本试验中随着鲜海带生物酶解有机液肥用量的增加,沙木蓼地径、株高、新梢增长量均呈现先增加后减小的趋势,在用量为6 mL/m2时,生长量达到最大,冲施肥和叶面肥的地径、株高、新梢生长量分别是0.34和0.23 cm、43.21和21.8 cm、75.27和66.71 cm,之后生长量随着肥料用量的增大而减小,并且冲施肥的效果更优。施肥能增加荒漠化土壤中的养分,提高土壤肥力,推动土壤养分循环,促进植物营养的吸收[25],从而使沙木蓼的生长量增加。但是随着肥料用量的增大,高肥水平的抑制效果明显。不同施肥方式中肥料用量对沙木蓼生长的影响出现差异的主要原因可能是:在毛乌素沙地,植物生长受营养元素限制时,鲜海带生物酶解有机液肥的添加会促进植物生长;在营养元素饱和或过量时,鲜海带生物酶解有机液肥对植物生长无影响或减缓植物生长,导致沙木蓼在高用量肥(12、15 mL/m2)的增长量降低。
3.3 鲜海带生物酶解有机液肥对沙木蓼群落物种丰富度以及物种多样性的影响
物种丰富度和物种多样性是生态环境稳定的重要标志[29]。施肥不仅可改善荒漠化土壤理化性质,促进植物生长,也是恢复和提高土壤肥力,改变植物群落变化的有效手段。有研究表明,在高寒典型草地施氮、磷肥时,物种丰富度随着施肥水平的增加显著下降[30],而本研究结果显示施肥使物种丰富度增加,可能是由于该地区物种丰富度基数低,植被群落间竞争引起的负效应要远远低于施肥驱动增加的丰富度[31],也可能是因为鲜海带生物酶解有机液肥,富含植物所需的生长刺激素、活性营养物质和微量元素,而不是单单添加氮、磷等元素。同时,在沙木蓼群落中的杨柴、油蒿和白蒿对鲜海带生物酶解有机液肥的响应敏感。在荒漠化治理进行植物措施时,对荒漠化土壤进行鲜海带生物酶解有机液肥添加不仅可以改善其植被的物种组成,还能提高沙木蓼的高度。
目前,关于营养添加对植物群落物种多样性影响的研究很多,而对于荒漠植物群落物种多样性影响的研究比较少。从本研究中可以看出,不同的鲜海带生物酶解有机液肥处理对沙木蓼植物群落多样性的影响各异。鲜海带生物酶解有机液肥肥增加了沙木蓼群落的Margalef物种丰富度指数、Simpson优势度指数和Shannon-Wiener多样性指数,冲施肥和叶面肥各处理的平均Margalef物种丰富度指数比对照的增加了53.97%、66.44%;冲施肥、叶面肥的Simpson优势度指数总体幅度都要比对照的增加了26.6%、22.02%;冲施肥和叶面肥的Shannon-Wiener多样性指数总体幅度都要相比对照增加了60.12%、55.25%。植物群落Pielou均匀度指数随着肥料用量的增加而减小,冲施肥和叶面肥的Pielou均匀度指数总体幅度都要比对照的降低了9.53%、11.67%。总的来说,施肥增加了物种数量和多样性。这与许多研究的施肥对植物群落多样性减少的结果相反。原因可能是鲜海带生物酶解有机液肥减缓了物种间及种内的竞争,相比氮、磷等肥富含的营养元素和活性营养物质多,促使沙木蓼群落的物种多样性增加。植物群落的多样性和它的生境条件与土壤营养有密切的联系,物种多样性是随着土壤资源类型分布的不同而变化[32]。毛乌素沙地土壤有机质含量低,土质瘠薄,鲜海带生物酶解有机液肥使土壤有机质增加,土壤表层结构改善,土壤向良性循环方向发展,从而群落多样性增加。
4. 结论
在毛乌素沙地施鲜海带生物酶解有机液肥不仅有效的增加了土壤的含水率、土壤全氮和有机质的含量,而且可以促进沙木蓼的生长,其地径、株高和新梢增长量都随着施肥量的增加呈先增加后减小的趋势。同时,增加了沙木蓼群落的物种组成、物种丰富度、Margalef物种丰富度指数、Simpson优势度指数和Shannon-Wiener多样性指数,降低了Pielou均匀度指数,但总体增加了沙木蓼群落物种多样性。由此可以建议,在干旱区、半干旱区沙地固沙造林时,鉴于沙地土壤瘠薄、保水保肥能力差等特点,可以适当进行鲜海带生物酶解有机液肥的使用,增加荒漠化土壤的养分,提高苗木成活率,促进植物生长,增加群落物种多样性;尤其是对于4年以上生的沙木蓼,为减少其枝条枯死现象,可以使用鲜海带生物酶解有机液肥促进沙木蓼生长,更好的发挥防风固沙的作用,促进植被恢复,改善荒漠化地区的生态环境。
-
![]()
图 1 各因子对不同树种BAI生长的偏依赖关系
Figure 1. Partial dependence of various factors on BAI growth of different tree species
表 1 样地信息统计表(1987)
Table 1 Statistical table of sample plot information (1987)
样地号
Sample plot No.面积/hm2
Area/ha平均胸径
Mean DBH/cm海拔
Altitude/m坡向
Slope aspect坡度
Slope degree/(°)树种组成
Tree species composition1 0.160 18.8 780 东北 Northeast 13 5云3冷1红1枫
5P3A1PK1B2 0.250 22.0 650 西南 Southwest 28 4冷2红2椴1枫1白
4A2PK2T1B1BP3 0.360 15.9 750 东北 Northeast 7 4冷3云1红1椴1枫
4A3P1PK1T1B4 0.312 16.3 780 东北 Northeast 10 5冷3云1红1枫
5A3P1PK1B5 0.200 20.4 634 东北 Northeast 26 3云3红2椴1冷1白
3P3PK2T1A1BP注:P. 红皮云杉 Picea koraiensis; A. 臭冷杉 Abies nephrolepis; PK. 红松 Pinus koraiensis; B. 枫桦 Betula costata; T. 椴树 Tilia amurensis; BP. 白桦 Betula platyphylla.  下载: 导出CSV
下载: 导出CSV
表 2 模型因子属性统计表
Table 2 Statistics table of model factor attributes
类别
Category因子
Factor最小值
Min.最大值
Max.平均值
Mean单木因子 Individual tree factor BA/(10−4 m2) 9.62 2341.40 295.64 竞争因子 Competitive factor BAL/(m2·ha−1) 0 11.10 6.08 CI 0.23 26.28 2.73 SDI 99.03 284.49 214.33 死亡率 Mortality rate/% 0 22 5.1 多样性 Diversity 树木大小多样性
Tree size diversityDIV_Size 2.44 2.87 2.67 U 0 1 0.50 树种多样性
Tree species diversityDIV_SP 1.54 2.05 1.74 M 0 1 0.67 气候 Climate 年平均温度
Average annual temperature/℃1.89 6.25 5.02 年平均降雨量
Average annual rainfall/mm393.90 735.38 588.15 注:BA. 单木胸高断面积;BAL. 大于对象木的林木断面积之和;CI. 对象木的竞争指数;SDI. Reineke的林分密度指数;DIV_Size. 树木大小多样性;U. 大小比数;DIV_SP. 林分树种多样性;M. 混交度。Notes: BA, basal area of individual tree; BAL, sum of basal area of forest tree that is greater than the object trees; CI, competition index of object tree; SDI, stand density index of Reineke; DIV_Size, tree size diversity; U, dominance degree; DIV_SP, tree species diversity of stand; M, mingling degree.
下载: 导出CSV
表 3 主要树种信息统计表
Table 3 Statistical table of main tree species information
树种
Tree species株数
Number of plant胸高断面积
BA/cm2胸高断面积生长量/(cm2·a−1)
BAI/(cm2·year−1)最小值
Min.q25 q50 q75 最大值
Max.平均值
Mean最小值
Min.q25 q50 q75 最大值
Max.平均值
Mean臭冷杉 Abies nephrolepis 2 375 19.6 91.6 243.3 447.7 1 562.3 311.2 0 2.7 6.5 11.7 40.6 8.0 云杉 Picea koraiensis 1 650 20.4 95.0 240.5 526.9 1 870.4 346.1 0 2.0 6.5 14.8 48.7 9.6 红松 Pinus koraiensis 898 9.6 59.4 124.3 411.9 2 341.4 293.8 0 1.5 4.2 11.6 47.6 8.1 椴树 Tilia amurensis 661 12.8 124.7 264.5 481.1 1 676.4 337.3 0 1.1 3.2 7.4 36.5 5.0 枫桦 Betula costata 899 23.8 50.3 78.5 118.3 1 809.6 109.8 0 0.3 1.8 3.9 21.6 2.8 白桦 Betula platyphylla 420 28.3 172.0 268.8 401.2 1 764.6 346.1 0 4.2 7.8 13.4 37.7 9.2 注:q25、q50、q75分别为下四分位数、中位数和上四分位数。Notes: q25, q50, and q75 are the lower quartiles, median, and the upper quartiles, respectively.
下载: 导出CSV
表 4 10折交叉验证结果
Table 4 10-fold blocked cross-validation results
树种 Tree species mtry R2 ± SD RMSE ± SD 臭冷杉 Abies nephrolepis 4 0.663 ± 0.052 3.897 ± 0.263 云杉 Picea koraiensis 4 0.683 ± 0.058 5.272 ± 0.445 红松 Pinus koraiensis 4 0.695 ± 0.064 5.067 ± 0.737 椴树 Tilia amurensis 4 0.459 ± 0.069 3.969 ± 0.294 枫桦 Betula costata 2 0.384 ± 0.075 2.523 ± 0.262 白桦 Betula platyphylla 5 0.568 ± 0.101 4.540 ± 0.719
下载: 导出CSV
表 5 模型因子相对重要性
Table 5 Relative importance of model factors
% 类别
Category因子
Factor臭冷杉
Abies
nephrolepis云杉
Picea
koraiensis红松
Pinus
koraiensis椴树
Tilia
amurensis枫桦
Betula
costata白桦
Betula
platyphylla单木因子
Individual tree factorBA 13.3 13.1 19.2 19.8 22.6 17.5 竞争因子
Competitive factorBAL 10.6 9.6 10.0 14.6 13.8 11.6 CI 12.6 10.1 11.4 5.8 8.0 10.2 SDI 8.3 9.3 8.5 10.6 7.0 14.0 死亡率 Mortality rate 8.9 8.2 4.8 10.6 6.7 9.7 多样性 Diversity 树木大小多样性
Tree size diversityDIV_Size 10.1 12.8 8.0 14.0 6.6 9.7 U 6.8 9.0 11.4 7.4 10.8 5.1 树种多样性
Tree species diversityDIV_SP 8.5 7.8 6.0 5.5 7.1 7.4 M 6.6 5.8 7.3 2.7 4.9 7.1 气候 Climate 年平均降雨量
Average annual rainfall5.1 6.0 5.8 2.0 5.0 2.3 年平均温度
Average annual temperature9.2 8.2 7.6 6.9 7.4 5.4
下载: 导出CSV
-
[1] Schlamadinger B, Bird N, Johns T, et al. A synopsis of land use, land-use change and forestry (lulucf) under the kyoto protocol and marrakech accords[J]. Environmental Science & Policy, 2007, 10(4): 271−282.
[2] 胡松. 湖南栎类天然林断面积生长模型研究[D]. 长沙: 中南林业科技大学, 2019. Hu S. Research on basal area growth model for oak natural forest in Hunan Province [D]. Changsha: Central South University of Forestry & Technology, 2019.
[3] Jevsenak J, Skudnik M. A random forest model for basal area increment predictions from national forest inventory data[J]. Forest Ecology and Management, 2021, 479: 118601.
[4] Jiang X, Huang J, Cheng J, et al. Interspecific variation in growth responses to tree size, competition and climate of western canadian boreal mixed forests[J]. Science of the Total Environment, 2018, 631−632: 1070−1078. doi: 10.1016/j.scitotenv.2018.03.099
[5] 江小雷, 岳静, 张卫国, 等. 生物多样性, 生态系统功能与时空尺度[J]. 草业学报, 2010, 19(1): 219−225. Jiang X L, Yue J, Zhang W G, et al. Biodiversity, ecosystem functioning and spatio-temporal scales[J]. Acta Prataculturae Sinica, 2010, 19(1): 219−225.
[6] Lei X, Wang W, Peng C. Relationships between stand growth and structural diversity in fpruce-dominated forests in new Brunswick, Canada[J]. Canadian Journal of Forest Research, 2009, 39(10): 1835−1847. doi: 10.1139/X09-089
[7] Subedi N, Sharma M. Climate-diameter growth relationships of black spruce and jack pine trees in Boreal Ontario, Canada[J]. Global Change Biology, 2013, 19(2): 505−516. doi: 10.1111/gcb.12033
[8] Chen L, Huang J, Stadt K, et al. Drought explains variation in the radial growth of white spruce in western Canada[J]. Agricultural and Forest Meteorology, 2017, 233: 133−142. doi: 10.1016/j.agrformet.2016.11.012
[9] Cortini F, Filipescu C, Groot A, et al. Regional models of diameter as a function of individual tree attributes, climate and site characteristics for six major tree species in Alberta, Canada[J]. Forests, 2011, 2(4): 814−831. doi: 10.3390/f2040814
[10] Ashraf M, Zhao Z, Bourque C, et al. Integrating biophysical controls in forest growth and yield predictions with artificial intelligence technology[J]. Canadian Journal of Forest Research, 2013, 43(12): 1162−1171. doi: 10.1139/cjfr-2013-0090
[11] 方匡南, 吴见彬, 朱建平, 等. 随机森林方法研究综述[J]. 统计与信息论坛, 2011, 26(3): 32−38. Fang K N, Wu J B, Zhu J P, et al. A review of technologies on random forests[J]. Journal of Statistics and Information, 2011, 26(3): 32−38.
[12] 高若楠, 苏喜友, 谢阳生, 等. 基于随机森林的杉木适生性预测研究[J]. 北京林业大学学报, 2017, 39(12): 36−43. Gao R N, Su X Y, Xie Y S, et al. Prediction of adaptability of Cunninghamia lanceolata based on random forest[J]. Journal of Beijing Forestry University, 2017, 39(12): 36−43.
[13] 董灵波, 梁凯富, 张一帆, 等. 基于Landsat 8时间序列数据的翠岗林场森林类型划分[J]. 应用生态学报, 2022, 33(9): 2339−2346. Dong L B, Liang K F, Zhang Y F, et al. Classification of forest types in Cuigang Forest Farm based on time series data of Landsat 8[J]. Chinese Journal of Applied Ecology, 2022, 33(9): 2339−2346.
[14] 徐丽, 欧阳勋志, 潘萍, 等. 基于GF-1 WFV与MODIS时空融合的南方森林植被类型识别[J]. 应用生态学报, 2022, 33(7): 1948−1956. Xu L, Ouyang X Z, Pan P, et al. Identification of forest vegetation types in southern China based on spatio- temporal fusion of GF-1 WFV and MODlS data[J]. Chinese Journal of Applied Ecology, 2022, 33(7): 1948−1956.
[15] 孙铭辰, 姜立春. 基于机器学习算法的樟子松立木材积预测[J]. 南京林业大学学报(自然科学版), 2023, 47(1): 31−37. Sun M C, Jiang L C. Standing volume prediction of Pinus sylvestris var. mongolica based on machine learning algorithm[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2023, 47(1): 31−37.
[16] Lh R. Perfecting a stand-density tndex for even-aged forests [J]. Journal of Agricultural Research, 1933, 46: 627−638.
[17] Hegyi F. A simulation model for managing Jack-pine stands simulation[J]. Royalcoll. For, Res. Notes, 1974, 30: 74−90.
[18] 惠刚盈, 胡艳波. 混交林树种空间隔离程度表达方式的研究[J]. 林业科学研究, 2001, 14(1): 23−27. doi: 10.3321/j.issn:1001-1498.2001.01.004 Hui G Y, Hu Y B. Measuring species spatial lsolation in mixed forests[J]. Forest Research, 2001, 14(1): 23−27. doi: 10.3321/j.issn:1001-1498.2001.01.004
[19] 惠刚盈. 一个新的林分空间结构参数: 大小比数[J]. 林业科学研究, 1999, 12(1): 4−9. Hui G Y. A new parameter for stand spatial structure: neighbourhood comparison[J]. Forest Research, 1999, 12(1): 4−9.
[20] Breiman L. Random forests[J]. Machine Learning, 2001, 45(1): 5−32. doi: 10.1023/A:1010933404324
[21] Arlot S, Celisse A. A survey of cross-validation procedures for model selection[J]. Statistics Surveys, 2010, 4: 40−79.
[22] Willmott C J, Matsuura K. Advantages of the mean absolute error (mae) over the root mean square error (rmse) in assessing average model performance[J]. Climate Research, 2005, 30(1): 79−82.
[23] 董雪婷, 张静, 张志东, 等. 树种相互作用、林分密度和树木大小对华北落叶松生产力的影响[J]. 应用生态学报, 2021, 32(8): 2722−2728. Dong X T, Zhang J, Zhang Z D, et al. Effects of tree species interaction, stand density, and tree size on the productivity of Larix principis-rupprechtii[J]. Chinese Journal of Applied Ecology, 2021, 32(8): 2722−2728.
[24] 吕沅杭, 伊利启, 王儒林, 等. 基于空间结构参数的大兴安岭天然落叶松单木直径生长模型[J]. 林业科学研究, 2021, 34(2): 81−91. Lü Y H, Yi L Q, Wang R L, et al. Diameter growth model using spatial structure parameters of natural Larix gmelinii stand in Daxing’anling Mountains, northeast China[J]. Forest Research, 2021, 34(2): 81−91.
[25] Lin S, Li Y, Li Y H, et al. Influence of tree size, local forest structure, topography, and soil resource availability on plantation growth in Qinghai Province, China[J]. Ecological Indicators, 2021, 120: 106957.
[26] Adame P, Hynynen J, Canellas I, et al. Individual-tree diameter growth model for rebollo oak (Quercus pyrenaica Willd.) coppices[J]. Forest Ecology and Management, 2008, 255(3): 1011−1022.
[27] 任玫玫, 杨华. 长白山云冷杉林优势树种的竞争[J]. 应用生态学报, 2016, 27(10): 3089−3097. Ren M M, Yang H. Competition among dominant tree species in a natural spruce-fir forest in Changbai Mountain[J]. Chinese Journal of Applied Ecology, 2016, 27(10): 3089−3097.
[28] Liu D, Zhou C, He X, et al. The effect of stand density, biodiversity, and spatial structure on stand basal area increment in natural spruce-fir-broadleaf mixed forests[J]. Forests, 2022, 13(2): 162.
[29] 陈梦飞. 青海东部黄土区青海云杉人工林空间结构对径向生长的影响[D]. 北京: 北京林业大学, 2019. Chen M F. Effects of spatial structure of Picea crassifolia plantationon radial growth in the loess area of eastern Qinghai[D]. Beijing: Beijing Forestry University, 2019.
[30] 吕延杰, 杨华, 张青, 等. 云冷杉天然林林分空间结构对胸径生长量的影响[J]. 北京林业大学学报, 2017, 39(9): 41−47. Lü Y J, Yang H, Zhang Q, et al. Effects of spatial structure on DBH increment of natural spruce-fir forest[J]. Journal of Beijing Forestry University, 2017, 39(9): 41−47.
[31] 余黎, 雷相东, 王雅志, 等. 基于广义可加模型的气候对单木胸径生长的影响研究[J]. 北京林业大学学报, 2014, 36(5): 22−32. Yu L, Lei X D, Wang Y Z, et al. Impact of climate on individual tree radial growth based on generalized additive model[J]. Journal of Beijing Forestry University, 2014, 36(5): 22−32.
[32] 欧强新, 雷相东, 沈琛琛, 等. 基于随机森林算法的落叶松–云冷杉混交林单木胸径生长预测[J]. 北京林业大学学报, 2019, 41(9): 9−19. Ou Q X, Lei X D, Shen C C, et al. Individual tree DBH growth prediction of larch-spruce-fir mixed forests based on random forest algorithm[J]. Journal of Beijing Forestry University, 2019, 41(9): 9−19.
[33] 覃鑫浩. 基于混合效应的兴安落叶松单木断面积生长模型研究[J]. 西北林学院学报, 2022, 37(4): 223−230. Qin X H. Growth model of individual-tree basal area for Larix gmelinii based on mixed effect approach[J]. Journal of Northwest Forestry University, 2022, 37(4): 223−230.
[34] 杜志, 陈振雄, 孟京辉, 等. 基于混合效应的马尾松单木断面积预估模型[J]. 中南林业科技大学学报, 2020, 40(9): 33−40. Du Z, Chen Z X, Meng J H, et al. Prediction model of individual-tree basal area for Pinus massoniana based on mixed effect[J]. Journal of Central South University of Forestry & Technology, 2020, 40(9): 33−40.
[35] Kindermann G E. The development of a simple basal area increment, model[J]. Nature Precedings, 2011, 6: 1−25.
[36] Forrester D. Linking forest growth with stand structure: tree size inequality, tree growth or resource partitioning and the asymmetry of competition[J]. Forest Ecology and Management, 2019, 447: 139−157. doi: 10.1016/j.foreco.2019.05.053
[37] Ni R, Baiketuerhan Y, Zhang C, et al. Analysing structural diversity in two temperate forests in northeastern China[J]. Forest Ecology and Management, 2014, 316: 139−147. doi: 10.1016/j.foreco.2013.10.012
[38] Mcroberts R E, Hahn J T, Hefty G J, et al. Variation in forest inventory field measurements[J]. Canadian Journal of Forest Research, 1994, 24(9): 1766−1770. doi: 10.1139/x94-228
[39] Andreassen K, Tomter S. Basal area growth models for individual trees of Norway spruce, scots pine, birch and other broadleaves in Norway[J]. Forest Ecology and Management, 2003, 180(1): 11−24.
-
期刊类型引用(17)
1. 张佳旺,董希斌,郭奔,刘慧,张莹,任允泽,滕弛,宋梓恺,张雨晨. 天然落叶松林不同演替阶段空间结构参数的多元分布. 东北林业大学学报. 2024(03): 1-9 .  百度学术
百度学术
2. 王迤翾,朱宁华,周光益,袁星明,江岱,董妍妍,颜润芝. 湘西石漠化区湿地松-马褂木人工混交林林分结构及土壤理化性质研究. 广西植物. 2024(03): 477-487 . 百度学术
3. 韩艳英,盛基峰,李垚,大布穷,叶彦辉. 西藏原始林芝云杉林空间结构参数分布特征. 西北林学院学报. 2023(01): 146-152 . 百度学术
4. 淦江,杜虎,宋同清,彭晚霞,曾馥平,黄国勤. 中亚热带喀斯特常绿落叶阔叶林空间结构特征. 西北植物学报. 2023(05): 846-855 . 百度学术
5. 张岚棋,李丽,杨华,谢伊. 基于AHP-CRITIC组合赋权法的长白山天然林空间结构优化调整. 北京林业大学学报. 2023(08): 74-83 . 本站查看
6. 黄露波 ,甄贞 ,赵颖慧 . 基于多源LiDAR的阔叶红松林林分空间结构特征分析. 中南林业科技大学学报. 2023(08): 36-50 . 百度学术
7. 娄明华,杨同辉,王卫兵,毛建方,徐婧,章建红. 四明山黄山松针阔混交林林分空间结构参数多元分布特征. 林业与环境科学. 2023(04): 12-20 . 百度学术
8. 罗建琼,缪宁,薛盼盼,张远东,王晖. 人工-天然混交林研究综述. 陆地生态系统与保护学报. 2023(06): 78-87 . 百度学术
9. 袁星明,朱宁华,周光益,党鹏,尚海. 湘西喀斯特地区42年生湿地松-樟树人工混交林空间结构研究. 中南林业科技大学学报. 2022(04): 49-58 . 百度学术
10. 王志康,祝乐,许晨阳,李艳,耿增超,王强,刘莉丽,秦一郎,杜旭光. 秦岭天然林凋落物去除对土壤团聚体稳定性及细根分布的影响. 生态学报. 2022(13): 5493-5503 . 百度学术
11. 刘磊,萨如拉,高明龙,王子瑞,铁牛. 大兴安岭北部白桦次生林空间结构多元分析. 西部林业科学. 2022(05): 118-126 . 百度学术
12. 和敬渊,王新杰,王开,郭韦韦,刘丽,王福增. 杨桦次生林林分空间结构参数多元分布研究. 北京林业大学学报. 2021(02): 22-33 . 本站查看
13. 薛卫星,郭秋菊,艾训儒,姚兰,朱江,黄阳祥,李玮宜,罗西,刘毅. 鹅掌楸天然林物种组成与林分空间结构特征研究. 林业科学研究. 2021(02): 166-173 . 百度学术
14. 崔玉华,韩有志,张梦弢,杨秀清,赵占合. 不同干扰强度下针阔混交林树种空间格局及种间关联性. 应用生态学报. 2021(06): 2053-2060 . 百度学术
15. 孙明港,王新杰. 长白山地区白桦红松混交林结构特征分析. 西北林学院学报. 2021(05): 18-27 . 百度学术
16. 徐美玲,农明川,欧光龙. 思茅松天然林林分空间结构分析. 西南林业大学学报(自然科学). 2020(03): 122-130 . 百度学术
17. 李燕,刘成功. 天目山天然混交林群落优势树种种群结构与空间格局. 基因组学与应用生物学. 2020(12): 5744-5757 . 百度学术
其他类型引用(12)










计量
- 文章访问数: 338
- HTML全文浏览量: 45
- PDF下载量: 44
- 被引次数: 29
