
Population dynamics of natural regeneration of rattan in secondary lowland rain forest in Hainan Island, southern China
-
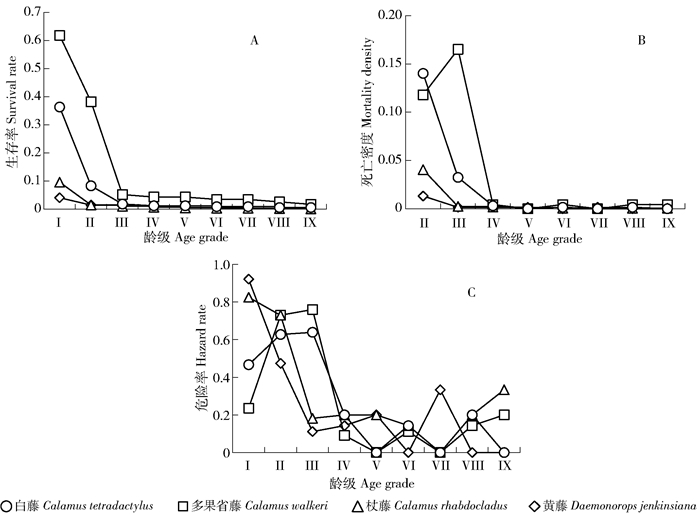
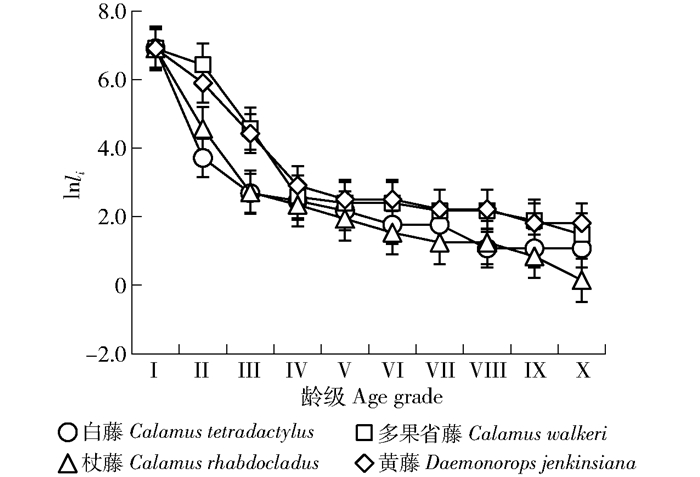
摘要:目的本文对海南岛甘什岭地区热带次生低地雨林棕榈藤的种群数量动态及波动周期进行分析,探究天然更新棕榈藤种群的生长动态规律,为棕榈藤资源保护和后期恢复提供参考。方法以海南岛甘什岭天然更新黄藤、杖藤、多果省藤和白藤种群为研究对象,采用空间代替时间,编制了棕榈藤种群生命表并进行动态分析;幂函数和指数函数对种群存活曲线拟合;3种生存函数预测藤种群生存概率;并通过谱分析来阐述藤种群的周期性波动规律。结果甘什岭天然更新棕榈藤种群以幼苗和幼藤为主,约占总样本的90.1%~97.7%,成年藤株数量少。白藤、杖藤间隔期死亡率和平均存活数随年龄增加逐渐减少,多果省藤和黄藤则为先增大后减小;白藤生命期望在2.5~4.5 m达到最大值,多果省藤、黄藤和杖藤则分布在4.5~6.5 m、6.5~8.5 m、8.5~10.5 m时最大。杖藤存活曲线趋于指数函数(Deevey-Ⅱ,R2=0.840,P=0.000 < 0.01),其余3种藤存活曲线趋于幂函数(Deevey-Ⅲ,R2>0.910,P=0.000 < 0.01)。4种棕榈藤生存率在幼苗期高,随生长呈逐渐降低趋势;除多果省藤外,死亡密度均呈锐减至相对稳定的下降趋势,早期危险率高于后期。谱分析表明,白藤波动振幅差异小,其余3种藤振幅差异大,其中杖藤和多果省藤在4.5~6.5 m时波动振幅最大,黄藤在2.5~4.5 m时波动振幅最大。结论甘什岭地区棕榈藤天然更新种群具有高繁殖和低存活率特点,反映出棕榈藤对该地区生境的适应策略,即通过高繁殖率弥补物种存活能力的不足,以提高物种保存几率。同时在攀援至林冠层过程的棕榈藤存活能力强,相对白藤,其余3种藤更易受外界环境影响。通过改善生境条件并结合人工培育壮苗措施提高更新苗存活率,有利于棕榈藤资源保护。Abstract:ObjectiveThis paper analyzes the population dynamics and undulation periods of rattan populations in Ganshiling tropical secondary lowland rain forest, Hainan Island of southern China, explores the growth dynamic regularity of rattan populations, and provides a theoretical basis for resource protection and later restoration for rattan.MethodThe Daemonorops jenkinsiana (DJ), Calamus rhabdocladus (CR), C. walkeri (CW) and C. tetradactylus (CT) distributing in Ganshiling tropical secondary lowland rainforest were chosen for the objects in this paper. We used the method of substituting space for time to compile and analyze life tables, matching surviving curve for each species according to power function and exponential function.The study also established 3 kinds of function for predicting the survival probability, and analyzed rattan's periodic fluctuation law by spectral method.ResultThe results showed that the main constituents of rattan populations were seedling and young vines which accounted for 90.1%-97.7% of total samples, only a few of adults were tracked in the region. The interval period mortality and average survival numbers of CT and CR decreased with growing. Meanwhile, CW and DJ increased firstly and then decreased. The life expectancy of CT would be maximized at 2.5-4.5 m, CW, DJ and CR of which would be maximized at 4.5-6.5 m, 6.5-8.5 m and 8.5-10.5 m, respectively. The survival curve of CR was more closed to exponential function (Deevey-Ⅱ type, R2=0.840, P=0.000 < 0.01) than power function, and the others were more closed to power function (Deevey-Ⅲ type, R2>0.910, P=0.000 < 0.01). All rattans had a high survival rate at early stage, and then reduced with growing. The others had a sharp decline to steady situation in death density except for CW, with high mortality in early stage than the later. The spectral analysis results indicated that wave amplitudes had significant differences among DJ, CR and CW except of CT, CR and CW had the biggest amplitude fluctuation at 4.5-6.5 m, and DJ had the biggest amplitude fluctuation at 2.5-4.5 m.ConclusionOur results indicated that natural regeneration of rattan in Ganshilin Region had characteristics of high breeding potential and low survival rate, which revealed the adaptive strategy of rattans in this region, in other words, making up insufficient of survival rate by means of high reproductive rate to increase the odds of species preservation. Meanwhile, survival ability is powerful before rattan climbing to the canopy, and the others relative to CT are more affected by external environment. Therefore, the measures increasing regenerate seedling survival rate by improving environment and manual plant strong seedling will be available for the protection of rattan resources.
-
Keywords:
- rattan /
- natural regeneration /
- life table /
- survival curve /
- spectral analysis
-
干旱是限制农林业生产的重要环境因子,国内外学者从分子、生理、生态等方面揭示植物对水分胁迫响应机制的研究逐步深入。探究植物在干旱胁迫下的响应机制,提高植物的抗旱能力,了解植物对干旱胁迫的应对策略,对于农林业的生产极为重要。
胡杨(Populus euphratica)作为荒漠河岸林的主要建群种之一,自然分布于新疆南部、柴达木盆地西部、河西走廊等地,对干旱、盐渍化等逆境具有广谱适应性,逆境相关基因资源丰富[1]。已有研究证实,PebHLH35通过调节拟南芥(Arabidopsis thaliana)的气孔发育和光合作用提高植物抗旱性[2];PeCHYR1基因通过介导植物叶片气孔关闭从而提高杨树耐旱性[3];peu-miR473a在拟南芥中异源表达增强其抗脱水能力从而提高抗旱性[4];peu-miR156j和peu-miR169o均响应脱水和高盐逆境胁迫[5],且peu-miR156j在拟南芥中过表达后可以增强植物耐盐性[6]。
MicroRNAs(miRNAs)是一类由20~25个核苷酸组成的非编码小分子RNA,广泛存在于真核生物体内,通过与靶mRNA相结合从而发挥剪切或沉默靶mRNA的作用,进而在转录后水平调控基因功能。至2017年6月,miRBase数据库中(http://www.miRbase.org/Rlease21)已收录来自动物、植物和病毒的28 645条miRNA,这些miRNA可以在植物的生长发育各个阶段以及植物响应逆境胁迫过程中发挥重要作用,尤其是干旱胁迫方面[7-13]。
已有研究证实,毛果杨(Populus trichocarpa)在过量铜胁迫下,miR1444表达量下调,而靶基因表达量上调;铜缺失情况下,miR1444家族成员表达量均上调,其靶基因下调,同时植物体内锌元素含量明显上升[14];miR1444a通过下调靶基因多酚氧化酶的表达从而参与植物对逆境胁迫的响应[15];miR1444a可能存在Cu-Zn传感模式参与毛果杨对锌胁迫的响应。本文研究的miR1444b和miR1444a均属于miR1444家族,但成熟体序列不同,预测基因启动子区顺式作用元件不相重合。为探究胡杨miR1444b的抗旱机能,本研究克隆得到peu-MIR1444b基因启动子序列,对测序结果进行顺式作用元件分析;克隆peu-MIR1444b基因,转化哥伦比亚野生型拟南芥(Arabidopsis thaliana, Col-0),通过相关生理指标测定初步探究干旱胁迫下植物miR1444b发挥的功能,为深入探讨miR1444b基因提高植物抗旱性的分子机制奠定基础,同时为研究miR1444a基因功能提供参考。
1. 材料与方法
1.1 供试材料
实验中用于基因克隆的胡杨材料为源自中国新疆地区,栽种于北京林业大学苗圃的1年生胡杨苗。拟南芥Col-0种子在超净台中用1%次氯酸钠消毒10 min后,无菌水冲洗5次,点播于1/2MS固体培养基(含有30 g/L蔗糖和6 g/L琼脂)上,4 ℃暗培养2 d后,置于22 ℃,16 h光照、8 h黑暗的环境中生长。将生长7 d的幼苗移栽于培养基质(营养土:草炭土:蛭石=2:2:1)中,于22 ℃,16 h光照、8 h黑暗,55%~65%的相对湿度的温室中生长。
引物合成和DNA测序均在生工生物工程(上海)股份有限公司完成,DNA提取试剂盒、RNA提取试剂盒和TOPO-Blunt Simple连接试剂盒均购自北京艾德莱生物科技有限公司;大肠杆菌感受态TOP10和农杆菌感受态GV3101购自北京博迈德生物技术有限公司;cDNA第1链合成试剂盒(KR201)、DNA琼脂糖凝胶回收试剂盒、质粒提取试剂盒、荧光定量PCR试剂盒和DNA marker连接试剂盒均购自天根生化科技有限公司;T4-DNA连接酶、限制性核酸内切酶BamHI和Sac I、Taq酶均购自宝生物工程有限公司(大连);植物转化表达载体pCAMBIA1301保存于北京林业大学国家工程实验室。其他化学试剂均为国产分析纯,购置于北京蓝弋化工产品有限责任公司和北京中科瑞普生物化工科技有限公司。
1.2 pro-peu-MIR1444b顺式作用元件分析
在NCBI(https://www.ncbi.nlm.nih.gov/)数据库中选取毛果杨MIR1444b基因前约2 000 bp的非编码序列,利用DNAMAN软件设计一对扩增引物,上游引物:5′-CTCATGAGGGTTCTTGTGCTC-3′,下游引物:5′-GTCAAGCCAGCCAATCAATG-3′。用DNA提取试剂盒提取胡杨叶片总DNA,以此为模板在57 ℃的退火温度下进行克隆。回收扩增产物,与载体pCAMBIA1301连接后测序。其结果利用在线软件plantCARE(http://bioinformatics.psb.ugent.be/web-tools/plantcare/html/)分析。
1.3 peu-MIR1444b基因的克隆和遗传转化
从miRBase数据库(www.mirbase.org)中下载MIR1444b茎环前体序列,利用DNAMAN软件设计一对扩增引物,同时在上下游引物的5′端分别加上BamHI(GGATCC)和Sac I(GAGCTC)酶切位点。上游引物为:5′-GGATCCACACTCCAGCTGGG-3′,下游引物为5′-GAGCTCAACTGGTGTCGTGGA-3′。以提取得到的胡杨叶片总DNA为模板,在60 ℃退火温度下克隆得到85 bp的前体片段,记作pre-peu-MIR1444b。连接零背景TOPO-Blunt Simple载体,将双酶切后含有粘性末端的表达载体片段和目的基因片段用T4-DNA连接酶连接,转化测序。
将经过测序验证的pre-peu-MIR1444b连接到含CaMV35S的过表达载体pCAMBIA1301上,转化拟南芥[16]。种子置于含有30 mg/L潮霉素的1/2MS筛选培养基中筛选得到T3代转基因株系(Transgenic lines, TG),进行后续生理指标测定实验。
1.4 转基因植株的RNA水平鉴定和靶基因初步筛选
取生长20 d的野生型和转基因拟南芥叶片液氮研磨,利用RNA提取试剂盒提取总RNA,并采用miRcute miRNA cDNA第1链合成试剂盒和FastKing RT Kit(With gDNase)逆转录试剂盒将RNA分别逆转录为与miRNA和mRNA互补的cDNA第1链。调节cDNA终浓度为80 ng/μL,进行实时荧光定量PCR(qRT-PCR),在RNA水平鉴定miR1444b的相对表达量。
利用软件psRNATarget(http://miRtarbase.mbc.nctu.edu.tw/php/index.php)预测peu-miR1444b在拟南芥数据库(TAIR10)中的靶基因。根据预测得到的靶基因序列设计上下游引物,通过qRT-PCR对其靶向关系进行验证。
1.5 渗透胁迫下拟南芥种子萌发率和幼苗根长的测定
将野生型(Col-0)和过表达的转基因拟南芥种子同时点播于甘露醇浓度为0或250 mmol/L的1/2MS固体培养基上,每个处理做3个重复,每个重复中每个株系点播56粒种子。暗培养2 d后置于光照16 h,黑暗8 h,22 ℃的恒温培养箱中培养,每12 h记录1次萌发率,连续记录4 d。同时选取在1/2MS固体培养基中萌发3 d的幼苗,在超净台中移栽到分别含有0或250 mmol/L甘露醇的1/2MS固体培养基上,测定拟南芥初始根长,竖直培养8 d再次测量根长[17]。同时将1/2MS固体培养基上生长8 d的拟南芥幼苗移栽到培养基质中,覆膜3 d后,于温室中正常培养。
1.6 自然干旱条件下拟南芥光合速率相关指标的测量
选取在同等条件下生长20 d、状况良好且一致的野生型和转基因株系进行自然干旱处理,浇足水后当天做为干旱第1天,设置9个重复。分别在正常浇水、干旱8 d、复水3 d时,利用Licor-6400便携式光合仪测量野生型和转基因株系的光合速率(Pn)、蒸腾速率(Tr)和气孔导度,并根据Pn和Tr计算植物叶片水分利用效率(WUE),其中WUE=Pn/Tr。
1.7 自然干旱条件下拟南芥PSⅡ最大光化学效率和POD活性的测定
选取经过暗适应30 min后的拟南芥叶片,利用双通道PAM-100测量系统Dual-PAM-100测定正常浇水、干旱8 d、复水3 d条件下的PSⅡ最大光化学效率。
以叶片为材料选用愈创木酚法测定正常浇水、干旱8 d和复水3 d时的过氧化物酶(POD)活性[18]。分别称取0.05 g野生型和peu-MIR1444b过表达植株的新鲜叶片,在预冷的研钵中加入0.5 mL预冷的磷酸缓冲液(PBS缓冲液pH7.8,50 mmol/L)冰浴研磨成匀浆,用0.5 mL相同的PBS冲洗研钵2次后转入2 mL离心管中。4 ℃、12 000 g下离心20 min,上清液即为酶粗提取液。在烧杯中加入50 mL PBS(pH6.0,0.2 mol/L)缓冲液和28 μL愈创木酚(2-甲氧基酚),加热搅拌至愈创木酚完全溶解,冷却后加入19 μL 30%的H2O2,混匀后即为反应混合液。取3 mL反应混合液加入40 μL酶粗提取液后立即在光吸收全波长酶标仪TECAN infinite M200中测定OD470值在40 s内的变化。以每分钟OD值变化(升高)0.01为1个酶活性单位(U),POD活性=(ΔA470·Vt)/(0.01WVst),单位为U/g。式中:ΔA470为反应时间内吸光度的变化,W为样品鲜质量(g),t为反应时间(min),Vt为提取酶液总体积(mL),Vs为测定时取用酶液体积(mL)。
1.8 数据分析
本文所有数据均使用SigmaPlot12.5和Excel软件统一处理,且利用SPSS17.0软件进行单因素方差分析(One Way ANOVA)。
2. 结果与分析
2.1 peu-MIR1444b基因启动子克隆与分析
以胡杨基因组为模板,利用PCR克隆得到1 886 bp胡杨MIR1444b基因启动子序列(pro-peu-MIR1444b)后送测,并通过plantCARE网站对测序结果进行比对分析,相似性达到99.61%。pro-peu-MIR1444b中不仅含有TATA-box和CAAT-box等核心元件,还有大量的光响应元件(AE-box, ATCT-motif, GA-motif, GAG-motif, GATA-motif, TCT-motif)和胁迫响应元件(MBS, MRE, TC-rich repeats),这表明miR1444b的表达可能与植物的生长发育、逆境响应以及激素调节等有关(表 1)。
表 1 peu-MIR1444b基因启动子顺式作用元件分析Table 1. Cis-acting element analysis of peu-MIR1444b gene promoter顺式作用元件
Sitename序列
Sequence功能
FunctionAE-box AGAAACAT Part of a module for light response ATCT-motif AATCTAATCT Part of a conserved DNA module involved in light responsiveness CAAT-box CAATT Common cis-acting element in promoter and enhancer regions CAT-box GCCACT Cis-acting regulatory element related to meristem expression GA-motif ATAGATAA Part of a light responsive element MBS CAACTG MYB binding site involved in drought-inducibility MRE AACCTAA MYB binding site involved in light responsiveness TATA-box TATA Core promoter element around -30 of transcription start TC-rich repeats ATTTTCTCCA Cis-acting element involved in defense and stress responsiveness TGA-element AACGAC Auxin-responsive element ARE TGGTTT Cis-acting regulatory element essential for anaerobic induction CGTCA-motif CGTCA Cis-acting regulatory element involved in MeJA-responsiveness GAG-motif AGAGAGTCCC Part of a light responsive element GATA-motif AAGATAAGATT Part of a light responsive element TC-rich repeats GTTTTCTTAC Cis-acting element involved in defense and stress responsiveness TCT-motif TCTTAC Part of a light responsive element TCATG-motif TGACG Cis-acting regulatory element involved in MeJA-responsiveness 2.2 peu-MIR1444b前体克隆及转化
以胡杨基因组DNA为模板克隆得到的peu-MIR1444b基因前体长85 bp,经过多次扩增测序后与毛果杨数据库中的MIR1444b基因进行比对分析,同源性达到98.82%(图 1),存在一个单核苷酸多态性(SNP)位点,MIR1444b在胡杨和毛果杨中保守。转化拟南芥后经DNA初步检测得到转基因株系。
![]() 图 1 胡杨MIR1444b前体与phytozome上预测的ptr-MIR1444b序列比对Figure 1. Comparison of P. euphratica MIR1444b precursor with that of P. trichocarpa
图 1 胡杨MIR1444b前体与phytozome上预测的ptr-MIR1444b序列比对Figure 1. Comparison of P. euphratica MIR1444b precursor with that of P. trichocarpa2.3 转基因株系的RNA水平验证
利用RNA提取试剂盒提取野生型和转基因株系RNA,加PolyA尾后进行反转录得到cDNA序列,利用miR1444b成熟体作为上游引物进行qRT-PCR[19]。结果表明,整合到拟南芥基因组中的peu-MIR1444b基因在转基因株系TG-1、TG-2、TG-5中表达水平最高,选取TG-1、TG-2、TG-5株系进行后续研究。
2.4 peu-miR1444b在拟南芥中的靶基因预测及表达分析
在软件psRNATarget中预测peu-miR1444b存在于拟南芥数据库中的靶基因(表 2),并通过qRT-PCR进行初步鉴定。结果显示,转基因株系中AT3G14570.2表达量显著下降(图 2),而AT1G23440.2和AT1G18830.1的表达量在转基因株系中没有出现明显的下降趋势。这初步表明葡聚糖合成酶Ⅳ基因(AT3G14570.2)作为miR1444b在拟南芥中的靶基因,受到peu-miR1444的负调控,而AT1G23440.2和AT1G18830.1的靶向关系有待进一步验证。
表 2 拟南芥中预测的靶基因Table 2. Potential target genes in A. thaliana靶基因名称
Target gene互补序列
Complementary sequence期望值
Expectation翻译产物
Translation productAT1G23440.2 GAACGUUGAUAGAGUGUGAU 3 类焦谷氨酰肽酶Ⅰ
Pyroglutamyl peptidase I-like, ATGSL04AT3G14570.2 GAGCUUUGACAGAAUGUGGA 3 葡聚糖合成酶Ⅳ
Glucan synthase-likeⅣAT1G18830.1 GAUAGUUCACCGAAUGUGAA 3 类WD40重复超家族蛋白
Transducin/WD40 repeat-like superfamily protein![]() 图 2 预测的miR1444b靶基因AT3G14570.2相对表达量分析Col-0为野生型,TG-1、TG-2和TG-5分别为转基因株系1、2和5号。每个实验设置3个重复,每个数值均为3组实验数据的平均值,误差线代表标准差。不同小写字母表示P < 0.05水平差异显著。下同。Figure 2. Analysis of relative gene expression of predicted miR1444b target gene AT3G14570.2Col-0 is wild type, TG-1, TG-2 and TG-5 are transgenic lines 1, 2 and 5, respectively. Three replicates were set up for each independent experiment, each of which was an average of three experimental sets of data and error bars representing standard deviation. Different lowercase letters indicate significant difference at P < 0.05 level. The same below.
图 2 预测的miR1444b靶基因AT3G14570.2相对表达量分析Col-0为野生型,TG-1、TG-2和TG-5分别为转基因株系1、2和5号。每个实验设置3个重复,每个数值均为3组实验数据的平均值,误差线代表标准差。不同小写字母表示P < 0.05水平差异显著。下同。Figure 2. Analysis of relative gene expression of predicted miR1444b target gene AT3G14570.2Col-0 is wild type, TG-1, TG-2 and TG-5 are transgenic lines 1, 2 and 5, respectively. Three replicates were set up for each independent experiment, each of which was an average of three experimental sets of data and error bars representing standard deviation. Different lowercase letters indicate significant difference at P < 0.05 level. The same below.2.5 干旱胁迫下植株萌发率、根长等测定
分别将野生型和转基因拟南芥种子点播到甘露醇浓度为0和250 mmol/L的1/2MS固体培养基中,统计种子萌发率和根的生长情况。结果表明,1/2MS培养基生长的野生型和转基因拟南芥萌发速率和根长均无明显差异(图 3A、3C),且种子在48 h内全部萌发。在甘露醇浓度为250 mmol/L的1/2 MS固体培养基中,野生型拟南芥在36和48 h萌发率显著高于转基因拟南芥。最终野生型拟南芥萌发率为71.43%,而转基因株系TG-1、TG-2和TG-5最终萌发率分别为81.55%、86.31%和90.48%。其中TG-2和TG-5均显著高于野生型植株20.83%和26.67%,TG-1萌发率高于野生型14.17%,差异不显著(图 3B)。在甘露醇模拟的干旱环境中,转基因拟南芥TG-1、TG-2和TG-5根长分别高于野生型43.30%、56.83%、52.60%,达到极显著水平(图 3D)。
![]() 图 3 拟南芥在甘露醇处理下的萌发率和根长统计分析A.1/2MS培养基上种子萌发率;B.1/2MS+250 mmol/L甘露醇培养基上种子萌发率;C.1/2MS培养基上生长8 d的根长;D.1/2MS培养基+250 mmol/L甘露醇培养基上生长8 d的根长。每个实验设置3个重复,每个数值均为3组实验数据的平均值,误差线代表标准差;下同。Figure 3. Analysis of A. thaliana germination rate and root length under mannitol treatmentA, seed germination rate on 1/2 MS medium; B, seed germination rate on 1/2 MS medium with 250 mmol/L mannitol; C, root length after 8 days of growth on 1/2 MS medium; D, root length after 8 days of growth on 1/2 MS with 250 mmol/L mannitol. Three replicates were set up for each independent experiment, each of which was an average of three experimental sets of data and error bars represented standard deviation; the same below.
图 3 拟南芥在甘露醇处理下的萌发率和根长统计分析A.1/2MS培养基上种子萌发率;B.1/2MS+250 mmol/L甘露醇培养基上种子萌发率;C.1/2MS培养基上生长8 d的根长;D.1/2MS培养基+250 mmol/L甘露醇培养基上生长8 d的根长。每个实验设置3个重复,每个数值均为3组实验数据的平均值,误差线代表标准差;下同。Figure 3. Analysis of A. thaliana germination rate and root length under mannitol treatmentA, seed germination rate on 1/2 MS medium; B, seed germination rate on 1/2 MS medium with 250 mmol/L mannitol; C, root length after 8 days of growth on 1/2 MS medium; D, root length after 8 days of growth on 1/2 MS with 250 mmol/L mannitol. Three replicates were set up for each independent experiment, each of which was an average of three experimental sets of data and error bars represented standard deviation; the same below.2.6 干旱胁迫下植株光合速率测定
将温室正常培养20 d的拟南芥幼苗(Col-0、TG-1、TG-2、TG-5)做自然干旱处理,浇足水后当天作为干旱第1天,利用Li-6400便携式光合仪分别检测正常浇水、干旱8 d和复水3 d后的植株光合速率并拍照。结果表明,干旱8 d后的转基因植株长势优于野生型,复水后转基因植株恢复能力更强(图 4A)。
![]() 图 4 自然干旱和复水后拟南芥光合指标测定A.干旱以及复水之后拟南芥的生长状况;B~D.正常浇水、干旱8 d、复水3 d野生型和转基因拟南芥的光合速率、气孔导度、叶片水分利用效率的变化。Figure 4. Measurement of A. thaliana photosynthesis indexes after natural soil drought and rewatered conditionsA, Growth status of A. thaliana after drought and rewatered conditions; B-D, measurement of photosynthetic rate, stomatal conductance and water use efficiency of wild-type and transgenic A. thaliana under normal watering, drought for 8 days and rewatered for 3 days.
图 4 自然干旱和复水后拟南芥光合指标测定A.干旱以及复水之后拟南芥的生长状况;B~D.正常浇水、干旱8 d、复水3 d野生型和转基因拟南芥的光合速率、气孔导度、叶片水分利用效率的变化。Figure 4. Measurement of A. thaliana photosynthesis indexes after natural soil drought and rewatered conditionsA, Growth status of A. thaliana after drought and rewatered conditions; B-D, measurement of photosynthetic rate, stomatal conductance and water use efficiency of wild-type and transgenic A. thaliana under normal watering, drought for 8 days and rewatered for 3 days.Licor-6400便携式光合仪测定结果表明,正常浇水条件下,转基因植株较野生型光合能力更强,其中TG-5的光合速率和气孔导度分别显著高于野生型8.51%、48.01%。转基因株系TG-1、TG-2和TG-5在干旱8 d时,光合速率分别高于野生型17.13%、28.80%和74.26%,复水3 d后,光合速率分别高于野生型77.47%、69.32%和118.73%,均存在显著差异(图 4B)。TG-1、TG-2和TG-5干旱8 d时气孔导度与Col-0差异不显著,但复水3 d后,气孔导度分别显著高于野生型63.98%、39.93%和90.29%,差异达到显著水平(图 4C)。
正常浇水条件下,TG-2和TG-5叶片水分利用效率极显著高于野生型91.91%、63.66%,TG-1与Col-0差异不显著。干旱8 d和复水3 d时,TG-2和TG-5叶片水分利用效率均显著高于Col-0,差异显著。TG-1在干旱8 d时叶片水分利用效率显著高于野生型48.66%,复水3 d时与野生型差异不显著。这些数据进一步说明,转基因株系在正常浇水、干旱8 d和复水3 d条件下具有更高的光合速率。
2.7 PSⅡ最大光化学效率测定
利用双通道PAM-100测量系统Dual-PAM-100测定野生型和转基因拟南芥在正常浇水、干旱8 d、复水3 d时的PSⅡ最大光化学效率。结果显示,正常浇水情况下,PSⅡ最大光化学效率在野生型和转基因拟南芥中无明显差异。转基因拟南芥在干旱处理8 d和复水3 d后PSⅡ最大光化学效率均显著高于野生型(图 5A)。
![]() 图 5 PSⅡ最大光化学效率和POD活性测定A.自然干旱和复水后拟南芥PSⅡ最大光化学效率;B.自然干旱和复水后过氧化物酶活性。Figure 5. PSⅡ maximum photochemical efficiency and POD activity determinationA, PSⅡ maximum photosynthetic efficiency of A. thaliana after natural drought and rewatering; B, peroxidase activity of A. thaliana after natural drought and rewatering.
图 5 PSⅡ最大光化学效率和POD活性测定A.自然干旱和复水后拟南芥PSⅡ最大光化学效率;B.自然干旱和复水后过氧化物酶活性。Figure 5. PSⅡ maximum photochemical efficiency and POD activity determinationA, PSⅡ maximum photosynthetic efficiency of A. thaliana after natural drought and rewatering; B, peroxidase activity of A. thaliana after natural drought and rewatering.方差分析显示,TG-2和TG-5在复水3 d后与正常浇水条件相比,PSⅡ最大光化学效率无显著差异。这表明转基因拟南芥具有较强的适应短期干旱能力。
2.8 干旱胁迫下植株POD活性的测定
过氧化物酶(Peroxidase,POD)是植物体响应干旱胁迫时产生的活性氧代谢清除系统成员之一,表征植物在逆境胁迫下的抗氧化能力。从图 5B可以看出,正常情况下野生型和转基因株系的POD活性差异不显著。干旱8 d后,Col-0、TG-1、TG-2和TG-5的POD活性增加百分比分别为180.89%、484.51%、462.74%和385.12%,转基因株系的POD活性增加百分比显著高于野生型。复水3 d后,转基因株系TG-1、TG-2和TG-5较干旱8 d时的POD活性下降百分比分别为28.29%、9.42%和29.56%,而野生型POD活性继续上升。结果表明, 转基因拟南芥可以在干旱胁迫条件下快速产生过氧化物酶,清除体内过量的活性氧,保护植株免受干旱胁迫的损害,而野生型拟南芥响应较慢。
3. 结论与讨论
miR1444是木本植物特异基因家族的一员,首先发现于毛果杨基因组中。随后在胡杨、北京杨(Populus×beijngensis)、川杨(P.szechwanica)、旱柳(Salix matsudana)等杨柳科(Salicaceae)植物中经过高通量测序也检测到miR1444的存在[11, 20-22]。miR1444基因家族的研究主要集中在逆境胁迫对其表达量的影响方面,尤其是铜、锌等金属胁迫,在干旱、盐等非生物胁迫中研究甚少。研究表明,胡杨脱水4~6 h后,miR1444a明显上调。ptr-miR1444b与ptr-miR1444a存在序列分歧,在毛果杨中分别调控不同的多酚氧化酶基因[23]。因此本文从胡杨基因组中克隆得到MIR1444b基因并转化模式植物拟南芥,研究其在拟南芥中的抗旱生理效应。
启动子中包含的顺式作用元件通过调控相关基因的表达,使植物适应不同的逆境胁迫。顺式作用元件ARE在厌氧诱导过程中起重要作用[24];ABA应答因子(ABRE)为干旱响应元件[25];ABF2和ABF3响应盐胁迫[26]。本文从胡杨基因组中克隆得到胡杨MIR1444b启动子序列,测序后利用plantCARE在线软件分析其顺式作用元件。分析结果显示,启动子区除包含光响应元件外,还含有受干旱诱导的MYB结合位点MBS,防卫和胁迫相关的TC-rich repeats元件。初步猜测peu-miR1444b的表达受干旱胁迫诱导,为MIR1444b基因功能研究奠定了理论基础。
利用生物信息学方法预测miR1444b在拟南芥中的靶基因并对其进行初步验证。验证结果说明, 编码葡聚糖合成酶Ⅳ的AT3G14570.2基因是miR1444b在拟南芥中的靶基因。葡聚糖合成酶Ⅳ具有葡聚糖合成酶活性,可催化尿苷二磷酸葡萄糖(uridine diphosphate glucose,UDPG)合成葡聚糖[27]。蔗糖合成酶可催化可逆反应:蔗糖+UDP⇌果糖+UDPG,催化活性偏向于蔗糖裂解反应[28]。因此植物体内UDPG可以在由UDP焦磷酸化酶催化UTP和葡萄糖-1-磷酸合成,也可以来源于蔗糖分解。转基因株系中MIR1444b基因的过表达,降低了其靶基因葡聚糖合成酶Ⅳ的表达水平,葡聚糖合成被抑制。故推测miR1444b对葡聚糖合成酶Ⅳ的负调控,可能导致植物积累UDPG,蔗糖合成酶水解活性被抑制,植物渗透调节能力增加,peu-MIR1444b过表达株系耐旱性提高。但其对干旱环境的响应机制仍需进一步验证。
本研究将胡杨基因组中克隆得到的peu-MIR1444b基因转化到拟南芥基因组中。在甘露醇模拟的干旱条件下,转基因植株虽萌发时间较晚,但最终萌发率和根长均显著高于野生型。这说明转基因植株可以较快响应干旱胁迫,通过减缓萌发速度适应干旱环境,萌发后为了维持自身正常生命活动,增加根长,增加植株的吸水能力,从而维持植株在逆境下的生长。
土壤干旱条件下,peu-MIR1444b过表达株系与野生型株系气孔导度差异不显著,但前者叶片水分利用效率、光合速率和PSⅡ最大光化学效率均显著高于野生型,因此peu-MIR1444b的过表达可以通过提高植物叶片水分利用效率从而提高植物的光合速率和PSⅡ最大光化学效率。复水3 d后,TG-1叶片水分利用效率与Col-0差异不显著,但气孔导度显著高于Col-0,TG-1通过提高气孔导度,进而提高了植物的光合速率和PSⅡ最大光化学效率。复水3 d后的TG-2和TG-5通过提高气孔导度和叶片水分利用效率从而提高植物的光合速率和PSⅡ最大光化学效率。综上所述,转基因株系通过提高气孔导度和叶片水分利用效率,从而提高植株对干旱环境的适应能力。
POD作用具有双重性,一方面在逆境初期表达,清除H2O2,表现为保护效应;另一方面在逆境后期表达,通过参与ROS的生成引发膜脂过氧化作用,表现为伤害效应[29]。POD活性的测定结果表明,peu-MIR1444b过表达株系能够在干旱环境中,快速产生较多的过氧化物酶,清除植物体内多余的H2O2,保护生物膜的功能进而维持自身较高的光合速率。复水后,转基因株系的POD活性开始下降,而野生型株系的POD活性继续上升,对自身产生伤害效应,故复水后野生型光合相关指标恢复能力显著弱于转基因株系。
综上所述,胡杨miR1444b在水分亏缺条件下通过促进根系生长增加植株的吸水能力,负调控葡聚糖合成酶Ⅳ提高植物渗透调节能力来维持植物光合效率,进而维持植株正常生长,在拟南芥中正调控植物抗旱性。
-
表 1 甘什岭调查样地概况
Table 1 Basic status of sample plots in Ganshiling Region
坡向
Slope aspect海拔
Altitude/m坡度
Slope grade/(°)林分密度/(株·hm-2)
Stand density/(plant·ha-1)凋落物厚
Litter thickness/cm郁闭度
Canopy density岩石裸露率
Exposed bedrock/%冠层高度
Canopy height/m半阳坡Semi-sunny slope 272~326 17~27 5 500~8 450 3.4~6.6 0.70~0.82 17~68 10.6~12.5 阳坡Sunny slope 201~322 3~31 5 400~19 100 2.1~7.1 0.68~0.91 0~22 8.8~17.2 半阴坡Semi-shady slope 219~335 5~30 1 200~12 500 2~7.5 0.63~0.90 0~75 8.4~12.2 阴坡Shady slope 208~278 3~31 6 050~8 000 2.4~5.9 0.60~0.82 0~50 10.4~11.9  下载: 导出CSV
下载: 导出CSV
表 2 棕榈藤种群静态生命表
Table 2 Static life table of four kinds of rattan population
种群
Population参数
Parameter龄级
Age gradeⅠ Ⅱ Ⅲ Ⅳ Ⅴ Ⅵ Ⅶ Ⅷ Ⅸ Ⅹ 白藤Calamus tetradactylus p 342 14 4 1 1 2 2 1 1 1 ˙p 342 14 5 4 3 2 2 1 1 1 li 1 000 41 15 12 9 6 6 3 3 3 di 328 9 1 1 1 0 1 0 0 — qi 1.0 0.6 0.2 0.3 0.3 0.0 0.5 0.0 0.0 — Li 178.0 9.5 4.5 3.5 2.5 2.0 1.5 1.0 1.0 1.0 Ti 204.5 26.5 17.0 12.5 9.0 6.5 4.5 3.0 2.0 1.0 ei 0.6 1.9 3.4 3.1 3.0 3.3 2.3 3.0 2.0 1.0 Ki 3.2 1.0 0.2 0.3 0.4 0.0 0.7 0.0 0.0 — 多果省藤Calamus walkeri p 456 282 44 4 5 6 2 3 3 5 ˙p 456 282 44 6 5 5 4 4 3 2 li 1 000 618 96 13 11 11 9 9 7 4 di 174 238 38 1 0 1 0 1 1 — qi 0.4 0.8 0.9 0.2 0.0 0.2 0.0 0.3 0.3 — Li 369.0 163.0 25.0 6.0 5.0 5.0 4.0 4.0 3.0 2.0 Ti 584.0 215.0 52.0 27.0 21.5 16.5 12.0 8.0 4.5 2.0 ei 1.3 0.8 1.2 4.5 4.3 3.3 3.0 2.0 1.5 1.0 Ki 0.5 1.9 2.0 0.2 0.0 0.2 0.0 0.3 0.4 — 杖藤Calamus rhabdocladus p 864 83 12 2 2 0 3 2 0 2 ˙p 864 83 13 9 6 4 3 3 2 1 lx 1 000 96 15 10 7 5 3 3 2 1 di 781 70 4 3 2 1 0 1 1 — qi 0.9 0.8 0.3 0.3 0.3 0.3 0.0 0.3 0.5 — Li 474.0 48.0 11.0 8.0 5.0 4.0 3.0 3.0 2.0 1.0 Ti 556.5 83.0 35.0 24.0 16.5 11.5 8.0 5.0 2.5 1.0 ei 0.6 1.0 2.7 2.7 2.8 2.9 2.7 1.7 1.3 1.0 Ki 2.3 1.9 0.4 0.4 0.4 0.3 0.0 0.4 0.7 — 黄藤Daemonorops jenkinsiana p 327 119 27 6 4 3 1 3 2 4 ˙p 327 119 27 6 4 4 3 3 2 2 li 1 000 364 83 18 12 12 9 9 6 6 di 208 92 21 2 0 1 0 1 0 — qi 0.6 0.8 0.8 0.3 0.0 0.3 0.0 0.3 0.0 — Li 223.0 73.0 17.0 5.0 4.0 4.0 3.0 3.0 2.0 2.0 Ti 334.6 111.7 38.6 22.0 17.0 13.0 9.5 6.5 4.0 2.0 ei 1.0 0.9 1.4 3.7 4.3 3.3 3.2 2.2 2.0 1.0 Ki 1.0 1.5 1.5 0.4 0.0 0.3 0.0 0.4 0.0 —
下载: 导出CSV
表 3 棕榈藤存活曲线模型
Table 3 Models of rattan survival curves
藤种Rattan species 方程式
Equation曲线类型
Curve typeR2 Radj2 F P 白藤Calamus tetradactylus Nx=6.942x-0.802 Deevey-Ⅲ 0.962 0.957 200.077 0.000** 多果省藤Calamus walkeri Nx=8.143x-0.688 Deevey-Ⅲ 0.920 0.910 92.504 0.000** 杖藤Calamus rhabdocladus Nx=9.261e-0.319x Deevey-Ⅱ 0.840 0.820 42.128 0.000** 黄藤Daemonorops jenkinsiana Nx=7.783x-0.638 Deevey-Ⅲ 0.957 0.951 175.910 0.000** 注:Nx为存活率,x为龄级。Note:Nx is survival rate, x is denoted age grade.
下载: 导出CSV
表 4 棕榈藤天然种群的周期性波动
Table 4 Periodic fluctuation of natural regeneration of rattan population
藤种Rattan species A1 A2 A3 A4 A5 白藤Calamus tetradactylus 1.168 1.435 1.131 1.223 1.362 多果省藤Calamus walkeri 1.500 3.805 4.429 4.154 1.098 杖藤Calamus rhabdocladus 1.829 3.538 3.906 2.161 2.662 黄藤Daemonorops jenkinsiana 1.343 3.118 0.692 0.208 0.686 注:A1~A5表示种群波动周期振幅,1~5表示波谱周期。Note: A1-A5 are denoted amplitudes of wave cycle of population, 1-5 are represented wave cycles.
下载: 导出CSV
-
[1] Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercus garryana) in British Columbia, Canada[J]. Forest Ecology and Management, 2000, 137(1): 209-219. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=a73d84970d763bf476b238e50db0cd54
[2] 张悦, 易雪梅, 王远遐, 等.采伐对红松种群结构与动态的影响[J].生态学报, 2015, 35(1):38-45. doi: 10.3969/j.issn.1673-1182.2015.01.008 Zhang Y, Yi X M, Wang Y X, et al. Impact of tree harvesting on the population structure and dynamics of Pinus koraiensis (Pinaceae) [J]. Acta Ecologica Sinica, 2015, 35(1): 38-45. doi: 10.3969/j.issn.1673-1182.2015.01.008
[3] 段桂芳, 单立山, 李毅, 等.甘肃中西部地区红砂种群结构及空间格局特征[J].水土保持研究, 2016, 23(1):67-74. http://d.old.wanfangdata.com.cn/Periodical/stbcyj201601013 Duan G F, Shan L S, Li Y, et al. Population structure and spatial pattern of Teaumuria soongorica population in midwest region of Gansu[J]. Research of Soil and Water Conservation, 2016, 23(1): 67-74. http://d.old.wanfangdata.com.cn/Periodical/stbcyj201601013
[4] 潘丽梅, 闫志刚, 马小军, 等.广西大明山自然保护区蛇足石杉种群生命表及生存分析[J].生态科学, 2015, 34(1):25-30. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=stkx201501004 Pan L M, Yan Z G, Ma X J, et al. Life table and survival analysis of Huperzia serrata population in Guangxi Damingshan Nature Reserve[J]. Ecological Science, 2015, 34(1): 25-30. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=stkx201501004
[5] 张婕, 上官铁梁, 段毅豪, 等.灵空山辽东栎种群年龄结构与动态[J].应用生态学报, 2014, 25(11):3125-3130. http://d.old.wanfangdata.com.cn/Periodical/yystxb201411008 Zhang J, Shangguan T L, Duan Y H, et al. Age structure and dynamics of Quercus wutaishanica population in Lingkong Mountain of Shanxi Province, China[J]. Chinese Journal of Applied Ecology, 2014, 25(11): 3125-3130. http://d.old.wanfangdata.com.cn/Periodical/yystxb201411008
[6] 张亚芳, 李登武, 王梅, 等.黄土高原不同地区杜松种群结构与动态[J].林业科学, 2015, 51(2):1-10. http://d.old.wanfangdata.com.cn/Periodical/lykx201502001 Zhang Y F, Li D W, Wang M, et al. Population structure and dynamics of Juniperus rigida in different regions of Loess Plateau[J]. Scientia Silvae Sinicae, 2015, 51(2): 1-10. http://d.old.wanfangdata.com.cn/Periodical/lykx201502001
[7] 胡喜生, 洪伟, 吴承祯, 等.木荷天然种群生命表分析[J].广西植物, 2007, 27(3):469-474. doi: 10.3969/j.issn.1000-3142.2007.03.019 Hu X S, Hong W, Wu C Z, et al. Analysis of the life table of natural population Schima superba[J]. Guihaia, 2007, 27(3): 469-474. doi: 10.3969/j.issn.1000-3142.2007.03.019
[8] 龙成, 杨小波, 龙文兴, 等.铜鼓岭热带常绿季雨矮林5种蒲桃属植物的种群结构及空间格局[J].林业科学, 2015, 51(2):18-27. http://d.old.wanfangdata.com.cn/Periodical/lykx201502003 Long C, Yang X B, Long W X, et al. Population structure and spatial patterns of five syzygium species in tropical evergreen monsoon elfin forest, Tongguling[J]. Scientia Silvae Sinicae, 2015, 51(2): 18-27. http://d.old.wanfangdata.com.cn/Periodical/lykx201502003
[9] 汤孟平, 沈丽芬, 赵赛赛, 等.基于谱分析的天目山毛竹林生长周期[J].林业科学, 2014, 50(9):184-188. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=lykx201409028 Tang M P, Shen L F, Zhao S S, et al. Growth cycle of Phyllostachys edulis stand in Tianmu Mountain based on spectrum analysis[J]. Scientia Silvae Sinicae, 2014, 50(9): 184-188. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=lykx201409028
[10] Dransfield J, Uhl N, Asmussen C, et al. The evolution and classification of palms[M]. London: Kew Publishing, 141-204.
[11] Putz F E. Growth habits and trellis requirements of climbing palms (Calamus spp.) in North-Eastern Queensland[J]. Australian Journal of Botany, 1990, 38(6): 603-608. doi: 10.1071/BT9900603
[12] Kidyoo A M, Mckey D. Flowering phenology and mimicry of the rattan Calamus castaneus(Arecaceae) in southern Thailand[J]. Botany, 2012, 90(9): 856-865. doi: 10.1139/b2012-058
[13] 王慷林.中国棕榈藤资源及其分布特征研究[J].植物科学学报, 2015, 33(3):320-325. http://d.old.wanfangdata.com.cn/Periodical/whzwxyj201503007 Wang K L. Resources and distribution of rattan in China[J]. Plant Science Journal, 2015, 33(3): 320-325. http://d.old.wanfangdata.com.cn/Periodical/whzwxyj201503007
[14] 刘晋仙, 陶建平, 王玉平, 等.海南霸王岭山地原始林与伐后林中木质藤本与支持木的多样性及其相互关系[J].林业科学, 2012, 48(5):15-19. http://d.old.wanfangdata.com.cn/Periodical/lykx201205003 Liu J X, Tao J P, Wang Y P, et al. Species diversity and tree-liana relationship in the tropical montane primary forest and post logged forest of Bawanglin, Hainan Island, China[J]. Scientia Silvae Sinicae, 2012, 48(5): 15-19. http://d.old.wanfangdata.com.cn/Periodical/lykx201205003
[15] 漆良华, 梁昌强, 毛超, 等.海南岛甘什岭热带低地次生雨林物种组成与地理成分[J].生态学杂志, 2014, 33(4):922-929. http://d.old.wanfangdata.com.cn/Periodical/stxzz201404011 Qi L H, Liang C Q, Mao C, et al. Species composition and geographic elements of the tropical lowland secondary rain forest of Ganshiling, Hainan Island, China[J]. Chinese Journal of Ecology, 2014, 33(4): 922-929. http://d.old.wanfangdata.com.cn/Periodical/stxzz201404011
[16] 王峥峰, 安树青, David G C, 等.海南岛吊罗山山地雨林物种多样性[J].生态学报, 1999, 19(1):61-67. doi: 10.3321/j.issn:1000-0933.1999.01.009 Wang Z F, An S Q, David G C, et al. Biodiverstiy of the montane rain forest in Diaoluo Mountain, Hainan[J]. Acta Ecologica Sinica, 1999, 19(1): 61-67. doi: 10.3321/j.issn:1000-0933.1999.01.009
[17] 赵科, 陶建平, 郝建辉, 等.海南岛霸王岭热带山地雨林木质藤本垂直结构特征[J].生态学报, 2010, 30(12):3178-3181. http://d.old.wanfangdata.com.cn/Periodical/stxb201012010 Zhao K, Tao J P, Hao J H, et al. Vertical structure of liana in a tropical montane rain forest in Bawangling Nature Reserve, Hainan Island, China[J]. Acta Ecologica Sinica, 2010, 30(12): 3178-3181. http://d.old.wanfangdata.com.cn/Periodical/stxb201012010
[18] 朱学雷, 安树青, 张立新, 等.海南五指山热带山地雨林主要种群结构特征分析[J].应用生态学报, 1999, 10(6):641-644. http://www.cnki.com.cn/Article/CJFDTOTAL-YYSB199906000.htm Zhu X L, An S Q, Zhang L X, et al. Population structure of tropical montane rainforest on Wuzhi Mountain of Hainan[J]. Chinese Journal of Applied Ecology, 1999, 10(6): 641-644. http://www.cnki.com.cn/Article/CJFDTOTAL-YYSB199906000.htm
[19] 路兴慧, 丁易, 臧润国, 等.海南岛热带低地雨林木本植物幼苗层组成与功能群恢复动态[J].林业科学研究, 2012, 25(6):726-731. doi: 10.3969/j.issn.1001-1498.2012.06.008 Lu X H, Ding Y, Zang R G, et al. Recovery dynamic of seedling composition and functional groups in the tropical lowland rain forest on Hainan Island, China[J]. Forest Research, 2012, 25(6): 726-731. doi: 10.3969/j.issn.1001-1498.2012.06.008
[20] 臧润国, 余世孝, 刘静艳, 等.海南霸王岭热带山地雨林林隙更新规律的研究[J].生态学报, 1999, 19(2):151-158. http://d.old.wanfangdata.com.cn/Periodical/stxb199902002 Zang R G, Yu S X, Liu J Y, et al. The gap phase regeneration in a tropical montane rain forest in Bawangling, Hainan Island[J]. Acta Ecologica Sinica, 1999, 19(2): 151-158. http://d.old.wanfangdata.com.cn/Periodical/stxb199902002
[21] 星耀武, 王慷林, 杨宇明.中国省藤属(棕榈科)区系地理研究[J].云南植物研究, 2006, 28(5):461-467. doi: 10.3969/j.issn.2095-0845.2006.05.003 Xing Y W, Wang K L, Yang Y M, et al. Floristic geography of calamus (Palmae Calamoideae) in China[J]. Acta Botanica Yunnanica, 2006, 28(5): 461-467. doi: 10.3969/j.issn.2095-0845.2006.05.003
[22] Ruppert N, Mansor A, Sah S A M. Rattan (Calamoideae) abundance and above-ground biomass at a primary rainforest of Peninsular Malaysia[J]. Plant Ecology & Diversity, 2015, 9(1): 63-67. http://cn.bing.com/academic/profile?id=811defffb04f4333feebabd6f9bed57c&encoded=0&v=paper_preview&mkt=zh-cn
[23] Watanabe N M, Suzuki E. Species diversity, abundance, and vertical size structure of rattans in Borneo and Java[J]. Biodiversity Conservation, 2008, 17(3): 523-538. doi: 10.1007/s10531-007-9268-1
[24] 殷谷丽, 唐建维, 杨成源, 等.四种省藤属植物的光合特征与叶片性状及生长的相关性[J].中南林业科技大学学报, 2010, 30(6):104-112. doi: 10.3969/j.issn.1673-923X.2010.06.019 Yin G L, Tang J W, Yang C Y, et al. Relationships among photosynthesis and leaf traits and growth of four rattan species seedlings[J]. Journal of Central South University of Forestry & Technology, 2010, 30(6): 104-112. doi: 10.3969/j.issn.1673-923X.2010.06.019
[25] Isnard S, Rowe N P. The climbing habit in palms: biomechanics of the cirrus and flagellum[J]. America Journal of Batany, 2008, 95(12): 1538-1547. http://cn.bing.com/academic/profile?id=0c18bcdf6690ff232187e1f898b30739&encoded=0&v=paper_preview&mkt=zh-cn
[26] Isnard S, Rowe N P. Mechanical role of the leaf sheath in rattans[J]. New Phytologist, 2010, 177(3): 643-652. http://cn.bing.com/academic/profile?id=9a8a14914a1d13e1a21c5952855bb092&encoded=0&v=paper_preview&mkt=zh-cn
[27] 曾炳山, 许煌灿, 尹光天, 等.黄藤藤丛结构和生长的研究[J].林业科学研究, 1993, 6(4):414-422. doi: 10.3321/j.issn:1001-1498.1993.04.001 Zeng B S, Xu H C, Yin G T, et al. A study on growth structure of clump of Daemonorops jenkinsiana[J]. Forest Research, 1993, 6(4): 414-422. doi: 10.3321/j.issn:1001-1498.1993.04.001
[28] 杨锦昌, 许煌灿, 尹光天, 等.黄藤人工林密度效应[J].林业科学, 2006, 42(4):57-61. http://d.old.wanfangdata.com.cn/Periodical/lykx200604010 Yang J C, Xu H C, Yin G T, et al. The density effect of Damonorops jenkinsiana plantation[J]. Scientia Silvae Sinicae, 2006, 42(4): 57-61. http://d.old.wanfangdata.com.cn/Periodical/lykx200604010
[29] 杨锦昌, 尹光天, 冯昌林, 等.单叶省藤人工林茎长分布模型的研制[J].浙江林业科技, 2010, 30(6):33-37. doi: 10.3969/j.issn.1001-3776.2010.06.007 Yang J C, Yin G T, Feng C L, et al. Establishment of stem length distribution model for the plantation of Calamus simplcifolius[J]. Journal of Zhejiang Por Sci & Tech, 2010, 30(6): 33-37. doi: 10.3969/j.issn.1001-3776.2010.06.007
[30] Svenning J C. Environmental heterogeneity, recruitment limitation and the mesoscale distribution of palms in a tropical montane rain forest(Maquipucuna, Ecuador) [J]. Journal of Tropical Ecology, 2001, 17: 97-113. doi: 10.1017/S0266467401001067
[31] Thonhofer J, Getto D, Straaten O V, et al. Influence of spatial and environmental variables on rattan palm (Arecaceae) assemblage composition in Central Sulawesi, Indonesia[J]. Plant Ecology, 2015, 216(1): 55-66. doi: 10.1007/s11258-014-0416-x
[32] 陈瑞国, 范少辉, 刘广路, 等.海南岛次生低地雨林棕榈藤伴生群落优势种生态位研究[J].西北植物学报, 2017, 37(6): 1226-1233. http://d.old.wanfangdata.com.cn/Periodical/xbzwxb201706021 Chen R G, Fan S H, Liu G L, et al. Niche characteristics of dominant species of rattan accompanying community in secondary lowland rain forest in Hainan Island, China[J]. Acta Botany Boreal.-Occident. Sin, 2017, 37(6): 1226-1233. http://d.old.wanfangdata.com.cn/Periodical/xbzwxb201706021
[33] 尹光天, 许煌灿, 张伟良.光照与藤苗生长的初步研究[J].林业科学研究, 1988, 1(5):548-552. http://www.cnki.com.cn/Article/CJFDTOTAL-LYKX198805012.htm Yin G T, Xu H C, Zhang W L. A preliminary study on the effect of different levels of light intensity on the growth of rattan seedlings[J]. Forest Research, 1988, 1(5): 548-552. http://www.cnki.com.cn/Article/CJFDTOTAL-LYKX198805012.htm
[34] 李建光, 李荣生, 尹光天, 等.竹柏林6年生单叶省藤的生长指标和光合特征[J].中南林业科技大学学报, 2012, 32(4):86-90. http://d.old.wanfangdata.com.cn/Periodical/znlxyxb201204018 Li J G, Li R S, Yin G T, et al. Growth index and photosynthetic characteristics of 6-year-old Calamus simplicifolius in Nageia nagi plantation[J]. Journal of Central South University of Forestry & Techology, 2012, 32(4): 86-90. http://d.old.wanfangdata.com.cn/Periodical/znlxyxb201204018
[35] 彭超, 王慷林, 李莲芳, 等.施肥对省藤移植苗木生长的影响[J].中南林业科技大学学报, 2016, 36(1):58-62. http://d.old.wanfangdata.com.cn/Periodical/znlxyxb201601010 Peng C, Wang K L, Li L F, et al. Effects of three fertilizers on growth of calamus transplanting seedlings[J]. Journal of Central South University of Forestry & Techology, 2016, 36(1): 58-62. http://d.old.wanfangdata.com.cn/Periodical/znlxyxb201601010
[36] Bøgh A. Abundance and growth of rattans in Khao Chong National Park, Thailand[J]. Forest Ecology and Management, 1996, 84: 71-80. doi: 10.1016/0378-1127(96)03738-3
[37] Li R S, Xu H C, Yang J C, et al. A review of relationship between rattan and water[J]. Forestry studies in China, 2002, 4(1): 65-68. http://en.cnki.com.cn/Article_en/CJFDTotal-BLDX200201010.htm
[38] Toledo-aceves T, Swaine M D. Biomass allocation and photosynthetic responses of lianas and pioneer tree seedlings to light[J]. Acta Oecologica, 2008, 34(1): 38-49. doi: 10.1016/j.actao.2008.03.003
[39] 寇亮, 尹光天, 杨锦昌, 等.不同滴灌处理对黄藤人工林生长的影响[J].中南林业科技大学学报, 2012, 32(4):100-104. doi: 10.3969/j.issn.1673-923X.2012.04.021 Kou L, Yin G T, Yang J C, et al. Effects of different drip irrigation on growth of Daemonorops margaritae plantation[J]. Journal of Central South University of Forestry & Techology, 2012, 32(4): 100-104. doi: 10.3969/j.issn.1673-923X.2012.04.021
[40] 张伟良, 尹光天, 许煌灿.白藤丛栽试验初报[J].林业科学研究, 1990, 3(1):81-85. http://www.cnki.com.cn/Article/CJFD1990-LYKX199001018.htm Zhang W L, Yin G T, Xu H C. A preliminary report of group-planting trial of Calamus tetradactylus[J]. Forest Research, 1990, 3(1): 81-85. http://www.cnki.com.cn/Article/CJFD1990-LYKX199001018.htm
[41] Svenning J C. Microhabitat specialization in a species-rich palm community in Amazonian Ecuador[J]. Journal of Ecology, 1999, 87(1): 55-65. doi: 10.1046/j.1365-2745.1999.00329.x
[42] 张菲菲, 李担, 汪佑宏, 等.棕榈藤材藤龄的判断[J].东北林业大学学报, 2017, 45(6):26-29. doi: 10.3969/j.issn.1000-5382.2017.06.006 Zhang F F, Li D, Wang Y H, et al. Judgment of rattan age[J]. Journal of Northeast Forestry University, 2017, 45(6): 26-29. doi: 10.3969/j.issn.1000-5382.2017.06.006
[43] 彭超, 范少辉, 刘广路, 等.海南岛低地次生雨林棕榈藤分布及其影响因子[J].生态学杂志, 2017, 36(10):2725-2733. http://d.old.wanfangdata.com.cn/Periodical/stxzz201710006 Peng C, Fan S H, Liu G L, et al. Distribution and impact factors of rattan in lowland secondary rain forest in Hainan Island, China[J]. Chinese Journal of Ecology, 2017, 36(10): 2725-2733. http://d.old.wanfangdata.com.cn/Periodical/stxzz201710006
-
期刊类型引用(9)
1. ZHANG Kai,ZHANG Hailong,TIAN Jianjin,QU Jianjun,ZHANG Xingxin,WANG Zhenghui,XIAO jianhua. Flow field, sedimentation, and erosion characteristics around folded linear HDPE sheet sand fence: Numerical simulation study. Journal of Mountain Science. 2024(01): 113-130 .  必应学术
必应学术
2. 马泽,蒙仲举,罗建国,阿拉腾陶格苏,赵东颖. 乌兰布和沙漠典型沿黄段格状沙障防风固沙效应. 水土保持通报. 2024(02): 1-10+21 . 百度学术
3. 闫德仁,黄海广,闫婷. 纱网沙障研究进展. 内蒙古林业科技. 2024(03): 50-53 . 百度学术
4. 潘加朋,张克存,安志山,张宏雪,薛承杰. 基于风洞模拟试验的典型机械防沙工程防护效益对比. 水土保持学报. 2023(06): 104-110 . 百度学术
5. 张帅,原伟杰,刘卉,王海霞,官昊慧,王鹿振. 附面层位移厚度对沙障防风效益评价的适用性研究——以聚乳酸(PLA)沙障为例. 干旱区地理. 2023(12): 1973-1983 . 百度学术
6. 李双立,黄海广,党晓宏,郭跃,万俊华. 基于CNKI数据库文献计量视角下沙障研究现状. 防护林科技. 2022(05): 75-78+85 . 百度学术
7. 韩雪莹,王涛,杨文斌,贾光普,刘静,杨宇. 中国沙障研究进展与热点分析:基于Vosviewer和Citespace的图谱量化分析. 中国沙漠. 2021(02): 153-163 . 百度学术
8. 祁帅,汪季,党晓宏,魏亚娟,丁奋谦. 3种低立式沙障内表层沉积物粒度特征研究. 干旱区研究. 2021(03): 875-881 . 百度学术
9. 厉静文,Dosmanbetov D A,郭浩,辛智鸣,刘朋飞,刘明虎. 不同配置乔灌混交林防风效益的风洞试验. 农业工程学报. 2020(11): 95-102 . 百度学术
其他类型引用(7)





计量
- 文章访问数: 1226
- HTML全文浏览量: 381
- PDF下载量: 21
- 被引次数: 16
