
Effects of forest tree species diversity on soil carbon and nitrogen contents in China
-
摘要:目的
揭示不同气候带森林生态系统中土壤碳氮含量对树种多样性的响应差异,并探讨影响这种响应的关键生物和非生物因素。
方法收集中国热带、亚热带、暖温带和中温带4个森林生态系统树种多样性,土壤碳、氮含量和土壤真菌多样性数据,针对处理组(多个树种)和对照组(单一树种)共计214组数据进行整合分析,使用随机效应模型计算多样性效应值,并分析不同气候带森林生态系统影响树种多样性效应值的生物和非生物变量的解释权重。
结果不同气候带森林土壤碳、氮含量对树种多样性存在差异性响应,随着纬度梯度增高,树种多样性对土壤碳、氮含量的影响逐渐减弱。热带和亚热带森林较强的树种多样性效应主要源于海拔和土壤pH,而非土壤真菌群落多样性,但是暖温带和中温带森林土壤真菌多样性则是调控树种多样性对土壤碳、氮含量影响的重要因素之一。
结论中国不同气候带森林生态系统的土壤碳、氮含量对树种多样性的响应格局一定程度上解释了局域尺度研究中森林树种多样性对土壤碳库影响机制的分歧,同时也说明树种多样性的变化对于热带和亚热带森林土壤碳库的影响可能更为剧烈。
Abstract:ObjectiveTo reveal the differences in the response of soil carbon and nitrogen content to tree species diversity in forest ecosystems in different climatic zones, and to explore the key biotic and abiotic factors affecting this response.
MethodWe collected the data of altitude, air temperature, soil properties (including soil carbon (C), nitrogen (N), C/N ratio), as well as fungal diversity from sample plots with both tree species mixtures and monocultures. In total, 214 sets of data were used to conduct meta analysis, with a random-effect model calculating effect size. Additionally, the explanatory weights of biotic and abiotic variables influencing the effect of tree richness were analyzed.
ResultWe found that the responses of soil C and N contents to tree richness varied across forest ecosystems, with the strength of tree richness effects on soil C and N contents gradually diminishing from tropical to mid-temperate forest ecosystems. Moreover, the effects of tree richness on soil C and N contents in tropical and subtropical forest ecosystems were primarily driven by altitude and soil pH, rather than soil fungal diversity. In contrast, cascading effects of tree richness on soil C and N contents were observed via modulating soil fungal diversity in warm-temperate and mid-temperate forest ecosystems.
ConclusionThe response patterns of soil carbon and nitrogen contents in forest ecosystems of different climate zones in China to tree species diversity, to some extent, explain the variability mechanisms of forest tree species diversity on soil carbon pools in local scale studies, and also indicate that changes in tree species diversity may have a more severe impact on soil carbon pools in tropical and subtropical forests.
-
森林生态系统占全球陆地面积30%[1],承载着陆地生态系统2/3的植物和动物物种多样性[2],蓄积了陆地生态系统45%的碳和50%的净初级生产力[3]。然而,世界森林资源报告最新数据显示,全球森林碳储量正面临持续下降的趋势,在1990—2020年的30年间,从668 Gt下降到662 Gt[3]。厘清森林碳储量变化的影响因素,对于提升森林质量、应对全球变化至关重要。
森林生物多样性在调控森林固碳、养分周转、维持生态系统稳定性方面至关重要[4−5]。其中,植物与土壤微生物多样性是影响森林土壤碳库的重要因素。大量研究表明森林植物多样性的增加能够调控森林植被生产力,从而提升植被碳储量,但是森林土壤碳储量的响应格局及其内在机制却仍不清楚。研究[6]发现森林植物多样性减少虽然会一定程度地降低土壤碳的积累;但也可能导致群落中生态位释放,减缓种间竞争,进而增加土壤碳含量[7]。植物多样性变化可以通过改变植物凋落物、根际分泌物输入等过程,直接改变土壤碳含量;也可以通过改变土壤微环境,或调控土壤微生物量、活性、群落多样性和组成,间接地影响土壤生态过程,改变土壤碳含量,影响土壤碳循环[8−10]。土壤真菌是土壤微生物群落中的重要组成部分,在森林生态系统有机碳形成过程和养分循环过程中起着重要作用,其多样性能够在一定程度上促进森林生产力的稳定性[11−12]。然而,植物多样性与真菌多样性对土壤碳含量的影响并非是孤立的,而是相互耦合、共同驱动的。植物与土壤真菌多样性往往形成级联效应,即植物群落变化改变土壤真菌多样性,进而影响土壤碳氮循环。当然,这种植物–土壤真菌–土壤碳的级联效应还可能受到气候因素、土壤养分和人类活动等多方面的干扰,因此也表现出一定的不确定性[13−14]。
本研究通过整合分析,收集中国热带、亚热带、暖温带和中温带4个气候带森林生态系统树种多样性,土壤碳、氮含量和土壤真菌多样性数据共计214组,旨在揭示不同气候带森林生态系统树种多样性与土壤真菌多样性影响土壤碳氮含量的耦合机制。土壤真菌对温度、海拔等环境因子的变化极为敏感,其在不同环境下的群落差异又将影响到植物所需养分的供给,从而影响植物生长。基于此,我们推测(1)不同气候带森林土壤碳库对树种多样性增加存在差异性响应,随着纬度梯度增高树种多样性对土壤碳库的影响会有所减弱。(2)考虑到森林树种多样性变化会显著改变土壤真菌群落,而真菌是参与土壤有机碳分解的重要土壤生物类群,尤其对顽拗有机养分的降解,森林树种多样性可能通过改变土壤真菌多样性而影响土壤碳、氮含量。
1. 材料与方法
1.1 资料来源
整合分析(meta-analysis)是近年来在生态学领域围绕所提出的问题,通过大量文献结论定量归纳的统计方法。本研究基于整合分析,通过对web of science、TreeDiv Net、中国知网等网站进行了相关文献检索,通过森林(forest)、树种多样性(tree richness)、真菌香农指数(fungal Shannon diversity)和土壤碳(soil organic carbon)等关键词进行文献筛选。文献筛选基本原则:剔除重复报告与信息不完整的文献;确保各试验样本具有重复;若同一篇文章中存在不同季节、土层深度的数据,则选取夏季表层土壤数据;对于用图形展示的数据,使用Getdata version 2.0软件进行数值化。通过逐层筛选,获取符合分析标准的文献48篇(详见附录1),包括对照组(单一树种)和处理组(多个树种)的数据共214组,其中热带森林生态系统9组、亚热带56组、暖温带130组和中温带19组。由此,提取了对照组和处理组中的土壤有机碳(soil organic carbon,SOC)、土壤全氮(total nitrogen,TN)、土壤碳氮比(carbon-nitrogen ratio,C/N)、土壤真菌多样性指数(即香农指数,H)的均值、标准差、样本量及其研究位点树种多样性;同时,收集了各文献中研究点经纬度、年平均温度、年平均降水量和海拔等气候和地形数据。
1.2 计算方法
在R version 4.1.3中进行了统计分析、差异分析和回归分析。由于标准差是meta分析中所需的重要参数,若文献中没有提供标准差,则使用metagear软件包估计标准差值,使用随机效应模型计算效应值(Hedges’d,d)[15]。
d=Xe−XcSJ J=1−34(Nc+Ne−2)−1 S=√(Nc−1)S2e+(Nc−1)S2cNc+Ne−2 Vd=Ne+NcNeNc+d22(Ne+Nc) 式中:Xe为处理组均值,Xc为对照组均值,J为校正偏差,N为样本量,Ne为处理组,Nc为对照组,S为对照组和处理组混合标准差,Se和Sc分别是处理和对照组标准差,Vd为方差。若d > 0,处理对试验结果具有正效应;若d = 0,处理对试验结果不产生影响;若d < 0,处理对试验结果具有负效应。
当各指标效应值的95%置信区间(confidence interval,CI)不与0重叠,则认为处理对该指标的影响达到显著性水平(P < 0.05)。此外,本研究使用Qt统计量进行异质性检验,Qt为组间异质性(Qb)和组内异质性(Qw)的总和。为了明确同一分组内不同处理间差异是否显著,对Qb值进行卡方检验。如果P < 0.05,即可认为同一分组内不同处理间差异显著;反之,则不显著。本文中效应值使用metafor软件包进行计算。
2. 结果与分析
2.1 不同森林生态系统类型树种多样性对土壤碳、氮含量的影响
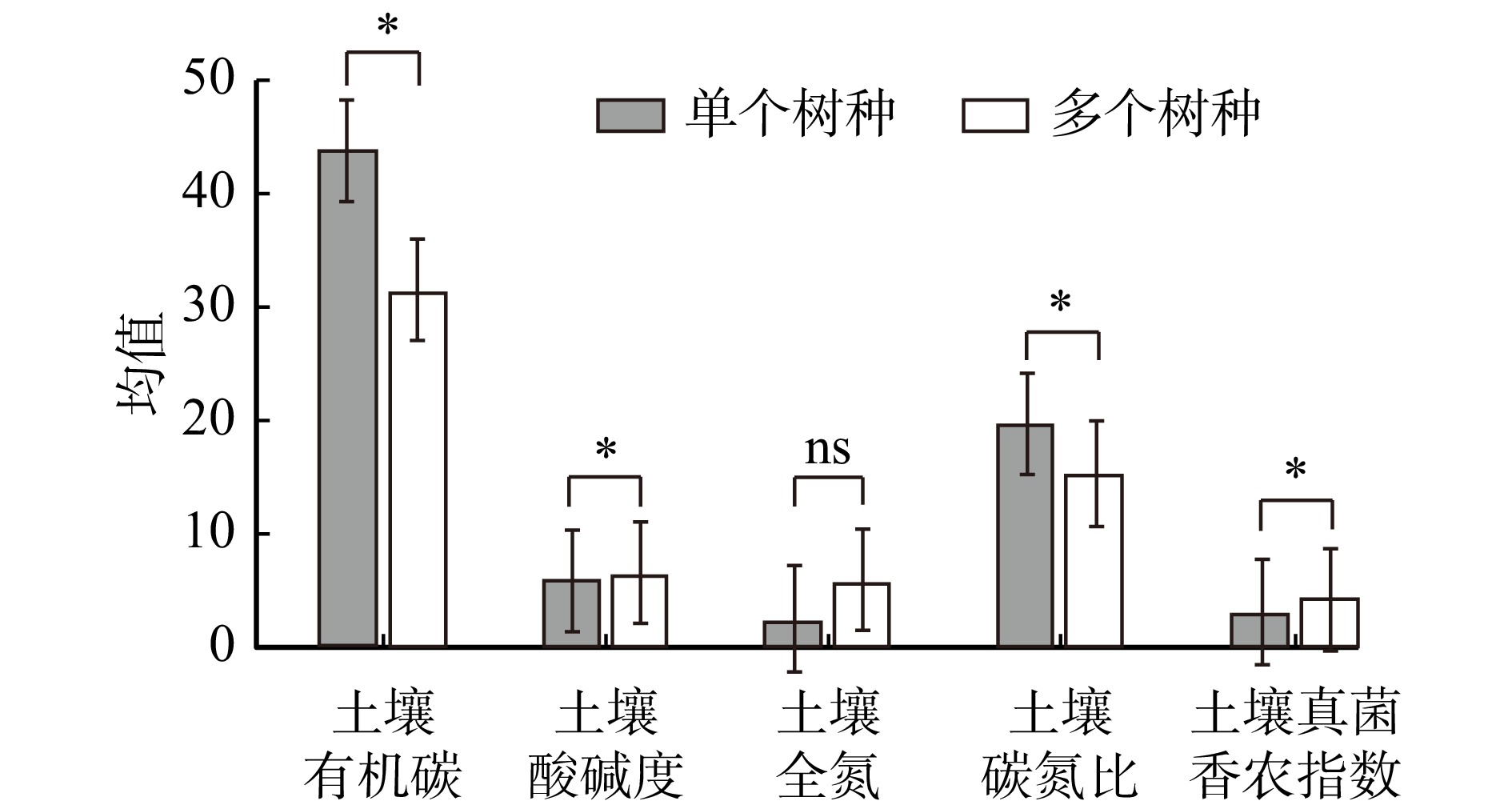
与单种样方相比,多物种混合样方具有较低的土壤碳氮比和土壤有机碳含量(P < 0.05);多物种混合样方内的土壤pH和土壤真菌香农指数显著高于单种样方(P < 0.05)。土壤全氮在两类样方间的差异不显著(图1)。
![]() 图 1 森林生态系统树种多样性对土壤碳、氮含量及土壤真菌多样性的影响土壤有机碳、土壤全氮的单位均为g/kg,pH、碳氮比和真菌香农指数无量纲。*表示在0.05水平上存在显著差异。ns表示差异不显著。Figure 1. Effects of tree species diversity on soil carbon, nitrogen contents, and soil fungal diversity in forest ecosystems
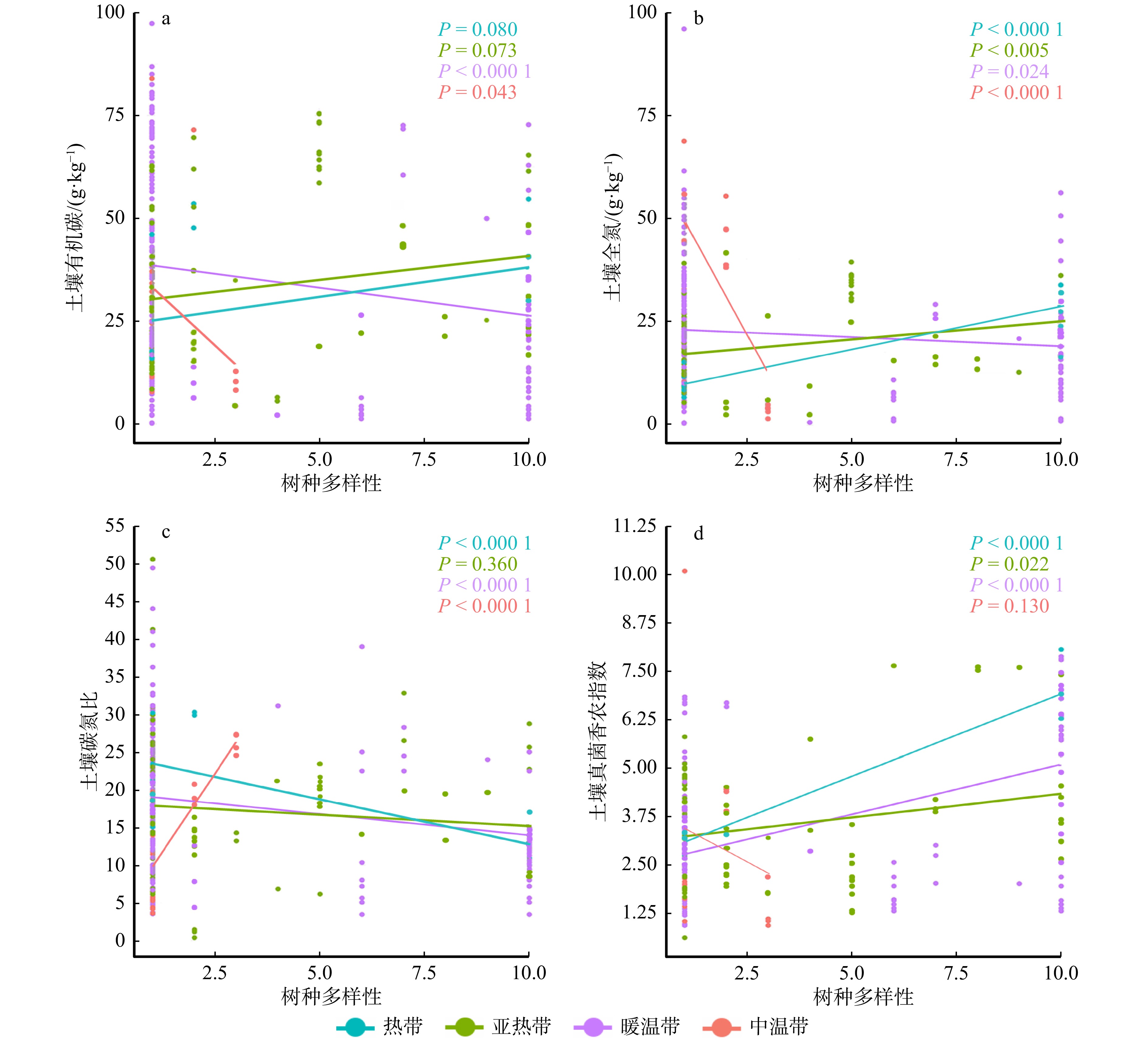
图 1 森林生态系统树种多样性对土壤碳、氮含量及土壤真菌多样性的影响土壤有机碳、土壤全氮的单位均为g/kg,pH、碳氮比和真菌香农指数无量纲。*表示在0.05水平上存在显著差异。ns表示差异不显著。Figure 1. Effects of tree species diversity on soil carbon, nitrogen contents, and soil fungal diversity in forest ecosystems在热带与亚热带中,森林土壤有机碳含量随着树种多样性的增加均呈现增加的趋势。而在暖温带和中温带中,土壤有机碳含量随树种多样性增加呈现减少的趋势(图2a)。与土壤有机碳基本一致,土壤全氮在热带与亚热带中,随树种多样性增加逐渐增加;在暖温带与中温带中,随树种多样性增加逐渐减少(图2b)。随着树种多样性的增加,土壤碳氮比在热带、亚热带和暖温带呈下降趋势,而在中温带呈上升趋势(图2c)。与土壤碳氮比的变化趋势相反,随着树种多样性的增加,土壤真菌香农指数在热带、亚热带和暖温带呈上升趋势,而在中温带呈下降趋势(图2d)。
![]() 图 2 不同森林生态系统树种多样性与土壤碳、氮和真菌多样性的回归分析Figure 2. Regression analysis of tree species diversity and soil carbon, nitrogen, and fungal diversity in forest ecosystems
图 2 不同森林生态系统树种多样性与土壤碳、氮和真菌多样性的回归分析Figure 2. Regression analysis of tree species diversity and soil carbon, nitrogen, and fungal diversity in forest ecosystems2.2 不同森林生态系统类型树种多样性调控土壤碳、氮含量的效应值
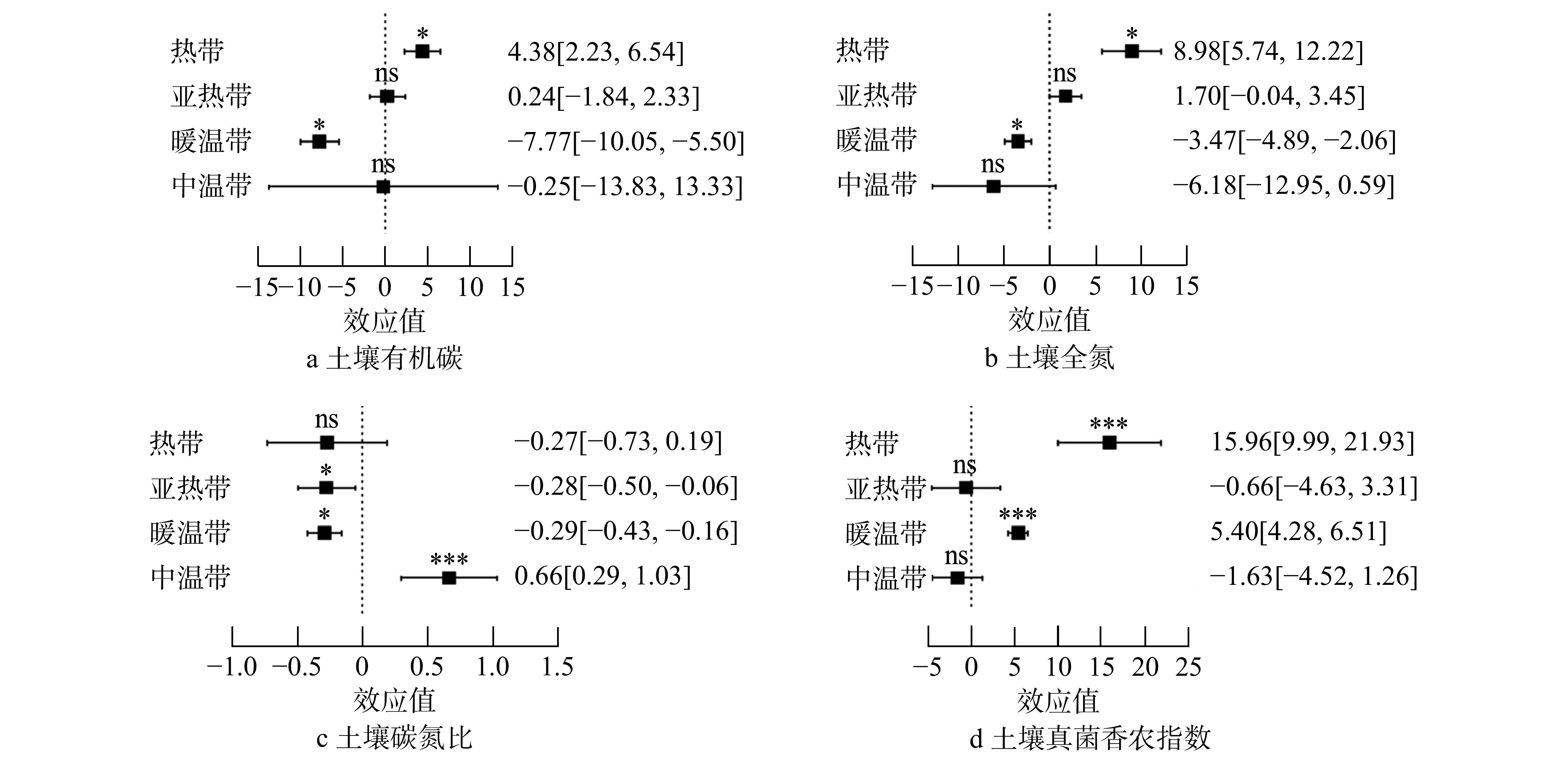
随着气候带温度降低,树种多样性对土壤碳、氮含量的作用呈现出由正效应向负效应的变化趋势(图3a、b);与之相反,树种多样性对土壤碳氮比的影响随着气候带温度的降低呈现逐渐增加的趋势(图3c)。在热带和亚热带森林生态系统,树种多样性对土壤碳、氮的累积表现出正效应,且热带所呈现的正效应较亚热带更为显著;暖温带和中温带森林树种多样性对土壤碳、氮的影响表现出负效应,其中,中温带森林树种多样性效应不显著。热带森林树种多样性对土壤碳氮比的影响不显著,而亚热带和暖温带森林树种多样性对土壤碳氮比呈现出显著的负效应。中温带森林树种多样性对土壤碳氮比的影响呈现出极显著的正效应。
![]() 图 3 森林生态系统树种多样性对土壤碳、氮和土壤真菌多样性的累计效应值方括号内数据是95%的置信区间。*表示影响显著(P<0.05),***表示影响极其显著( P<0.001);ns表示影响不显著。下同。Figure 3. Cumulative effect values of tree species diversity on soil carbon, nitrogen, and soil fungal diversity in forest ecosystems
图 3 森林生态系统树种多样性对土壤碳、氮和土壤真菌多样性的累计效应值方括号内数据是95%的置信区间。*表示影响显著(P<0.05),***表示影响极其显著( P<0.001);ns表示影响不显著。下同。Figure 3. Cumulative effect values of tree species diversity on soil carbon, nitrogen, and soil fungal diversity in forest ecosystems将气候、地形和土壤理化因子引入随机效应模型(表1),分析发现随着海拔增加,土壤有机碳、全氮含量和碳氮比的多样性效应值显著降低;随着纬度增加,土壤有机碳、全氮含量的多样性效应值显著降低;随着年平均温度的增加,土壤有机碳含量的多样性效应值显著降低,但土壤全氮的多样性效应值变化不显著;随着土壤pH的增加,土壤有机碳、全氮的多样性效应值则显著增加。
表 1 森林生态系统树种多样性效应值的影响因素Table 1. Influencing factors of tree species diversity effect values in forest ecosystems指标 土壤有机碳效应值 土壤全氮效应值 土壤碳氮比效应值 土壤真菌香农指数效应值 海拔 −0.010 1*** −0.005 1*** −0.000 3** 0.003 2* 纬度 −0.667 9* −0.627 1*** 0.019 3 −0.553 8** 年平均温度 −0.848 1* −0.201 7 −0.026 9 −0.721 7** 土壤真菌香农指数 0.960 4 −0.256 7 0.097 9* 土壤pH 2.123 1** 2.248 6*** −0.048 7 −0.529 1 土壤有机碳 −0.044 8 土壤全氮 −1.168 5 土壤碳氮比 0.031 2 注: **表示影响极显著(P < 0.01)。 通过最优模型诊断,进一步解析各变量对多样性效应值的解释权重(表2)。就总体数据来看,对于土壤有机碳效应值变化,解释权重最高的是海拔,其次是土壤pH。与此类似,亚热带和暖温带森林土壤有机碳效应值受到海拔和土壤pH的影响较大,但在热带和中温带森林却没有类似的结果。暖温带森林土壤真菌多样性对土壤有机碳效应值的解释权重较大。土壤全氮效应值变化,解释权重最高的是海拔和土壤pH,在暖温带中体现最明显。中温带森林土壤全氮效应值受海拔、土壤pH和真菌香农指数的影响都比较大。而在热带和亚热带森林中并没有类似的结果。对于土壤碳氮比效应值变化,解释权重最高的是海拔,其次是土壤真菌多样性,在暖温带森林中土壤碳氮比效应值受到海拔和真菌多样性的影响较大,而在热带、亚热带和中温带中没有明显的影响。
表 2 树种多样性效应值影响因子的解释权重Table 2. Explanatory weight of the factors affecting the effect values of tree species diversity指标 土壤有机碳效应值 土壤全氮效应值 土壤碳氮比效应值 土壤真菌香农指数效应值 总体 热带 亚热带 暖温带 中温带 总体 热带 亚热带 暖温带 中温带 总体 热带 亚热带 暖温带 中温带 总体 热带 亚热带 暖温带 中温带 海拔 0.993 0.500 0.989 1.000 0.281 1.000 0.500 0.291 0.998 0.986 0.960 0.500 0.279 0.783 0.261 0.690 0.500 0.249 0.306 0.413 土壤pH 0.965 0.141 0.831 1.000 0.194 1.000 0.291 0.386 1.000 0.940 0.352 0.102 0.422 0.272 0.222 0.320 0.722 0.420 0.640 0.725 土壤真菌
香农指数0.420 0.078 0.271 1.000 0.354 0.323 0.085 0.586 0.335 0.964 0.713 0.090 0.259 0.726 0.187 年平均温度 0.500 0.500 0.240 0.379 0.433 0.370 0.500 0.681 0.272 0.431 0.590 0.500 0.244 0.349 0.285 0.410 0.500 0.953 0.339 0.392 纬度 0.545 0.996 0.623 0.970 土壤有机碳 0.960 0.738 0.960 0.966 0.559 土壤全氮 0.896 0.120 0.290 0.834 0.471 土壤碳氮比 0.326 0.044 0.303 0.317 0.325 2.3 不同森林生态系统类型树种多样性调控土壤真菌多样性的效应值分析
随着气候带温度降低,树种多样性对土壤真菌多样性的影响呈现较大波动(图3d)。在热带与暖温带森林生态系统,树种多样性对土壤真菌香农指数表现出极显著的正效应,而在亚热带和中温带中,树种多样性对土壤真菌香农指数的影响并不显著(图3d)。土壤真菌香农指数的多样性效应值并未受到土壤碳、氮及二者比值的显著影响,而随着海拔的升高或年平均温度的降低,土壤真菌香农指数的多样性效应值显著增加,随着纬度增加,土壤真菌香农指数的多样性效应值显著降低(表1)。
纬度、土壤有机碳和全氮对土壤真菌香农指数的多样性效应值变化解释权重最大(表2)。真菌香农指数对树种多样性的响应在各气候带均受到土壤有机碳的一定影响,在暖温带中同时受到土壤全氮的较大影响。在热带和中温带森林中土壤真菌香农指数多样性效应值受到土壤pH的作用权重较大,而在亚热带森林中,年平均温度在解释土壤真菌香农指数多样性效应值变化中占了重要比重。
3. 结论与讨论
本研究基于中国热带、亚热带、暖温带和中温带森林生态系统,收集了214组样地数据并进行整合分析,探索不同气候带森林生态系统树种多样性对土壤碳、氮含量的影响,解析树种与土壤真菌多样性的级联效应及其调控土壤碳、氮含量的微生物学机制。主要研究结果表明,中国森林树种多样性对土壤碳、氮含量的影响,除了受到地形和土壤理化性质等非生物因素的影响,还受到土壤真菌多样性的调控,并且后者的作用存在生态系统差异性。这些研究结果能够加深我们对中国森林生态系统树种多样性与生态系统功能关系的理解,助力森林生物多样性保护与“双碳目标”等国家重大战略需求。
3.1 不同气候带森林土壤碳、氮含量对树种多样性的差异性响应
已有结果表明,树种多样性的增加能够显著提高植物生产力,增加土壤有机碳等生态系统碳储量,同时在湿润气候条件下树种多样性效应可能更强[16−18]。这与本研究热带和亚热带森林中,随着树种多样性增加,土壤碳、氮含量逐渐增加的结果一致,说明树种多样性增加能够有效促进热带或亚热带森林土壤碳的累积。然而,在中国暖温带和中温带森林生态系统,土壤碳、氮含量对树种多样性的响应与全球尺度整合分析结果呈现出的趋势不同。说明地域差异可能引起环境因素的改变,导致土壤碳、氮储量对树种多样性变化的差异性响应。由此可以看出,不同气候带森林生态系统土壤碳、氮含量受其特定的生物与非生物环境的影响。
土壤本底资源的丰富程度是森林树种多样性对土壤碳含量效应值的重要影响因素之一[19−20]。一般而言,在土壤相对肥沃的区域,森林土壤所受到的树种多样性效应可能存在较大的不确定性。一方面,树种多样性的增加可以改善生物与非生物环境,促进土壤有机碳的降解,从而降低土壤有机碳含量;另一方面,树种多样性也可能对土壤碳含量无显著影响[21]。然而,在土壤贫瘠的地区,树种多样性往往会增强树种间的资源利用的互补性,促进森林生产力,增加凋落物等有机养分输入量,从而同步促进土壤碳的累积[19−20]。在中国热带和亚热带,尤其是热带森林生态系统,由于降雨的淋洗作用,土壤有机养分大量流失,导致土壤环境相对贫瘠。这种贫瘠的土壤条件限制了热带森林土壤生物的活性与多样性。然而,随着树种多样性的增加,输入土壤的资源多样性也随之增加,这不仅促进了土壤生物对资源的互补利用,还提高了土壤有机养分的周转与累积[22]。在暖温带和中温带森林生态系统,土壤养分含量较高,随着树种多样性增加,植物对土壤养分资源的利用竞争可能会加剧,从而限制养分资源的进一步增加。这可能导致土壤有机养分的减少,进而影响土壤碳、氮含量[16]。
3.2 调控森林树种多样性效应的非生物因素
在本研究中,森林生态系统树种多样性对土壤碳、氮含量的影响受到土壤pH和海拔等非生物因素的影响较大,这与以往的研究结果[23−25]相似。
土壤pH的变化对土壤碳、氮养分周转和累积的影响是至关重要的。一方面,土壤pH可以通过影响地上植被生长或群落组成而改变土壤碳储量[26];另一方面,土壤pH能够调控土壤微生物量、活性和多样性,进而影响土壤碳、氮周转过程及其储量[27]。本研究发现较高的森林土壤pH值条件下,树种多样性对土壤碳、氮含量的作用更强。在不同气候带,亚热带和暖温带森林土壤有机碳的多样性效应受土壤pH的影响较为突出;而暖温带和中温带土壤有机氮的多样性效应受土壤pH的调控则更强。
海拔是温度、水分和光照条件等多种环境因子的综合体,其产生的环境变化影响土壤碳、氮对树种多样性的响应与适应。海拔能够重新分配热量和降水,并使土壤养分有效性呈现梯度分布[28−32],这可能造成不同海拔高度森林土壤有机碳含量对树种多样性表现出差异性响应。本研究发现,随着海拔增加,树种多样性对土壤碳、氮含量和土壤碳氮比的影响均呈现递减趋势。这表明,低海拔森林树种多样性对土壤碳、氮含量的作用更强。低海拔森林生态系统通常具有更高树种的多样性和更充沛的水热条件,这些条件有利于树木的生长,进而增加凋落物种类和数量[33]。物理过程(如降雨淋洗)能够进一步提升低海拔森林凋落物的质量和数量,从而促进土壤碳和氮含量的累积[34−35]。
3.3 森林树种多样性与真菌多样性对土壤碳、氮含量的耦合调控机制
植物多样性假说指出,树种多样性的增加能够促进包括真菌在内的土壤微生物多样性的提升,这在已有的一些研究中已得到验证[36]。土壤真菌多样性的增加能够增强土壤有机养分的互补利用,加速土壤碳、氮养分的周转,改变土壤碳的固持[37−38]。不同气候带森林树种多样性不仅可以通过改变环境因素影响土壤碳、氮含量,也可以通过改变土壤真菌多样性来调控土壤有机碳、氮的累积。
本研究发现,尽管基于总体数据,森林生态系统土壤真菌多样性并未改变土壤碳、氮含量的多样性效应,但是从变量的解释权重来看,暖温带森林树种多样性通过改变土壤真菌多样性而作用于土壤碳含量。暖温带树种多样性可能通过增加土壤真菌多样性,促进其对土壤有机碳养分互补性利用,从而使土壤有机碳含量降低。此外,基于总体数据的分析发现,森林土壤碳氮比的树种多样性效应值随土壤真菌多样性的增加而增加,说明树种和土壤真菌多样性在影响土壤碳氮比上也存在较强的耦合作用。树种多样性对土壤真菌群落的影响有直接和间接途径。树种多样性可以改变凋落物的质量和数量,直接改变真菌分解底物,进而影响土壤真菌多样性。树种多样性的增加往往也有利于改善土壤微环境,为更多的真菌种类提供有利的生长环境[12]。土壤真菌群落的变化会进一步影响土壤有机碳的周转、利用与稳定性[39]。真菌群落对环境变化敏感,当群落稳定性较低时,真菌会降低对土壤碳的分解速率,并通过增加对氮的利用维持群落更替[40]。树种多样性增加显著提升了生态系统功能,使环境更加稳定,进而提高了植物生产力和根系的质量与数量,增强土壤有机碳稳定性。同时,土壤微生物群落的活性及组成也受到影响,其生长与周转速度加快,导致大量真菌和其他微生物类群生物量以残体碳的形态被固存,促进了土壤碳库的积累[41−42]。然而,本研究还表明,尽管热带和亚热带森林树种多样性对土壤碳、氮含量的影响较为强烈,但这些影响主要受海拔和土壤pH等非生物因素的调控,并非源于植物与土壤真菌多样性的级联效应。
近年来,森林的树种多样性及其碳汇功能的影响因素和维持机制受到广泛关注。森林微生物群落功能,以及森林植物与土壤微生物、碳、氮的相互作用仍是当前研究的热点问题之一[43−45]。当前全球尺度整合分析发现,树种多样性和土壤微生物均对土壤有机碳含量及其稳定性产生一定的影响,土壤真菌与树种多样性的纬度格局存在相关关系[39,46]。然而,大尺度环境下树种与土壤真菌的级联效应对土壤碳、氮含量的调控机制的研究仍较为匮乏。大量样地试验发现,不同树种多样性样地的土壤碳、氮含量对其驱动因子的响应不完全相同,得到的结论并不一致,甚至截然相反[47−48]。本研究揭示了中国不同气候带森林生态系统的土壤碳、氮含量对树种多样性存在明显的差异性响应。环境因素和土壤真菌多样性对这种响应的调控强度和机制也存在显著差异,这在一定程度上解释了局域尺度研究中森林树种多样性对土壤碳库影响机制的分歧。尽管这些研究结果有助于增强我们对森林生态系统中植物多样性、真菌多样性与土壤碳、氮含量间互作关系的理解,但由于相关数据的局限性,要明确各气候带森林生态系统树种多样性对土壤碳、氮含量的调控机制及其影响因素,仍需要进一步扩大数据收集范围,并进行更深入的研究。
-
![]()
图 1 森林生态系统树种多样性对土壤碳、氮含量及土壤真菌多样性的影响
土壤有机碳、土壤全氮的单位均为g/kg,pH、碳氮比和真菌香农指数无量纲。*表示在0.05水平上存在显著差异。ns表示差异不显著。
Figure 1. Effects of tree species diversity on soil carbon, nitrogen contents, and soil fungal diversity in forest ecosystems
![]()
图 2 不同森林生态系统树种多样性与土壤碳、氮和真菌多样性的回归分析
Figure 2. Regression analysis of tree species diversity and soil carbon, nitrogen, and fungal diversity in forest ecosystems
![]()
图 3 森林生态系统树种多样性对土壤碳、氮和土壤真菌多样性的累计效应值
方括号内数据是95%的置信区间。*表示影响显著(P<0.05),***表示影响极其显著( P<0.001);ns表示影响不显著。下同。
Figure 3. Cumulative effect values of tree species diversity on soil carbon, nitrogen, and soil fungal diversity in forest ecosystems
表 1 森林生态系统树种多样性效应值的影响因素
Table 1 Influencing factors of tree species diversity effect values in forest ecosystems
指标 土壤有机碳效应值 土壤全氮效应值 土壤碳氮比效应值 土壤真菌香农指数效应值 海拔 −0.010 1*** −0.005 1*** −0.000 3** 0.003 2* 纬度 −0.667 9* −0.627 1*** 0.019 3 −0.553 8** 年平均温度 −0.848 1* −0.201 7 −0.026 9 −0.721 7** 土壤真菌香农指数 0.960 4 −0.256 7 0.097 9* 土壤pH 2.123 1** 2.248 6*** −0.048 7 −0.529 1 土壤有机碳 −0.044 8 土壤全氮 −1.168 5 土壤碳氮比 0.031 2 注: **表示影响极显著(P < 0.01)。  下载: 导出CSV
下载: 导出CSV
表 2 树种多样性效应值影响因子的解释权重
Table 2 Explanatory weight of the factors affecting the effect values of tree species diversity
指标 土壤有机碳效应值 土壤全氮效应值 土壤碳氮比效应值 土壤真菌香农指数效应值 总体 热带 亚热带 暖温带 中温带 总体 热带 亚热带 暖温带 中温带 总体 热带 亚热带 暖温带 中温带 总体 热带 亚热带 暖温带 中温带 海拔 0.993 0.500 0.989 1.000 0.281 1.000 0.500 0.291 0.998 0.986 0.960 0.500 0.279 0.783 0.261 0.690 0.500 0.249 0.306 0.413 土壤pH 0.965 0.141 0.831 1.000 0.194 1.000 0.291 0.386 1.000 0.940 0.352 0.102 0.422 0.272 0.222 0.320 0.722 0.420 0.640 0.725 土壤真菌
香农指数0.420 0.078 0.271 1.000 0.354 0.323 0.085 0.586 0.335 0.964 0.713 0.090 0.259 0.726 0.187 年平均温度 0.500 0.500 0.240 0.379 0.433 0.370 0.500 0.681 0.272 0.431 0.590 0.500 0.244 0.349 0.285 0.410 0.500 0.953 0.339 0.392 纬度 0.545 0.996 0.623 0.970 土壤有机碳 0.960 0.738 0.960 0.966 0.559 土壤全氮 0.896 0.120 0.290 0.834 0.471 土壤碳氮比 0.326 0.044 0.303 0.317 0.325
下载: 导出CSV
-
[1] Hansen M C, Potapov P V, Moore R, et al. High-resolution global maps of 21st-century forest cover change[J]. Science, 2013, 342: 850−853. doi: 10.1126/science.1244693
[2] Food and Agriculture Organization of the United Nations, Forestry Department. Global forest resources assessment 2020: main report[M]. Rome: Food and Agriculture Organization of the United Nations, 2020.
[3] Huang L F, Zhu Z C, Huang M T, et al. Projection of gross primary productivity change of global terrestrial ecosystem in the 21st century based on optimal ensemble averaging of CMIP6 models[J]. Climate Change Research, 2021, 17(5): 514−524.
[4] Liu X J, Trogisch S, He J S, et al. Tree species richness increases ecosystem carbon storage in subtropical forests[J/OL]. Biological Sciences, 2018, 285: 20181240[2023−02−12]. https://doi.org/10.1098/rspb.2018.1240.
[5] Qiao X T, Geng Y, Zhang C, et al. Spatial asynchrony matters more than alpha stability in stabilizing ecosystem productivity in a large temperate forest region[J]. Global Ecology and Biogeography, 2022, 31(6): 1133−1146. doi: 10.1111/geb.13488
[6] Chen X L, Chen H Y, Chen C, et al. Effects of plant diversity on soil carbon in diverse ecosystems: a global meta-analysis[J]. Biological Reviews, 2020, 95(1): 167−183. doi: 10.1111/brv.12554
[7] Wang Q, Bai W, Sun Z X, et al. Does reduced intraspecific competition of the dominant species in intercrops allow for a higher population density?[J]. Food and Energy Security, 2021, 10(2): 285−298.
[8] Jia Y F, Zhai G Q, Zhu S S, et al. Plant and microbial pathways driving plant diversity effects on soil carbon accumulation in subtropical forest[J/OL]. Soil Biology & Biochemistry, 2021, 161: 108375[2023−02−12]. https://doi.org/10.1016/j.soilbio.2021.108375.
[9] Gillespie L M, Hättenschwiler S, Milcu A, et al. Tree species mixing affects soil microbial functioning indirectly via root and litter traits and soil parameters in European forests[J]. Functional Ecology, 2021, 35: 2190−2204. doi: 10.1111/1365-2435.13877
[10] Domeignoz-Horta L A, Pold G, Liu X J, et al. Microbial diversity drives carbon use efficiency in a model soil[J/OL]. Nature Communications, 2020, 11, 3684[2020−07−23]. https://doi.org/10.1038/s41467-020-17502-z.
[11] Wu Y T, Deng M F, Huang J S, et al. Global patterns in mycorrhizal mediation of soil carbon storage, stability, and nitrogen demand: a meta-analysis[J/OL]. Soil Biology & Biochemistry, 2022, 166, 108578[2022−03−01]. https://doi.org/10.1016/j.soilbio.2022.108578.
[12] Wei Y Q, Xiong X, Ryo M, et al. Repeated litter inputs promoted stable soil organic carbon formation by increasing fungal dominance and carbon use efficiency[J]. Biology and Fertility of Soils, 2022, 58(6): 619−631. doi: 10.1007/s00374-022-01647-8
[13] Jing X, Chen X, Fang J Y, et al. Soil microbial carbon and nutrient constraints are driven more by climate and soil physicochemical properties than by nutrient addition in forest ecosystems[J/OL]. Soil Biology & Biochemistry, 2020, 141[2020−02−01]. https://doi.org/10.1016/j.soilbio.2019.107657.
[14] Li J, Pei J, Pendall E, et al. Rising temperature may trigger deep soil carbon loss across forest ecosystems[J/OL]. Advanced Science, 2020, 6; 7(19): 2001242[2023−02−12]. https://doi.org/10.1002/advs.202001242.
[15] Wolfgang V. Conducting meta-analyses in R with the metafor package[J]. Journal of Statistical Software, 2010, 36: 1−48.
[16] Lange M, Eisenhauer N, Sierra C, et al. Plant diversity increases soil microbial activity and soil carbon storage[J/OL]. Nature Communications, 2015, 6, 6707[2015−04−07]. https://doi.org/10.1038/ncomms7707.
[17] Cong W F, van Ruijven J, Mommer L, et al. Plant species richness promotes soil carbon and nitrogen stocks in grasslands without legumes[J]. Journal of Ecology, 2014, 102(5): 1163−1170. doi: 10.1111/1365-2745.12280
[18] Xu S, Eisenhauer N, Ferlian O, et al. Species richness promotes ecosystem carbon storage: evidence from biodiversity-ecosystem functioning experiments[J/OL]. Proceedings of the Royal Society B-Biological Sciences, 2020, 287: 20202063[2020−11−25]. https://doi.org/10.1098/rspb.2020.2063.
[19] Hong S, Yin G, Piao S, et al. Divergent responses of soil organic carbon to afforestation[J]. Nature Sustainability, 2020, 3: 694−700. doi: 10.1038/s41893-020-0557-y
[20] Sayer E, Heard M, Grant H, et al. Soil carbon release enhanced by increased tropical forest litterfall[J]. Nature Climate Change, 2011, 1: 304−307. doi: 10.1038/nclimate1190
[21] Crowther T W, Riggs C, Lind E M, et al. Sensitivity of global soil carbon stocks to combined nutrient enrichment[J]. Ecology Letters, 2019, 22: 936−945. doi: 10.1111/ele.13258
[22] Wang Y F, Du J Q, Pang Z, et al. Unimodal productivity-biodiversity relationship along the gradient of multidimensional resources across Chinese grasslands[J]. National Science Review, 2022, 9(12): 1−14.
[23] 李婷婷, 唐永彬, 周润惠, 等. 云顶山不同人工林林下植物多样性及其与土壤理化性质的关系[J]. 生态学报, 2021, 41(3): 1168−1177. Li T T, Tang Y B, Zhou R H, et al. Understory plant diversity and its relationship with soil physicochemical properties in different plantations in Yunding Mountain[J]. Acta Ecologica Sinica, 2021, 41(3): 1168−1177.
[24] Palandrani C, Alberti G. Tree derived soil carbon is enhanced by tree species richness and functional diversity[J]. Plant and Soil, 2020, 446: 457−469. doi: 10.1007/s11104-019-04381-7
[25] 徐文仕. 植物多样性对亚热带森林土壤可溶性有机碳氮含量的影响[D]. 上海: 华东师范大学, 2022. Xu W S. Effects of plant diversity on soil labile carbon and nitrogen contents in a subtropical forest[D]. Shanghai: East China Normal University, 2022.
[26] Campo J, Merino A. Variations in soil carbon sequestration and their determinants along a precipitation gradient in seasonally dry tropical forest ecosystems[J]. Global Change Biology, 2016, 22(5): 1942−1956. doi: 10.1111/gcb.13244
[27] Chen S P, Wang W T, Xu W T, et al. Plant diversity enhances productivity and soil carbon storage[J]. Proceedings of the National Academy of Sciences, 2018, 115(16): 4027−4032.
[28] 胡靓达, 周海菊, 黄永珍, 等. 不同杉木林分类型植物多样性及其土壤碳氮关系的研究[J]. 生态环境学报, 2022, 31(3): 451−459. Hu L D, Zhou H J, Huang Y Z, et al. A study on plant species diversity and soil carbon and nitrogen in different Cunninghamia lanceolata stand types[J]. Ecology and Environment Sciences, 2022, 31(3): 451−459.
[29] Feng J, Tang M, Zhu B. Soil priming effect and its responses to nutrient addition along a tropical forest elevation gradient[J]. Global Change Biology, 2021, 27: 2793−2806. doi: 10.1111/gcb.15587
[30] 于文睿南, 潘畅, 郭佳欢, 等. 杉木人工林表土有机质含量及其对土壤养分的影响[J]. 中国生态农业学报(中英文), 2021, 29(11): 1931−1939. Yu-Wen R N, Pan C, Guo J H, et al. Topsoil organic matter and its effect on the soil nutrients contents of Cunninghamia lanceolata plantations[J]. Chinese Journal of Eco-Agriculture, 2021, 29(11): 1931−1939.
[31] Fu H, Yuan G, Ge D, et al. Cascading effects of elevation, soil moisture and soil nutrients on plant traits and ecosystem multi-functioning in Poyang Lake wetland, China[J/OL]. Aquatic Sciences, 2020, 82(2) [2020−03−04]. https://doi.org/10.1007/s00027-020-0711-7.
[32] Nobuhiko S, Kiyoshi U, Toshihide H. Plant functional diversity and soil properties control elevational diversity gradients of soil bacteria[J/OL]. FEMS Microbiology Ecology, 2019, 95(4) [2019−02−28]. https://doi.org/10.1093/femsec/fiz025.
[33] Dai Z, Zang H, Chen J, et al. Metagenomic insights into soil microbial communities involved in carbon cycling along an elevation climosequences[J]. Environmental Microbiology, 2021, 23: 4631−4645. doi: 10.1111/1462-2920.15655
[34] Hättenschwiler S. Effects of tree species diversity on litter quality and decomposition[M]// Scherer-Lorenzen M, Körner C, Schulze E D. Forest diversity and function: temperate and boreal systems. Berlin: Springer, 2005: 176.
[35] Handa I, Aerts R, Berendse F, et al. Consequences of biodiversity loss for litter decomposition across biomes[J]. Nature, 2014, 509: 218−221. doi: 10.1038/nature13247
[36] Shen C C, Wang J, He J Z, et al. Plant diversity enhances soil fungal diversity and microbial resistance to plant invasion [J/OL]. Applied and Environmental Microbiology, 2021, 87(11)[2021−05−11]. https://doi.org/10.1128/AEM.00251-21.
[37] Zeng X M, Feng J, Chen J, et al. Microbial assemblies associated with temperature sensitivity of soil respiration along an altitudinal gradient [J/OL]. Science of the Total Environment, 2022, 820, 153257[2022−05−10]. https://doi.org/10.1016/j.scitotenv.2022.153257.
[38] Zhu X F, Jackson R D, de Lucia E H, et al. The soil microbial carbon pump: from conceptual insights to empirical assessments[J]. Global Change Biology, 2020, 26(11): 6032−6039. doi: 10.1111/gcb.15319
[39] Ye X D, Luan J W, Wang H, et al. Tree species richness and N-fixing tree species enhance the chemical stability of soil organic carbon in subtropical plantations [J/OL]. Soil Biology & Biochemistry, 2022, 174, 108828 [2022−11−01]. https://doi.org/10.1016/j.soilbio.2022.108828.
[40] Xie L L, Yin C Y. Seasonal variations of soil fungal diversity and communities in subalpine coniferous and broadleaved forests [J/OL]. Science of the Total Environment, 2022, 846, 157409[2022−11−10]. https://doi.org/10.1016/j.scitotenv.2022.157409.
[41] Orracha S, Gernot B, Christoph R, et al. Fungal biomass and microbial necromass facilitate soil carbon sequestration and aggregate stability under different soil tillage intensities [J/OL]. Applied Soil Ecology, 2022, 179 [2022−11−01]. 104599, https://doi.org/10.1016/j.apsoil.2022.104599.
[42] Xu Y, Gao X, Pei J, et al. Crop root vs. shoot incorporation drives microbial residue carbon accumulation in soil aggregate fractions[J]. Biology and Fertility of Soils, 2022, 58: 843−854. doi: 10.1007/s00374-022-01666-5
[43] Ma X Y, Wang T X, Shi Z, et al. Long-term nitrogen deposition enhances microbial capacities in soil carbon stabilization but reduces network complexity [J/OL]. Microbiome, 2022, 10(1): 112[2022−07−28]. https://doi.org/10.1186/s40168-022-01309-9.
[44] Mishra S, Hättenschwiler S, Yang X D, et al. The plant microbiome: a missing link for the understanding of community dynamics and multifunctionality in forest ecosystems [J/OL]. Applied Soil Ecology, 2020, 145, 103345 [2020−01−01]. https://doi.org/10.1016/j.apsoil.2019.08.007.
[45] Ren C J, Wang J Y, Bastida F, et al. Microbial traits determine soil C emission in response to fresh carbon inputs in forests across biomes[J]. Global Change Biology, 2022, 28(4): 1516−1528. doi: 10.1111/gcb.16004
[46] Zhong Y L, Chu C J, Myers J A, et al. Arbuscular mycorrhizal trees influence the latitudinal beta-diversity gradient of tree communities in forests worldwide [J/OL]. Nature Communications, 2021, 12(1), 3137[2021−05−25]. https://doi.org/10.1038/s41467-021-23236-3.
[47] Craig M E, Geyer K M, Beidler K V, et al. Fast-decaying plant litter enhances soil carbon in temperate forests but not through microbial physiological traits [J/OL]. Nature Communications, 2022, 13, 1229[2022−03−09]. https://doi.org/10.1038/s41467-022-28715-9.
[48] Gutierrez C C, Sánchez F D, Velasco J, et al. Similarity in the difference: changes in community functional features along natural and anthropogenic stress gradients[J]. Ecology, 2015, 96(9): 2458−2466. doi: 10.1890/14-1447.1
-
其他相关附件
-
DOCX格式
2022-0400附录 1 点击下载(44KB)
-
计量
- 文章访问数: 353
- HTML全文浏览量: 112
- PDF下载量: 162
