
Genetic structure variation of Pinus tabuliformis plantations in Beijing and the differences with Shanxi mountain populations
-
摘要:目的本研究旨在揭示与阐明北京主要油松人工林种群的遗传多样性、遗传结构与亲缘关系,引种地对种群遗传结构及生长的影响,尝试追溯其可能的种源,为北京地区油松人工林培育与种质遗传管理提供参考。方法试验采用7对多态性高且扩增稳定的核基因组EST-SSR引物,以北京地区20世纪营造的油松人工林8个种群和皇家园林的古油松3个种群及山西省五大山系的油松种群为试验对象,分析种群遗传多样性、遗传结构和种群间的遗传距离等。结果结果表明:3类种群中,北京人工林种群间遗传结构差异最大(FST = 0.066),山西五山系群体间遗传结构差异次之(FST = 0.033),古油松种群遗传结构差异最小(FST = 0.023);北京油松人工林和古油松种群均偏离Hardy-Weinberg平衡,且呈现杂合子过剩状态,其中古油松种群杂合子过剩更多;在遗传距离0.020处北京人工林、古油松和山西五山系共16个油松种群聚为3类,各种群之间的遗传距离较近,遗传差异较小,相似程度较高;EST-SSR位点J10、J20、J42和J50在北京人工林、古油松和山西五山系中位点扩增频率差别大可用于未知种源的油松种群种源溯源。结论北京油松人工林种群较古油松种群有更丰富的遗传多样性和更大的遗传结构差异;外来种质在引入地胁迫生境下的适合度差异导致部分植株被淘汰,因此保留下来的种群遗传结构发生一定程度的改变;试验为后续北京地区油松人工林的评价、培育和种质种源管理打下了基础。Abstract:ObjectiveThe purpose of this study was to reveal the genetic diversity, genetic structure and genetic relationship of Pinus tabuliformis populations in Beijing, and to explore the impact of introduced land on population genetic structure and growth, and to provide references for the cultivation and germplasm resource management of P. tabuliformis in Beijing.MethodBased on 7 pairs of highly and stable polymorphically nuclear genomic EST-SSR primers, the population genetic diversity, genetic structure and genetic distance were analyzed among 8 P. tabuliformis populations planted in last century in Beijing, 3 ancient P. tabuliformis populations and 5 mountain populations from Shanxi Province of northern China.ResultThe genetic structure difference ( FST = 0.066) was the maximum among Beijing plantation populations, the second was population from Shanxi Province (FST = 0.033), and the lowest was ancient populations (FST = 0.023). Both of the Beijing populations and ancient populations deviated from Hardy-Weinberg equilibrium and showed heterozygote excess, and the maximum heterozygote excess was in ancient P. tabuliformis populations. All of the populations could be clustered into 3 categories when genetic distance was 0.020 based on genetic distance analysis, showing near genetic distances among different populations, the level of evolutionary divergence and genetic differentiation were relatively low. The EST-SSR loci site amplification frequency of J10, J20, J42 and J50 in Beijing populations, ancient P. tabuliformis and Shanxi populations showed significant differences, which could be used for source tracing for unknown provenances.ConclusionThe Beijing populations of P. tabuliformis have richer genetic diversity and higher genetic differentiation than ancient P. tabuliformis; some introduced provenances eventually phased out because of the difference in fitness under stress environment, resulting in the genetic structure has a certain amount of change for the retained population. Our study provides important theoretical foundation for the follow-up studies on the evaluation, cultivation and germplasm management of P. tabuliformis populations in Beijing.
-
土壤微生物作为森林生态系统中最活跃生命组分之一,是地球化学循环的关键环节,在有机质分解、养分元素循环和土壤肥力维持等方面有着重要的作用[1-4]。因此,土壤微生物被认为是评价土壤质量和指示生产力的重要指标。土壤微生物群落对土壤微环境因子变化十分敏感,往往能对土壤水热条件和养分元素浓度等因素的改变迅速做出反应,从而引起微生物群落结构的变化[5-8]。在森林生态系统中,地上植被多样性的变化被认为是影响土壤微生物群落结构的重要因素之一[9-12],不仅可通过改变地上植物生物量、凋落叶和细根数量、组成和分解速率等来直接影响土壤微生物群落[13-15],还可通过影响土壤水热条件、养分元素含量和林下微气候等因子间接影响微生物群落结构[16-18]。关于环境因子对人工林土壤微生物群落调控机制的研究受到越来越多生态学家的关注。
华北落叶松(Larix principis-rupprechtii)是中国北方造林的重要树种之一,在木材生产、水土保持和固碳释氧等方面发挥重要作用[19]。然而,由于不合理经营管理措施引起了诸如土壤肥力退化、人工林养分循环效率低下、林下植物多样性较差等一系列生态问题[20-22]。虽然近年来针对华北落叶松人工林展开了一系列的研究,但多数研究主要集中在土壤肥力、人工林生产力和林地生物多样性等方面[23-25],关于土壤微生物群落结构对林龄梯度变化的响应机制及关键影响因子的研究相对较少。因此,本文以太岳山不同林龄华北落叶松人工林为研究对象,通过磷脂脂肪酸(phospholipid fatty acids,PLFA)法分析其土壤微生物群落结构变化特征,探讨林下植被和土壤理化性质对土壤微生物群落驱动机制,以期进一步了解太岳山不同林龄华北落叶松林土壤微生物群落结构特征和生态功能,为该地区林地资源的可持续经营提供科学的理论依据和实践指导。
1. 研究区概况与研究方法
1.1 研究区概况
本研究位于山西省长治市沁源县好地方林场(36°40′N、112°05′E),该地区年平均气温约为6.1 ℃,最高温在夏季7月份,约为15 ℃,最低温在12月末至1月初,约为−11 ℃。年降雨量为700 mm左右,雨季约从当年5月持续至9月,但降水期多集中于6—8月份。全年无霜期仅有125 d左右,其生长季从4月末5月初持续至10月中上旬。该地区森林类型主要涵盖针叶林、针阔混交林和阔叶林,其中针叶林是该地区森林的主要类型。本项研究选取的华北落叶松人工林样地分别为Lar30、Lar40和Lar50,其中Lar50华北落叶松人工林(林龄为53年)主要在20世纪70年代左右种植,Lar40华北落叶松人工林(林龄为42年)主要在20世纪80年代初种植,Lar30华北落叶松人工林(林龄为33年)是在20世纪90年代初期种植的。栽植所选取的苗木为生长良好,基径超过0.6 cm,苗高在25 cm以上的2年生苗,完成生根粉浸根处理后在山区沿等高线进行栽植。林下主要灌木植物有绣线菊(Spiraea salicifolia)、黄刺玫(Rosa xanthine)和胡枝子(Lespedeza bicolor)等;草本植物主要有唐松草(Thalictrum aquilegiifolium var. sibiricum)、糙苏(Phlomis umbrosa)和玉竹(Polygonatum odoratum)等。
1.2 样地设置和土壤样品采集
2014年7月,以Lar30、Lar40和Lar50华北落叶松人工林群落为研究对象,共设置12块20 m × 30 m的固定样地进行人工林基础信息的采集和记录,包括样地坡向、坡度、海拔、林分密度、林下植被等信息(表1)。林下灌木层和草本层香农多样性指数、丰富度指数和均匀度指数的计算按照Dang等[5]的方法进行。2018年8月份在不同林龄人工林各样地中通过S型采样法对0~10 cm土层进行9点采样。在去除土壤中明显凋落物、石砾和植物残体等杂物后对鲜土进行过筛(土筛孔径为2 mm)并分为2份尽快装入保温箱送回实验室进行相关指标测定。
表 1 不同林龄华北落叶松人工林样地基本情况Table 1. Basic situation of larch plantation sample plots with different forest ages项目
Item林龄梯度 Plantation age gradient Lar30 Lar40 Lar50 林龄/a Stand age
/year33 42 53 坡度 Slope
/(°)25 25 25 坡向
Aspect东南 Southeast 东南 Southeast 东南 Southeast 平均树高 Mean tree height
/m16.3 ± 0.8 18.8 ± 1.3 16.1 ± 1.4 平均胸径 Mean DBH
/cm16.4 ± 0.5 18.7 ± 1.6 22.6 ± 2.8 郁闭度
Canopy density0.28 ± 0.02 0.35 ± 0.01 0.39 ± 0.01 林分密度/(株·hm−2)
Plantation density/(tree·ha−1)1158 ± 87 1341 ± 62 803 ± 24 凋落物层生物量/(t·hm−2) Biomass of litter layer/(t·ha−1) 43.2 ± 10.8 106.9 ± 18.3 50.9 ± 8.2 注:Lar30.林龄33年人工林;Lar40.林龄42年人工林;Lar50.林龄53年人工林。下同。Notes: Lar30, 33-year-old plantation; Lar40, 42-year-old plantation; Lar50, 53-year-old plantation. The same below. 1.3 土壤理化性质测定
将带回实验室土壤中的一份进行自然风干、研磨并根据待测定指标的实验标准进行筛分,另一部分土壤则存入0 ~ 4 ℃条件的冰箱中保存待用。土壤有机碳含量通过重铬酸钾外加热法进行测定[26];土壤全氮和全磷分别采样浓硫酸−混合加速剂消化法和浓硫酸−高氯酸消化法对土样进行消化处理后[26],在连续流动分析仪器上(AA3,SEAL Analytical GmbH,Germany)测定。土壤铵态氮、硝态氮的测定首先称取10 g已经过2 mm土筛的新鲜土样于250 mL广口瓶中,加入1 mol/L氯化钾溶液100 mL,塞紧瓶塞后经过振荡器震荡1 h以上[26]。将浸提液过滤后采用连续流动分析仪器分析土壤铵态氮和硝态氮含量。土壤有效磷含量测定步骤为称取5 g土样并加入0.5 mol/L碳酸氢钠溶液振荡1 h以上,而后将浸提液用无磷滤纸过滤[26],最后通过连续流动分析仪器进行有效磷含量测定[26]。
土壤pH值使用pH计(Five Easy,Mettler-Toledo,Switzerland)测定[26],土壤水分含量的测定则是采用烘干法测定烘干10 g新鲜土样至恒质量后的质量损失来计算土壤水分含量[26]。土壤温度则在采集土样时通过土壤温度测定仪(Li-COR Biosciences,Lincoln,NE,USA)进行测定。
1.4 土壤微生物群落结构
通过PLFA方法对不同林龄土壤微生物群落结构进行测定,主要过程包括从土样中提取脂质、采样碱性甲醇分解并回收甲酯化的脂肪酸样品。而后根据已知含量的甲酯(19∶0)作为内标测定PLFAs含量。其中,革兰氏阳性菌选择 i13∶0,i14∶0,a14∶0,a15∶0,i15∶0,i16∶0,a17∶0,i17∶0,i18∶0 表征[27-28];革兰氏阴性菌选择 10∶0 2OH,15∶1 w4c,15∶1 w6c,16∶0 2OH,16∶1 w6c,16∶1 w7c,16∶1 w9c,cy17∶0,17∶1 w8c,18∶1 w5c,18∶1 w7c, cy19∶0 表征[27, 29-30],真菌选择 16∶1 w5c,18∶1 w9c,18∶2 w6c,18∶2 w9c表征[27, 30-31];放线菌选择 10 Me 16∶0,10 Me 17∶0,10 Me 18∶0表征[31-32]。
1.5 数据处理
采用Microsoft Office Excel 2011软件对相关数据进行初步录入,使用统计软件 SPSS 20.0 对数据进行描述性统计分析和方差分析等,RDA分析在Canoco Software 5. 0中进行,图表绘制通过Sigmaplot 12.0 进行。
2. 结果与分析
2.1 土壤理化性质
土壤pH值变化范围在6.28 ~ 6.56之间,且随着林龄增加表现出逐步上升的变化趋势,特别是Lar50人工林中的土壤pH值相对较高,但不同林龄间土壤pH值差异未达显著水平(P > 0.05)。Lar40和Lar50人工林中土壤含水率均显著高于Lar30人工林(P < 0.05)。土壤温度则随着林龄的增加呈逐步上升的变化趋势,且在Lar50林分中达到峰值。土壤有机碳、全氮、铵态氮和硝态氮含量均在Lar50人工林中显著高于Lar30(P < 0.05),土壤全磷含量在Lar40和Lar50人工林中显著高于Lar30(P < 0.05),但土壤有效磷含量在Lar50人工林中则显著低于Lar30和Lar40(P < 0.05)(表2)。
表 2 不同林龄华北落叶松人工林土壤理化性质Table 2. Soil physicochemical properties of larch plantations with different forest ages土壤理化性质
Soil physical and chemical property林龄梯度
Plantation age gradientLar30 Lar40 Lar50 pH值
pH value6.28 ± 0.09a 6.35 ± 0.11a 6.56 ± 0.08a 含水率
Water content/%51.34 ± 2.54a 60.18 ± 5.16b 65.49 ± 3.78c 温度
Temperature/℃10.95 ± 0.87a 11.75 ± 0.29ab 13.45 ± 0.92b 有机碳
Organic carbon/(g·kg−1)25.12 ± 1.67a 26.88 ± 2.04ab 30.49 ± 2.30b 全氮
Total nitrogen/(g·kg−1)3.84 ± 0.21a 4.11 ± 0.26b 4.39 ± 0.17c 全磷
Total phosphorus/(g·kg−1)0.46 ± 0.02a 0.71 ± 0.11b 0.74 ± 0.03b 硝态氮
Nitrate nitrogen/(mg·kg−1)9.37 ± 1.13a 10.28 ± 0.56ab 12.94 ± 1.86b 铵态氮
Ammonium nitrogen/(mg·kg−1)3.97 ± 0.24a 4.38 ± 0.31b 5.16 ± 0.57c 有效磷
Available phosphorus/(mg·kg−1)4.98 ± 0.07a 4.92 ± 0.39a 4.56 ± 0.28b 注:不同字母表示在不同林龄间存在显著差异(P < 0.05)。Note: different letters mean significant differences among 3 plantation ages (P < 0.05). 2.2 林下植物物种多样性
由表3可知,华北落叶松人工林草本层香农多样性指数、丰富度指数和均匀度指数在Lar40和Lar50人工林中相对较高。灌木层香农多样性指数、丰富度指数和均匀度指数也在Lar50人工林中较高,其次依次为Lar40和Lar30。但总体上看,林龄增长虽然对华北落叶松人工林林下植物物种多样性存在一定促进作用但均未达显著差异水平(P > 0.05)。
表 3 不同林龄林下植物物种多样性Table 3. Species diversity of understory plants at different forest ages林龄梯度
Plantation age gradient草本层多样性
Herb layer diversity灌木层多样性
Diversity of shrub layer香农多样性指数
Shannon index丰富度指数
Richness index均匀度指数
Evenness index香农多样性指数
Shannon index丰富度指数
Richness index均匀度指数
Evenness indexLar30 3.18 ± 0.19a 25.67 ± 4.04a 0.90 ± 0.01a 1.60 ± 0.06a 6.33 ± 2.08a 0.90 ± 0.01a Lar40 3.21 ± 0.10a 28.33 ± 3.51a 0.92 ± 0.02a 1.74 ± 0.08a 6.67 ± 1.53a 0.91 ± 0.02a Lar50 3.23 ± 0.25a 30.67 ± 3.79a 0.94 ± 0.01a 1.83 ± 0.33a 7.00 ± 2.00a 0.91 ± 0.02a 注:不同字母表示各指标在不同林龄处理间存在显著差异(P < 0.05)。Note: different letters indicate that there are significant differences among varied forest age treatments (P < 0.05). 2.3 土壤微生物群落结构
如表4所示:不同林龄对土壤革兰氏阳性细菌、革兰氏阴性细菌、真菌和放线菌含量存在不同程度的影响。具体而言:在不同林龄华北落叶松人工林中,土壤革兰氏阳性细菌含量为4.39 ~ 6.37 nmol/g,Lar50和Lar40林分比Lar30分别高28.02%和45.10%;土壤革兰氏阴性细菌含量为3.52 ~ 4.84 nmol/g,Lar50和Lar40林分比Lar30分别高20.74%和37.50%;土壤放线菌含量为1.60 ~ 2.12 nmol/g,Lar50和Lar40林分比Lar30分别高11.88%和32.50%。土壤真菌含量为0.74 ~ 0.80 nmol/g,但各林龄间差异性未达显著水平。此外,本研究还比较了不同林龄华北落叶松人工林土壤微生物群落革兰氏阳性细菌与革兰氏阴性细菌比以及真菌与细菌比。二者在不同林龄间从高到低依次均为Lar30 > Lar50 > Lar40。
表 4 不同林龄土壤微生物群落特征Table 4. Characteristics of soil microbial community in different forest agesPLFA生物量
PLFA biomass林龄梯度
Plantation age gradientLar30 Lar40 Lar50 革兰氏阳性细菌 Gram-positive bacteria (G+)/(nmol·g−1) 4.39 ± 1.34a 5.62 ± 1.25b 6.37 ± 1.04c 革兰氏阴性细菌 Gram-negative bacteria (G−)/(nmol·g−1) 3.52 ± 1.06a 4.25 ± 1.46b 4.84 ± 1.20c 真菌 Fungi (F)/
(nmol·g−1)0.74 ± 0.08a 0.78 ± 0.03a 0.80 ± 0.04a 放线菌 Actinmycete (ACT)/
(nmol·g−1)1.60 ± 0.25a 1.79 ± 0.46b 2.12 ± 0.34c 革兰氏阳性细菌∶革兰氏阴性细菌 Gram-positive bacteria ∶ Gram-negative bacteria (G+∶G−) 1.42 ± 0.18a 1.26 ± 0.27a 1.39 ± 0.21a 真菌∶细菌
Fungi ∶ bacteria (F∶B)0.11 ± 0.01a 0.08 ± 0.02a 0.09 ± 0.01a 注:不同字母表示不同林龄间存在显著差异(P < 0.05)。Note: different letters indicate significant differences among varied forest ages (P < 0.05). 2.4 不同林龄对土壤微生物群落结构关键影响因子分析
经过置换检验筛选后采用RDA分析,探讨影响土壤微生物群落结构的关键影响因子。由图1可知土壤pH值、含水率、温度、养分元素含量和林下植物物种多样性对微生物群落结构的影响,结果表明:Lar30、Lar40和Lar50林分间土壤微生物群落明显分开,说明林龄增长导致林地土壤微生物群落结构出现明显变化;第1轴和第2轴的特征值分别为0.718和0.143,即共同解释了土壤微生物群落总变异的86.1%。土壤有效磷含量与第1轴呈负相关,土壤含水率、温度、硝态氮含量、灌木层丰富度指数和草本层丰富度指数与第1轴呈正相关,分别解释了微生物群落结构变异程度的11.8%、22.7%、18.4%、10.6%、7.9%和5.6%,这说明土壤含水率、温度、有效磷含量、硝态氮含量、灌木层和草本层丰富度指数是不同林龄人工林表层土壤中微生物群落结构变化的主要影响因子。
![]() 图 1 土壤微生物群落结构影响的冗余分析ST. 土壤温度;SWC. 土壤含水率;SAP. 土壤有效磷;NO3−-N. 土壤硝态氮;SR. 灌木层物种丰富度指数;HR. 草本层物种丰富度指数。ST, soil temperature; SWC, soil water content; SAP, soil available phosphorus; NO3−-N, nitrate nitrogen; SR, richness index of shrub; HR, richness index of herb.Figure 1. Redundancy analysis of relationships of soil microbial community structure to soil factors and understory vegetation diversity
图 1 土壤微生物群落结构影响的冗余分析ST. 土壤温度;SWC. 土壤含水率;SAP. 土壤有效磷;NO3−-N. 土壤硝态氮;SR. 灌木层物种丰富度指数;HR. 草本层物种丰富度指数。ST, soil temperature; SWC, soil water content; SAP, soil available phosphorus; NO3−-N, nitrate nitrogen; SR, richness index of shrub; HR, richness index of herb.Figure 1. Redundancy analysis of relationships of soil microbial community structure to soil factors and understory vegetation diversity3. 讨 论
3.1 林龄对土壤理化性质和林下植物物种多样性影响
土壤作为森林生态系统的重要组成部分,pH值、含水率、温度和养分元素含量等理化性质对人工林中植物生长具有重要作用[15, 22]。前人研究指出林龄变化可以通过影响地上植被对土壤养分利用效率来影响土壤养分浓度[25],同时林分群落结构和物种多样性的差异等也会间接改变诸如透光率、穿透雨量、凋落物产量等林下微环境因素,从而促进土壤养分含量的积累,提高土壤养分水平[6, 17, 33]。在本研究中,Lar50和Lar40林地土壤pH值、含水率、温度和养分元素浓度总体上高于Lar30,表明林龄的增加可能更有利于提高土壤物理性质和促进土壤养分含量积累,这与罗达等[2]和Chen等[34]等在云杉(Picea asperata)人工林和油松(Pinus tabuliformis)人工林中进行的研究结果相似。此外,本研究结果还表明土壤有效磷含量在Lar30和Lar40林地土壤中显著高于Lar50,这可能是由于地上植被对土壤有效磷的需求随林龄增长而增加,从而不利于土壤有效磷含量的积累,因此在Lar50华北落叶松人工林中出现磷缺乏现象,应通过合理施用磷肥以保证林木生长对磷素的需求。
在本研究中,随着人工林林龄增加,林下灌木层和草本层物种多样性总体也呈上升趋势(表3)。通常在人工林幼龄或中龄阶段,林下植被尚处于发展阶段,灌木层和草本层各物种间存在剧烈养分竞争和排斥现象,从而导致林下物种多样性下降[25, 33];随着林龄增加,林分结构逐步趋于稳定,此时各种适应人工林环境的灌木层和草本层物种逐渐共存,林下物种多样性也相对较高[7, 25, 33]。
3.2 土壤理化性质和林下植被对土壤微生物结构的影响
本研究结果表明林龄增加能有效提高人工林土壤中革兰氏阳性细菌、革兰氏阴性细菌和放线菌含量,同时土壤理化性质、灌木层和草本层物种丰富度指数是驱动不同林龄人工林土壤微生物群落结构变化的主要因素。Chen等[34]和Liu等[35]等研究认为,林龄的增长往往有利于提高地上植被生物量、改善土壤水热环境和提高土壤养分水平,从而增加土壤微生物活性,进而提高土壤微生物生物量;且随着林龄的增加,地表凋落物数量、地下细根生物量及其分解效率等均会发生变化,这可能会加快养分元素归还效率并影响土壤中微生物群落结构和各菌群含量,这与本文研究结果相似。Fu等[36]研究提出人工林乔木层物种相对简单,林下植被多样性的变化可能与土壤微生物群落间关系更为密切。多数研究结果也提出林下植物多样性增加会丰富凋落物和细根的组成和质量,且与乔木相比,灌、草凋落叶和细根生物量对微生物群落PLFA含量影响更大,进而促进土壤微生物生物量的积累。此外,林下植被多样性还与根系分泌物数量密切相关,土壤微生物诸如细菌、真菌和放线菌等菌群的活性也受到根系分泌物的影响[33, 37]。本文研究结果也发现不同林龄人工林灌木层和草本层丰富度指数也与土壤微生物群落结构密切相关,这与Fu等[36]研究得出结论相似。
林龄增加总体上提高了土壤微生物菌群丰度,但针对不同林龄人工林土壤真菌与细菌比值和革兰氏阳性细菌与革兰氏阴性细菌比值的研究结果表明:不同林龄人工林土壤真菌与细菌比值在0.08 ~ 0.11之间,表明细菌在该地区华北落叶松人工林土壤微生物群落中占据支配地位。而不同林龄间二者比例的差异未达显著变化水平,这可能是由于与Lar30人工林相比,Lar40和Lar50人工林虽然对土壤养分元素含量存在积极的促进作用,但该地区土壤真菌群落和细菌群落总体上受营养胁迫的程度较小,林龄增长对细菌群落促进作用尽管相对高于真菌,但总体并未对土壤真菌丰度和细菌丰度产生显著影响。不同林龄人工林土壤革兰氏阳性细菌与革兰氏阴性细菌比值则在1.26 ~ 1.42之间,其中Lar50人工林土壤革兰氏阳性细菌与革兰氏阴性细菌比相对高于其余两种人工林,表明在细菌群落中,革兰氏阳性细菌是华北落叶松林土壤细菌的主要菌群。这可能是因为革兰氏阳性细菌对难分解基质利用效率相对较高,使得其在适宜的水热条件和养分情况下能够更有效的生长[1, 6, 10]。
4. 结 论
林龄增加有效改善了土壤水热状况和土壤养分含量,特别是在Lar50人工林中含水率、温度、有机碳、全氮、全磷、硝态氮、铵态氮含量均显著高于Lar30人工林,但有效磷含量则在Lar30和Lar40人工林中则相对较高。此外与Lar30人工林相比,Lar40和Lar50人工林土壤中革兰氏阳性细菌、革兰氏阴性细菌、真菌和放线菌含量较高,但真菌与细菌和革兰氏阳性细菌与革兰氏阴性细菌比值在研究区内分别为0.08 ~ 0.11和1.26 ~ 1.42之间,这表明在太岳山华北落叶松人工林群落中,土壤细菌是占据支配地位的微生物群落,同时革兰氏阳性细菌也是细菌群落中的主要菌群。土壤含水率、温度、有效磷含量、硝态氮含量、灌木层和草本层丰富度指数是不同林龄人工林土壤中微生物群落结构变化的主要影响因子,在分析土壤微生物群落影响因子时,不仅要考虑土壤水热条件和养分含量对土壤微生物群落结构的驱动作用,还需进一步分析地上植被特征对微生物群落的影响。
-
![]()
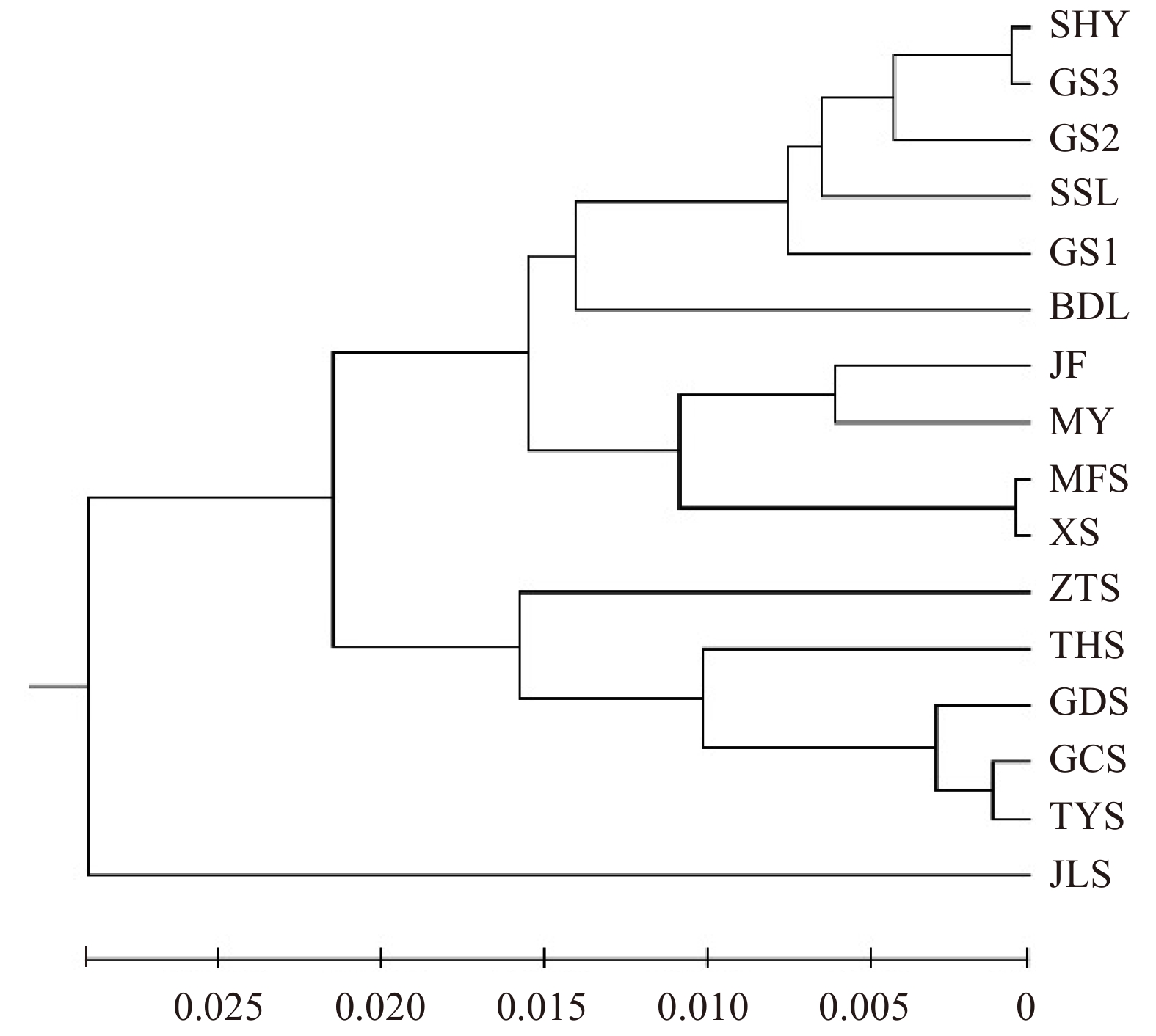
图 1 7对引物分析北京油松种群和山西油松种群的Nei’s遗传距离UPGMA聚类图
Figure 1. UPGMA clustering map of Nei’s genetic distancefor Beijing and Shanxi P. tabuliformis populations analysed by 7 pairs of primers
表 1 供试油松种群基本情况
Table 1 General information of sampling stands of Pinus tabuliformis
项目
Item种群
Population经度
Longitude纬度
Latitude海拔
Altitude/m树龄/a
Stand age/year年均高
Annual average height/cm样本数
Sample plant number北京人
工林
Beijing plantationJF 116°28′ E 39°54′ N 450 69 14.493 36 JLS 115°59′ E 39°54′ N 751 43 24.419 36 MFS 116°01′ E 40°03′ N 180 45 27.333 30 MY 116°49′ E 40°29′ N 75 45 15.778 36 XS 116°05′ E 40°03′ N 145 64 12.969 35 SSL 116°16′ E 40°15′ N 170 30 21.667 30 BDL 115°56′ E 40°22′ N 676 45 17.111 33 SHY 116°32′ E 40°40′ N 600 59 20.339 36 古油松 Ancient P. tabuliformis GS1 116°10′ E 39°59′ N 135 > 100 — 31 GS2 116°14′ E 40°17′ N 180 > 200 — 102 GS3 116°04′ E 39°52′ N 300 > 200 — 18 山西五
山系
Five mountain populations in Shanxi ProvinceGCS 112°01′ E 38°36′ N 1 650 21 24.762 52 GDS 111°42′ E 37°29′ N 1 655 21 40.476 66 THS 113°31′ E 37°39′ N 1 412 21 27.143 32 TYS 112°04′ E 37°37′ N 1 496 21 33.333 20 ZTS 112°01′ E 35°44′ N 1 519 21 32.381 18 注:JF. 鹫峰林场; JLS. 九龙山林场; MFS. 妙峰山林场; MY. 密云水库; XS. 西山林场; SSL. 十三陵林场; BDL. 八达岭林场; SHY. 石湖峪林场; GS1. 以香山公园为中心的皇家园林; GS2. 明十三陵; GS3. 妙峰山; GCS. 管涔山; GDS. 关帝山; THS. 太行山; TYS. 太岳山; ZTS. 中条山. GS1, GS2, GS3树龄过大,暂不考虑胸径与树高的关系。下同。Notes: JF, Jiufeng Forest Farm; JLS, Jiulongshan Forest Farm; MFS, Miaofengshan Forest Farm; MY, Miyun Reservoir; XS, Xishan Forest Farm; SSL, Shisanling Forest Farm; BDL, Badaling Forest Farm; SHY, Shihuyu Forest Farm; GS1, royal garden centered by Xiangshan Park; GS2, Ming Shisanling Tombs; GS3, Miaofengshan Mountain; GCS, Guancenshan Mountain; GDS, Guandishan Mountain; THS, Taihangshan Mountain; TYS, Taiyuanshan Mountain; ZTS, Zhongtiaoshan Mountain. Since GS1, GS2, and GS3 are too old, the relationship between DBH and tree height is not considered. The same below.  下载: 导出CSV
下载: 导出CSV
表 2 油松PCR检测SSR引物
Table 2 SSR primers for PCR detection of P. tabuliformis
引物名称
Primer name前引物序列
Former primer sequence (5′→3′)后引物序列
Back primer sequence (5′→3′)荧光修饰
Fluorescent modification片段长度
Fragment length/bpJ9 GTTTGCAGTGAAAGCATGAAAG GCACCAATTCCTTCTCAAATTC HEX 244 ~ 253 J10 GTCGACACTCCAGGGTAGATTC ATATCATCAGCTAATTGTGCGG TAMRA 254 ~ 257 J20 CACCTCCGTAGTTTGATGTTCC CGATGTATCGTGTACACAGCCT FAM 150 ~ 170 J29 AGTCCGAATGTCTTCTTTCTGC TATGGAACGAATCAGAGATGACG FAM 182 ~ 200 J42 AACCTGTCATCCAGTTCCTGTT TTGTCAAATTCCAATTCAGCAC TAMRA 251 ~ 269 J48 GAAGAGGAAGACGAAATGGATG CTTTACATTTACCGCCTCTGCT ROX 262 ~ 268 J50 TCATCCATTTCAATAGCACGAC GTAGCTGCTTGGCCTGATTATC HEX 235 ~ 244 注:引自参考文献[16]。Note: cited from reference [16].
下载: 导出CSV
表 3 16个油松种群各位点等位标记频率
Table 3 Allele frequency of each locus in the 16 populations of P. tabuliformis
引物 Primer 等位标记 Allele marker 北京人工林
Beijing plantation古油松
Ancient P. tabuliformis山西五山系
Five mountain populations
in Shanxi ProvinceJF JLS MFS MY XS SSL BDL SHY GS1 GS2 GS3 GCS GDS THS TYS ZTS J9 A 0.281 0.375 0.267 0.235 0.242 0.014 0.106 0.043 0.133 0.189 0.222 0.144 0.242 0.219 0.150 0.167 B 0.719 0.625 0.733 0.765 0.758 0.987 0.894 0.957 0.867 0.811 0.778 0.856 0.758 0.781 0.850 0.833 J10 A 0.941 0.985 0.981 0.956 0.982 0.865 0.879 0.917 0.983 0.995 0.972 0.692 0.677 0.609 0.675 0.750 B 0.059 0.015 0.019 0.044 0.018 0.135 0.121 0.083 0.017 0.005 0.028 0.308 0.323 0.391 0.325 0.250 J20 A 0.015 — 0.019 0.106 0.056 0.028 0.016 — 0.035 0.026 — 0.040 0.074 0.048 0.079 0.094 B 0.632 0.786 0.904 0.697 0.852 0.778 0.688 0.889 0.931 0.871 0.941 0.890 0.787 0.774 0.842 0.594 C 0.353 0.214 0.077 0.167 0.093 0.153 0.297 0.111 0.035 0.103 0.059 0.070 0.131 0.129 0.079 0.313 D — — — 0.030 — 0.042 — — — — — — 0.008 0.048 — — J29 A — — — 0.015 — 0.015 0.206 0.063 0.024 0.014 — 0.010 0.015 — — — B 1.000 0.881 1.000 0.970 1.000 0.941 0.677 0.906 0.857 0.892 1.000 0.933 0.879 0.984 0.950 0.944 C — 0.095 — 0.015 — — 0.118 0.031 0.119 0.081 — 0.048 0.099 — 0.050 0.056 D — 0.024 — — — 0.044 — — — 0.014 — 0.010 0.008 0.016 — — J42 A 0.167 0.500 0.017 0.329 — 0.014 0.210 0.014 0.050 0.015 0.028 0.010 — 0.016 — — B 0.046 — 0.017 0.014 0.016 0.069 0.032 0.014 0.050 0.069 0.056 0.039 0.048 0.177 0.025 0.028 C 0.015 — 0.017 0.043 0.047 0.264 0.210 0.306 0.383 0.157 0.250 0.206 0.318 0.403 0.225 0.222 D 0.773 0.500 0.948 0.614 0.938 0.653 0.548 0.667 0.517 0.760 0.667 0.745 0.635 0.403 0.750 0.750 J48 A 0.206 0.208 0.100 0.177 0.132 0.176 0.197 0.194 0.242 0.142 0.222 0.200 0.269 0.313 0.290 0.028 B 0.721 0.736 0.800 0.691 0.765 0.757 0.727 0.722 0.710 0.784 0.722 0.750 0.723 0.688 0.684 0.972 C 0.074 0.056 0.100 0.132 0.103 0.068 0.076 0.083 0.048 0.074 0.056 0.050 0.008 — 0.026 — J50 A 0.221 0.157 0.232 0.177 0.206 0.216 0.242 0.250 0.183 0.314 0.361 0.125 0.156 0.188 0.175 0.167 B 0.779 0.843 0.768 0.824 0.750 0.743 0.758 0.722 0.817 0.686 0.639 0.875 0.828 0.797 0.825 0.833 C — — — — 0.029 — — — — — — — 0.008 0.016 — — D — — — — 0.015 0.041 — 0.028 — — — — 0.008 — — — 合计Total 23 17 16 17 20 18 21 19 19 19 19 16 20 22 19 17 16 注:各等位SSR标记排序按照扩增产物片段从小到大的顺序;山西省五山系油松地理种群的各位点多样性数据来源于武文斌试验原始数据[13]。Notes: each allelic SSR markers are sorted according to the order of amplification product fragments from small to large; the diversity data of the geographical distribution of P. tabuliformis in Shanxi Province are derived from the original data of professor Wu Wenbin[13].
下载: 导出CSV
表 4 16个油松种群遗传多样性
Table 4 Genetic diversity of 16 P. tabuliformis populations
项目 Item 种群 Population Na Ne I Ho He D 北京人工林
Beijing plantationJF 2.429 1.513 0.503 0.367 0.310 0.184 JLS 2.286 1.536 0.503 0.417 0.323 0.291 MFS 2.429 1.297 0.354 0.237 0.204 0.162 MY 2.857 1.569 0.562 0.343 0.327 0.049 XS 2.571 1.341 0.406 0.210 0.227 − 0.075 SSL 3.000 1.479 0.528 0.210 0.294 − 0.286 BDL 2.714 1.732 0.661 0.271 0.397 − 0.317 SHY 2.857 1.437 0.482 0.414 0.278 0.489 极值比
Extremum ratio1.312 1.335 1.867 1.985 1.946 — 古油松
Ancient P. tabuliformisGS1 2.857 1.487 0.496 0.239 0.285 − 0.161 GS2 2.857 1.423 0.488 0.310 0.277 0.119 GS3 2.286 1.467 0.449 0.303 0.227 0.094 极值比
Extremum ratio1.249 1.044 1.104 1.297 1.028 — 山西五山系
Five mountain populations in Shanxi ProvinceGCS 2.857 1.439 0.504 0.302 0.291 0.038 GDS 3.143 1.608 0.608 0.315 0.370 − 0.149 THS 2.714 1.731 0.617 0.400 0.382 0.047 TYS 2.429 1.494 0.517 0.428 0.321 0.333 ZTS 2.286 1.479 0.479 0.311 0.297 0.047 极值比
Extremum ratio1.374 1.202 1.288 1.417 1.312 — 注:Na. 观测等位标记数;Ne. 有效等位标记数;I. Shannon’s信息指数;Ho. 观测杂合度;He. 期望杂合度;D. Hardy-Weinberg平衡偏离指数;极值比:在同一大类地理种群中某一参数值的最大值与最小值的比值。Notes: Na, number of observed alleles; Ne, effective alleles; I, Shannon’s information index; Ho, observed heterozygosity; He, expected heterozygosity; D, Hardy-Weinberg balance deviation index; extremum ratio: ratio of the maximum value to the minimum value of a parameter value in the same large geographic population.
下载: 导出CSV
表 5 16个油松种群中的F统计量
Table 5 F-statistics in 16 P. tabuliformis populations
参数
ParameterJ9 J10 J20 J29 J42 J48 J50 平均值 Mean 北京人工林 Beijing plantation 古油松
Ancient P. tabuliformis山西五山系
Five mountain populations
in Shanxi ProvinceFIS − 0.282 − 0.360 0.217 0.228 − 0.057 − 0.193 0.016 − 0.076 − 0.071 − 0.102 − 0.068 FIT − 0.217 − 0.155 0.266 0.287 0.068 − 0.162 0.005 − 0.001 − 0.001 − 0.077 − 0.033 FST 0.051 0.151 0.062 0.078 0.119 0.026 0.021 0.070 0.066 0.023 0.033 Nm 4.657 1.402 3.763 2.973 1.860 9.328 11.593 3.314 3.552 10.806 7.311 注:FIS. 群体内的固定指数;FIT. 总群体的固定指数;FST. 群体间遗传分化系数;Nm. 基因流。Notes: FIS, fixed index in population; FIT, fixed index of total population; FST, coefficient of genetic differentiation among populations ; Nm, gene flow.
下载: 导出CSV
表 6 北京11个油松种群和山西五山系油松种群的遗传一致度和Nei’s遗传距离
Table 6 Genetic identity and genetic distance of 11 P. tabuliformis populations in Beijing and P. tabuliformis populations in five mountain populations in Shanxi Province
种群 Population JF JLS MFS MY XS SSL BDL SHY GS1 GS2 GS3 GCS GDS THS TYS ZTS JF — 0.971 0.981 0.988 0.983 0.967 0.966 0.962 0.950 0.977 0.969 0.960 0.958 0.931 0.960 0.970 JLS 0.030 — 0.951 0.986 0.949 0.932 0.956 0.935 0.946 0.952 0.948 0.932 0.934 0.911 0.927 0.927 MFS 0.019 0.051 — 0.975 0.999 0.969 0.950 0.973 0.961 0.991 0.983 0.971 0.959 0.922 0.967 0.962 MY 0.012 0.014 0.026 — 0.976 0.969 0.972 0.967 0.966 0.976 0.970 0.962 0.958 0.936 0.959 0.958 XS 0.017 0.053 0.001 0.024 — 0.974 0.953 0.976 0.964 0.992 0.983 0.973 0.962 0.926 0.970 0.964 SSL 0.034 0.070 0.032 0.031 0.027 — 0.978 0.996 0.983 0.984 0.981 0.984 0.978 0.965 0.983 0.976 BDL 0.034 0.046 0.051 0.028 0.048 0.022 — 0.979 0.973 0.972 0.961 0.962 0.963 0.942 0.959 0.964 SHY 0.039 0.067 0.028 0.033 0.024 0.004 0.021 — 0.991 0.989 0.989 0.982 0.976 0.959 0.980 0.964 GS1 0.059 0.055 0.040 0.035 0.037 0.017 0.028 0.009 — 0.982 0.985 0.972 0.974 0.960 0.970 0.948 GS2 0.023 0.049 0.009 0.025 0.008 0.016 0.029 0.011 0.018 — 0.994 0.973 0.968 0.939 0.970 0.965 GS3 0.032 0.053 0.018 0.030 0.017 0.019 0.040 0.012 0.016 0.006 — 0.970 0.969 0.953 0.971 0.951 GCS 0.041 0.070 0.029 0.039 0.027 0.016 0.039 0.019 0.028 0.027 0.030 — 0.994 0.975 0.998 0.976 GDS 0.043 0.069 0.041 0.043 0.039 0.022 0.038 0.025 0.027 0.033 0.032 0.006 — 0.987 0.995 0.976 THS 0.072 0.093 0.081 0.066 0.077 0.036 0.060 0.042 0.041 0.063 0.049 0.025 0.013 — 0.978 0.953 TYS 0.041 0.076 0.033 0.042 0.030 0.018 0.042 0.020 0.031 0.030 0.030 0.002 0.005 0.022 — 0.972 ZTS 0.030 0.076 0.039 0.043 0.036 0.025 0.037 0.036 0.053 0.036 0.050 0.025 0.024 0.048 0.029 — 注:对角线以上为遗传相似度,对角线以下为遗传距离。Notes: above diagonal represents Nei’s genetic identity, below diagonal represents genetic distance.
下载: 导出CSV
-
[1] White T L, Adams W T, Neale D B. Forest genetics[M]. Cambridge: CABI Publishing, 2007.
[2] Wilkinson D M. Is local provenance important in habitat creation?[J]. Journal of Applied Ecology, 2001, 38(6): 1371−1373.
[3] Hugron S, Bussières J, Rochefort L. Tree plantations within the context of ecological restoration of peatlands: a practical guide[M]. Québec: Université Laval, 2013.
[4] 徐化成. 油松[M]. 北京: 中国林业出版社, 1993. Xu H C. Chinese pine[M]. Beijing: China Forestry Publishing House, 1993.
[5] 毛爱华, 李建祥, 张超英, 等. 19年生侧柏种源变异及选择研究[J]. 北京林业大学学报, 2010, 32(1):63−68. Mao A H, Li J X, Zhang C Y, et al. Geographic variation and provenance selection of Platycladus orientalis in a 19-year-old testing plantation[J]. Journal of Beijing Forestry University, 2010, 32(1): 63−68.
[6] 徐化成, 孙肇凤, 郭广荣, 等. 油松天然林的地理分布和种源区的划分[J]. 林业科学, 1981, 17(3):258−270. Xu H C, Sun Z F, Guo G R, et al. Geographic distribution of Pinus tabuliformis Carr. and classification of provenance regions[J]. Scientia Silvae Sinicae, 1981, 17(3): 258−270.
[7] 徐化成, 郭广荣, 冯林, 等. 油松天然林的生长与地理-气候因素的关系[J]. 北京林学院学报, 1981, 3(4):9−13. Xu H C, Guo G R, Feng L, et al. The relativity between natural Pinus tabuliformis forest growth and geographical climatic factors[J]. Journal of Beijing Forestry University, 1981, 3(4): 9−13.
[8] 云希和. 油松在北京平地栽植问题的初步探讨[J]. 园艺学报, 1965, 4(4):230−232. Yun X H. Preliminary discussion on the cultivation of Pinus tabuliformis in Beijing[J]. Journal of Horticulture, 1965, 4(4): 230−232.
[9] 史宇, 余新晓, 张佳音, 等. 北京山区油松人工林单木材积生长量BP神经网络模型[J]. 东北林业大学学报, 2010, 38(2):20−22. doi: 10.3969/j.issn.1000-5382.2010.02.008 Shi Y, Yu X X, Zhang J Y, et al. BP neural network model for volume growth of single trees in Pinus tabulaeformis plantations in Beijing mountain area[J]. Journal of Northeast Forestry University, 2010, 38(2): 20−22. doi: 10.3969/j.issn.1000-5382.2010.02.008
[10] 《北京市林业志》编委会. 北京林业志[M]. 北京: 中国林业出版社, 1993. Editorial Board of Beijing Forestry Journal. Beijing forestry annals[M]. Beijing: China Forestry Publishing House, 1993.
[11] 张新波, 李悦, 袁虎威, 等. 山西油松天然林分21年子代生长性状遗传变异研究[J]. 北京林业大学学报, 2014, 36(3):104−109. Zhang X B, Li Y, Yuan H W, et al. Genetic variations of growth traits in a 21-year-old stand progeny of Shanxi natural Pinus tabuliformis forests[J]. Journal of Beijing Forestry University, 2014, 36(3): 104−109.
[12] 富裕华, 饶九欢, 周学仁, 等. 山西油松种群特点和亲子代性状的变异[J]. 山西农业科学, 1995, 23(3):40−45. Fu Y H, Rao J H, Zhou X R, et al. The characteristics of species colony and character variation of parents and filial generation of Pinus tabulaemis in Shanxi provindce[J]. Shanxi Agricultural Sciences, 1995, 23(3): 40−45.
[13] 武文斌, 贺快快, 狄皓, 等. 基于SSR标记的山西省油松山脉地理种群遗传结构与地理系统[J]. 北京林业大学学报, 2018, 40(10):51−59. Wu W B, He K K, Di H, et al. Genetic structure and geographic system of geographical population of Pinus tabuliformis mountain range based on SSR in Shanxi Province of northern China[J]. Journal of Beijing Forestry University, 2018, 40(10): 51−59.
[14] 张新波, 刘海荣, 毕理智, 等. 21年生油松种源性状变异和选择的研究[J]. 山西林业科技, 2013, 42(1):20−23, 39. doi: 10.3969/j.issn.1007-726X.2013.01.007 Zhang X B, Liu H R, Bi L Z, et al. Variation and choice of provenance trait of 21 year Pinus tabuliformis[J]. Shanxi Forestry Science and Technology, 2013, 42(1): 20−23, 39. doi: 10.3969/j.issn.1007-726X.2013.01.007
[15] 王春玲, 王久丽. 北京市古树名木管理系统介绍[J]. 中国花卉园艺, 2008(4):16−17. Wang C L, Wang J L. Introduction of Beijing ancient and famous tree management system[J]. Chinese Flower Gardening, 2008(4): 16−17.
[16] 程祥, 张梅, 毛建丰, 等. 有限种群油松种子园的遗传多样性与交配系统[J]. 北京林业大学学报, 2016, 38(9):8−15. Cheng X, Zhang M, Mao J F, et al. Gene diversity and mating system of Pinus tabuliformis in finite population seed orchard[J]. Journal of Beijing Forestry University, 2016, 38(9): 8−15.
[17] Wright S. Evolution and the genetics of population: variability within and among natural populations[M]. Chicago: University of Chicago Press, 1978.
[18] Vendramin G G, Fady B, Gonzalez-Martinez S C, et al. Genetically depauperate but widespread: the case of an emblematic Mediterranean pine[J]. Evolution, 2008, 62(3): 680−688. doi: 10.1111/j.1558-5646.2007.00294.x
[19] 滑留帅, 陈宏, 杨晓冰, 等. 固原本地黄牛及其利杂群体PIT-1基因多态性与育肥性状的关系[J]. 西北农林科技大学学报(自然科学版), 2007, 35(12):1−5. Hua L S, Chen H, Yang X B, et al. Association of polymorphism of Pit-1 gene with fattening traits in Guyuan local cattle and crossbreed of Limousin
× local cattle[J]. Journal of Northwest A&F University (Natural Science Edition), 2007, 35(12): 1−5. [20] Valente C, Alvarez L, Marques P I, et al. Genes from the TAS1R and TAS2R families of taste receptors: looking for signatures of their adaptive role in human evolution[J]. Genome Biology & Evolution, 2018, 10(4): 1139−1152.
[21] Bulmer M G. The maintenance of the genetic variability of polygenic characters by heterozygous advantage[J]. Genetical Research, 1973, 22(1): 9−12. doi: 10.1017/S0016672300012799
[22] Amandine D, Anne V, Magali F, et al. Progressive ataxia of Charolais cattle highlights a role of KIF1C in sustainable myelination[J/OL]. PLoS Genetics, 2018, 14(8): e1007550 [2019−09−30]. https://doi.org/10.1371/journal.pgen.1007550.
[23] 许玉兰, 蔡年辉, 白青松, 等. 基于微卫星分子标记的云南松及其近缘种遗传关系分析[J]. 西南林业大学学报, 2017, 37(1):1−9. doi: 10.11929/j.issn.2095-1914.2017.01.001 Xu Y L, Cai N H, Bai Q S, et al. Genetic relationship of Pinus yunnanensis with related pine species based on microsatellite molecular markers[J]. Journal of Southwest Forestry University, 2017, 37(1): 1−9. doi: 10.11929/j.issn.2095-1914.2017.01.001
[24] Li J J, Xiong C, He X, et al. Using SSR-HRM to identify closely related species in herbal medicine products: A case study on licorice[J/OL]. Frontiers in Pharmacology, 2018, 9: 407 [2019−09−10]. http://www.frontiersin.org/articles/10.3389/fphar.2018.00407/full.
[25] 徐化成, 唐季林. 油松种子发芽的生态学及其与种源的关系[J]. 林业科学, 1989, 25(6):492−501. Xu H C, Tang J L. Seed germination ecology of Pinus tabulaeformis in relation to their provenances[J]. Scientia Silvae Sinicae, 1989, 25(6): 492−501.
[26] 李明, 王树香, 高宝嘉. 油松天然次生林居群遗传多样性及与产地地理气候因子的关联分析[J]. 生态学报, 2013, 33(12):3602−3610. doi: 10.5846/stxb201211271680 Li M, Wang S X, Gao B J, et al. Analysis of genetic diversity of Chinese pine (Pinus tabuliformis) natural secondary forest population and correlation with their habitat ecological factors[J]. Acta Ecologica Sinica, 2013, 33(12): 3602−3610. doi: 10.5846/stxb201211271680
[27] 隋宏大. 树高测量综合技术比较研究[D]. 北京: 北京林业大学, 2009. Sui H D. Comparative study on comprehensive technologies of tree height measurement[D]. Beijing: Beijing Forestry University, 2009.
[28] 袁虎威, 梁胜发, 符学军, 等. 山西油松第二代种子园亲本选择与配置设计[J]. 北京林业大学学报, 2016, 38(3):47−54. Yuan H W, Liang S F, Fu X J, et al. Parental selection and deployment design in the second-generation seed orchard of Chinese pine in Shanxi Province
[J]. Journal of Beijing Forestry University, 2016, 38(3): 47−54. [29] Latta R G, Linhart Y B, Fleck D, et al. Direct and indirect estimates of seed versus pollen movement within a population of ponderosa pine[J]. Evolution, 1998, 52(1): 61−67. doi: 10.1111/j.1558-5646.1998.tb05138.x
[30] Chen K M, Abbott R J, Milne R I, et al. Phylogeography of Pinus tabulaeformis Carr. (Pinaceae), a dominant species of coniferous forest in northern China[J]. Molecular Ecology, 2008, 17(19): 4276−4288. doi: 10.1111/j.1365-294X.2008.03911.x
[31] Xia H H, Wang B, Pan J, et al. Combining mitochondrial and nuclear genome analyses to disst the effects of colonization, environment, and geography on population structure in Pinus tabuliformis[J]. Evolutionary Applications, 2018, 10(11): 1931−1945.
-
期刊类型引用(11)
1. 孙嘉雯,施秀珍,邵雅琪,冯鼎,王建青,刘宝,邹秉章,王思荣,黄志群. 亚热带天然次生林演替对土壤氮矿化的影响机制. 应用生态学报. 2025(01): 59-66 .  百度学术
百度学术
2. 李永涛,王振猛,杨庆山,张子龙,董爱新,周健,曹琪琪,张军. 黄河三角洲柽柳人工林土壤微生物群落结构及多样性分析. 中南林业科技大学学报. 2024(09): 80-93 . 百度学术
3. 樊容源,叶绍明,吴昊,李梓华,李林欣,余春和. 不同林龄桉树人工林土壤细菌群落结构与功能多样性变化特征. 北京林业大学学报. 2024(12): 41-52 . 本站查看
4. TONG Shan,CAO Guangchao,ZHANG Zhuo,ZHANG Jinhu,YAN Xin. Soil microbial community diversity and distribution characteristics under three vegetation types in the Qilian Mountains, China. Journal of Arid Land. 2023(03): 359-376 . 必应学术
5. 刘泽勋,庄家尧,刘超,郑康,陈玲. 大同铅锌尾矿不同污染程度土壤细菌群落分析及生态功能特征. 环境科学. 2023(07): 4191-4200 . 百度学术
6. 王玥. 不同林分密度对卓尼县落叶松林下植物多样性的影响. 南方农业. 2023(09): 273-275 . 百度学术
7. 王明涛,赵玉红,苗彦军,孙磊,王向涛,包赛很那,王敬龙,雷变霞. 藏北一年生人工草地弃耕不同年限的土壤微生物群落特征研究. 中国草地学报. 2023(07): 100-109 . 百度学术
8. 庞丹波,吴梦瑶,赵娅茹,杨娟,董立国,吴旭东,陈林,李学斌,倪细炉,李静尧,梁咏亮. 贺兰山东坡不同海拔土壤微生物群落特征及其影响因素. 应用生态学报. 2023(07): 1957-1967 . 百度学术
9. 张胜男,高海燕,王志波,闫德仁,王春颖,唐凯,冯福应. 华北落叶松-白桦混交林土壤细菌群落结构与多样性及其影响因素. 东北林业大学学报. 2023(10): 98-105 . 百度学术
10. 高禾苗,任爱天,张绍聪,袁龙义. 西藏高原不同树龄核桃林土壤细菌多样性及其影响因素分析. 北京林业大学学报. 2023(11): 100-109 . 本站查看
11. 刘明慧,柳叶,任悦,高广磊,丁国栋,张英,赵珮杉,刘轩. 科尔沁沙地樟子松人工林土壤真菌共现网络及其与土壤因子的关系. 生态学报. 2023(23): 9912-9924 . 百度学术
其他类型引用(6)

计量
- 文章访问数: 2869
- HTML全文浏览量: 1647
- PDF下载量: 75
- 被引次数: 17
