
An intermittent conventional drying schedule for 30 mm-thick Eucalyptus obliqua lumbers
-
摘要:目的 产自澳大利亚的斜叶桉木材渗透性极低,常规干燥过程中极易开裂、皱缩,难以保证质量。本研究在常规干燥工艺中引入间歇处理,制定常规间歇干燥基准,缩短斜叶桉锯材干燥时间的同时提升其干燥质量。方法 采用100 ℃试验法测定斜叶桉木材干燥缺陷等级,拟定干燥初终期条件。结合Keylwerth研究,用图表法得到初步常规干燥基准,干燥30 mm厚斜叶桉锯材。根据干燥初期锯材表面的皱缩程度,设置间歇处理,修订原基准。依据GB/T 6491—2012《锯材干燥质量》,从含水率梯度、弯曲、开裂程度、皱缩等方面评价锯材干燥质量。结果 本研究中斜叶桉试件平均基本密度为(558 ± 21) kg/m3。综合弦切和径切试件测得的缺陷等级,评定斜叶桉木材干燥缺陷等级为初期开裂4级,内裂5级和截面变形5级。据此拟定30 mm厚斜叶桉锯材常规干燥基准的初始温度为46 ℃,初期干湿球温差为1.5 ℃,后期最高温度为67 ℃。干燥过程中试件平均含水率为37.7%、34.4%和24.4%时,观察到严重皱缩,做了3次间歇处理,处理时长分别为16、8和8 h。试件从61.0%的平均初含水率干燥到10.8%的终含水率,共耗时20 d,平均干燥速率为0.10%/h。试件厚度方向含水率偏差为0.70%,可见干燥缺陷指标中顺弯、翘弯、扭曲、纵裂、内裂均达到一级标准,但试件横弯和皱缩较严重,仅能达到三级标准。结论 100 ℃试验法表明:由皱缩导致的截面变形为斜叶桉木材最为严重的干燥缺陷,且径切板的皱缩程度要大于弦切板。在依据100 ℃试验制定的初步干燥基准中加入间歇处理,能够较为有效地缓解和抑制皱缩。本研究中干燥的30 mm厚斜叶桉锯材仍然存在少量皱缩,未达到GB/T 6491—2012中锯材干燥质量二级指标。但是,干燥时间比传统的大气干燥 + 常规干燥的联合干燥方式减少约90%,且干燥质量有所提高。本研究推荐的常规间歇干燥基准可为斜叶桉锯材的实际应用提供科学依据。Abstract:Objective The Eucalyptus obliqua wood from Australia has extremely low permeability and is extremely prone to crack and shrink in the conventional drying process, and it is difficult to ensure the quality. In this study, the intermittent treatment was introduced into the conventional kiln drying process, and an intermittent conventional drying schedule was established, which shortened the drying time of E. obliqua lumbers and improved the drying quality as well.Method The drying characteristics and possible drying defects were obtained by the 100 ℃-test method proposed by Tarazawa. Combined with Keylwerth’s chart method, the preliminary drying schedule for 30 mm thick E. obliqua lumbers was proposed. Intermittent treatments were applied based on the extent of collapse observed on the sample surface at the early stage of drying. The final drying quality of lumber samples was evaluated according to GB/T 6491−2012 “Drying Quality of Lumbers” from the aspect of moisture content gradient, degree of bending, checking and collapse.Result The average basic density of E. obliqua samples in this study was (558 ± 21) kg/m3. Based on the defect grades from the flatsawn and quartersawn 100 ℃-test samples, the final defect grade of E. obliqua wood was evaluated as “Grade-4” for the initial checking, “Grade-5” for the internal checking and “Grade-5” for the cross-section deformation. The initial temperature of the conventional drying schedule for 30 mm thick E. obliqua lumber was set at 46 ℃ with the wet-bulb depression of 1.5 ℃, and the final maximum temperature was set at 67 ℃. During the drying process, severe collapse was observed when the average moisture content of the specimens was 37.7%, 34.4% and 24.4%, and three batches of intermittent treatments were followed with the treatment time of 16, 8 and 8 h, respectively. The specimens were dried from an average initial moisture content of 61.0% to a final one of 10.8% in a total of 20 d, with an average drying rate of 0.10%/h. The moisture content deviation in the thickness direction of the specimen was 0.70%. Most drying defects such as spring, bow, twisting, surface and internal checks of the specimens met the first-class standard, but cupping and collapse of the specimen were more serious, only meeting Grade-3 standard.Conclusion The 100 ℃-test shows that the cross-section deformation caused by collapse is the most serious drying defect of E. obliqua woods, and the collapse extent of the quartersawn samples is greater than that of the flatsawn samples. Adding intermittent treatments to the preliminary conventional drying schedule based on the 100 ℃-test could effectively alleviate and prevent further collapse development effectively. A small amount of collapse still occurs on the 30 mm-thick E. obliqua specimens dried in this study, therefore not meeting the Grade-2 drying quality in GB/T 6491−2012, which is usually required by most enterprises in China. However, the drying time was reduced approximately 90% compared with that of the traditional combined air and conventional kiln drying method, and the drying quality was improved. The intermittent drying schedule recommended in this study can provide insights into the actual drying of E. obliqua woods in the industry.
-
Keywords:
- Eucalyptus obliqua /
- 100 ℃-test /
- intermittent drying schedule /
- collapse
-
板栗(Castanea mollissima)属于壳斗科(Fagaceae)栗属(Castanea)植物,是我国重要的木本粮食树种之一,其果实营养价值高,除富含淀粉外,还有蛋白质、脂肪、矿物质和维生素等营养物质,深受消费者喜爱,具有较高的经济价值[1]。目前我国板栗种植规模和总产量均居世界第一,但由于板栗喜光,枝条顶端优势强,结果部位外移快且自交不亲和,其结实率低,效益低下[2]。因此,培育具有优质、高产、短枝型等优良性状的板栗新品种具有重要的现实意义。
现阶段分子辅助育种已被广泛应用到植物育种工作中,但板栗新品种的培育仍以杂交育种为主[3],如:新品种‘明丰2号’(C. mollissima ‘Mingfeng 2’)是以‘燕明’(C. mollissima ‘Yanming’)为母本、‘燕山早丰’(C. mollissima ‘Yanshanzaofeng’)为父本杂交培育而来[4];‘南垂5号’(C. mollissima ‘Nanchui5’)则是以母本‘垂枝’(C. mollissima ‘Chuizhi’)与父本‘南沟1号’(C. mollissima ‘Nangou1’)杂交培育得来[5]。在杂交授粉过程中,如何提高成功率,以获取丰富的杂种资源是板栗育种亟待解决的问题。较高活性的花粉和柱头是授粉成功的关键[6],准确掌握花粉与柱头活性检测方法,探明其日变化规律,明确最适杂交授粉时间,可有效提高板栗杂交育种成功率。
目前,植物花粉活力检测方法多样,如2,3,5-三苯基氯化四氮唑(TTC)染色法、碘–碘化钾(I2-KI)法、MTT染色法、离体萌发法、墨水染色法等[7−9]。在柱头可授性检测方面,多数学者采用联苯胺–过氧化氢法[10−11]检测柱头可授性,部分研究采用MTT染色法[12]、直接授粉法等[13]。前期板栗花粉活力研究发现,不同板栗品种采用不同花粉活力测定方法,结果存在较大差异。陈可可[14]采用TTC染色法检测5个板栗品种花粉活力发现,‘泰安薄壳栗’(C. mollissima ‘Taianbaoke’)花粉活力最高可达72.18%;覃腾[15]以广西4个板栗品种为研究对象,采用离体萌发法检测其花粉活力,发现‘封开2号’(C. mollissima ‘Fengkai2’)萌发率最高,但仅为15.76%。在板栗柱头可授性研究方面,王倩[16]采用联苯胺–过氧化氢法检测了不同时期燕山板栗柱头可授性,发现盛花期柱头可授性最强;周晶[17]采用联苯胺–过氧化氢法检测板栗‘燕山红’(C. mollissima ‘Yanshanhong’)柱头可授性发现,柱头可授性高峰值出现于盛花期单日08:00—12:00。以上研究对花粉活力和柱头可授性的测定方法和日变化规律进行了初步探索,但并未将花粉活力、柱头可授性日变化规律与其坐果率、结实率、空苞率等进行综合分析。
为准确把握板栗最适授粉时间,提高板栗杂交育种成功率,为后期的良种培育提供充足的杂种资源,本研究首次以河北省迁西县主栽国审板栗品种‘燕山早丰’(果肉质地细腻、味香甜、熟食品质上等、丰产、质感效应较强)为母本[18−19],父本选择具有多粒、矮化等特异性状的‘王钱大栗’(C. mollissima ‘Wangqiandali’)、‘短枝芽变’(C. mollissima ‘Duanzhiyabian’)、‘三屯红栗’(C. mollissima ‘Santunhongli’),采用联苯胺–过氧化氢法、MTT法检测母本柱头可授性,采用TTC染色法、I2-KI法、醋酸洋红染色法、离体萌发法等检测花粉活力,通过研究盛花期花粉活力、柱头可授性、不同时间授粉对结实情况的影响,筛选‘燕山早丰’柱头可授性和杂交花粉活力的适宜检测方法,探明两者的日变化规律,筛选出最适杂交授粉时间。研究结果将为板栗人工杂交育种的成功提供理论指导,同时对解析板栗生殖生物学特性及繁殖机制具有重要意义。
1. 材料与方法
1.1 试验地概况
试验地位于河北省唐山市迁西县北京林业大学经济林(板栗)育种与栽培试验实践基地(40°18′ N,118°54′ E),属暖温带大陆性季风气候,年平均气温10.1 ℃,最低月平均气温−6.5 ℃,最高月平均气温25.4 ℃。年平均降水量817 mm,其中5—10月降雨量657.6 mm,占全年降水量的88%。年平均相对湿度59%,年平均无霜期176 d,全年日照2 581.5 h,日照充足。该地属于低山丘陵区,土壤质地为沙壤土,pH 6.44,有机质含量2.89 mg/g,土壤肥力为中等水平[20]。
1.2 试验材料
材料为11 年生板栗品种‘燕山早丰’‘王钱大栗’‘短枝芽变’‘三屯红栗’,长势优良,无病虫害,株行距4 m × 3 m,平均树高4.3 m,冠幅4.0 m × 4.5 m,树体为三主枝,开张角度 > 60°,树形为自然开心形。
1.3 研究方法
1.3.1 试验设计
参照周晶[17]研究结果,选择盛花期进行人工授粉。选取长势一致且健壮的板栗试验树60株,包括‘燕山早丰’45株,‘王钱大栗’‘短枝芽变’‘三屯红栗’各5株。40株‘燕山早丰’作为母本进行柱头可授性检测与授粉试验,5株‘燕山早丰’用于花粉活力检测;‘王钱大栗’‘短枝芽变’‘三屯红栗为父本,用于花粉活力日变化规律研究与授粉试验。各杂交授粉组合为 ‘燕山早丰’♀ × ‘王钱大栗’♂、‘燕山早丰’♀ × ‘短枝芽变’♂、‘燕山早丰’♀ × ‘三屯红栗’♂和1个自交组合‘燕山早丰’♀ × ‘燕山早丰’♂。
1.3.2 人工授粉
每授粉组合选择10株母树,每棵母树去雄套袋后,于盛花期单日08:00—18:00每隔2 h进行杂交授粉。采用随机区组设计,以单株为试验小区,共10个重复。各授粉组合每株母树每授粉时段在冠层中部东、南、西、北4个方向外围各均匀选取3个长势相近且健壮的雌花簇,1个授粉时段每母株共选取12个雌花簇,每个授粉组合每时段授粉120个雌花簇,4个组合共选取2 400个雌花簇。授粉时去掉套袋,用毛笔蘸取花粉轻轻点涂雌花柱头上,授粉完成后立即套袋。
1.3.3 花粉活力测定方法筛选
在上午08:00—10:00,采集‘燕山早丰’雄花盛花期的雄花序,每株树均从东、南、西、北4个方向取样,混合后分别采用TTC染色法[21]、醋酸洋红染色法[22]、I2-KI染色法[23]、离体萌发培养法[24]测定花粉活力,并以清水作为对照,筛选板栗花粉活力适宜检测方法。
1.3.4 花粉活力日变化规律研究
在08:00—18:00每隔2 h采集‘燕山早丰’‘王钱大栗’‘短枝芽变’‘三屯红栗’的雄花盛花期的花粉,选择1.3.3筛选出的花粉活力检测方法进行花粉活力日变化规律研究。
1.3.5 柱头可授性检测方法筛选
从40株‘燕山早丰’授粉母株中随机选取15株,在雌花盛花期每株树冠层中部东、南、西、北4个方向各均匀选取3个雌花簇,在上午08:00—10:00采集柱头,分别利用联苯胺–过氧化氢法[25]和MTT染色法[26]测定柱头可授性,并以清水作为对照,筛选板栗柱头可授性适宜检测方法。联苯胺–过氧化氢法判定标准:若柱头周围产生气泡数量多且体积较大,则柱头可授性很强(+++);若柱头周围产生大量小气泡,则柱头可授性较强(++);若柱头周围产生少量气泡,则柱头可授性较弱(+);若柱头无气泡产生,则柱头无可授性(−)。MTT染色法判定标准:若柱头具有可授性,则柱头被染成深蓝色或黑色,颜色由浅变深,柱头可授性由弱变强;若没有可授性,则柱头没有颜色反应。
1.3.6 柱头可授性日变化规律研究
在08:00—18:00每隔2 h采集‘燕山早丰’盛花期的柱头,选择1.3.5筛选出的柱头可授性检测方法进行柱头可授性日变化规律研究。
1.3.7 结实情况统计
授粉20 d后去除纸袋,使球苞自然生长,统计坐果率、落果率,9月上旬果实成熟后,计算结实率、空苞率。坐果率为未凋落雌花簇数与授粉雌花簇数的比值,落果率为凋落雌花簇数与授粉雌花簇数的比值,空苞率为空苞数与授粉雌花簇数的比值,结实率为果实成熟时的结实苞数与授粉雌花簇数的比值[15]。
1.4 数据分析
试验数据采用Excel软件进行整理,SPSS 26.0进行方差分析。
2. 结果与分析
2.1 花粉活力适宜检测方法筛选
花粉在清水处理的情况下,颜色均为淡黄色,接近半透明,分为形状规则的卵圆形与形状不规则两种形态(图1a)。0.5%和2%的TTC染色法的染色效果均不明显,延长染色时间后花粉依旧呈淡黄色(图1b,1c),故此方法不适合板栗花粉活力检测。醋酸洋红染色法效果明显,有活力的花粉呈鲜红色,形状呈规则的卵圆形,而无活力花粉未着色,体积小且形状不规则(图1d);该方法操作简单,染色时间短,效果好,有活力花粉与无活力花粉较易辨别,可作为板栗花粉活力检测方法。I2-KI染色法染色效果明显,有活力花粉颜色呈深棕色,无活力花粉不着色,但两者之间存在色浅、黄褐色的花粉,花粉活力较难判别,故此方法也不适合板栗花粉活力检测(图1e)。离体萌发培养法可以直接观察到花粉的萌发情况(图1f),可判断花粉是否具有活力,但花粉离体培养耗时较长,且培养条件较为严苛,易染菌,需要在无菌且温度恒定的环境下培养,花粉整体萌发率较低。
![]() 图 1 不同方法测定‘燕山早丰’花粉活力效果a. 清水对照;b. 0.5%TTC染色法;c. 2%TTC染色法;d. 醋酸洋红染色法;e. I2-KI染色法;f. 离体萌发法Figure 1. Effects of different methods on measuring pollen viability of ‘Yanshanzaofeng’
图 1 不同方法测定‘燕山早丰’花粉活力效果a. 清水对照;b. 0.5%TTC染色法;c. 2%TTC染色法;d. 醋酸洋红染色法;e. I2-KI染色法;f. 离体萌发法Figure 1. Effects of different methods on measuring pollen viability of ‘Yanshanzaofeng’综合反应条件、反应时间、判断标准等方面,发现醋酸洋红染色法更适合进行板栗盛花期单日花粉活力规律研究。
2.2 不同板栗品种盛花期单日花粉活力变化规律
采用醋酸洋红染色法测定不同板栗品种单日内花粉活力变化规律,结果如表1所示。各品种盛花期单日内花粉活力均保持在75%以上,但不同品种板栗花粉在单日内不同时间活力存在差异。‘短枝芽变’和‘三屯红栗’单日内花粉活力波动较小,最高值均出现在10:00—12:00。‘燕山早丰’‘王钱大栗’单日内花粉活力波动相对较大,但花粉活力最高值也出现在10:00—12:00,最低值出现时段存在差异。其中,‘燕山早丰’10:00—12:00花粉活力显著高于16:00—18:00,‘王钱大栗’10:00—12:00花粉活力显著高于其余时间花粉活力。综上,以这4个品种为父本时,应该选择上午10:00—12:00采集花粉,此时花粉活力最高。
表 1 不同板栗品种盛花期花粉活力变化Table 1. Changes in pollen viability of different chestnut cultivars at full flowering stage% 时段 ‘燕山早丰’ ‘短枝芽变’ ‘王钱大栗’ ‘三屯红栗’ 08:00—10:00 84.78ab 90.47a 83.71b 88.58a 10:00—12:00 90.71a 91.32a 87.26a 88.97a 12:00—14:00 80.65ab 88.93a 75.34c 82.23a 14:00—16:00 81.14ab 90.63a 79.49c 83.76a 16:00—18:00 78.63b 86.89a 78.26c 80.53a 注:不同小写字母表示同一品种单日内不同时间花粉活力存在显著差异(P < 0.05)。下同。 2.3 柱头可授性适宜检测方法筛选
采用联苯胺–过氧化氢法检测‘燕山早丰’柱头可授性,由于板栗柱头形状结构限制,染色效果不佳,仅有柱头尖端可观察到轻微蓝色反应;但其表面气泡反应明显,区分度较高。柱头表面只产生少量气泡,且气泡体积较小,柱头可授性较弱,标记为+,记1分(图2a-1);柱头表面产生较多小气泡,柱头可授性较强,标记为++,记2分(图2a-2);柱头表面迅速产生大量气泡,且均匀附着整个柱头,柱头可授性最强,标记为+++,记3分(图2a-3)。
![]() 图 2 联苯胺–过氧化氢法(a)和MTT染色法(b)检测‘燕山早丰’柱头可授性Figure 2. Detecting stigma receptivity of ‘Yanshanzaofeng’ according to benzidine hydrogen peroxide method (a) and MTT method (b)
图 2 联苯胺–过氧化氢法(a)和MTT染色法(b)检测‘燕山早丰’柱头可授性Figure 2. Detecting stigma receptivity of ‘Yanshanzaofeng’ according to benzidine hydrogen peroxide method (a) and MTT method (b)采用MTT染色法检测柱头可授性,短时间内柱头表面无明显颜色变化,避光染色1 h后,染色情况较为明显,有活力的柱头顶部变为黑色,且随着柱头可授性增强,染色长度逐渐增大。具体而言,仅有柱头尖端部分染色,染色面积微小,呈点状,此柱头可授性较弱(图2b-1);柱头尖端面积染色面积增大,其柱头底部也部分染色,即柱头可授性较强(图2b-2);柱头尖端染色部分延伸至柱头中部,且花柱上端也有部分染色,此柱头可授性极强(图2b-3)。
综上,联苯胺–过氧化氢法和MTT染色法都可以判断板栗柱头可授性强弱。综合考虑染色时长、可操作性等因素,在实际生产中,采用联苯胺–过氧化氢法检测板栗柱头可授性更适宜。因而,选择联苯胺–过氧化氢法进行板栗盛花期单日柱头可授性变化规律研究。
2.4 ‘燕山早丰’盛花期单日柱头可授性变化规律
表2显示:‘燕山早丰’盛花期单日内不同时段柱头均具有可授性,整体呈现出“上升—下降—上升—下降”的变化趋势。上午08:00—10:00柱头就已经具有较强可授性,10:00—12:00可授性达到最高,柱头表面具有大量气泡;随后急剧下降,12:00—14:00可授性要明显低于其他时段,之后可授性又重新升高,于14:00—16:00达到日变化第2个高峰,16:00—18:00可授性降低(图3)。
表 2 联苯胺–过氧化氢法检测‘燕山早丰’盛花期单日不同时间柱头可授性Table 2. Detecting single day stigma receptivity during full flowering period of ‘Yanshanzaofeng’ by benzidine-hydrogen peroxide method时段 柱头反应 柱头可授性 08:00—10:00 浸泡在溶液中3 ~ 5 min后,柱头表面出现大量气泡,且体积较大 +++ 10:00—12:00 浸泡溶液3 min左右,柱头表面出现大量气泡,气泡密集且体积较大 +++ 12:00—14:00 浸泡溶液5 min后,柱头表面出现少量气泡,体积较小 + 14:00—16:00 浸泡溶液5 min后,柱头表面出现气泡,气泡密集 +++ 16:00—18:00 浸泡溶液5 min后,柱头表面出现大体积气泡,但数量较少 ++ 注:+++. 柱头可授性极强;++. 柱头可授性较强;+. 柱头具可授性。 ![]() 图 3 联苯胺–过氧化氢法检测‘燕山早丰’盛花期单日柱头可授性a. 08:00—10:00;b. 10:00—12:00;c. 12:00—14:00;d. 14:00—16:00;e. 16:00—18:00Figure 3. Detecting single day stigma receptivity during blooming period of ‘Yanshanzaofeng’ by benzidine-hydrogen peroxide method
图 3 联苯胺–过氧化氢法检测‘燕山早丰’盛花期单日柱头可授性a. 08:00—10:00;b. 10:00—12:00;c. 12:00—14:00;d. 14:00—16:00;e. 16:00—18:00Figure 3. Detecting single day stigma receptivity during blooming period of ‘Yanshanzaofeng’ by benzidine-hydrogen peroxide method2.5 盛花期单日不同授粉时间对板栗结实情况的影响
各杂交组合中,不同时间授粉落果率与坐果率均无显著差异,但结实率与空苞率在不同授粉时间存在显著差异(图4)。 ‘燕山早丰’ × ‘王钱大栗’杂交组合在盛花期08:00—10:00与10:00—12:00时段授粉,其结实率分别高达79.30%与80.36%,显著高于12:00—14:00与16:00—18:00两个授粉时段(图4);且在10:00—12:00时段授粉,3粒坚果球苞结实率相较于其他授粉时段最高,达55.10%(表3);空苞率在10:00—12:00授粉时段最低,仅为14.74%,而16:00—18:00授粉时段的空苞率高达37.88%,二者差异达到显著水平。
![]() 图 4 各授粉组合盛花期单日不同时段授粉结果a.‘燕山早丰’♀ × ‘王钱大栗’♂;b.‘燕山早丰’♀ × ‘短枝芽变’♂;c.‘燕山早丰’♀ × ‘三屯红栗’♂;d.‘燕山早丰’♀ × ‘燕山早丰’♂。不同小写字母表示同一授粉组合单日内不同时间授粉后结实情况存在显著差异(P < 0.05)。Figure 4. Pollination results of different combinations at varied times in a single day during blooming period表 3 各授粉组合盛花期单日不同时间授粉结实情况Table 3. Pollination and seed setting at different times of a single day during blooming period of each pollination combination
图 4 各授粉组合盛花期单日不同时段授粉结果a.‘燕山早丰’♀ × ‘王钱大栗’♂;b.‘燕山早丰’♀ × ‘短枝芽变’♂;c.‘燕山早丰’♀ × ‘三屯红栗’♂;d.‘燕山早丰’♀ × ‘燕山早丰’♂。不同小写字母表示同一授粉组合单日内不同时间授粉后结实情况存在显著差异(P < 0.05)。Figure 4. Pollination results of different combinations at varied times in a single day during blooming period表 3 各授粉组合盛花期单日不同时间授粉结实情况Table 3. Pollination and seed setting at different times of a single day during blooming period of each pollination combination授粉组合 授粉时段 结实率(1粒)/% 结实率(2粒)/% 结实率(3粒)/% 总结实率/% ‘燕山早丰’♀ × ‘王钱大栗’♂ 08:00—10:00 10.76a 14.71a 53.83a 79.30a 10:00—12:00 9.64a 15.62a 55.10a 80.36a 12:00—14:00 12.24a 17.78a 30.58b 60.60bc 14:00—16:00 5.21a 20.96a 44.24ab 70.42ab 16:00—18:00 12.35a 12.92a 29.86b 55.12c ‘燕山早丰’♀ × ‘短枝芽变’♂ 08:00—10:00 10.59a 8.07ab 40.69a 59.35a 10:00—12:00 4.98a 7.40ab 43.72a 56.10a 12:00—14:00 7.13a 3.25b 26.20ab 36.59b 14:00—16:00 5.67a 12.61a 34.56a 52.85a 16:00—18:00 7.28a 8.01ab 23.62ab 38.90ab ‘燕山早丰’♀ × ‘三屯红栗’♂ 08:00—10:00 3.25b 19.76a 53.41a 76.42a 10:00—12:00 10.60a 17.28ab 50.98a 78.86a 12:00—14:00 14.65a 17.31ab 29.02b 60.98ab 14:00—16:00 13.82a 17.32ab 39.06ab 70.20ab 16:00—18:00 11.38a 16.50b 29.02b 56.91b ‘燕山早丰’♀ × ‘燕山早丰’♂ 08:00—10:00 23.80a 4.70a 2.82a 31.32a 10:00—12:00 21.12a 4.17ab 2.50ab 27.79ab 12:00—14:00 17.63a 3.25ab 1.95ab 22.83ab 14:00—16:00 20.07a 3.96ab 2.38ab 26.40ab 16:00—18:00 16.84a 2.73b 1.64b 21.21b 注:不同小写字母表示同一授粉组合单日内不同时段授粉后结实情况存在显著差异(P < 0.05)。 在‘燕山早丰’ × ‘短枝芽变’杂交组合中,结实率在08:00—10:00授粉时段最高(59.35%),10:00—12:00时段的结实率与08:00—10:00时段相比略有下降(56.10%),并显著高于12:00—14:00授粉时段(图4);3粒坚果球苞结实率以10:00—12:00时段最高(43.72%)(表3);空苞率则以10:00—12:00授粉时段最低(27.64%),12:00—14:00与16:00—18:00的空苞率则高达46.34%与44.59%,均显著高于10:00—12:00(图4)。
在‘燕山早丰’ × ‘三屯红栗’杂交组合中,08:00—10:00、10:00—12:00授粉时段的结实率较高,达76.42%、78.86%,16:00—18:00结实率最低(56.91%),与上午差异达到显著水平(图4);3粒坚果球苞结实率以08:00—10:00时段最高(53.41%),10:00—12:00时段略有下降(50.98%)(表3);空苞率则在10:00—12:00授粉时段最低(8.94%),显著低于12:00—14:00和16:00—18:00授粉时段。
在‘燕山早丰’ × ‘燕山早丰’自交组合中,结实率均在30%左右(图4)。在08:00—10:00授粉时段,结实率最高(31.32%),空苞率最低(30.38%);在16:00—18:00授粉时段结实率最低(21.21%)(图4)。结实的球苞大多只含有1粒坚果,2粒坚果球苞与3粒坚果球苞结实率均以08:00—10:00最高,16:00—18:00最低;空苞率则以12:00—14:00最高,达53.54%(表3)。相比其他杂交组合,自交组合中的结实率较低,落果率与空苞率较高;但是结实率最高值出现在上午授粉时段,这与其他杂交组合相同(图4)。
综合不同板栗品种授粉组合在不同时时段授粉后的结实指标可以得出,结实率和3粒坚果球苞结实率最高值均出现在上午。因此,为提高板栗杂交授粉结实率,应选择上午进行板栗杂交授粉。
3. 讨 论
3.1 花粉活力检测
前人系统研究花粉活力发现,不同植物有不同的适宜检测方法。如:花生(Arachis hypogaea)花粉活力的最适检测方法为离体萌发法与红墨水染色法[7],薄壳山核桃(Carya illinoinensis)的花粉采用MTT染色法着色效果最佳[8],多花黄精(Polygonatum cyrtonema)的花粉活力最适检测方法为TTC染色法[27]。TTC试剂是依靠呼吸作用产生的NADH2或NADPH2对花粉进行染色。本试验中TTC试剂无法使板栗花粉着色,这可能是由于板栗自身呼吸作用产生的脱氢酶较少,无法达到理想的染色状态。此外,利用I2-KI对花粉进行染色时,染色效果会因直链淀粉与支链淀粉的比例不同而产生差异[28]。当花粉直链淀粉含量高时,会被试剂染为蓝色,如芝麻(Sesamum indicum)[23]、油茶(Camellia oleifera)[29];反之则为紫红色,如花生[7]、百合(Lilium lancifolium)[30]等。醋酸洋红染液则是根据花粉内物质积累量对花粉进行染色,正常花粉物质积累较多,对醋酸洋红的吸附作用与亲和力强,易被染为深红色,反正,呈淡红色或无色,如油菜(Brassica campestris)[31]。本研究中,醋酸洋红染色法、I2-KI染色法均可将‘燕山早丰’花粉成功染色,但是醋酸洋红染色法操作简单,染色快速,有活力花粉与无活力花粉易辨别,结果准确;而I2-KI染色法中存在浅色、黄褐色的介于有活力和无活力之间的花粉,活力界定具有主观性,花粉活力较难准确判别。因而,建议生产上使用醋酸洋红染色法检测‘燕山早丰’花粉活力。
3.2 柱头可授性检测
利用联苯胺–过氧化氢法检测柱头可授性,主要是依靠柱头表面有活力的过氧化氢酶,试剂中的过氧化氢在酶的作用下产生强氧化性的原子氧,将联苯胺氧化变色,并产生气泡。故当柱头表面气泡体积较大,数量较多时,柱头具有较强可授性,反之则柱头可授性较弱。此方法已广泛用于植物柱头可授性检验,如川贝母(Fritillaria cirrhosa)[32]、兜兰(Paphiopedilum)[33]等。MTT染色法检测柱头可授性主要依据柱头表面活性酶与试剂反应变色原理,可授性较强的柱头被染为深色,且染色面积较大,反之柱头颜色较浅[34−35]。本试验采用联苯胺–过氧化氢法和MTT染色法检验‘燕山早丰’的柱头可授性,其反应结果与其他植物基本相同,但MTT染色法检验板栗柱头可授性耗时较长,可能导致错过最适授粉时间。因此,在生产上可选取联苯胺–过氧化氢法快速检测柱头可授性,为杂交授粉提供指导。
3.3 花粉活力与柱头可授性日变化规律
花粉活力是决定授粉成功的重要因素,柱头是植物承接花粉的部位,二者对于准确把握植物授粉时间有重要意义,共同决定植物育种成功率[36]。不同植物的花粉活力与柱头可授性变化规律不同[23,37],因此,植物育种授粉前,应准确掌握花粉活力和柱头的变化规律,由此得到最适授粉时间,提高育种成功率。
本研究发现,上午08:00—12:00柱头可授性与花粉活力均较强,而在正午12:00—14:00柱头可授性较弱,且此时花粉活力显著降低,这可能与空气温度、湿度变化有关。板栗柱头为针状形,在雌花成熟过程中可观察到柱头尖端有蛋白质、脂类、多糖等分泌物溢出[38],中午温度升高,空气湿度降低,导致柱头分泌物内酶活性降低,柱头可授性降低。这与温度对甜樱桃(Prunus avium)柱头可授性的影响结果[39]相一致。此外,正午高温导致板栗花粉活力降低,这与辣椒(Capsicum annuum)的研究结果[40]相似。正午过后,气温降低,日照强度减弱,柱头内酶活性增强,柱头可授性与花粉活力回升;傍晚气温继续下降导致柱头可授性与花粉活力又重新降低。本研究得出的柱头可授性变化规律与周晶[17]对板栗柱头可授性的研究结果基本一致,且本研究中板栗花粉活力检测方法和变化规律方面的内容也弥补了板栗相关研究的缺失。
3.4 杂交授粉时间与结实情况
授粉时间显著影响授粉受精完成情况,如周建辉等[41]对厚皮甜瓜(Cucumis melo)进行不同时间杂交授粉,发现06:00—12:00授粉种子发芽率较高。对西瓜(Citrullus lanatus)[42]、西葫芦(Cucurbita pepo)[43]的研究也证实上午授粉比下午效果好,主要表现为坐果率较高,结籽数量多。此外,对玉米(Zea mays)进行不同时间授粉处理,发现不同自交系均在09:00—11:00和16:00—18:00授粉果穗的结实性状较好[44]。这与本研究得到的板栗08:00—12:00与14:00—16:00杂交授粉结实情况较好的结果相似,即最适授粉时间均避开了正午时段。除可以根据授粉结实情况筛选植物授粉时间外,也可根据柱头可授性和花粉活力筛选植物的适宜授粉时间。对青杨派树种进行柱头可授性检测,发现最佳授粉期为花序顶部露出芽鳞84 ~ 96 h[45]。本研究中,上午10:00—12:00进行授粉结实效果优良,这与前期柱头可授性与花粉活力变化规律的结果相符合,进一步验证了此时段为板栗最适杂交授粉时间。
通过将3个杂交授粉组合与自交授粉结实情况相对比,可以看出‘燕山早丰’存在明显的自交不亲和现象,其自交落果率与空苞率均较高,而结实率较低。不同杂交授粉组合间结实情况也存在差异,以‘王钱大栗’‘三屯红栗’为父本的授粉组合的结实率和3粒坚果球苞结实率与以‘短枝芽变’为父本的授粉组合存在较大差距,即存在明显的花粉直感效应,这与前人的研究结果[19]一致。后续在进行板栗杂交育种和种质优化时,宜选取性状优良的品种为父本,以丰富子代板栗优良性状。
4. 结 论
本研究首次以国审板栗品种‘燕山早丰’为试验对象,筛选其柱头可授性与花粉活力的高效便捷检测方法,并在此基础上以其为母本进行了多组合的分时段授粉,从而筛选出板栗适宜的杂交授粉时段。研究得到以下主要结论。
(1)板栗‘燕山早丰’柱头可授性最适检测方法为联苯胺–过氧化氢法,花粉活力最适检测方法为醋酸洋红染色法。
(2)盛花期单日内柱头可授性呈“上升—下降—上升—下降”趋势,且以10:00—12:00时间段柱头可授性最强,此时段不同父本板栗品种的花粉活力也最高。
(3)不同杂交组合中,总结实率与3粒坚果球苞结实率最高值均出现在08:00—12:00。
综合上述结果,实际生产中建议在上午10:00—12:00对‘燕山早丰’杂交授粉。研究结果不仅为优化板栗人工杂交授粉技术,提高板栗杂交授粉成功率,以获得充足的杂种资源提供了理论指导,同时有助于丰富板栗生殖生物学内容,揭示其繁殖机制。本研究设置的授粉试验时间梯度为2 h,在后续研究中可将时间梯度细化,以期筛选出更加精确的最适授粉时间,从而获得更理想的结实效果。
-
![]()
图 1 30 mm厚斜叶桉干燥曲线和干燥过程中温湿度变化
Figure 1. Drying curves for the 30 mm-thick E. obliqua lumbers and the corresponding temperature and relative humiditychange during drying process
![]()
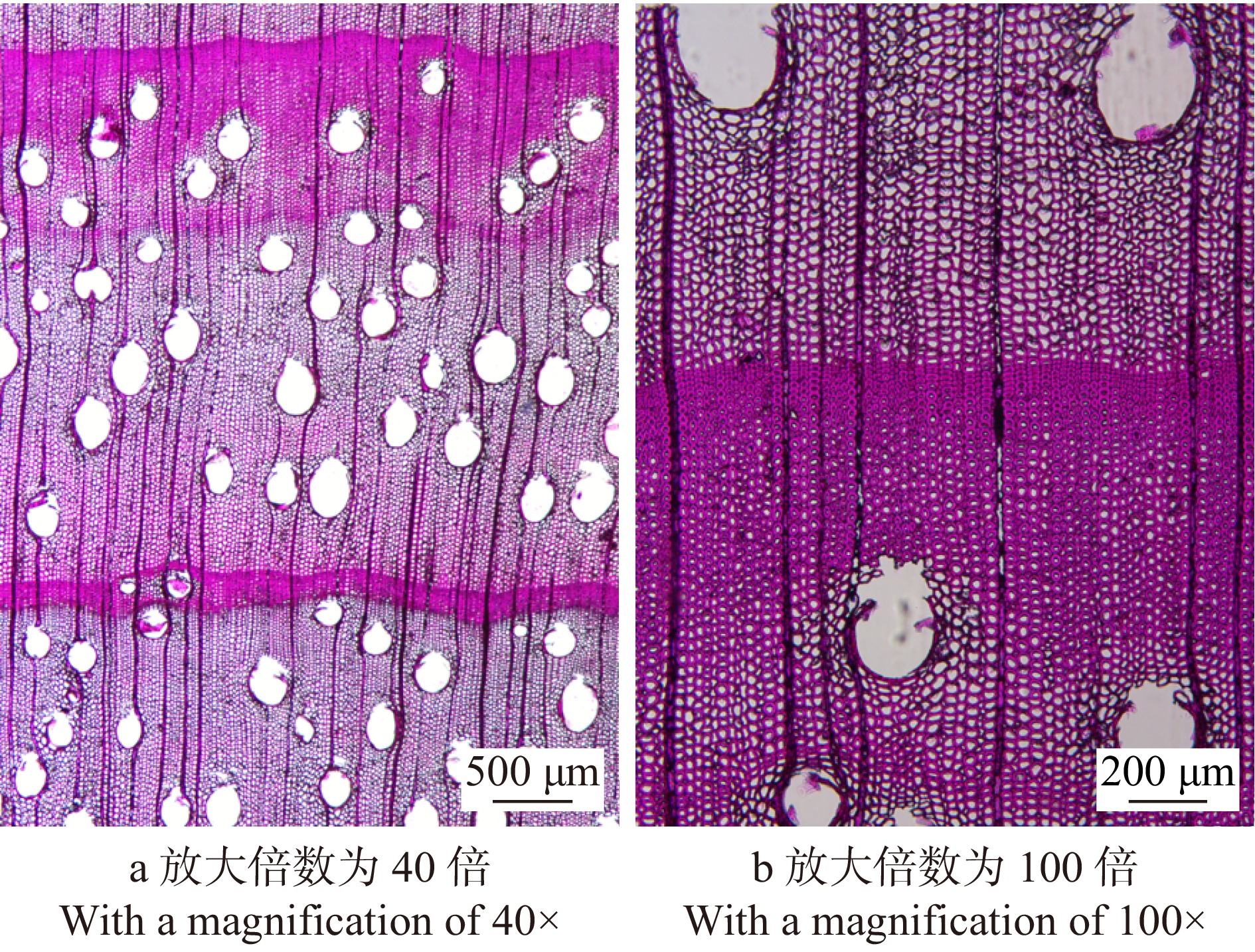
图 2 呈现出严重的皱缩(A)和内裂(B)的100 ℃试验径切板试件
Figure 2. Radial 100 ℃-test sample showing serious collapse (A) and internal checks (B)
![]()
图 4 无严重皱缩、内裂的100 ℃试验试件
Figure 4. 100 ℃-test sample with no serious collapse and internal checks
表 1 斜叶桉100 ℃实验干燥缺陷等级
Table 1 Grades of drying defects in the 100 ℃-test for E. obliqua samples
试件编号
Sample No.初期开裂情况
Initial checking condition内裂情况
Internal checking condition初期开裂等级
Initial checking
grade内裂等级
Internal checking
grade截面变形等级
Cross-section deformation
grade截面变形
Cross-section deformation/
mmT1 短细表裂7条;短端表裂3条
7 short and fine surface checks and 3 short end-surface checks宽裂4条;细裂16条
4 wide checks and 16 fine checks2 5 4 3.31 T2 短细表裂3条;短端表裂1条;
贯通裂1条
3 short and fine surface checks; 1 short end-surface check; 1end splitting宽裂5条;细裂7条
5 wide checks and 7 fine checks4 4 4 2.64 T3 短细表裂16条;长端表裂2条,短端表裂3条
16 short and fine surface checks; 2 long and 3 short end-surface checks宽裂7条;细裂12条
7 wide checks and 12 fine checks4 5 4 3.39 T4 短端表裂1条
1 short and fine surface check宽裂3条;细裂4条
3 wide checks and 4 fine checks1 3 5 3.84 T5 长细表裂4条,短细表裂28条;长端表裂1条,短端表裂4条;贯通裂1条
4 long and fine surface checks, 28 short and fine surface checks; 1 long and 4 short end-surface checks; 1 end splitting宽裂6条;细裂15条
6 wide checks and 15 fine checks4 5 4 2.50 T6 短细表裂16条;短端表裂5条;
贯通裂1条
16 short and fine surface checks;
5 short end-surface checks; 1 end splitting宽裂7条;细裂17条
7 wide checks and 17 fine checks4 5 4 2.65 T7 短细表裂5条;短端表裂2条
5 short and fine surface checks;
2 short end-surface checks宽裂8条;细裂13条
8 wide checks and 13 fine checks2 5 4 3.41 T8 短细表裂5条;短端表裂3条
5 short and fine surface checks;
3 short end-surface checks宽裂9条;细裂11条
9 wide checks and 11 fine checks2 5 4 2.86 T9 短细表裂3条;短端表裂5条
3 short and fine surface checks;
5 short end-surface checks宽裂7条;细裂15条
7 wide checks and 15 fine checks2 5 4 2.86 T10 短细表裂1条;短端表裂4条
1 short and fine surface checks;
4 short end-surface checks宽裂4条;细裂13条
4 wide checks and 13 fine checks2 4 4 2.14 R1 无裂纹
No check宽裂2条;细裂15条
2 wide checks and 15 fine checks1 4 5 4.87 R2 无裂纹
No check宽裂3条;细裂11条
3 wide checks and 11 fine checks1 4 5 5.04 R3 无裂纹
No check宽裂4条;细裂10条
4 wide checks and 10 fine checks1 4 5 5.46 R4 无裂纹
No check宽裂1条;细裂15条
1 wide check and 15 fine checks1 4 5 4.87 R5 长侧裂1条;长端侧裂1条;短端表裂3条
1 long lateral check;1 long end-lateral check; 3 short end-surface checks宽裂1条;细裂12条
1 wide check and 12 fine checks1 4 5 3.90 R6 短细侧裂5条
5 short and fine lateral check宽裂3条;细裂9条
3 wide check and 9 fine checks1 4 5 4.54 R7 短细侧裂1条
1 short and fine lateral check宽裂1条;细裂12条
1 wide check and 12 fine checks1 4 5 4.60 R8 短细侧裂1条
1 short and fine lateral check细裂12条
12 fine checks1 4 5 5.00 注:表中T1 ~ T10为弦切试件,其中T4偏径切;R1 ~ R8为径切试件,其中R5、R6略偏弦切。Notes: T1–T10 in the above table are flatsawn samples, of which T4 is partially quartersawn; R1–R8 are quartersawn samples, of which R5 and R6 are partially flatsawn.  下载: 导出CSV
下载: 导出CSV
表 2 100 ℃试验缺陷程度与干燥条件的关系
Table 2 Recommended initial and final drying conditions based on the 100 ℃-test
初步拟定干燥条件
Initially assumed
drying condition100 ℃试验等级100 ℃-test grade 最终干燥条件
Final drying condition初期开裂(4级)
Initial checking (grade 4)内裂(5级)
Internal checking (grade 5)截面变形(5级)
Cross-section deformation (grade 5)初始温度
Initial temperature/℃50 45 45 46 初期干湿球温差
Initial wet-bulb depression/℃2.0 2.5 2.5 1.5 后期最高温度
Final highest temperature/℃80 70 70 67
下载: 导出CSV
表 3 30 mm 厚斜叶桉锯材试用干燥基准
Table 3 Preliminary drying schedule for 30 mm-thick E. obliqua lumbers
含水率调节
Moisture content (MC) adjustment干球温度
Dry-bulb temperature/℃干湿球温度差
Wet-bulb depression/℃平衡含水率
Equilibrium MC/%相对湿度
Relative humidity/%干燥至50%
Dry to 50%46 1.5 20.0 92 干燥至40%
Dry to 40%50 1.5 20.0 92 从40%干燥至35%
From 40% to 35% by drying50 2.5 18.0 87 从35%干燥至30%
From 35% to 30% by drying52 4.0 15.0 80 从30%干燥至25%
From 30% to 25% by drying55 7.0 10.5 68 从25%干燥至20%
From 25% to 20% by drying58 11.0 8.0 54 从20%干燥至15%
From 20% to 15% by drying63 17.0 5.5 40 干燥至15%以下
Below 15% by drying67 22.0 4.0 30
下载: 导出CSV
表 4 30 mm 厚斜叶桉锯材干燥质量
Table 4 Drying quality for 30 mm-thick E. obliqua lumbers
项目
Item终含水率
Final moisture content/%厚度上含水率偏差
Moisture content deviation on thickness/%翘曲度
Warp degree/%扭曲度
Twist degree/%纵裂度
Longitudinal fissure degree/%内裂个数
Number of internal check皱缩深度
Collapse depth/mm顺弯 Bow 横弯 Cupping 翘弯 Spring 平均值
Mean value10.8 0.7 0 1.56 0.32 0 0 0 1.34 等级 Grade 2 1 1 3 1 1 1 1 3
下载: 导出CSV
表 5 30 mm 厚斜叶桉锯材推荐干燥基准
Table 5 Recommended drying schedule for 30 mm-thick E. obliqua lumbers
处理
Treatment干球温度
Dry-bulb
temperature/℃干湿球温度差
Web-bulb
depression/℃平衡含水率
Equilibrium MC/%相对湿度
Relative humidity/%处理时间
Treatment time/hMC干燥至40%
Dry to 40% of MC46 3 16.0 85 间歇处理1
Intermittent treatment 125 0 30.0 100 12 MC从40%干燥至35%
From 40% to 35% of MC by drying50 3 16.0 83 间歇处理2
Intermittent treatment 225 0 30.0 100 12 MC从35%干燥至30%
From 35% to 30% of MC by drying50 4 14.0 79 MC从30%干燥至25%
From 30% to 25% of MC by drying50 5 12.5 76 间歇处理3
Intermittent treatment 325 2 17.2 85 12 MC从25%干燥至20%
From 25% to 20% of MC52 6 11.5 71 MC从20%到15%
From 20% to 15% of MC by drying54 6 11.5 72 中间处理
Intermediate treatment54 3 16.0 84 12 MC干燥至15%以下
Below 15% of MC by drying60 8 10.0 65 终了调湿处理
Final humidifying treatment65 3 15.5 86 8
下载: 导出CSV
-
[1] Wallis N K. Australian timber handbook[M]. Sydney: Angus and Robertson, 1963.
[2] Redman A L, Bailleres H, Turner I, et al. Mass transfer properties (permeability and mass diffusivity) of four Australian hardwood species[J]. BioResources, 2012, 7(3): 3410−3424.
[3] Redman A L, Bailleres H, Turner I, et al. Characterisation of wood-water relationships and transverse anatomy and their relationship to drying degrade[J]. Wood Science and Technology, 2016, 50(4): 739−757. doi: 10.1007/s00226-016-0818-0
[4] Redman A L, Bailleres H, Perré P, et al. A relevant and robust vacuum-drying model applied to hardwoods[J]. Wood Science and Technology, 2017, 51(4): 701−719. doi: 10.1007/s00226-017-0908-7
[5] 周桥芳, 胡传双, 涂登云, 等. 大花序桉干燥特性与干燥工艺研究[J]. 林业机械与木工设备, 2018, 46(1): 5. doi: 10.3969/j.issn.2095-2953.2018.01.009 Zhou Q F, Hu C S, Tu D Y, et al. Drying characteristics and drying process of Eucalyptus cloeziana wood[J]. Forestry Machinery & Woodworking Equipment, 2018, 46(1): 5. doi: 10.3969/j.issn.2095-2953.2018.01.009
[6] 刘媛, 卢翠香, 黄腾华, 等. 桉树无性系大径材干燥特性分析[J]. 南方农业学报, 2017, 48(5): 5. doi: 10.3969/j.issn.2095-1191.2017.05.023 Liu Y, Lu C X, Huang T H, et al. Drying characteristics of Eucalyptus clones with large-diameter[J]. Journal of Southern Agriculture, 2017, 48(5): 5. doi: 10.3969/j.issn.2095-1191.2017.05.023
[7] 刘元, 吴义强, 乔建政, 等. 桉树人工林木材的干燥特性及干燥基准研究[J]. 中南林业科技大学学报, 2002, 22(4): 44−49. doi: 10.3969/j.issn.1673-923X.2002.04.015 Liu Y, Wu Y Q, Qiao J Z, et al. A study of drying characteristics and drying schedules of Eucalyptus plantation wood[J]. Journal of Central South Forestry University, 2002, 22(4): 44−49. doi: 10.3969/j.issn.1673-923X.2002.04.015
[8] 赵荣军, 江泽慧, 费本华, 等. 澳大利亚桉树木材加工利用研究现状[J]. 世界林业研究, 2003, 16(3): 58−61. doi: 10.3969/j.issn.1001-4241.2003.03.012 Zhao R J, Jiang Z H, Fei B H, et al. Present studies on processing and utilization of eucalypt wood in Australia[J]. World Forestry Research, 2003, 16(3): 58−61. doi: 10.3969/j.issn.1001-4241.2003.03.012
[9] 戴于龙, 黄月瑞, 刘渝, 等. 推广预测木材干燥基准的百度试验法的可行性探讨[J]. 北京林学院学报, 1985, 7(1): 19−29. Dai Y L, Huang Y R, Liu Y, et al. Feasibility of a new method for determination of kiln-drying schedule of wood[J]. Journal of Beijing Forestry College, 1985, 7(1): 19−29.
[10] 寺泽真. 木材干燥的简单决定法[J]. 木材工业(日本), 1965(5): 216−221. Terazawa S. A simple method for determination of kiln-drying schedule of wood[J]. Wood Industry (Japan), 1965(5): 216−221.
[11] Yang L, Liu H, Cai Y, et al. A novel method of studying the collapsed cell of eucalyptus wood using X-ray CT scanning[J]. Drying Technology, 2019, 37(12): 1597−1604. doi: 10.1080/07373937.2018.1519572
[12] Yang L, Liu H. A review of Eucalyptus wood collapse and its control during drying[J]. BioResources, 2018, 13(1): 2171−2181.
[13] Phonetip K, Ozarska B, Belleville B, et al. Comparing two intermittent drying schedules for timber drying quality[J]. Drying Technology, 2019, 37(2): 186−197. doi: 10.1080/07373937.2018.1445638
[14] Phonetip K, Brodie G I, Ozarska B, et al. Drying timber in a solar kiln using an intermittent drying schedule of conventional laboratory kiln[J]. Drying Technology, 2019, 37(10): 1300−1312.
[15] 高建民, 王喜明. 木材干燥学 [M]. 第二版. 北京: 科学出版社, 2018. Gao J M, Wang X M. Wood drying[M]. 2nd ed. Beijing: Science Press, 2018.
[16] 何定华, 林文洁. 24种南方阔叶树材的干燥特性及窑干基准[J]. 木材工业, 1990, 4(4): 12−18. doi: 10.19455/j.mcgy.1990.04.003 He D H, Lin W J. The drying characteristics and kiln drying schedules on 24 species of hardwood from south China[J]. China Wood Industry, 1990, 4(4): 12−18. doi: 10.19455/j.mcgy.1990.04.003
[17] 何清慧. 木材干燥基准简易确定法: 百度试验法[J]. 木材工业, 1998, 12(6): 39−41. He Q H. A simple method for determination of kiln-drying schedule of wood: 100 ℃-test[J]. China Wood Industry, 1998, 12(6): 39−41.
[18] Redman A L, Bailleres H, Gilbert B P, et al. Finite element analysis of stress-related degrade during drying of Corymbia citriodora and Eucalyptus obliqua[J]. Wood Science and Technology, 2018, 52(1): 67−89. doi: 10.1007/s00226-017-0955-0
[19] Shi J, Avramidis S. Dried cell wall nanopore configuration of Douglas-fir, western red cedar and aspen heartwoods[J]. Wood Science Technology, 2018, 52(4): 1025−1037.
[20] 广东省雷州林业局, 南京林产工业学院木材干燥实验室, 中国林业科学研究院热带林业科学研究所. 雷州桉木的高温干燥试验[J]. 南京林产工业学院学报, 1979, 22(1/2): 1−13. The Forestry Office of Leizhou, Guangdong Province, Laboratory of Wood Drying, Nanjing Technological College of Forest Products, Institute of Tropical Forest, Chinese Academy of Forest Science. On the high-temperature drying of Eucalyptus wood[J]. Journal of Nanjing Technological College of Forest Products, 1979, 22(1/2): 1−13.







计量
- 文章访问数: 559
- HTML全文浏览量: 165
- PDF下载量: 76
