
Tensile creep characteristics of compression wood and normal wood under different temperatures and loads
-
摘要:目的
探究马尾松应压木和正常木的拉伸蠕变特性对温度和载荷的响应规律,旨在为马尾松木材的高附加值利用提供数据参考和基础理论依据。
方法以应压木和正常木早材组织切片为研究对象,分别采用X射线衍射仪和动态力学分析仪测定试材的微纤丝角和拉伸应力–应变曲线;在50 ~ 170 ℃范围内,利用动态力学分析仪分别获得3个应力水平和不同恒定温度水平下应压木和正常木的拉伸蠕变曲线。
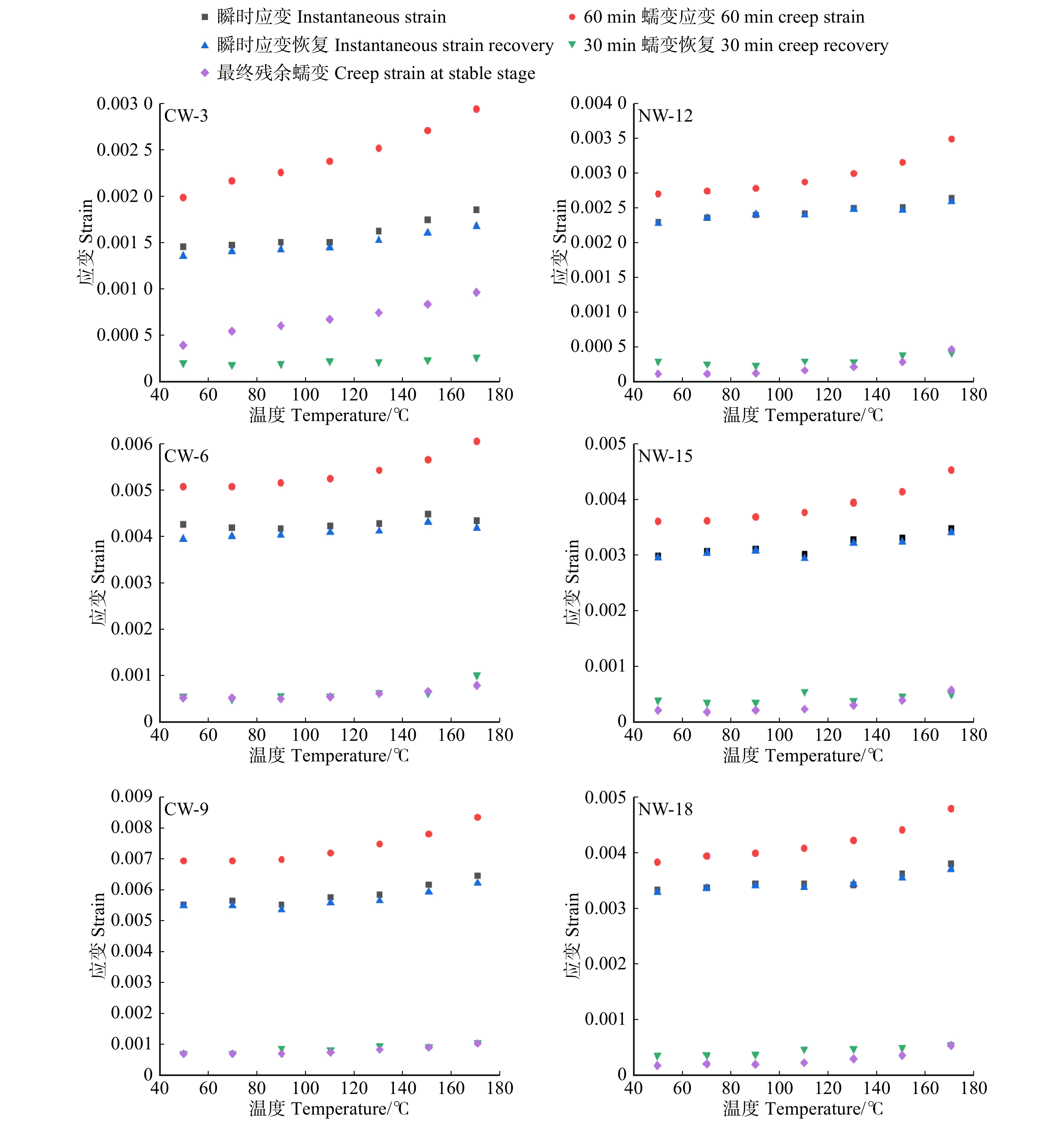
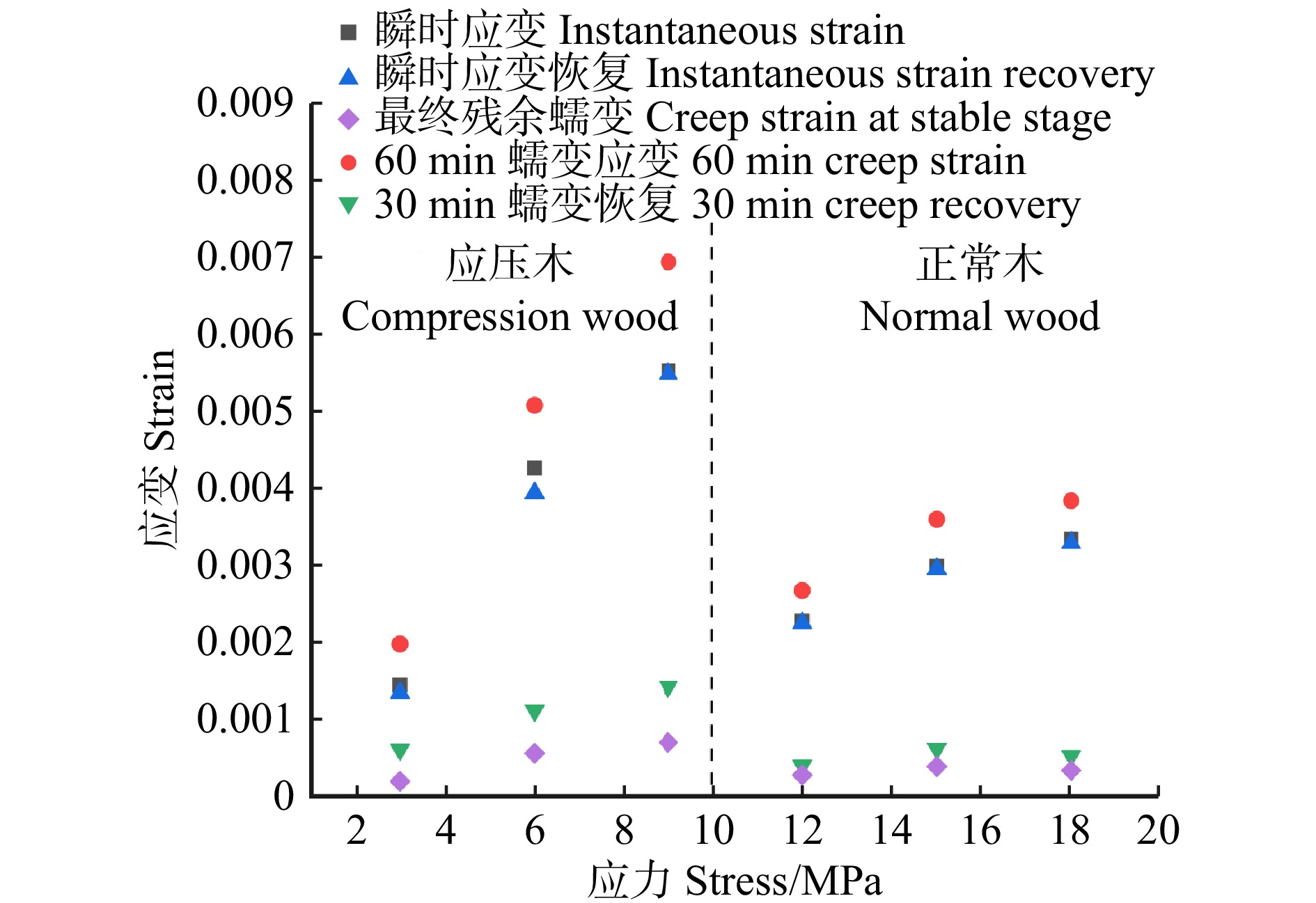
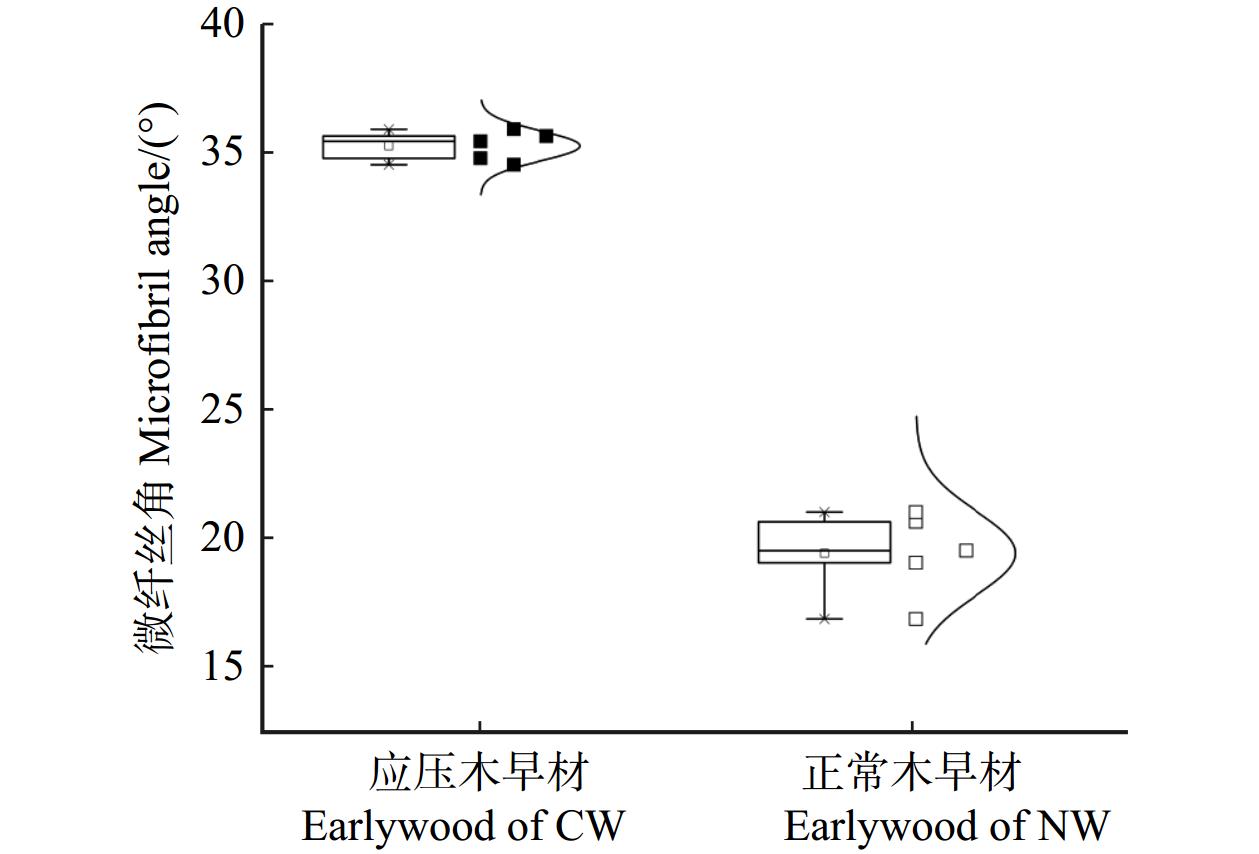
结果与正常木相比,应压木的基本密度、微纤丝角、拉伸应变、拉伸蠕变柔量和最终残余应变均较大,而拉伸应力和弹性模量较小。应压木或正常木的瞬时应变均略大于瞬时应变恢复应变量。应压木与正常木试样的瞬时应变、瞬时应变恢复量、60 min蠕变应变量和30 min蠕变恢复量均分别随温度、施加载荷的增大而增加;最终残余应变随温度升高而增加,而对施加载荷的响应无明显规律。在50 ~ 170 ℃范围内,单位应力增加引起应压木与正常木试样的瞬时应变增加量分别为0.063% ~ 0.076%和0.016% ~ 0.020%,拉伸黏弹变形增加量分别为0.078% ~ 0.095%和0.019% ~ 0.028%。
结论与正常木相比,单位应力对应压木的拉伸瞬时应变和黏弹变形增加量影响更为明显,应压木中较大的微纤丝角导致其抵抗外力变形的能力较弱。升高温度加剧了单位应力对应压木与正常木瞬时应变和拉伸黏弹变形的影响。
Abstract:ObjectiveThis research was carried to explore the difference in tensile creep behavior between compression wood (CW) and normal wood (NW) from masson pine (Pinus massoniana) under different temperature and load levels, aiming to provide the data reference and theoretical basis for high value-added utilization of plantation wood.
MethodThe tissue sections of earlywood (EW) between CW and NW were used as research objects, and the microfiber angle (MFA) and tensile stress-strain curve were characterized by X-ray diffraction and dynamic mechanical analyzer, respectively. Dynamic mechanical analyzer was also used to determine tensile creep for CW and NW specimens in the temperature range of 50 to 170 ℃ and at three stress levels.
ResultCompared with NW, the basic density, MFA, tensile strain, creep compliance and creep strain at stable stage of CW were larger, while the tensile stress and modulus of elasticity were smaller. The instantaneous strain of CW or NW specimens was slightly higher than instantaneous recovery strain. The instantaneous strain, instantaneous recovery strain, 60 min creep strain, and 30 min creep recovery of CW and NW specimens increased with increasing temperature and stress level, respectively. Furthermore, the creep strain at stable stage increased with increasing temperature, while its response to load level was not obvious. In the temperature range from 50 to 170 ℃, the increase of instantaneous strain caused by per unit stress for CW and NW was 0.063%−0.076% and 0.016%−0.020%, respectively. And the increase of creep strain in 60 min caused by per unit stress for CW and NW was 0.078%−0.095% and 0.019%−0.028%, respectively.
ConclusionCompared with NW, per unit stress shows obvious influence on the increase of tensile instantaneous strain and viscoelastic deformation, and CW with larger MFA has less ability to resist external deformation. The increasing temperature intensifies the effect of per unit stress on instantaneous strain and tensile viscoelastic deformation for NW and CW specimens.
-
Keywords:
- tensile strain /
- creep /
- loads /
- stresses /
- viscoelasticity /
- masson pine /
- compression wood
-
杨树(Populus spp.)是一种成材快、适应性强的多年生木本植物,并且是世界上分布最广的树种之一。我国杨树人工林的种植面积已持续多年位居世界第一,其天然种约有50种,分布于全国22个省区。由于杨树生长快且轮伐期短,在工业和生活中有多种用途,是建立速生丰产林的主要树种。然而,随着人工林面积不断扩大和种植树种单一化,林地生态系统变得脆弱,易发生大规模病害,造成了严重的经济损失。杨树烂皮病在华北、西北等地广泛发生,某些林场发病率超过50%。杨树溃疡病死亡率可高达30%,而黑斑病曾对南京地区的响叶杨(P. adenopoda)造成毁灭性影响。此外,落叶病曾导致杨树生长量减少约25%,木材损失约30%[1]。胶孢炭疽菌(Colletotrichum gloeosporioides)是十大植物真菌病原体之一在全球热带、亚热带和温带地区均有分布。在我国北方,由胶孢炭疽菌引起的炭疽病频繁发生,尤其是在北京、陕西、河北、甘肃等地区,毛白杨(P. tomentosa)和北京杨(P. × beijingensis)受害最为严重[2]。
目前关于炭疽菌生物学特性、相关基因表达调控、侵染机制等方面已有相关研究。炭疽菌的侵染机制可以分为胞内半活体侵染和角质层下层侵染,这两种侵染方式前期都需分生孢子萌发产生芽管和附着胞[3−4]。张晓林等[5]采用光学显微镜、扫描和透射电镜技术系统地揭示了胶孢炭疽菌在杨树叶片上的侵染特征,确认了胶孢炭疽菌侵染杨树的类型为胞内半活体侵染型。炭疽菌的分生孢子产生附着胞,附着胞成熟后形成的侵染钉穿透角质层和表皮细胞壁,最终导致次生菌丝在寄主表皮和叶肉细胞内扩展,而次生菌丝的产生是由活体营养寄生阶段转为死体营养阶段的标志。大量次级菌丝在植物细胞生长,这些菌丝分泌大量细胞壁降解酶破坏植物组织为自身生长发育获取营养,最终导致叶片坏死。已有研究揭示胶孢炭疽菌的分子调控机制,如胶孢炭疽菌CgSho1与CgMsb2共同调节病原菌信号识别和致病性[6];CgRac1是不对称发育的主要调节因子,参与病原菌形态发生和核分裂过程,其活性改变会影响胶孢炭疽菌发育并导致非致病性[7]。在拮抗胶孢炭疽菌侵染方面,研究表明芽孢杆菌可以通过合成水解酶和抗菌化合物抑制胶孢炭疽菌生长,将分离自芽孢杆菌的环四肽作用于胶孢炭疽菌,结果显示环四肽具有对胶孢炭疽菌显著抗性[8],Zhang等[9]研究发现施加外源油菜素内酯(BR)显著抑制了胶孢炭疽菌的传播,提高了苯丙氨酸解氨酶(PAL)活性,表明外源BR可能通过激活苯丙烷合成来提高抗病性。
植物响应胶孢炭疽菌侵染的机制也有相关研究,如草莓(Fragaria × ananassa)[10]、茶树(Camellia sinensis)[11]、柱花草(Stylosanthes spp.)[12]、鳄梨(Persea americana)[13]等。陈哲等[10]研究发现草莓与炭疽菌互作过程中,类黄酮生物合成、谷胱甘肽代谢、植物激素信号转导等植物抵御胁迫相关的途径富集到不少基因。Fang等[14]利用病毒介导的基因沉默技术构建HSP17.4沉默株系,发现草莓HSP17.4能诱导水杨酸和茉莉酸途径协同作用,从而提高对炭疽病的抗性。Jiang等[12]结合转录组学和代谢组学解析柱花草(Stylosanthes spp.)对炭疽菌的防御反应,差异表达基因功能富集分析结果表明黄酮类生物合成途径中酶的基因强烈响应病原菌诱导。Mehmood等[15]以抗性草莓为试验材料,转录组数据表明胶孢炭疽菌侵染后宿主防御基因与萜类合成基因共表达,代谢组学分析结果表明萜类化合物在叶片中快速积累。然而,杨树响应胶孢炭疽菌胁迫的分子机制的研究相对薄弱,转录调控机制、关键抗病通路目前仍不清晰。基于以上科学问题,本研究以毛白杨无性系LM50为研究材料,对胶孢炭疽菌侵染后的杨树叶片进行高通量转录组测序,分析病原菌侵染下转录水平变化,发掘杨树抗病相关通路和关键基因,为杨树抗病新品种的选育提供重要理论依据。
1. 试验材料和方法
1.1 试验材料
以毛白杨(Populus tomentosa)无性系LM50为研究材料,将生长1个月的组培苗经过炼苗后移栽到灭菌过的大田土中,置于温室条件(光照12 h/d,相对湿度约60%,温度(25 ± 2) ℃)下连续培养3个月,选取长势相近的幼苗作为试验材料,3棵苗子为一组。
试验所用菌株为胶孢炭疽菌BJ12,分离自北京林业大学校内毛白杨叶片,保存于北京林业大学林木花卉遗传育种实验室。病原菌侵染采用菌饼接种法,胶孢炭疽菌在PDA培养基活化后,置于25 ℃培养箱生长5 ~ 7 d,用孔径1 cm的打孔器打孔,将有菌面贴于叶片表面,侵染6 d后剪取叶片,对照接种无菌PDA培养基做同样处理[16]。对照和处理选取相同位置的叶片,采集到的样品液氮速冻处理后保存于−80 ℃冰箱备用。
1.2 叶片酶活性测定
酶粗提液的制备参照费莉玢[17]所用方法,每个处理重复3次,提取液保存于−80℃冰箱备用。丙二醛(MDA)含量和3种抗氧化酶,包括多酚氧化酶(PPO)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)均采用酶联免疫法,绘制标准曲线后计算得到样品浓度,具体试验步骤按照上海沪鼎生物科技有限公司酶联免疫分析试剂盒操作。
1.3 转录组测序和生物信息学分析
叶片RNA提取和纯度检测参照宋润先等[18]采用的方法,以检测合格的RNA构建文库,利用Illumina HiSeqTM4000高通量平台进行双端测序获得原始测序数据,文库构建和测序工作交由诺禾致源有限公司完成。
以毛果杨(Populus trichocarpa;version 3.1)基因组序列[19]作为参考,从phytozome(https://phytozome-next.jgi.doe.gov/)开放平台下载相关文件,同时对测序得到的原始数据进行过滤质控,得到高质量转录组数据(clean reads)。利用Hisat2(version 2.0.9)将clean reads比对到参考基因组[20],使用Cuffdiff软件进行差异基因分析[21]。为最大程度降低假阳性,筛选差异表达基因的条件设定为:差异倍数(fold change,FC) > 1.5,显著性P < 0.05。使用网页版工具agriGO(http://systemsbiology.cau.edu.cn/agriGOv2/index.php/)进行差异基因GO富集分析[22];利用TBtools(version 1.068)进行KEGG富集分析[23];使用MapMan(version 3.5.1)分析差异基因响应模式[24]。
1.4 差异表达基因的qRT-PCR验证
基于转录组数据的分析结果,选取6个可能与植物抗性相关的上调差异表达基因进行qRT-PCR分析,验证转录组测序分析数据的可靠性,以0 h样品为对照,试验流程参照Chen等[25]使用的方法。数据处理使用2−ΔΔCT计算基因的相对表达量[26]。通过在线网站(https://sg.idtdna.com/Scitools/Applications/RealTimePCR/)设计引物,并由生工生物工程(上海)有限公司合成。
2. 结果和分析
2.1 表型和生理指标测定
侵染第6 天对杨树对照及试验组叶片生理指标进行分析,测定MDA含量及PPO、SOD、CAT活性。MDA含量和CAT、SOD活性与对照相比均有不同程度增加,且差异极显著(图1)。MDA含量比对照高出213.41%,CAT、SOD和PPO活性分别比对照高出393.16%、151.39%和116.56%。
![]() 图 1 胶孢炭疽菌胁迫下杨树叶片表型变化及MDA浓度 (i) 及PPO (j)、CAT (k)、SOD (l) 活性变化a为胶孢炭疽菌接种方式; b ~ h分别为侵染后0、1、2、3、4、5、6 d的叶片;a ~ h叶片标尺 = 2 cm。*、**、***分别表示对照和处理之间差异显著性水平为P<0.05、P<0.01、P<0.001。 a, inoculation method of Colletotrichum gloeosporioides; b−h, leaf symptoms of 0 to 6 d after inoculation; leaf bar = 2 cm. *, **, and *** denote significant differences between controls and treatments at the P < 0.05, P < 0.01, and P < 0.001 levels, respectively.Figure 1. Leaf symptoms and change of MDA (i), PPO (j), CAT (k), SOD (l) activity after inoculation
图 1 胶孢炭疽菌胁迫下杨树叶片表型变化及MDA浓度 (i) 及PPO (j)、CAT (k)、SOD (l) 活性变化a为胶孢炭疽菌接种方式; b ~ h分别为侵染后0、1、2、3、4、5、6 d的叶片;a ~ h叶片标尺 = 2 cm。*、**、***分别表示对照和处理之间差异显著性水平为P<0.05、P<0.01、P<0.001。 a, inoculation method of Colletotrichum gloeosporioides; b−h, leaf symptoms of 0 to 6 d after inoculation; leaf bar = 2 cm. *, **, and *** denote significant differences between controls and treatments at the P < 0.05, P < 0.01, and P < 0.001 levels, respectively.Figure 1. Leaf symptoms and change of MDA (i), PPO (j), CAT (k), SOD (l) activity after inoculation2.2 测序结果统计
如表1所示,各样品最终得到clean data至少3 Gbp,过滤条件下,数据总体通过率平均达到97.4%以上,各样品Q20占比至少98%,Q30占比均在93%以上。质控过程中,GC含量保持在43.7% ~ 45.1%之间,表明测序技术对样本数据没有较高的偏好性,且样品本身无污染。以上结果说明本次测序的cDNA文库质量高,可以进行后续的生物信息学分析研究。
表 1 过滤质控结果统计表Table 1. Statistics about filter and quality control results样本名称
Sample name数据量
Data amount/Gbp合格片段比率
Percentage of reads passed filter/%碱基错误率
Base error rate/%Q20/% Q30/% GC含量
GC content/%0h_1 3.76 98.76 0.04 98.25 94.69 44.70 0h_2 3.77 98.50 0.04 98.27 94.74 45.10 0h_3 2.93 98.14 0.04 98.25 94.69 44.60 6d_1 3.00 98.12 0.04 98.16 94.58 44.78 6d_2 3.56 98.16 0.04 98.20 94.63 44.54 6d_3 3.60 97.40 0.04 98.12 94.48 43.70 注:Q20为质量值达到20的碱基占总碱基的比例;Q30为质量值达到30的碱基占总碱基的比例。Notes: Q20, proportion of base with a mass value of 20 to the total base; Q30, proportion of base with a mass value of 30 to the total base. 2.3 差异表达分析
表 2 显著性差异表达基因统计分析Table 2. Statistical analysis of significantly differential expressed genesFC 上调基因数目
Up-regulated gene number下调基因数目
Down-regulated gene number> 1.5 2 262 2 285 1.5 ~ 3 1 584(70.0%) 1 807(79.1%) 3 ~ 5 542(23.96%) 417(18.25%) > 5 136(6.0%) 61(2.7%) 注:FC.差异倍数。括号内数据为上调基因数量或下调基因数量占差异基因数量的百分比。Notes: FC, fold change. In parentheses, the data is the percentage of the number of up-regulated genes or down-regulated genes in the number of differential genes. 为探究病原菌侵染下杨树转录水平的变化,首先对差异基因表达进行严格筛选,条件为差异倍数(fold change,FC) > 1.5且P < 0.05,结果表明共有4 547个基因在转录水平上存在显著性差异,其中上调基因为2 262个,下调基因为2 285个(表2)。根据FC进行分类,差异基因倍数变化大多集中在1.5 ~ 5.0倍之间。大于5.0倍的差异基因中,上调基因共136个,如PR-1(FC = 9.9)、WRKY51(FC = 5.3)、细胞色素P450(FC = 5.1)等;下调基因共61个,包括聚半乳糖醛酸酶(FC = −5.8)、果胶裂解酶(FC = −5.3)等。
2.4 差异表达基因的GO富集分析
对筛选到的差异基因进行GO富集分析,4 547个基因分别注释到细胞组分、分子功能和生物学过程3大类中。其中,富集到差异基因最多的3个分子功能条目是催化活性、转录活性、氧化还原酶活性;生物学过程富集到差异基因最多的是代谢途径、细胞代谢途径和大分子代谢途径3个条目;线粒体、细胞壁合成和代谢是细胞组分中富集到差异基因最多的2个条目。
进一步分析表明,上调差异表达基因富集到135个通路中(P < 0.05),细胞组分、分子功能和生物学过程分别占总数的10%、35%和55%(图2a)。生物学过程显著富集到的GO条目包括氧化还原、代谢过程、呼吸气体交换、胁迫响应、多细胞生物进程等,分子功能显著富集到氧化还原酶活性、血红素结合、四吡咯结合、铁离子结合、催化活性等条目中;而线粒体组分、线粒体、线粒体膜、细胞器等GO条目富集显著到细胞组分(图2c)。下调差异表达基因富集到213个通路中,细胞组分、分子功能和生物学过程分别占总数的5%、45%和50%(图2b)。碳代谢、微管运动、脂代谢、细胞多糖代谢、葡聚糖代谢等是差异显著的生物学过程条目,分子功能通路中显著富集条目包括催化活性、水解酶活性、微管马达活性、纤维素合酶等,细胞组分显著富集到细胞壁、外部封装结构和质外体等(图2d)。
![]() 图 2 差异基因GO功能富集分析a、c为上调基因;b、d为下调基因。a and c are up-regulation genes; b and d are down-regulation genes.Figure 2. GO function enrichment of different experssed genes (DEGs)
图 2 差异基因GO功能富集分析a、c为上调基因;b、d为下调基因。a and c are up-regulation genes; b and d are down-regulation genes.Figure 2. GO function enrichment of different experssed genes (DEGs)2.5 差异表达基因的KEGG富集分析
对差异表达基因进行KEGG通路富集分析,共注释到204条KEGG通路上,其中50条通路被显著富集(P < 0.05),选取前30条作图(图3)。富集到的通路中,代谢途径包含类目最多,包括糖类、脂类、氨基酸、不饱和脂肪酸等多种物质,这与GO富集得到的结果一致。代谢途径上调差异表达基因主要富集到谷胱甘肽、苯丙烷、类黄酮、氨基酸、亚麻酸、萜类化合物等,下调差异表达基因则富集到脂质、碳、卟啉和叶绿素、淀粉和蔗糖、长链脂肪酸、磷酸肌醇、维生素、不饱和脂肪酸等(表3)。
![]() 图 3 差异基因KEGG富集分析Q. 富集程度。下同。Q, enrichment degree. The same below.Figure 3. KEGG enrichment analysis of DEGs表 3 差异表达基因KEGG富集分析Table 3. KEGG enrichment analysis of DEGs
图 3 差异基因KEGG富集分析Q. 富集程度。下同。Q, enrichment degree. The same below.Figure 3. KEGG enrichment analysis of DEGs表 3 差异表达基因KEGG富集分析Table 3. KEGG enrichment analysis of DEGsKO号 KO ID 代谢途径 Metabolic pathway 基因数量 Gene number Q 上调基因 Up-regulation gene Ko09105 氨基酸代谢 Amino acid metabolism 55 0.026 Ko00940 苯丙烷生物合成 Phenylpropanoid biosynthesis 34 < 0.001 Ko09106 其他氨基酸代谢 Metabolism of other amino acids 34 0.026 Ko09108 辅酶因子和维生素代谢 Metabolism of cofactors and vitamins 30 0.136 Ko09109 萜类和多酮类代谢 Metabolism of terpenoids and polyketides 27 0.057 Ko00480 谷胱甘肽代谢 Glutathione metabolism 25 0.000 Ko00630 乙醛酸和二羧酸代谢 Glyoxylate and dicarboxylate metabolism 14 0.055 Ko00941 类黄酮生物合成 Flavonoid biosynthesis 13 0.001 Ko00592 亚麻酸代谢 alpha-Linolenic acid metabolism 11 0.053 Ko00900 萜类化合物生物合成 Terpenoid backbone biosynthesis 11 0.059 Ko00350 酪氨酸代谢 Tyrosine metabolism 10 0.181 Ko00280 缬氨酸、亮氨酸和异亮氨酸降解 Valine, leucine and isoleucine degradation 9 0.117 Ko00910 氮代谢 Nitrogen metabolism 8 0.056 下调基因 Down-regulation gene Ko09101 碳代谢 Carbohydrate metabolism 92 < 0.001 Ko09103 脂代谢 Lipid metabolism 58 < 0.001 Ko09110 其他次生代谢产物的生物合成 Biosynthesis of other secondary metabolites 36 0.023 Ko09108 辅酶因子和维生素的代谢 Metabolism of cofactors and vitamins 31 0.022 Ko09106 其他氨基酸代谢 Metabolism of other amino acids 25 0.188 Ko00500 淀粉和蔗糖代谢 Starch and sucrose metabolism 23 0.001 Ko00940 苯丙烷的生物合成 Phenylpropanoid biosynthesis 23 0.062 Ko00040 戊糖、葡萄糖醛酸转换 Pentose and glucuronate interconversions 15 0.030 Ko00860 卟啉和叶绿素代谢 Porphyrin and chlorophyll metabolism 13 < 0.001 Ko00061 脂肪酸生物合成 Fatty acid biosynthesis 12 0.008 Ko00562 磷酸肌醇代谢 Inositol phosphate metabolism 12 0.018 Ko00564 甘油磷脂 Glycerophospholipid metabolism 12 0.145 Ko00062 长链脂肪酸 Fatty acid elongation 10 0.008 Ko00460 氰基氨基酸代谢 Cyanoamino acid metabolism 9 0.070 Ko00561 甘油酯代谢 Glycerolipid metabolism 8 0.187 Ko00511 其他多糖降解 Other glycan degradation 7 0.012 苯丙烷合成途径共有57个基因发生变化,其中34个基因上调;谷胱甘肽代谢途径36个基因发生变化,其中25个上调;类黄酮合成途径20个基因发生变化,其中14个上调。值得注意的是,苯丙烷代谢是参与植物防御反应的主要次级代谢通路之一,它的分支又是类黄酮的合成途径,因此对类黄酮和苯丙烷生物合成通路中的基因进行了注释分析[27]。
苯丙烷类化合物合成通路中的关键酶,其富集到的基因大多呈上调表达趋势,主要包括PAL(Potri.010G224100)、4CL(Potri.003G188500、Potri.001G036900)、CHS(Potri.014G145100)、LDOX(Potri.004G030700)、CCoAOMT(Potri.001G304800)、CAD(Potri.009G095800)、POX(Potri.005G135300、Potri.013G156500、Potri.014G143200)等。另有少量基因呈下调表达趋势,如PAL(Potri.006G126800)、4CL(Potri.018G094200)、FLS(Potri.004G139500、Potri.004G139700)等。
2.6 杨树响应炭疽菌侵染的转录因子及MapMan分析
为揭示转录因子响应胶孢炭疽菌侵染的表达模式,将差异基因和毛果杨转录因子数据库进行比对,共有322条基因被注释为38个家族的转录因子(表4)。MYB、ERF、bHLH转录因子数量最多,分别为36、34、31个,值得注意的是WRKY转录因子富集到27个,且全部上调表达(图4)。
表 4 转录因子统计表Table 4. Statistics of transcription factors转录因子家族
Transcription factor family数量
Number转录因子家族
Transcription factor family数量
Number转录因子家族
Transcription factor family数量
NumberMYB 36 SBP 7 Nin-like 2 ERF 34 GRF 6 LSD 1 bHLH 31 HSF 6 ARR-B 1 WRKY 27 ZF-HD 5 SRS 1 NAC 23 TALE 5 BES1 1 bZIP 14 CO-like 5 Whirly 1 HD-ZIP 13 Trihelix 5 AP2 1 C2H2 13 TCP 4 C3H 1 MYB_related 11 LBD 4 E2F/DP 1 GRAS 10 DBB 4 NF-YA 1 Dof 10 B3 4 WOX 1 GATA 8 YABBY 3 RAV 1 ARF 8 MIKC_MADS 3 G2-like 8 STAT 2 对筛选到的4 547个基因进行MapMan分析,以差异基因生物学功能划分为不同层级,得到杨树–病原菌互作的调控网络[23]。结果显示在病原菌诱导下,与激素信号(生长素、乙烯、茉莉酸、水杨酸)、细胞壁、抗逆、次级代谢、MAPK、转录因子等关联的基因被激活(图5)。植物在受到病原菌侵染时启动保护酶系统,保护宿主免受ROS造成的氧化损伤,POD则作为重要抗氧化酶清除体内自由基。显然,在杨树抵御病原菌胁迫的过程中需要大量POD的积累,MapMan结果显示氧化还原相关的差异基因大多呈上调表达趋势,如氧化还原态、过氧化物和谷胱甘肽-S-转移酶,其中,10个编码POD合成的基因受到活性氧信号激发大量表达。ROS引发植物产生过敏反应(hypersensitive response,HR),宿主体内激素含量随之大量增加,这些激素作为二级信号进而激活防御反应,即PR基因的大量表达[28]。本研究结果中19个PR基因上调表达,编码抗菌蛋白CC-NBS-LRR、NBS-LRR、TIR-NBS-LRR等在抗病过程中的积累。
![]() 图 5 MapMan分析杨树–胶孢炭疽菌互作相关的差异表达基因Figure 5. DEGs involved in poplar-C. gloeosporioides interaction analysis by MapMan
图 5 MapMan分析杨树–胶孢炭疽菌互作相关的差异表达基因Figure 5. DEGs involved in poplar-C. gloeosporioides interaction analysis by MapMan此外,次级代谢产物在抗病过程中也发挥关键作用,代谢通路差异基因注释到多种酶,包括CHS、DFR、LDOX、CCR、CLA等苯丙烷合成关键酶上调表达,这与KEGG富集结果相似。MapMan结果表明大量转录因子也被激活表达,其中,11个MYB转录因子和WRKY家族全部20个转录因子上调表达,包括WRKY75(FC = 7.7)、WRKY51(FC = 5.3)、WRKY70(FC = 5.3)、MYB62(FC = 4.5)、MYB3(FC = 3.8)、WRKY18(FC = 3.7)。
2.7 qRT-PCR验证
为检验转录组数据的真实性和可靠性,根据数据分析结果选取6个关键基因,通过qRT-PCR检测其表达水平。分析结果表明,选取的基因表达趋势与转录组测序结果一致,表明RNA-seq结果可靠(图6)。
苯丙烷代谢途径起始于苯丙氨酸,首先在苯丙氨酸解氨酶(PAL)的作用下转化为反式肉桂酸,再经过肉桂酸羟化酶(C4H)催化形成对香豆酸,然后经4-香豆酸–辅酶A连接酶(4CL)将其转化为对香豆酰辅酶A。随后分为两个分支,经查尔酮合成酶(CHS)、查尔酮异构酶(CHI)、二氢黄酮醇-4-还原酶(DFR)、无色花青素还原酶(LDOX)、黄酮醇合酶(FLS)等催化形成类黄酮化合物,另一分支经咖啡酰辅A-O-甲基转移酶(CCoAOMT)、肉桂醇脱氢酶(CAD)、过氧化物酶(POX)等催化形成木质素[29]。PAL既是苯丙氨酸代谢途径的关键酶也是限速酶,CHS是连接苯丙烷生物合成和类黄酮生物合成途径的关键酶,为黄酮、黄酮醇及其他物质的合成提供了基本的碳架结构,4CL则调控苯丙氨酸代谢途径的最后一步反应,催化形成木质素类和黄酮类化合物。由图可知,苯丙烷合成通路的关键酶基因在接种胶孢炭疽菌后表达上调,包括4CL(Potri.018G094200)、PAL(Potri.010G224100)、POX(Potri.005G135300)、CHS(Potri.014G145100)。此外,转录因子MYB62(Potri.008G122100)和ABR1(Potri.005G087200)作为植物抗逆过程中的重要调控因子,在病原菌胁迫条件下表达量显著提高。
3. 讨 论
植物受到病原菌胁迫后,会激活多条抗逆通路来抵御病原菌侵染,KEGG富集结果表明这些抗逆通路大多与生长、代谢和激素相关。转录因子在宿主生长发育调控及抗逆途径中起到重要作用,广泛存在于植物体内,能够与基因启动子区域特异性结合,激活或抑制下游基因的表达。ERF转录因子家族作为逆境信号参与到胁迫信号交叉途径中[30],同时,在水杨酸、茉莉酸、乙烯、脱落酸等植物激素胁迫应答反应中起到重要作用[31]。WRKY同样是植物的重要转录因子家族,可以作为防御基因的调节因子,且是不同信号途径的靶标[32]。拟南芥AtERF2转录因子是茉莉酸胁迫响应基因的正调控因子,介导了对尖孢镰刀菌(Fusarium oxysporum)的抗性,小麦植株ERF转录因子可以激活PR基因,将其转入烟草植株中过表达,提高了对链格孢菌(Alternaria alternate)的抗病性[33]。ERF转录因子也可以通过ABA信号通路,负调控对真菌病原菌的抗性,如过量ABA导致水稻内源ET水平降低,从而提高了对稻瘟病的敏感性[34]。MapMan分析结果表明,杨树与胶孢炭疽菌互作过程中,激素相关基因大多受到病原菌正向调控,如89%乙烯相关基因上调表达,表明乙烯水平可能作为信号分子影响下游基因的表达,进而实现抗病目的。然而,病原菌胁迫下的激素信号分子并不是独立发挥作用的,激素信号分子之间存在相互串扰。本研究中WRKY和ERF转录因子与激素信号分子之间的调控关系尚不明确,有待进一步研究。
活性氧(ROS)含量提高是植物响应病原菌侵染的重要特征之一,持续累积的ROS会对宿主细胞造成损伤[35]。随着ROS水平的变化,植物清除系统中的抗氧化酶活性也发生大幅变化[36]。如木薯叶片被病原菌侵染后,CAT活性随侵染时间延长而增大,PPO活性在侵染初期变化不显著,随着时间的推移迅速增强,表明相关酶机制启动以应对病原菌侵染[37]。炭疽菌的侵染同样可以导致茶树叶片CAT、SOD活性提高[38]。MDA是参与细胞膜过氧化作用的主要成分之一,含量越高对细胞膜伤害越大。杨树叶片受到炭疽菌侵染6 d后MDA含量显著提高,说明此时胶孢炭疽菌已经对植物的活性氧清除系统产生破坏,并导致了细胞膜脂化。在此过程中,参与植物体内抗氧化及活性氧清除过程的过氧化物酶、双加氧酶、抗坏血酸过氧化物酶、无色花青素还原酶等基因表达上调,酶活测定结果同样表明抗氧化酶(CAT、PPO、SOD)活性相比对照得到显著提高。说明杨树受到胶孢炭疽菌侵染后,抗氧化酶在清除ROS、维持宿主体内ROS平衡,缓解病原菌胁迫造成的氧化损伤中发挥了重要作用。
苯丙烷代谢途径激活是植物响应病原菌侵染的标志之一,该途径在多种酶催化作用下分别合成黄酮类化合物和木质素,苯丙氨酸作为多种酚类化合物(如黄酮类化合物、异黄酮类化合物、植物激素)的前体,在宿主防御反应中起到重要作用[39−40]。类黄酮通过清除体内自由基来保护植物免受外界侵扰,木质素沉积在细胞壁上可以提高组织机械强度并有效抑制病原菌生长,单宁可以有效抑制真菌的体外生长[41]。黄瓜CsMYB60促进关键基因Cs4CL、CsCHS的表达,调控黄酮醇和单宁合成,诱导单宁在根部积累,进而增强对根腐病的抗性[42−43]。在茶树抗病品种“ZC108”中,CAD相关基因的表达水平高于感病品种“LJ43”,且在受到炭疽菌侵染后,PAL调控基因显著上调表达[44]。木质素通过加固细胞壁的形式参与植物防御反应,如番茄被V. dahliae侵染后,抗病品种木质素含量比感病品种高4%[45]。本研究中,胶孢炭疽菌侵染杨树叶片后苯丙烷合成通路及类黄酮合成通路受到强烈诱导,PAL、4CL、CHS等关键酶合成相关基因表达量提高,因此推测,胶孢炭疽菌侵染可能导致了植物体内类黄酮含量提高和木质素积累,这些抗病物质的增加提高了杨树抗病性。
需注意的是,本研究荧光定量PCR结果与转录组测序结果虽然呈现相同表达趋势,但表达倍数存在差异。通过比较重复性、准确性、数据统计等多方面因素,RNA-seq在测定基因表达水平方面整体水平更高[46−47]。研究发现qRT-PCR检测结果与RNA-seq测序结果大多是一致的,但也有相当一部分基因的表达倍数存在差异,检测结果的相关性最低可低至0.58,仅中度相关[48]。此外,qRT-PCR检测结果受到模板、特异性引物、仪器设备、扩增效率等多种因素影响。qRT-PCR检测到的信号取决于模板的质量,本研究所用模板cDNA是在本实验室制备的,而转录组测序所用模板cDNA为公司制备,二者之间可能存在差异。引物特异性和扩增效率密切相关,也是影响qRT-PCR结果的重要原因[49]。本研究所用引物和模板可能是导致qRT-PCR检测结果低于RNA-seq结果的重要因素。
病原菌不同菌株之间的致病性强弱不同,菌饼接种方式虽然比较老,但对于部分未建立孢子液接种方法和孢子液侵染效率较低的植物来说,菌饼接种方式更加高效。如利用果生炭疽菌孢子液侵染苹果叶片,发现抗病品种叶片表面没有形成次生菌丝,孢子液侵染不能使抗病品种发病[50]。王杰等[16]对红叶石楠炭疽病病原进行研究时发现,菌饼法的侵染效果更好。此外,后续实验结果也证明,本研究得到的数据是可靠的。我们利用生物信息学分析筛选到多个关键基因,包括WRKY18和bZIP44。目前已通过农杆菌介导的遗传转化技术成功构建杨树bZIP44过表达株系,表型观测结果显示过表达株系的平均株高在生长1个月、2个月和3个月时,分别高出对照30%、21%和14%。杨树叶片侵染结果表明过表达株系的抗病性得到显著提高(P < 0.05),通过酵母双杂交实验及DAP-seq试验已经筛选到的bZIP44互作蛋白及下游靶基因(研究结果未发表)。
为进一步探究关键基因bZIP44在杨树抗病过程中的具体作用机制,后续我们将继续开展分子实验对其进行功能研究,解析关键基因调控网络,结合代谢组学验证黄酮类化合物和木质素等抑菌物质在杨树响应胶孢炭疽菌侵染过程中的作用。综上所述,本研究获得的杨树响应病原菌胁迫的生理指标及转录组数据,将为今后解析杨树与病原菌互作机制提供重要的数据支撑,同时也为杨树抗逆新品种的培育提供理论基础。
4. 结 论
本研究对受到胶孢炭疽菌侵染的杨树叶片进行转录组测序,筛选得到4 547个差异表达的基因,并对差异表达基因进行生物信息学分析。结果表明:杨树受到病原菌侵染后,一方面利用抗氧化酶系统清除宿主体内不断累积的ROS;另一方面编码黄酮类化合物和木质素等抗病物质的合成,从而提高杨树对胶孢炭疽菌的抗病性。胶孢炭疽菌侵染导致活性氧含量增加,抗氧化物酶基因随之上调表达,激活了植物体内清除氧系统(CAT、PPO、SOD)以维持活性氧代谢平衡。同时,转录组数据及qRT-PCR结果均表明类黄酮、木质素等抑菌物质调控基因被诱导,可能促进了抑菌物质的合成和积累,进而达到抑制了病原菌生长的目的。
-
![]()
图 1 马尾松应压木(CW)和正常木(NW)早材试样微纤丝角的箱线–正态图
Figure 1. Boxplots of microfibril angle of earlywood between compression wood (CW) and normal wood (NW) from masson pine
![]()
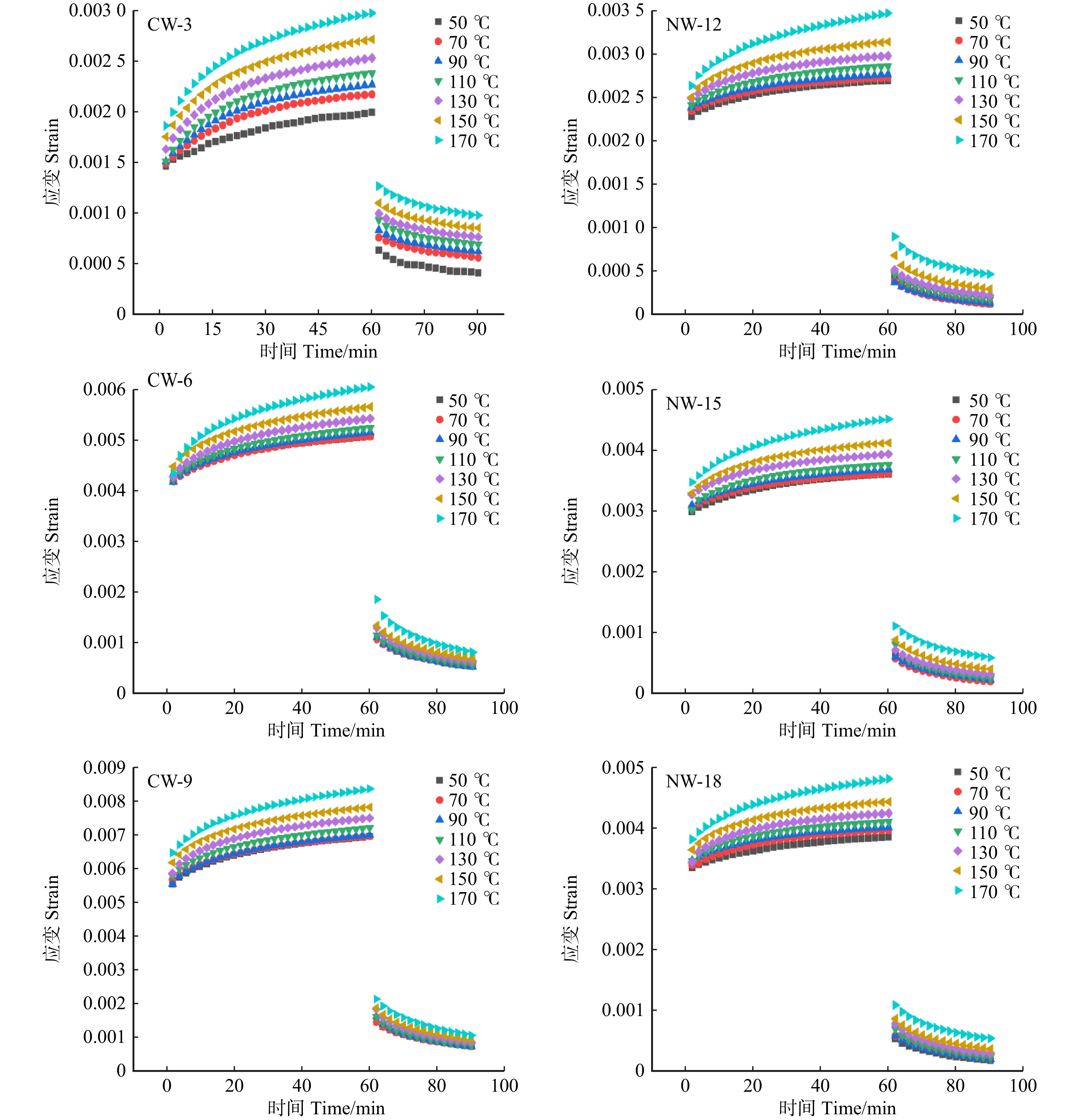
图 2 不同温度水平下应压木和正常木早材试样拉伸蠕变曲线
CW-3、CW-6和CW-9分别代表蠕变应力值3、6和9 MPa的应压木。NW-12、NW-15和NW-18分别代表蠕变应力值12、15和18 MPa的正常木。下同!CW-3, CW-6, and CW-9 represent compressive wood with creep stress values of 3, 6, and 9 MPa, respectively. NW-12, NW-15, and NW-18 represent normal wood with creep stress values of 12, 15, and 18 MPa, respectively. The same below!
Figure 2. Tensile creep curves of earlywood between CW and NW at different temperature levels
![]()
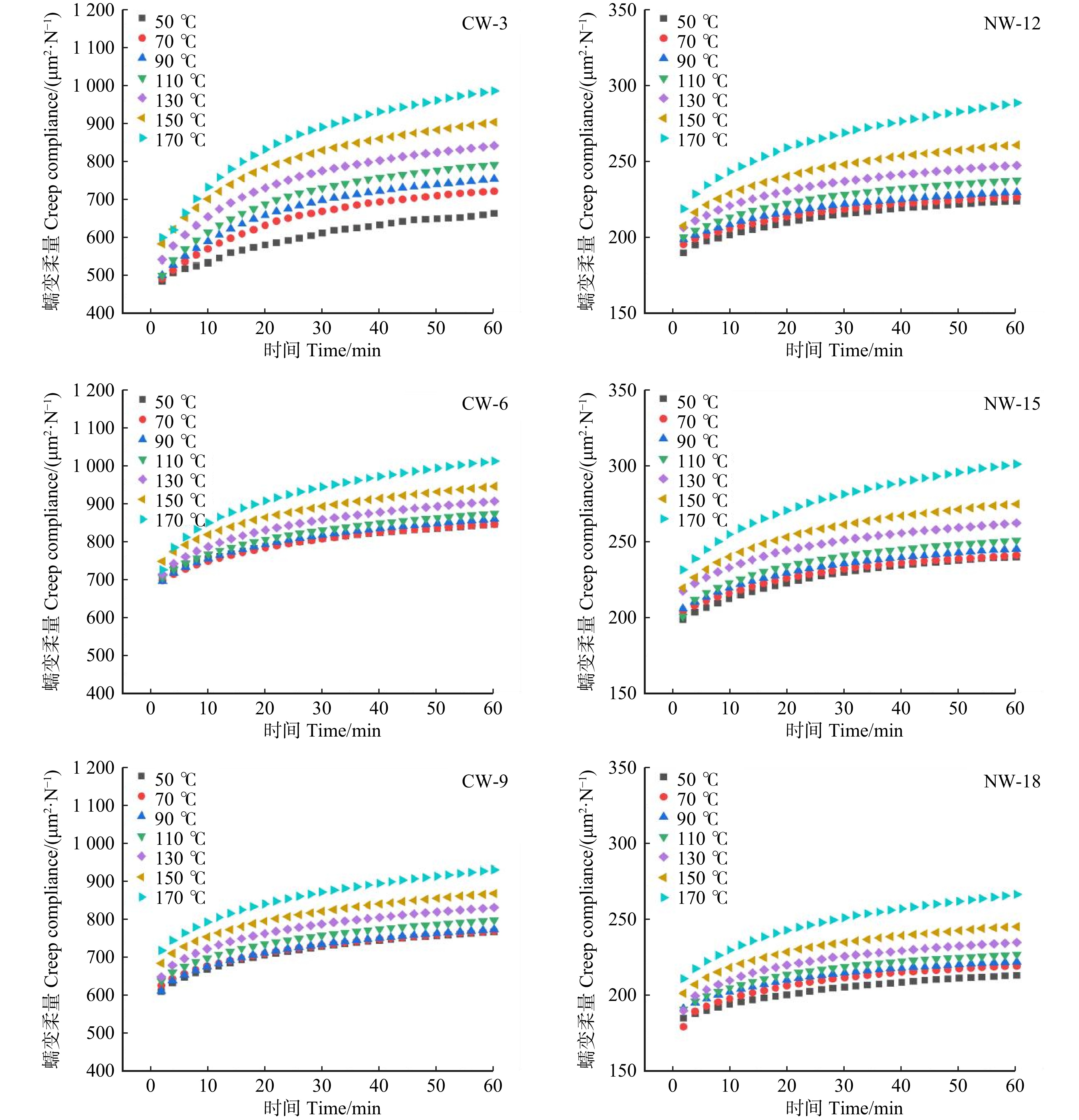
图 3 不同温度水平下应压木和正常木早材试样拉伸蠕变柔量
Figure 3. Tensile creep compliance of EW between CW and NW at different temperature levels
![]()
图 4 不同温度水平下应压木和正常木早材试样拉伸蠕变参数
Figure 4. Tensile creep parameters of EW between CW and NW at different temperature levels
![]()
图 5 不同应力水平下应压木和正常木早材试样拉伸蠕变参数(50 ℃)
Figure 5. Tensile creep parameters of EW between CW and NW at different stress levels at 50 ℃
![]()
图 6 不同应力水平下应压木和正常木早材试样拉伸蠕变参数(70 ~ 170 ℃)
Figure 6. Tensile creep parameters of EW between CW and NW at different stress levels in the temperature range of 70 to 170 ℃
表 1 马尾松应压木和正常木早材试样拉伸力学参数
Table 1 Tensile mechanical parameters of earlywood between CW and NW from masson pine
试样
Specimen拉伸弹性模量 Tensile elastic modulus 应力 Stress 应变 Strain 均值
Mean value/MPaCV/% 均值
Mean value/MPaCV/% 均值
Mean value/%CV/% 应压木早材 Earlywood of CW 1 164 10.98 15.97 0.30 1.47 14.65 正常木早材 Earlywood of NW 2 680 5.06 19.38 1.66 0.65 7.53  下载: 导出CSV
下载: 导出CSV
-
[1] Stevanic J S, Salmén L. Molecular origin of mechano-sorptive creep in cellulosic fibres[J]. Carbohydrate Polymers, 2020, 230: 115615. doi: 10.1016/j.carbpol.2019.115615
[2] Salmén L, Stevanic J S, Olsson A M. Contribution of lignin to the strength properties in wood fibres studied by dynamic FTIR spectroscopy and dynamic mechanical analysis (DMA)[J]. Holzforschung, 2016, 70(12): 1155−1163. doi: 10.1515/hf-2016-0050
[3] Peng H, Salmén L, Stevanic J S, et al. Structural organization of the cell wall polymers in compression wood as revealed by FTIR microspectroscopy[J]. Planta, 2019, 250(1): 163−171. doi: 10.1007/s00425-019-03158-7
[4] Salmén L, Bergström E. Cellulose structural arrangement in relation to spectral changes in tensile loading FTIR[J]. Cellulose, 2009, 16: 975−982. doi: 10.1007/s10570-009-9331-z
[5] Roszyk E, Mania P, Moliński W. The influence of microfibril angle on creep of Scotch pine wood under tensile stress along the grains[J]. Wood Research, 2012, 57(3): 347−358.
[6] 王东, 林兰英, 傅峰, 等. 傅里叶变换红外光谱研究拉伸过程中应压木主要化学组分的响应规律[J]. 光谱学与光谱分析, 2020, 40(11): 3585−3589. Wang D, Lin L Y, Fu F, et al. Chemical construction changes of compression wood main components in longitudinal tension by the FTIR analysis[J]. Spectroscopy and Spectral Analysis, 2020, 40(11): 3585−3589.
[7] Peng H, Salmén L, Jiang J L, et al. Creep properties of compression wood fiber[J]. Wood Science and Technology, 2020, 54: 1497−1510. doi: 10.1007/s00226-020-01221-1
[8] Purusatama B D, Choi J K, Lee S H, et al. Microfibril angle, crystalline characteristics, and chemical compounds of reaction wood in stem wood of Pinus densiflora[J]. Wood Science and Technology, 2020, 54(1): 123−137. doi: 10.1007/s00226-019-01140-w
[9] 李珠, 殷方宇, 蒋佳荔, 等. 杉木应压木和对应木的水分吸附特性比较研究[J]. 木材科学与技术, 2022, 36(5): 37−42. doi: 10.12326/j.2096-9694.2022018 Li Z, Yin F Y, Jiang J L, et al. Comparative studies on water vapor sorption characteristics between compression wood and opposite wood of Chinese fir[J]. Chinese Journal of Wood Science and Technology, 2022, 36(5): 37−42. doi: 10.12326/j.2096-9694.2022018
[10] Engelund E T, Salmén L. Tensile creep and recovery of Norway spruce influenced by temperature and moisture[J]. Holzforschung, 2012, 66(8): 959−965. doi: 10.1515/hf-2011-0172
[11] 邓彪, 罗迎社, 李贤军, 等. 荷载、含水率及温度对桉树木材抗弯蠕变性能的影响[J]. 中南林业科技大学学报, 2013, 33(5): 142−131. Deng B, Luo Y S, Li X J, et al. Effects of load, moisture content and temperature on flexural creep behavior of Eucalyptus[J]. Journal of Central South University of Forestry & Technology, 2013, 33(5): 142−131.
[12] 林金星, 李正理. 马尾松正常木与应压木的比较解剖[J]. 植物学报, 1993, 35(3): 201−205. Lin J X, Li Z L. Comparative anatomy of normal wood and compression wood of masson pine (Pinus massoniana)[J]. Acta Botanica Sinica, 1993, 35(3): 201−205.
[13] Wang J F, Wang X, He Q, et al. Time-temperature-stress equivalence in compressive creep response of Chinese fir at high-temperature range[J]. Construction and Building Materials, 2020, 235: 117809. doi: 10.1016/j.conbuildmat.2019.117809
[14] Li Z, Zhan T Y, Eder M, et al. Comparative studies on wood structure and microtensile properties between compression and opposite wood fibers of Chinese fir plantation[J]. Journal of Wood Science, 2021, 64(1): 12.
[15] Hou J F, Jiang Y Q, Yin Y Q, et al. Experimental study and comparative numerical modeling of creep behavior of white oak wood with various distributions of earlywood vessel belt[J]. Journal of Wood Science, 2021, 67(1): 57. doi: 10.1186/s10086-021-01989-1
[16] 尹业桥, 侯俊峰, 姜志宏, 等. 早材管孔分布对环孔材栎木蠕变特性的影响[J]. 林业工程学报, 2021, 6(3): 54−60. Yin Y Q, Hou J F, Jiang Z H, et al. Effect of earlywood vessel distribution on creep characteristics of ring-porous oak wood[J]. Journal of Forestry Engineering, 2021, 6(3): 54−60.
[17] 王聪, 吴强, 林鹏, 等. 不同纹理方向栎木微小无疵试样板材蠕变特性[J]. 林业科学, 2018, 54(4): 76−83. doi: 10.11707/j.1001-7488.20180409 Wang C, Wu Q, Lin P, et al. Orthotropic creep performance of small flawless oak board[J]. Scientia Silvae Sinicae, 2018, 54(4): 76−83. doi: 10.11707/j.1001-7488.20180409
[18] Peng H, Zhan T Y, Jiang J L, et al. Comparison of the time-moisture and time-temperature equivalences in the creep properties of Chinese fir[J]. Wood Material Science & Engineering, 2022, 17(6): 911−917.
[19] 蔡绍祥, 李延军, 王思群, 等. 马尾松木材轴向管胞细胞壁黏弹性变异研究[J]. 林产工业, 2021, 58(11): 1−7. Cai S X, Li Y J, Wang S Q, et al. Study on the variation of viscoelasticity of cell wall of masson pine tracheid at axial direction[J]. China Forest Products Industry, 2021, 58(11): 1−7.
[20] Keckes J, Burgert I, Klaus F, et al. Cell-wall recovery after irreversible deformation of wood[J]. Nature Materials, 2003, 2(12): 810−814. doi: 10.1038/nmat1019
-
期刊类型引用(10)
1. 王政,杨茗麟,肖泽芳,王海刚,王永贵,梁大鑫,谢延军. 高熔点石蜡处理木材的抗紫外光老化性能. 东北林业大学学报. 2021(04): 85-93 .  百度学术
百度学术
2. 张燕,张朔,薄宇轩,陈瑞瑶,赵喜龙. 传统烫蜡技艺传承及现代技术延伸. 林业机械与木工设备. 2021(04): 65-69 . 百度学术
3. 崔蒙蒙,吴智慧. 上蜡硬木家具表面性能评价方法研究. 林产工业. 2020(07): 24-29 . 百度学术
4. 李景奎,王亚男,牟洪波,戚大伟. 磁控溅射法制备纳米氧化锌/木材复合材料及其物理性能变化. 北京林业大学学报. 2019(01): 119-125 . 本站查看
5. 张海洋,鞠泽辉,何倩,詹先旭,梅长彤,卢晓宁. 线性振动摩擦作用下木材表面材色与硬度的变化. 西北林学院学报. 2019(02): 227-233 . 百度学术
6. 李景鹏,吴再兴,任丹静,陈玉和. 无机纳米材料在木竹材防霉防腐中的研究进展. 竹子学报. 2019(02): 16-23 . 百度学术
7. 崔蒙蒙,吴智慧,黄琼涛. 硬木家具上蜡装饰表面耐光性研究. 涂料工业. 2018(12): 54-63 . 百度学术
8. 杨小军,徐汀竹,张璐,郁岗,马澜,庞康颖. 户外用蜡浸注木工艺及物理力学性能. 森林与环境学报. 2017(04): 496-501 . 百度学术
9. 崔蒙蒙,吴智慧. 家具烫蜡技艺的研究与发展现状分析. 林产工业. 2017(02): 3-6 . 百度学术
10. 袁哲,曹积微,强明礼. 热软化及干燥处理对棕榈藤材色的影响. 西南林业大学学报. 2016(06): 158-163 . 百度学术
其他类型引用(7)







计量
- 文章访问数: 205
- HTML全文浏览量: 53
- PDF下载量: 65
- 被引次数: 17
